Pathological Study on Epithelial-Mesenchymal Transition in Silicotic Lung Lesions in Rat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
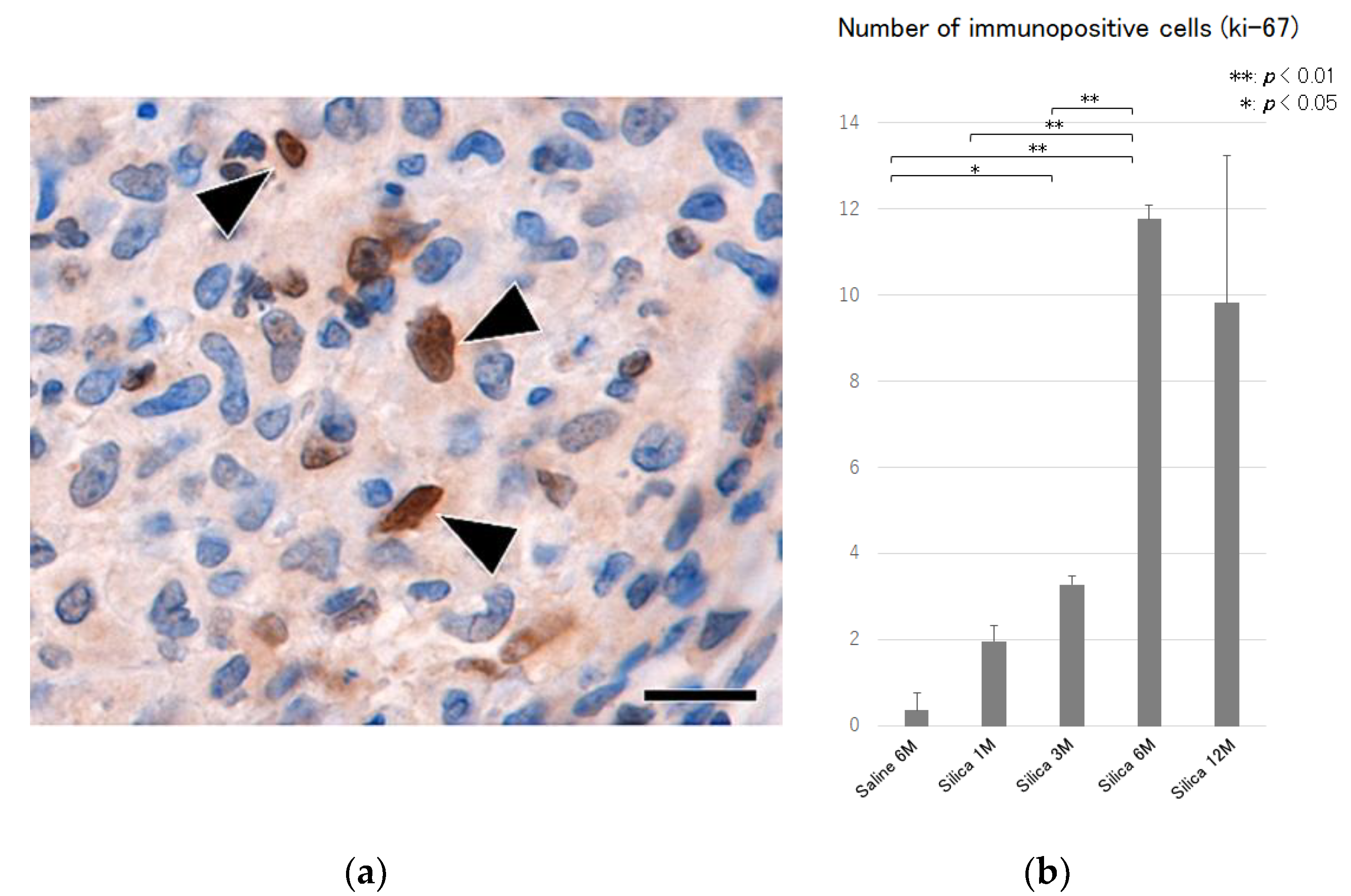{kind=link}
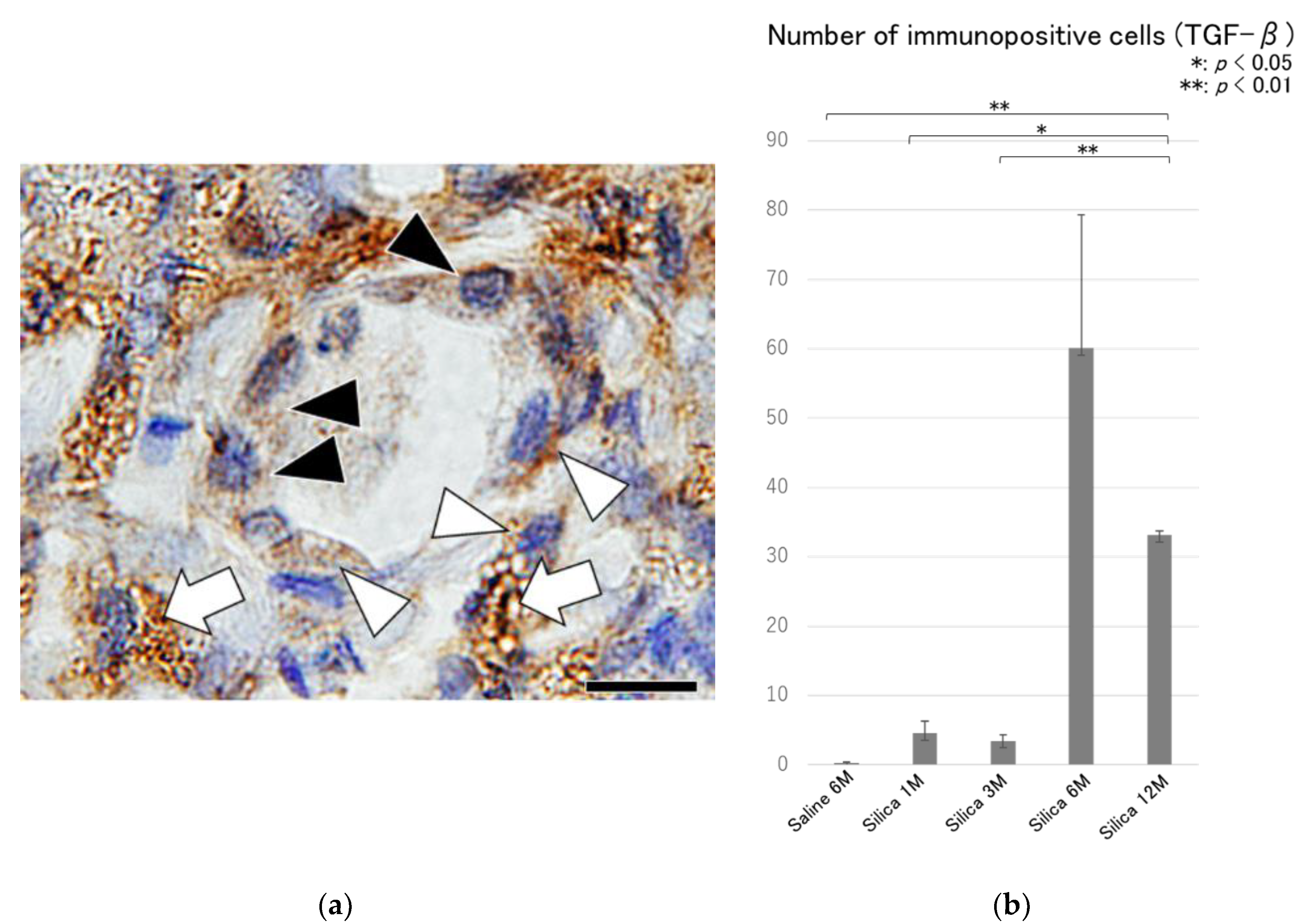{kind=link}
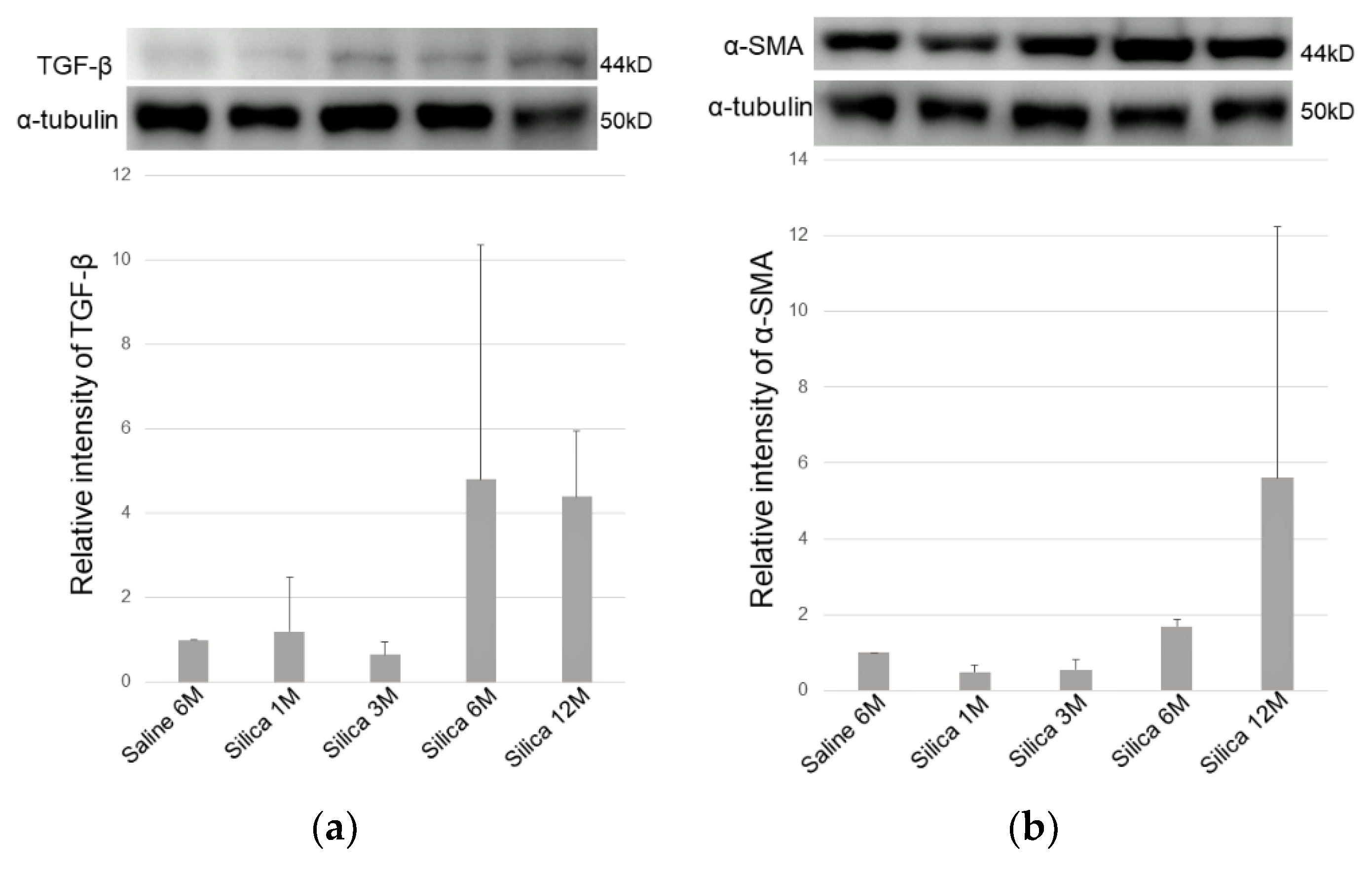{kind=link}
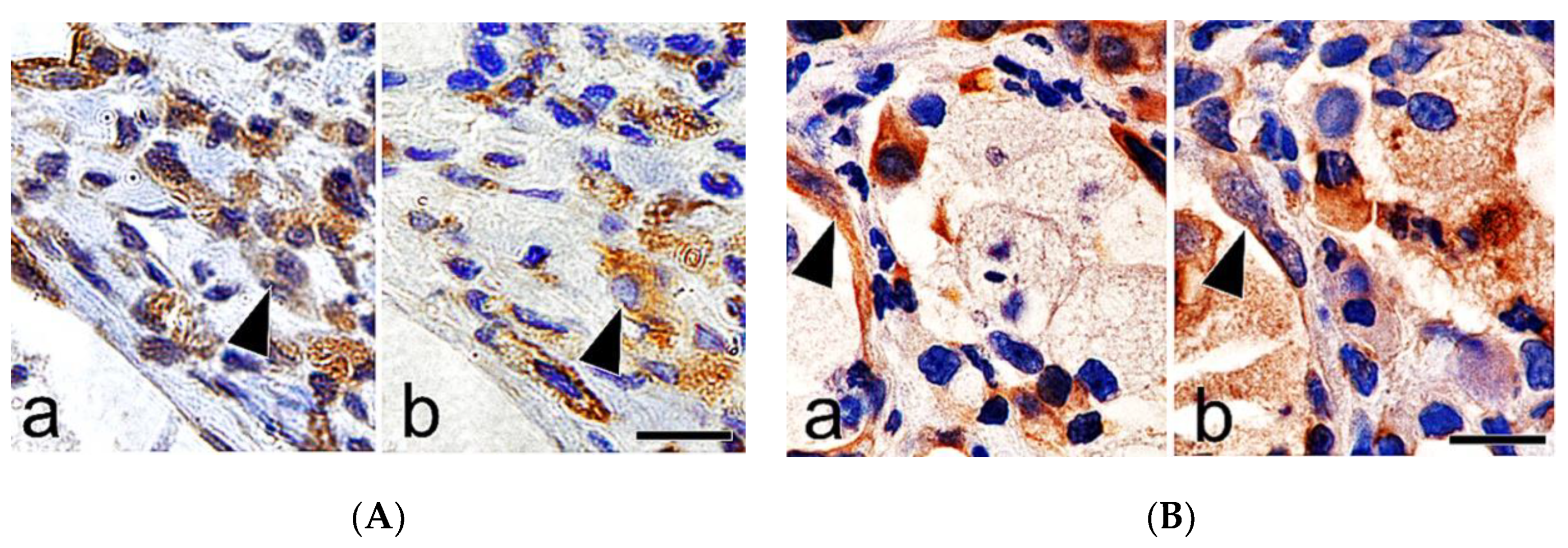{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Silica Particle Suspensions
2.3. Study Protocol
2.4. Pathological Examination
2.5. Immunohistochemistry
2.6. Statistical Analysis
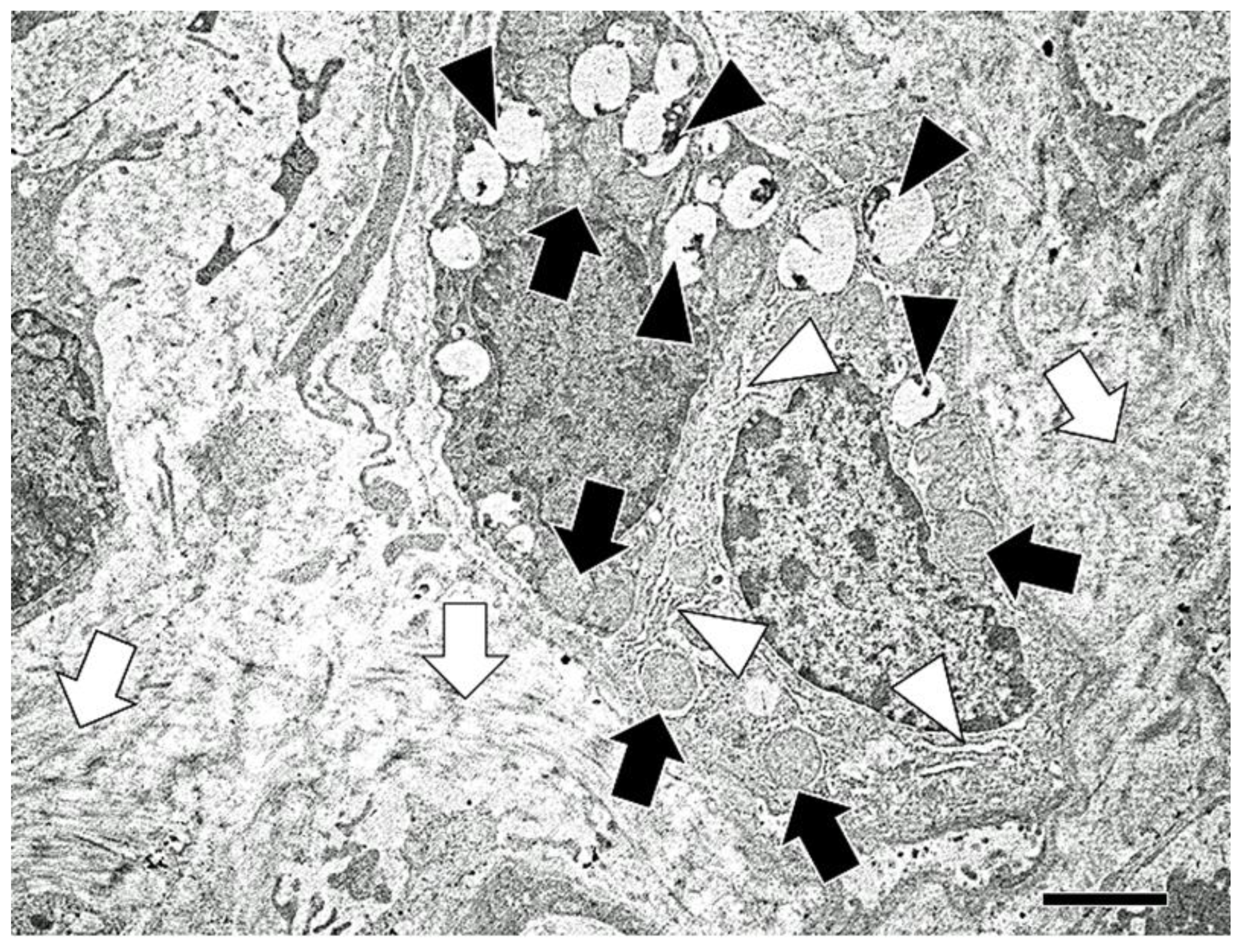
2.7. Transmission Electron Microscopy
