Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA and RNA Extraction
2.3. Reverse-Transcription Reaction
2.4. PCR for Fowl Adenovirus
2.5. PCR for Chicken Parvovirus
2.6. RT-PCR for Chicken Astrovirus and Avian Nephritis Virus
2.7. RT-PCR for Coronavirus
2.8. RT-PCR for Avian Rotavirus
2.9. RT-PCR for Avian Reovirus
2.10. Evaluation of RT-PCR and PCR Products
2.11. Sequencing and Phylogenetic Analysis
2.12. Statistical Analysis
3. Results
3.1. Single and Multiple Viral Infections
3.2. Age and type of birds
3.3. Clinical Signs
3.4. Geographic Origin
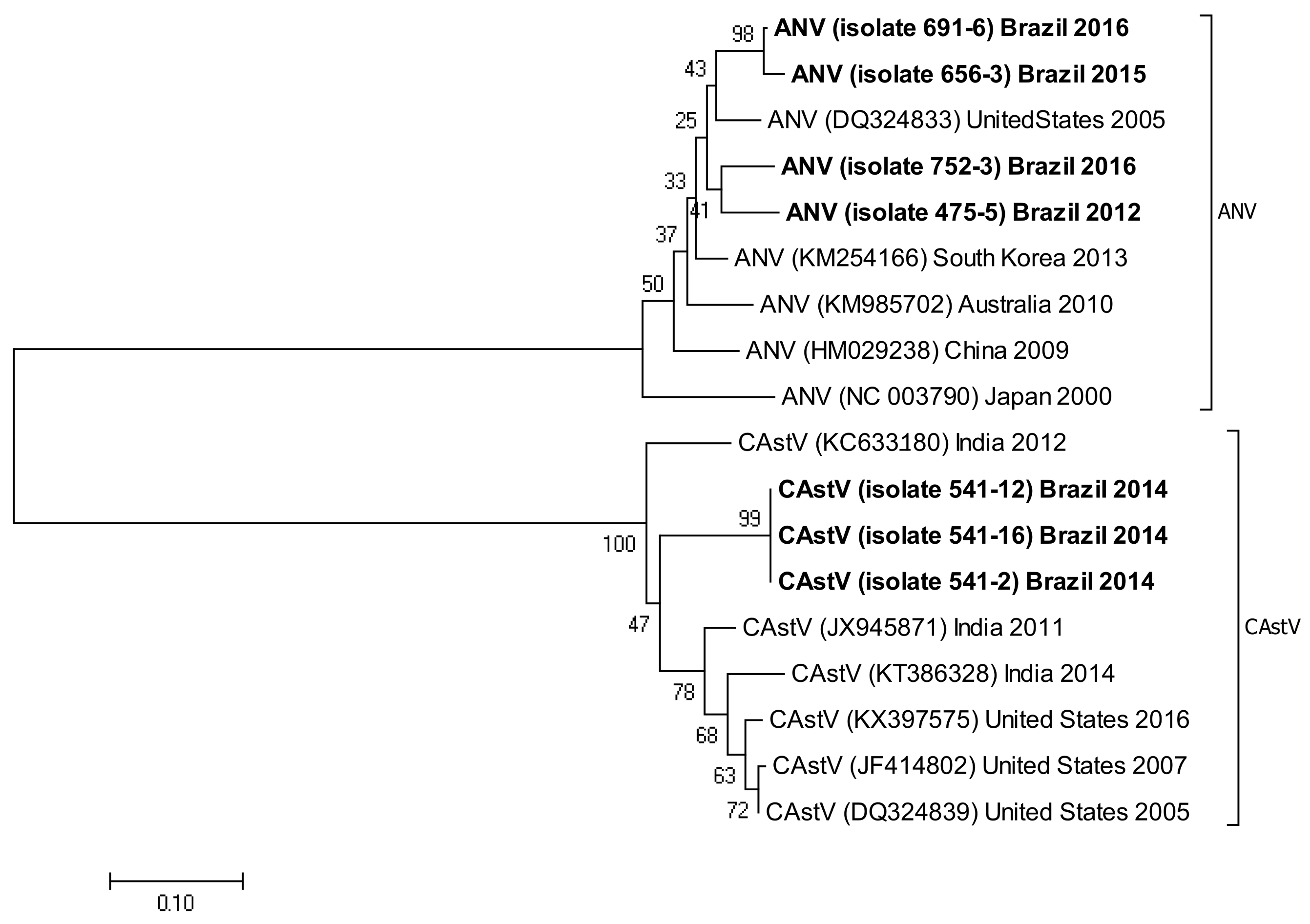
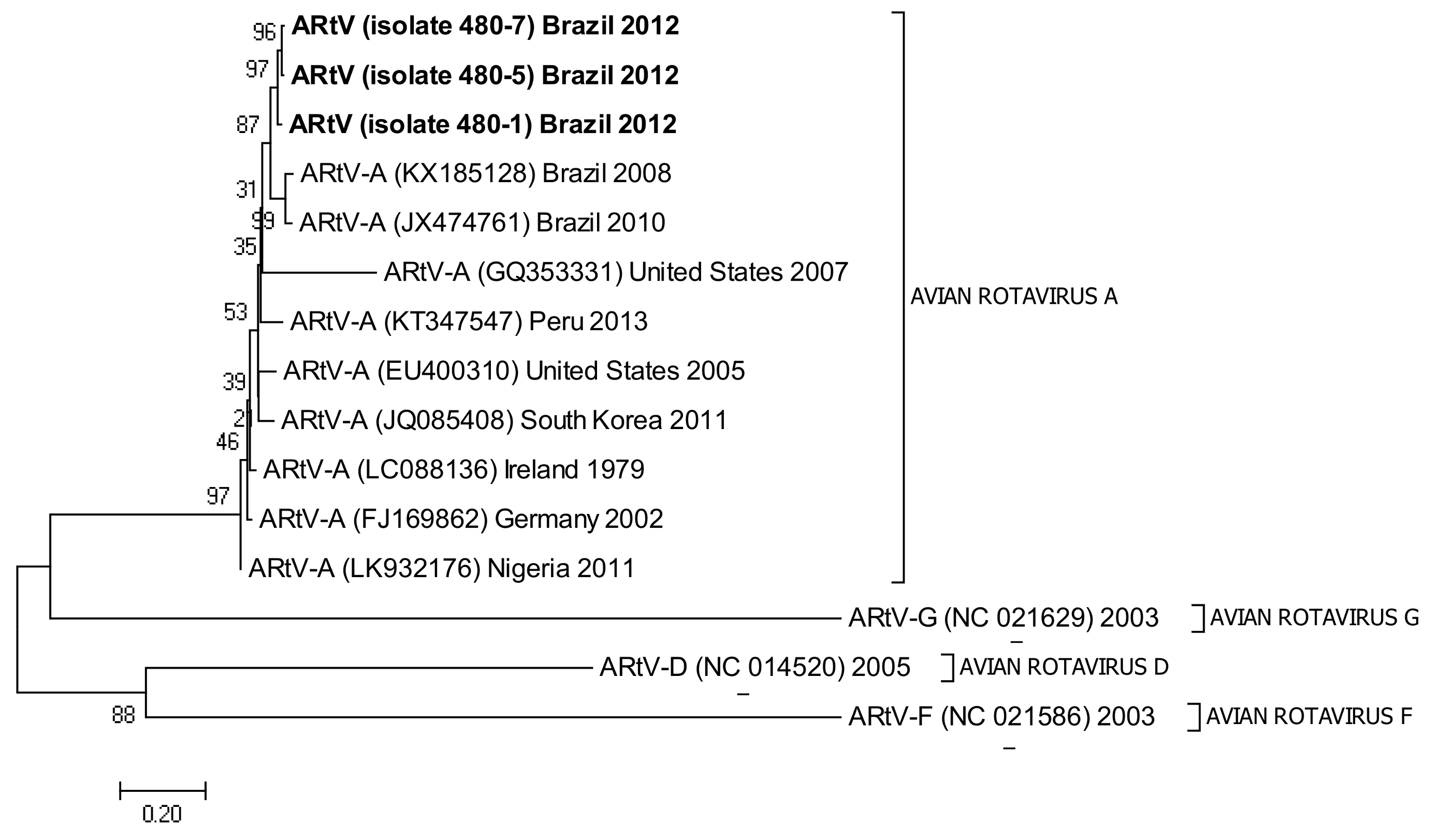
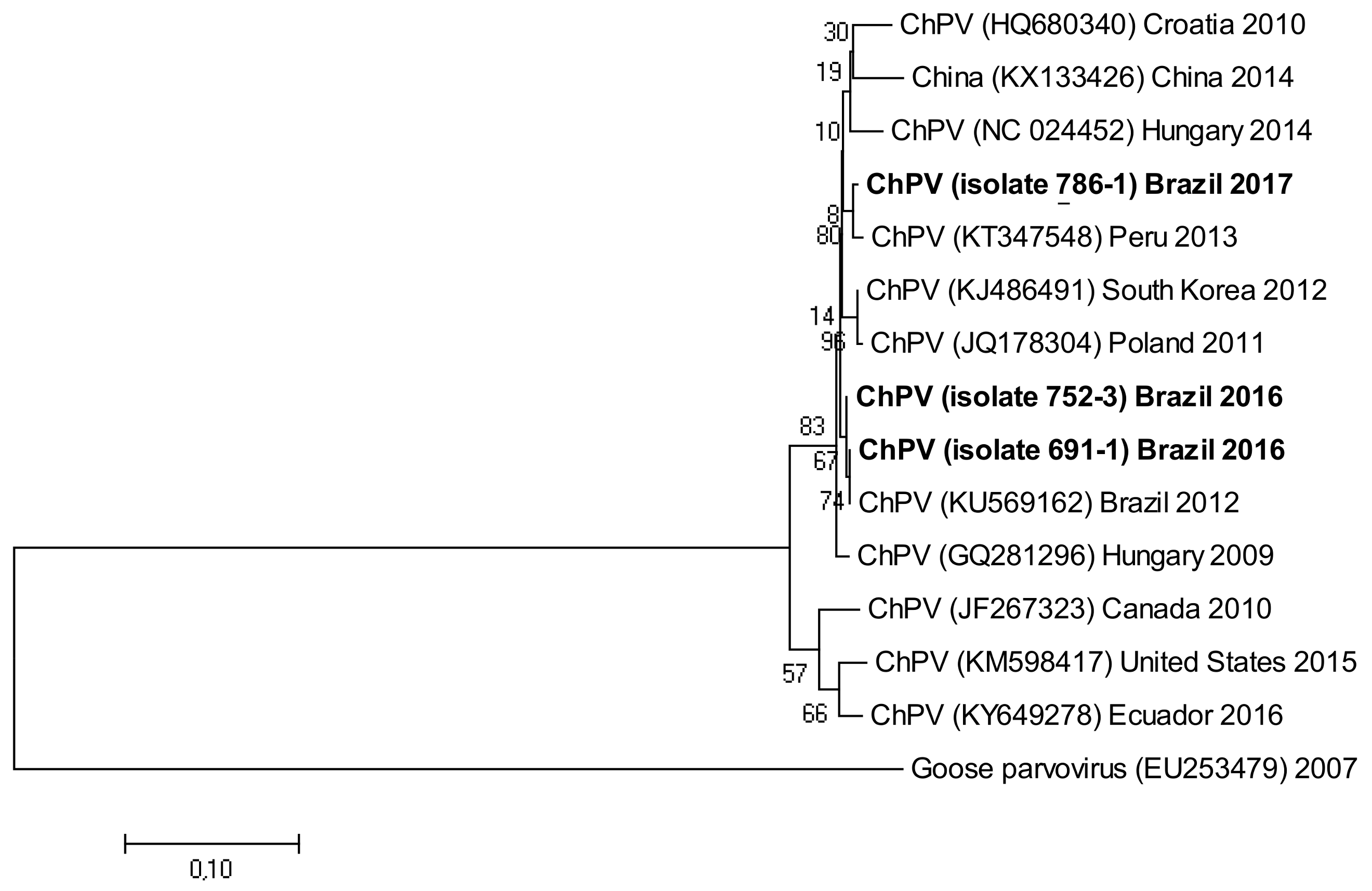
3.5. Phylogenetic Analysis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Otto, P.; Liebler-Tenorio, E.M.; Elschner, M.; Reetz, J.; Löhren, U.; Diller, R. Detection of rotaviruses and intestinal lesions in broiler chicks from flocks with runting and stunting syndrome (RSS). Avian Dis. 2006, 50, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.I.; El-Gazzar, M.; Sellers, H.S.; Dorea, F.; Susan, M.; Kim, T.; Collett, S.; Mundt, E. Investigation into the aetiology of runting and stunting syndrome in chickens. Avian Pathol. 2012, 41, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Devaney, R.; Trudgett, J.; Trudgett, A.; Meharg, C.; Smyth, V. A metagenomic comparison of endemic viruses from broiler chickens with runting stunting syndrome and from normal birds. Avian Pathol. 2016, 45, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, L.F.N.; Parra, S.H.S.; Astolfi-Ferreira, C.S.; Carranza, C.; Torre, D.I.; Pedroso, A.C.; Ferreira, A.J.P. Detection of enteric viruses in pancreas and spleen of broilers with runting-stunting syndrome (RSS). Pesq. Vet. Bras. 2016, 36, 595–599. [Google Scholar] [CrossRef]
- Jindal, N.; Patnayak, D.P.; Ziegler, A.F.; Lago, A.; Goyal, S.M. A retrospective study on poult enteritis syndrome in Minnesota. Avian Dis. 2009, 53, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Decaesstecker, M.; Charlier, G.; Meulemans, G. Epidemiological study of enteric viruses in broiler chickens: Comparison of tissue culture and direct electron microscopy. Avian Pathol. 1988, 17, 477–486. [Google Scholar] [CrossRef] [PubMed]
- Roussan, D.A.; Shaheen, I.A.; Khawaldeh, G.Y.; Totanji, W.S.; Al-Rifai, R.H. Simultaneous detection of astrovirus, rotavirus, reovirus and adenovirus type I in broiler chicken flocks. Pol. J. Vet. Sci. 2012, 15, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Moura-Alvarez, J.; Chacon, J.V.; Scanavini, L.S.; Nuñez, L.F.; Astolfi-Ferreira, C.S.; Jones, R.C.; Piantino Ferreira, A.J. Enteric viruses in Brazilian turkey flocks: Single and multiple virus infection frequency according to age and clinical signs of intestinal disease. Poult. Sci. 2013, 92, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Mettifogo, E.; Nuñez, L.F.; Chacón, J.L.; Santander Parra, S.H.; Astolfi-Ferreira, C.S.; Jerez, J.A.; Jones, R.C.; Piantino Ferreira, A.J. Emergence of enteric viruses in production chickens is a concern for avian health. Sci. World J. 2014, 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Meulemans, G.; Boschmans, M.; Berg, T.P.; Decaesstecker, M. Polymerase chain reaction combined with restriction enzyme analysis for detection and differentiation of fowl adenoviruses. Avian Pathol. 2001, 30, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D.; Mawditt, K.; Welchman, D.D.B.; Britton, P.; Gough, R.E. Coronaviruses from pheasants (Phasianus colchicus) are genetically closely related to coronaviruses of domestic fowl (infectious bronchitis virus) and turkeys. Avian Pathol. 2002, 31, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Day, J.M.; Spackman, E.; Pantin-Jackwood, M. A Multiplex RT-PCR test for the differential identification of turkey astrovirus type 1, turkey astrovirus type 2, chicken astrovirus, avian nephritis virus, and avian rotavirus. Avian Dis. 2007, 51, 681–684. [Google Scholar] [CrossRef]
- Pantin-Jackwood, M.J.; Day, J.M.; Jackwood, M.W.; Spackman, E. Enteric viruses detected by molecular methods in commercial chicken and turkey flocks in the United States between 2005 and 2006. Avian Dis. 2008, 52, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Zsak, L.; Strother, K.O.; Kisary, J. Partial genome sequence analysis of parvoviruses associated with enteric disease in poultry. Avian Pathol. 2008, 37, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Isolation of high-molecular-weight DNA using organic solvents. Cold Spring Harb. Protoc. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Zsak, L.; Strother, K.O.; Day, J.M. Development of a polymerase chain reaction procedure for detection of chicken and turkey parvoviruses. Avian Dis. 2009, 53, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, L.F.N.; Santander Parra, S.H.; Carranza, C.; Astolfi-Ferreira, C.S.; Buim, M.R.; Piantino Ferreira, A.J. Detection and molecular characterization of chicken astrovirus associated with chicks that have an unusual condition known as “white chicks” in Brazil. Poult. Sci. 2016, 95, 1262–1270. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Felsentein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [PubMed]
- Koo, B.S.; Lee, H.R.; Jeon, E.O.; Han, M.S.; Min, K.C.; Lee, S.B.; Mo, I.P. Molecular survey of enteric viruses in commercial chicken farms in Korea with a history of enteritis. Poult. Sci. 2013, 92, 2876–2885. [Google Scholar] [CrossRef] [PubMed]
- Zsak, L.; Cha, R.M.; Day, J.M. Chicken parvovirus—Induced runting-stunting syndrome in young broilers. Avian Dis. 2013, 57, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Jindal, N.; Patnayak, D.P.; Chander, Y.; Ziegler, A.F.; Goyal, S.M. Detection and molecular characterization of enteric viruses in breeder turkeys. Avian Pathol. 2010, 39, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Palade, E.A.; Kisary, J.; Benyeda, Z.; Mándoki, M.; Balka, G.; Jakab, C.; Végh, B.; Demeter, Z.; Rusvai, M. Naturally occurring parvoviral infection in Hungarian broiler flocks. Avian Pathol. 