2.8. Western Blot Analysis
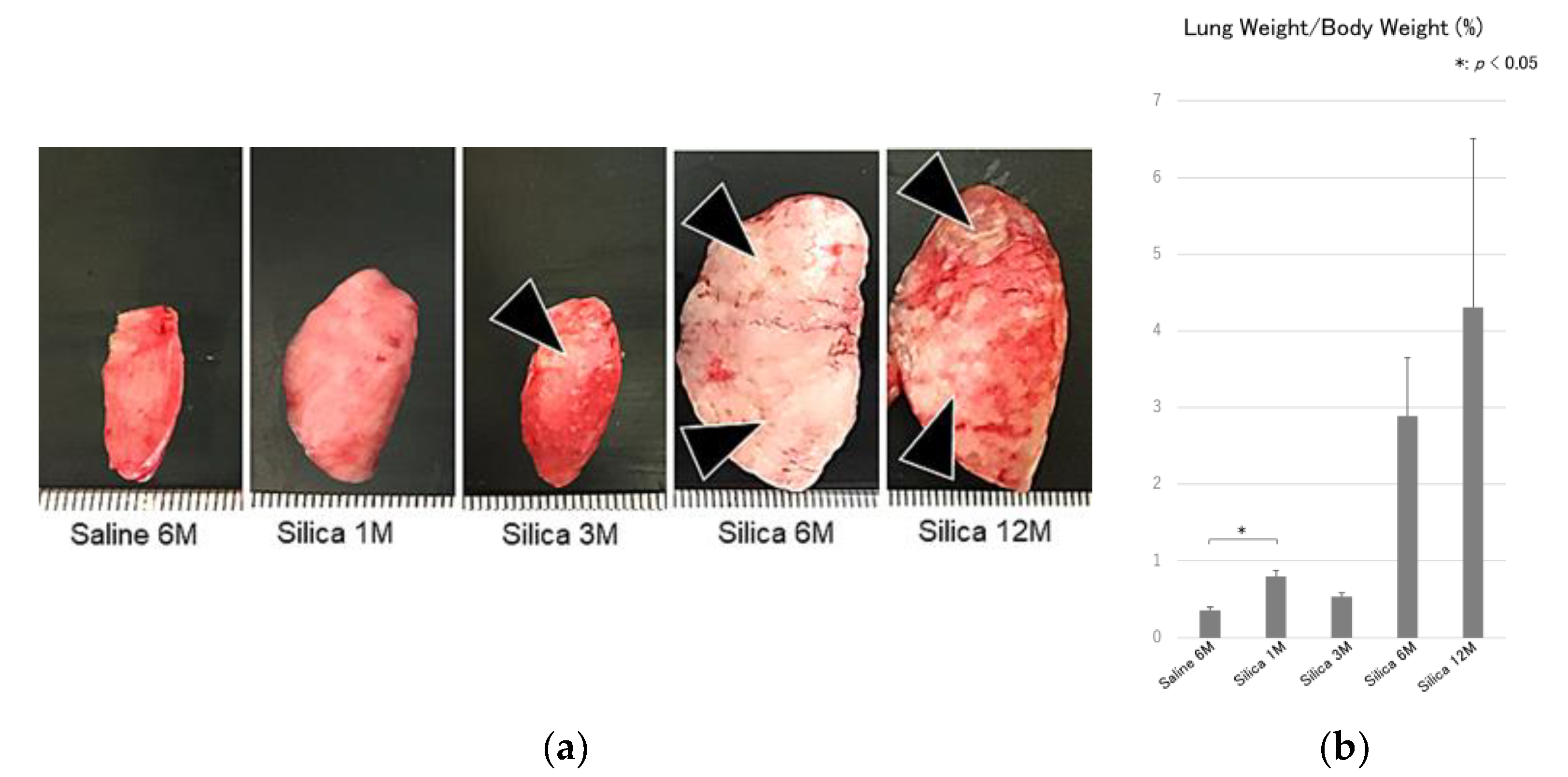
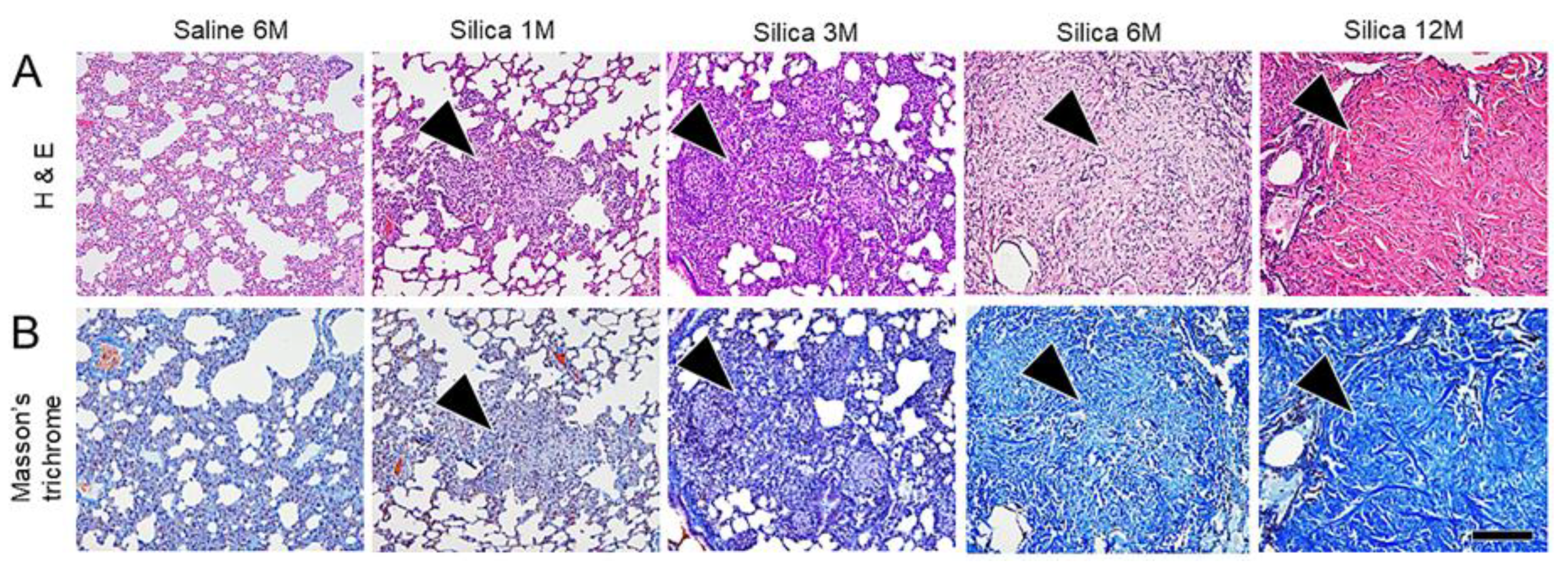
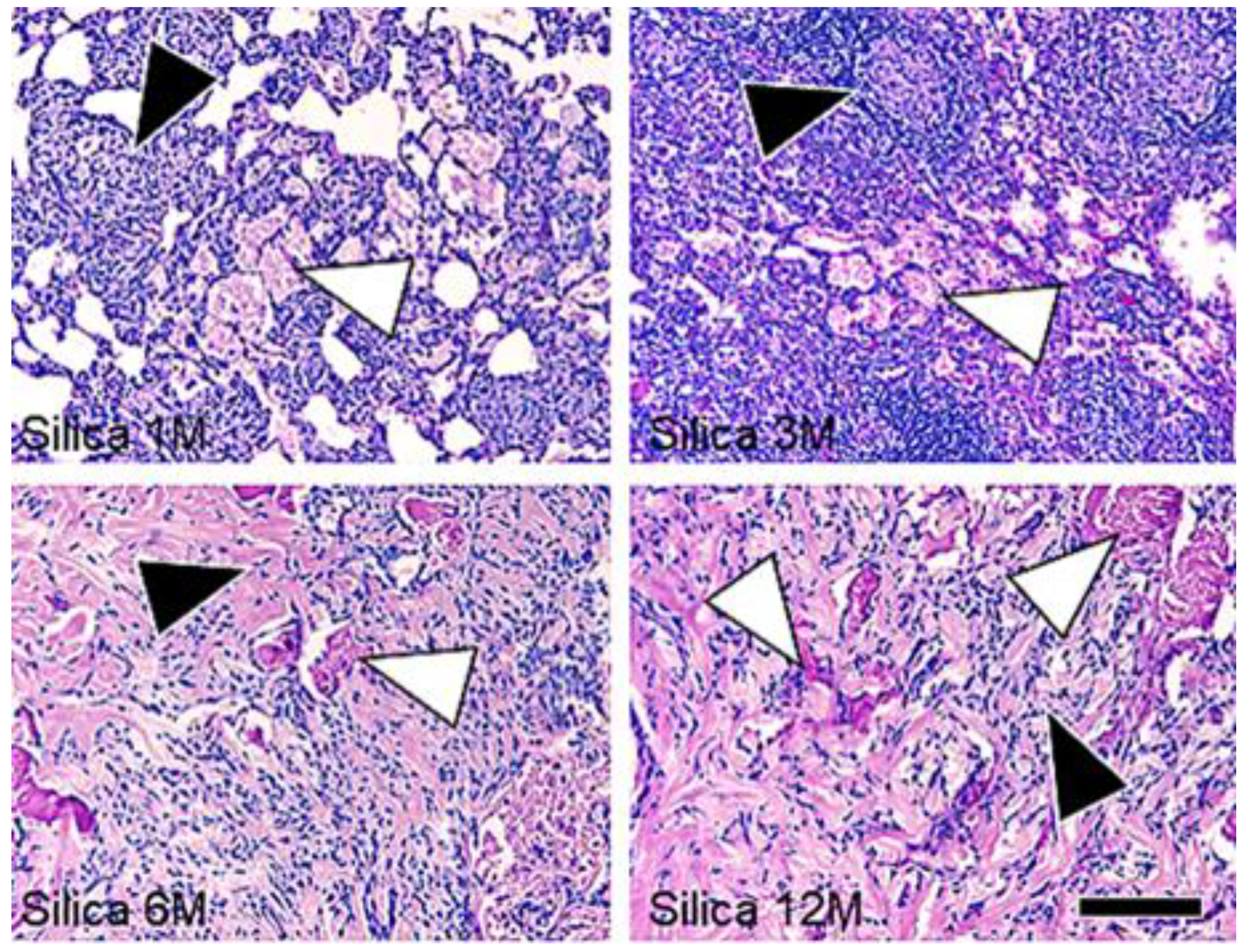
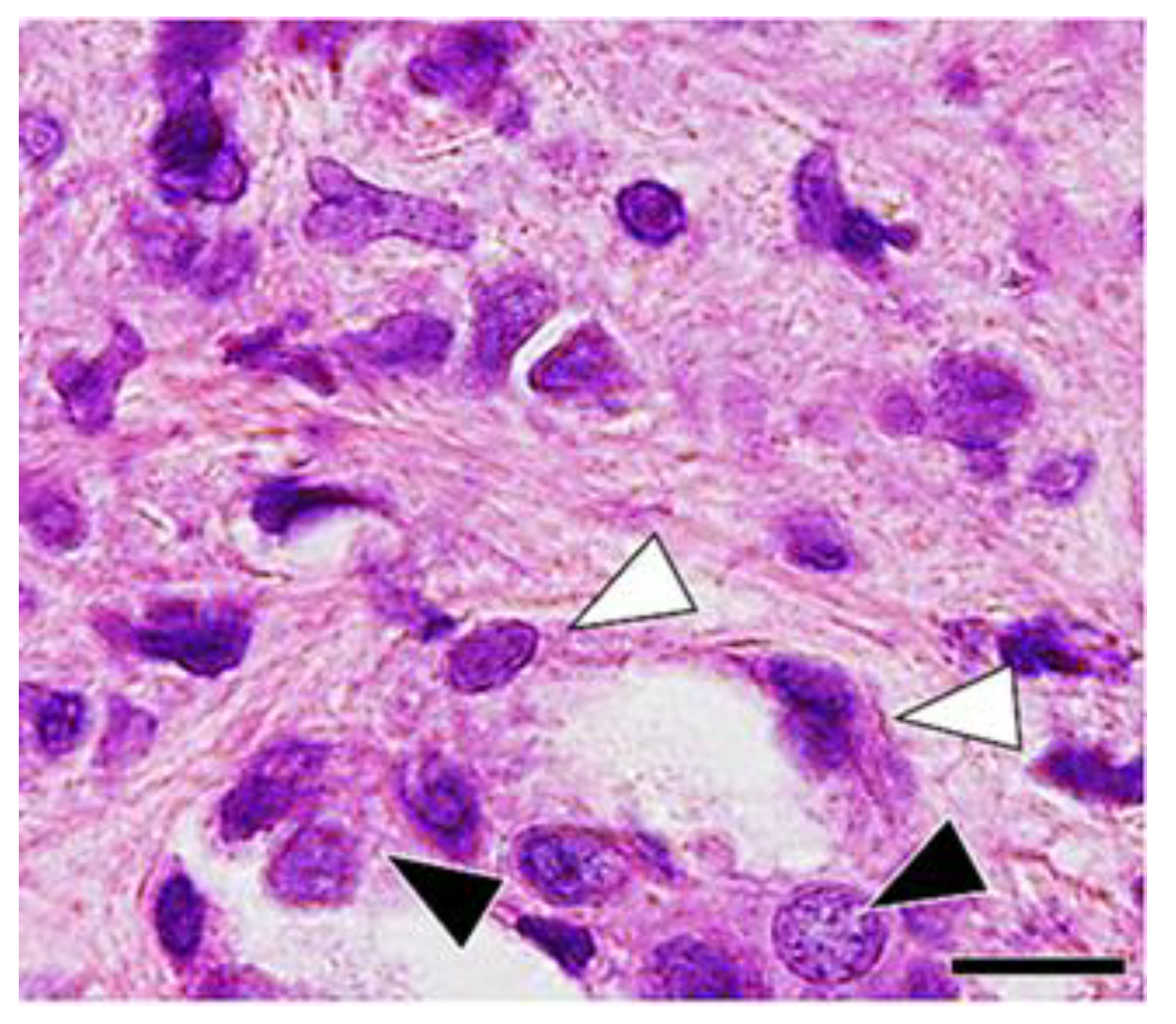
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leung, C.C.; Yu, I.T.; Chen, W. Silicosis. Lancet 2012, 379, 2008–2018. [Google Scholar] [CrossRef]
- Greenberg, M.I.; Waksman, J.; Curtis, J. Silicosis: A review. Dis. Mon. 2007, 53, 394–416. [Google Scholar] [CrossRef] [PubMed]
- Jalloul, A.S.; Banks, D.E. The health effects of silica exposure. In Environmental and Occupational Medicine, 4th ed.; Rom, W.N., Ed.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 365–387. [Google Scholar]