2011, 40, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Strother, K.O.; Mundt, E.; Zsak, L.; Day, J.M.; Spackman, E. Molecular characterization of avian astroviruses. Arch. Virol. 2011, 156, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Finkler, F.; Lima, D.A.; Cerva, C.; Cibulski, S.P.; Teixeira, T.F.; Santos, H.F.; Franco, A.C. Chicken parvovirus viral loads in cloacal swabs from malabsorption syndrome-affected and healthy broilers. Trop. Anim. Health Prod. 2016, 48, 1685–1689. [Google Scholar] [CrossRef] [PubMed]
- Pantin-Jackwood, M.J.; Spackman, E.; Woolcock, P.R. Molecular characterization and typing of chicken and turkey astroviruses circulating in the United States: Implications for diagnostics. Avian Dis. 2006, 50, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Sun, Q.; Zhang, G.; Sun, W.; Liu, X.; Xiao, Y.; Shang, Y.; Liu, S. Epidemiological investigation of outbreaks of fowl adenovirus infections in commercial chickens in China. Transbound Emerg. Dis. 2018, 65, e121–e126. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; Gonzalez, C.; Cerda, L.; Hess, M.; Reyes, E.; Geissea, C. Chicken anemia virus and fowl adenoviruses: Association to induce the inclusion body hepatitis/hydropericardium syndrome. Avian Dis. 2000, 44, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, L.F.N.; Piantino Ferreira, A.J. Viral agents related to enteric disease in commercial chicken flocks, with special reference to Latin America. World’s Poult. Sci. J. 2013, 69, 853–864. [Google Scholar] [CrossRef]
- Goodwin, M.A.; Hill, D.L.; Dekich, M.A.; Putnam, M.R. Multisystemic adenovirus infection in broiler chicks with hypoglycemia and spiking mortality. Avian Dis. 1993, 37, 625–627. [Google Scholar] [CrossRef] [PubMed]
- Ojkić, D.; Krell, P.J.; Tuboly, T.; Nagy, É. Characterization of fowl adenoviruses isolated in Ontario and Quebec, Canada. Can. J. Vet. Res. 2008, 72, 236–241. [Google Scholar] [PubMed]
- Romanova, N.; Corredor, J.C.; Nagy, É. Detection and quantitation of fowl adenovirus genome by a real-time PCR assay. J. Virol. Methods. 2009, 159, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Absalon, A.E.; Morales-Garzon, A.; Vera-Hernandez, P.F.; Cortes-Espinosa, D.V.; Uribe-Ochoa, S.M.; Garcia, L.J.; Lucio-Decanini, E. Complete genome sequence of a non-pathogenic strain of Fowl Adenovirus serotype 11: Minimal genomic differences between pathogenic and non-pathogenic viruses. Virology 2017, 501, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.E.; Reavill, D.R. Lesions of the Avian Pancreas. Vet. Clin. North Am. Exot. Anim. Pract. 