- Mossman, B.T.; Churg, A. Mechanisms in the pathogenesis of asbestosis and silicosis. Am. J. Respir. Crit. Care Med. 1998, 157, 1666–1680. [Google Scholar] [CrossRef] [PubMed]
- Muhle, H.; Kittel, B.; Ernst, H.; Mohr, U.; Mermelstein, R. Neoplastic lung lesions in rat after chronic exposure to crystalline silica. Scand. J. Work Environ. Health 1995, 21, 27–29. [Google Scholar] [PubMed]
- Chang, C.C.; Huang, H.C.; Chen, C.Y.; Tsai, P.J. Silica exposure causes epithelial-mesenchymal transition and pulmonary fibrosis. Epidemiology 2011, 22, S184. [Google Scholar] [CrossRef]
- Chapman, H.A. Epithelial-mesenchymal interactions in pulmonary fibrosis. Ann. Rev. Physiol. 2011, 73, 413–435. [Google Scholar] [CrossRef] [PubMed]
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.M.; Dedhar, S.; Kalluri, R.; Thompson, E.W. The epithelial-mesenchymal transition: New insights in signaling, development, and disease. J. Cell Biol. 2006, 172, 973–981. [Google Scholar] [CrossRef]
- Acloque, H.; Thiery, J.P.; Nieto, M.A. The physiology and pathology of the EMT. Meeting on the Epithelial–Mesenchymal Transition. EMBO Rep. 2008, 9, 322–326. [Google Scholar] [CrossRef]