2014, 17, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Marusak, R.A.; Guy, J.S.; Abdul-Aziz, T.A.; West, M.A.; Fletcher, O.J.; Day, J.M.; Zsak, L.; Barnes, H.J. Parvovirus-associated cerebellar hypoplasia and hydrocephalus in day old broiler chickens. Avian Dis. 2010, 54, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, L.F.; Sá, L.R.; Parra, S.H.; Astolfi-Ferreira, C.S.; Carranza, C.; Ferreira, A.J. Molecular detection of chicken parvovirus in broilers with enteric disorders presenting curving of duodenal loop, pancreatic atrophy, and mesenteritis. Poult. Sci. 2016, 95, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Hua, X.G.; Yuan, L.; Cui, L.; Shan, T.L.; Dai, X.; Zhu, A.L.; Yu, Y.; Zhu, C.X.; Yang, Z.B. Sequence analyses of the representative chinese-prevalent strain of avian nephritis virus in healthy chicken flocks. Avian Dis. 2011, 55, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Hungarica, A.V.; Vasudevan, G.; Nadu, T.; Sciences, A. Detection and partial genetic characterisation of a novel variant of avian nephritis virus in Indian poultry flocks showing diverse clinical signs. Acta Vet. Hung. 2015, 63, 499–507. [Google Scholar]
- Chamings, A.; Hewson, K.A.; Rourke, D.O.; Ignjatovic, J.; Chamings, A.; Hewson, K.A.; Noormohammadi, A.H. High-resolution melt curve analysis to confirm the presence of co-circulating isolates of avian nephritis virus in commercial chicken flocks. Avian Pathol. 2015, 44, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Wang, H.; Zhang, Y.; Guan, Z.; Wang, T.; Xu, C.; Yang, X. Comparative dynamic distribution of avian infectious bronchitis virus M41, H120, and SAIBK strains by quantitative real-time RT-PCR in SPF chickens. Biosci. Biotechnol. Biochem. 2012, 76, 2255–2260. [Google Scholar] [CrossRef] [PubMed]
- Day, J.M.; Oakley, B.B.; Seal, B.S.; Zsak, L. Comparative analysis of the intestinal bacterial and RNA viral communities from sentinel birds placed on selected broiler chicken farms. PLoS ONE 2015, 10, e0117210. [Google Scholar] [CrossRef] [PubMed]
- Christensen, N.H.; Saifuddin, M. A primary epidemic of inclusion body hepatitis in broilers. Avian Dis. 1989, 33, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Toro, H.; Prusas, C.; Raue, R.; Cerda, L.; Geissea, C.; González, C.; Hess, M. Characterization of fowl adenoviruses from outbreaks of inclusion body hepatitis/hydropericardium syndrome in Chile. Avian Dis. 1999, 43, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Lim, T.H.; Lee, H.J.; Lee, D.H.; Lee, Y.N.; Park, J.K.; Youn, H.N.; Kim, M.S.; Youn, H.S.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. Identification and virulence characterization of fowl adenoviruses in the Republic of Korea. Avian Dis. 2011, 55, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Maartens, L.H.; Joubert, H.W.; Aitchison, H.; Venter, E.H. Inclusion body hepatitis associated with an outbreak of fowl adenovirus type 2 and type 8b in broiler flocks in South Africa. J. S. Afr. Vet. Assoc. 2014, 85, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Niczyporuk, J.S. Phylogenetic and geographic analysis of fowl adenovirus field strains isolated from poultry in Poland. Arch. Virol. 2016, 161, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Domanska-Blicharz, K.; Jacukowicz, A.; Lisowska, A.; Minta, Z. Genetic characterization of parvoviruses circulating in turkey and chicken flocks in Poland. Arch. Virol. 2012, 157, 2425–2430. [Google Scholar] [CrossRef] [PubMed]