- Micalizzi, D.S.; Farabaugh, S.M.; Ford, H.L. Epithelial-mesenchymal transition in cancer: Parallels between normal development and tumor progression. J. Mammary Gland Biol. Neoplasia 2010, 15, 117–134. [Google Scholar] [CrossRef]
- Bartis, D.; Mise, N.; Mahida, R.Y.; Eickelberg, O.; Thickett, D.R. Epithelial-mesenchymal transition in lung development and disease: Does it exist and is it important? Thorax 2014, 69, 760–765. [Google Scholar] [CrossRef] [PubMed]
- Khalil, N.; O’Connor, R.N.; Unruh, H.W.; Warren, P.W.; Flanders, K.C.; Kemp, A.; Bereznay, O.H.; Greenberg, A.H. Increased production and immunohistochemical localization of transforming growth factor-beta in idiopathic pulmonary fibrosis. Am. J. Respir. Cell Mol. Biol. 1991, 5, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Sime, P.J.; Xing, Z.; Graham, F.L.; Csaky, K.G.; Gauldie, J. Adenovector-mediated gene transfer of active transforming growth factor-beta1 induces prolonged severe fibrosis in rat lung. J. Clin. Investig. 1997, 100, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Doerner, A.M.; Zuraw, B.L. TGF-beta1 induced epithelial to mesenchymal transition (EMT) in human bronchial