- Koo, B.S.; Lee, H.R.; Jeon, E.O.; Han, M.S.; Min, K.C.; Lee, S.B.; Bae, Y.J.; Cho, S.H.; Mo, J.S.; Kwon, H.M.; Sung, H.W.; Kim, J.N.; Mo, I.P. Genetic characterization of three novel chicken parvovirus strains based on analysis of their coding sequences. Avian Pathol. 2015, 44, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Hewson, K.A.; Rourke, D.O.; Noormohammadi, A.H.; Hewson, K.A.; Rourke, A.D.O.; Noormohammadi, A.H. Detection of avian nephritis virus in Australian chicken flocks. Avian Dis. 2010, 54, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Jindal, N.; Mor, S.K.; Goyal, S.M. Enteric viruses in turkey enteritis. Virusdisease. 2014, 25, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Mitake, H.; Yamada, D.; Nagai, M.; Okadera, K.; Ito, N.; Okada, K.; Nakagawa, K.; Mizutani, T.; Sugiyama, M. Genome sequences of rotavirus A strains Ty-1 and Ty-3, isolated from turkeys in Ireland in 1979. Genome Announc. 2016, 4, e01565-15. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Oni, O.O.; Sausy, A.; Owoade, A.A.; Adeyefa, C.A.O.; Muller, C.P.; Hübschen, J.M.; Snoeck, C.J. Molecular epidemiology of avian rotaviruses group A and D shed by different bird species in Nigeria. Virol. J. 2017, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Coronavirus avian infectious bronchitis virus. Vet. Res. 2007, 38, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Seger, W.; Ghalyanchi Langeroudi, A.; Karimi, V.; Madadgar, O.; Vasfi Marandi, M.; Hashemzadeh, M. Prevalence of avian infectious bronchitis virus in broiler chicken farms in south of Iraq, 2014–2015. Vet. Res. Forum 2016, 7, 317–321. [Google Scholar] [PubMed]
- Hutton, S.; Bettridge, J.; Christley, R.; Habte, T.; Ganapathy, K. Detection of infectious bronchitis virus 793B, avian metapneumovirus, Mycoplasma gallisepticum and Mycoplasma synoviae in poultry in Ethiopia. Trop. Anim. Health Prod. 2017, 49, 317–322. [Google Scholar] [CrossRef]
- Lima, D.A.; Cibulski, S.P.; Finkler, F.; Teixeira, T.F.; Varela, A.P.M.; Cerva, C.; Loiko, M.R.; Scheffer, C.M.; Dos Santos, H.F.; Mayer, F.Q.; Roehe, P.M. Fecal virome of healthy chickens reveals a large diversity of the eukaryote viral community, including novel circular ssDNA viruses. J. Gen. Virol. 2017, 98, 690–703. [Google Scholar] [PubMed]
- Espinoza, L.; Beserra, L.A.; Soares, R.M.; Gregori, F.; Espinoza, L.L.; Beserra, A.L.; Gregori, F. Turkey astrovirus type 1 (TAstV-1) and chicken astrovirus (CAstV) detection in Brazilian chicken flocks. Avian Dis. 2016, 60, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, L.L.; Beserra, L.A.; Soares, R.M.; Gregori, F. Avian nephritis virus (ANV) on Brazilian chicken farms: Circulating genotypes and intra-genotypic diversity. Arch. Virol. 2016, 161, 3455–3462. [Google Scholar] [CrossRef] [PubMed]
- Rios, R.L.; Marin, S.Y.; Gomes, A.M.; Resende, J.S.; Gomes, A.D.; Horta, R.S.; Resende, M. The occurrence of orthoreovirus, rotavirus and chicken anemia virus in chickens of the poultry industry in Minas Gerais, Brazil. Arq. Bras. Med. Vet. Zootec. 2012, 64, 1613–1620. [Google Scholar] [CrossRef]
- Beserra, L.A.R.; Barbosa, B.R.P.; Bernardes, N.T.C.G.; Brandao, P.E.; Gregori, F. Occurrence and characterization of rotavirus A in broilers, layers, and broiler breeders from Brazilian poultry farms. Avian Dis. 2014, 58, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, L.Y.; Brandão, P.E.; Chacón, J.L.; Saidenberg, A.B.S.; Assayag, M.S.; Jones, R.C.; Ferreira, A.J.P. Molecular characterization of infectious bronchitis virus strains isolated from the enteric contents of Brazilian laying hens and broilers. Avian Dis. 2007, 51, 974–978. [Google Scholar] [CrossRef] [PubMed][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Gene Target * | Primer Name | Primer Sequence | bp | Reference |
|---|---|---|---|---|---|
| FAdV-I | Hexon | Hexon A | 5’-CAARTTCAGRCAGACGGT-3’ | 897 | [10] |
| Hexon B | 5’-TAGTGATGMCGSGACATCAT-3’ | ||||
| ChPV | NS | PVF1 | 5’-TTCTAATAACGATATCACTCAAGTTTC-3’ | 561 | [17] |
| PVR1 | 5’-TTTGCGCTTGCGGTGAAGTCT GGCTCG-3’ | ||||
| IBV | UTR | UTR 11 | 5’-GCTCTAACTCTATACTAGCCTA-3’ | 179 | [11] |
| UTR 31 | 5’-GGGCGTCCAAGTGCTGTACCC-3’ | ||||
| UTR 41 | 5’-ATGTCTATCGCCAGGGAAATGTC-3’ | ||||
| CAstV | ORF 1b | CAS pol 1F | 5’-GAYCARCGAATGCGRAGRTTG-3’ | 362 | [12] |
| CAS pol 1R | 5’-TCAGTGGAAGTGGGKARTCTAC-3’ | ||||
| ANV | ORF 1b | ANV pol 1F | 5’-GYTGGGCGCYTCYTTTGAYACCRT-3’ | 473 | [12] |
| ANV pol 1R | 5’-CRTTTGCCCKRTARTCTTTRTGAY-3’ | ||||
| AReo | S4 | S4-F13 | 5’-GTGCGTGTTGGAGTTTCCCG-3’ | 1,120 | [13] |
| S4-R1133 | 5’-TACGCCATCCTAGCTGGA-3’ | ||||
| ARtV | NSP4 | NSP4-F30 | 5’-GTGCGGAAAGATGGAGAAC-3’ | 630 | [12] |
| NSP4-R660 | 5’-GTTGGGGTACCAGGGATTAA-3’ |
| Number of Viruses | FAdV-I | ANV | CAstV | IBV | ChPV | AReo | ARtV | Single and Multiple Positive Samples |
|---|---|---|---|---|---|---|---|---|
| 1 virus | 22 | 19 | 15 | 164 | 8 | 1 | 4 | 233/270 (86.3%) |
| 2 viruses | 3 | 6 | 7 | 14 | 1 | 0 | 3 | 17/270 (6.3%) |
| 3 viruses | 3 | 12 | 1 | 14 | 10 | 0 | 5 | 15/270 (5.6%) |
| 4 viruses | 0 | 4 | 3 | 3 | 2 | 0 | 4 | 4/270 (1.5%) |
| 5 viruses | 0 | 1 | 1 | 1 | 1 | 0 | 1 | 1/270 (0.4%) |
| Number of positive samples for each virus | 28 | 42 | 27 | 196 | 22 | 1 | 17 | - |
| % of samples positive for each virus (n = 333) | 8.4% | 12.6% | 8.1% | 58.9% | 6.6% | 0.3% | 5.1% | - |
| Patterns | Virus Examined in 270 Digestive Organs | Number of Samples in Each Pattern (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| FAdV-I | ChPV | CAstV | ANV | IBV | AReo | ARtV | ||||
| 1 | + | - | - | - | - | - | - | 22 | 8.1% | |
| 2 | + | - | - | + | - | - | - | 1 | 0.4% | |
| 3 | + | - | - | - | + | - | - | 2 | 0.7% | |
| 4 | + | + | - | - | + | - | - | 2 | 0.7% | |
| 5 | + | - | - | + | + | - | - | 1 | 0.4% | |
| 6 | - | - | - | + | - | - | - | 19 | 7% | |
| 7 | - | - | - | + | + | - | - | 3 | 1.1% | |
| 8 | - | - | + | + | - | - | - | 1 | 0.4% | |
| 9 | - | - | - | + | - | - | + | 1 | 0.4% | |
| 10 | - | + | - | + | + | - | - | 7 | 2.6% | |
| 11 | - | - | - | + | + | - | + | 3 | 1.1% | |
| 12 | - | + | - | + | - | - | + | 1 | 0.4% | |
| 13 | - | - | + | + | + | - | + | 2 | 0.7% | |
| 14 | - | + | + | + | - | - | + | 1 | 0.4% | |
| 15 | - | + | - | + | + | - | + | 1 | 0.4% | |
| 16 | - | + | + | + | + | - | + | 1 | 0.4% | |
| 17 | - | - | + | - | - | - | - | 15 | 5.6% | |
| 18 | - | - | + | - | + | - | - | 6 | 2.2% | |
| 19 | - | - | + | - | + | - | + | 1 | 0.4% | |
| 20 | - | - | - | - | + | - | - | 164 | 60.7% | |
| 21 | - | + | - | - | + | - | - | 1 | 0.4% | |
| 22 | - | - | - | - | + | - | + | 2 | 0.7% | |
| 23 | - | + | - | - | - | - | - | 8 | 3% | |
| 24 | - | - | - | - | - | - | + | 4 | 1.5% | |
| 25 | - | - | - | - | - | + | - | 1 | 0.4% | |
| Age of Flocks | Layers | Breeders | ||||
|---|---|---|---|---|---|---|
| Number of Positive Samples | % of Samples Positive for Each Week of Age | Number of Positive Samples | % of Samples Positive for Each Week of Age | |||
| 1–10 weeks |  | 49 | 16.3% |  | 37 | 18.9% |
| 11–20 weeks | 18.4% | 0% | ||||
| 21–30 weeks | 4.1% | 0% | ||||
| 31–40 weeks | 16.3% | 13.5% | ||||
| 41–50 weeks | 0% | 18.9% | ||||
| >51 weeks | 22.4% | 37.8% | ||||
| Not reported | 22.4% | 10.8% | ||||
| Age of Flocks | Number of Positive Samples | % of Samples Positive for Each Day of Age | |
|---|---|---|---|
| 1–7 days |  | 163 | 7.4% |
| 8–14 days | 15.3% | ||
| 15–21 days | 14.7% | ||
| 22–28 days | 6.7% | ||
| 29–35 days | 4.9% | ||
| 36–42 days | 32.5% | ||
| >43 days | 11.7% | ||
| Not reported | 6.7% | ||
| Item | Positive Samples with Single and Multiple Viral Infections | Clinical Signs | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 Virus | 2 Viruses | 3 Viruses | 4 Viruses | 5 Viruses | Respiratory Problems | Digestive Problems | Poor Production * | No Clinical Signs | Not Reported | |
| Broilers (n = 163) | 129 (55.4%) | 14 (82.4%) | 15 (100%) | 4 (100%) | 1 (100%) | 12 (30.8%) | 9 (100%) | 55 (79.7%) | 1 (20%) | 86 (58%) |
| Layers (n = 49) | 49 (21%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 23 (59%) | 0 (0%) | 2 (2.9%) | 0 (0%) | 24 (16.2%) |
| Breeders (n = 37) | 34 (14.6%) | 3 (17.6%) | 0 (0%) | 0 (0%) | 0 (0%) | 3 (7.7%) | 0 (0%) | 12 (17.4%) | 4 (80%) | 18 (12.2%) |
| Not reported (n = 21) | 21 (9%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 1 (2.6%) | 0 (0%) | 0 (0%) | 0 (0%) | 20 (13.5%) |
| Number of positive samples (n = 270) | 233 | 17 | 15 | 4 | 1 | 39 | 9 | 69 | 5 | 148 |
| % of positive samples independently of chicken line | 86.3% | 6.3% | 5.6% | 1.5% | 0.4% | 14.4% | 3.3% | 25.6% | 1.9% | 54.8% |
| Sample | FAdV-I | ChPV | CAstV | ANV | IBV | AReo | ARtV |
|---|---|---|---|---|---|---|---|
| Liver (n = 39) | 19 | 3 | 1 | 1 | 14 | 0 | 2 |
| Intestines (n = 89) | 4 | 14 | 19 | 27 | 52 | 1 | 4 |
| Pancreas (n = 25) | 5 | 5 | 7 | 14 | 13 | 0 | 11 |
| Caecal tonsils (n = 20) | 0 | 0 | 0 | 0 | 20 | 0 | 0 |
| Cloacal swabs (n = 97) | 0 | 0 | 0 | 0 | 97 | 0 | 0 |
| Total (viruses = 333) | 28 | 22 | 27 | 42 | 196 | 1 | 17 |
| Chicken Lines | Age of Birds | FAdV-I | ChPV | CAstV | ANV | IBV | AReo | ARtV | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Layers | 1–10 weeks |  | 0 | | 0 | | 0 | | 0 |  | 49 | | 0 | | 0 |
| 11–20 weeks | |||||||||||||||
| 21–30 weeks | |||||||||||||||
| 31–40 weeks | |||||||||||||||
| 41–50 weeks | |||||||||||||||
| >51 weeks | |||||||||||||||
| N/R | |||||||||||||||
| Breeders | 1–10 weeks |  | 12 | | 0 |  | 10 | | 0 |  | 16 | | 0 |  | 2 |
| 11–20 weeks | |||||||||||||||
| 21–30 weeks | |||||||||||||||
| 31–40 weeks | |||||||||||||||
| 41–50 weeks | |||||||||||||||
| >51 weeks | |||||||||||||||
| N/R | |||||||||||||||
| Broilers | 1–7 days |  | 14 |  | 20 |  | 17 |  | 41 |  | 116 | | 0 |  | 15 |
| 8–14 days | |||||||||||||||
| 15–21 days | |||||||||||||||
| 22–28 days | |||||||||||||||
| 29–35 days | |||||||||||||||
| 36–42 days | |||||||||||||||
| >43 days | |||||||||||||||
| N/R | |||||||||||||||
| Not reported neither age nor line | 2 | 2 | 0 | 1 | 15 | 1 | 0 | ||||||||
| Total viruses = 333 * | 28 | 22 | 27 | 42 | 196 | 1 | 17 | ||||||||
| Brazilian States—Samples Origin | Total Positive Samples * | % of Samples Positive for Each State | Virus Strains Examined in This Study ** | ||||||
|---|---|---|---|---|---|---|---|---|---|
| FAdV-I | ChPV | CAstV | ANV | IBV | AReo | ARtV | |||
| Bahia | 6 | 2.2% | 3 (13.6) A | 6 (14.3) | 5 (2.6) | ||||
| Ceara | 5 | 1.9% | 2 (9.1) | 2 (4.8) | 1 (0.5) | ||||
| Espirito Santo | 3 | 1.1% | 3 (1.5) | ||||||
| Goias | 13 | 4.8% | 13 (6.6) | ||||||
| Mato Grosso | 11 | 4.1% | 11 (5.6) | ||||||
| Minas Gerais | 50 | 18.5% | 4 (18.2) | 20 (74.1) | 14 (33.3) | 33 (16.8) | 11 (64.7) | ||
| Paraiba | 4 | 1.5% | 1 (4.5) | 3 (7.1) | |||||
| Pernambuco | 9 | 3.3% | 1 (3.6) | 2 (9.1) | 5 (11.9) | 1 (0.5) | 1 (5.9) | ||
| Piaui | 1 | 0.4% | 1 (0.5) | ||||||
| Santa Catarina | 7 | 2.6% | 7 (25) | ||||||
| São Paulo | 82 | 30.4% | 18 (64.3) | 3 (13.6) | 7 (16.7) | 65 (33.2) | 3 (17.6) | ||
| Not informed | 79 | 29.3% | 2 (7.1) | 7 (31.8) | 7 (25.9) | 5 (11.9) | 63 (32.1) | 1 (100) | 2 (11.8) |
| Total | 270 | 100% | 28 (100) | 22 (100) | 27 (100) | 42 (100) | 196 (100) | 1 (100) | 17 (100) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De la Torre, D.I.; Nuñez, L.F.; Astolfi-Ferreira, C.S.; Piantino Ferreira, A.J. Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks. Vet. Sci. 2018, 5, 38. https://doi.org/10.3390/vetsci5020038
De la Torre DI, Nuñez LF, Astolfi-Ferreira CS, Piantino Ferreira AJ. Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks. Veterinary Sciences. 2018; 5(2):38. https://doi.org/10.3390/vetsci5020038
Chicago/Turabian StyleDe la Torre, David I., Luis F. Nuñez, Claudete S. Astolfi-Ferreira, and Antonio J. Piantino Ferreira. 2018. "Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks" Veterinary Sciences 5, no. 2: 38. https://doi.org/10.3390/vetsci5020038
APA StyleDe la Torre, D. I., Nuñez, L. F., Astolfi-Ferreira, C. S., & Piantino Ferreira, A. J. (2018). Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks. Veterinary Sciences, 5(2), 38. https://doi.org/10.3390/vetsci5020038