epithelial cells is enhanced by IL-1beta but not abrogated by corticosteroids. Respir. Res. 2009, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Z.; Pan, H.Y.; Wang, D.X.; Deng, Z.T.; Ye, X.L. TGF-beta1 induces human bronchial epithelial cell-to-mesenchymal transition in vitro. Lung 2009, 187, 187–194. [Google Scholar] [CrossRef]
- Kasai, H.; Allen, J.T.; Mason, R.M.; Kamimura, T.; Zhang, Z. TGF-beta1 induces human alveolar epithelial to mesenchymal cell transition (EMT). Respir. Res. 2005, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforming growth factor-beta l: Potential role in idiopathic pulmonary fibrosis. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef]
- Wu, Z.; Yang, L.; Cai, L.; Zhang, M.; Cheng, X.; Yang, X.; Xu, J. Detection of epithelial to mesenchymal transition in airways of a bleomycin induced pulmonary fibrosis model derived from an alpha-smooth muscle actin-Cre transgenic mouse. Respir. Res. 2007, 8, 1. [Google Scholar] [CrossRef]
- Mauderly, J.L.; Tesarek, J.E.; Sifford, L.J.; Sifford, L.J. Respiratory measurements of unsedated small laboratory mammals using nonrebreathing valves. Lab. Anim. Sci. 1979, 29, 323–329. [Google Scholar]
- Naota, M.; Mukaiyama, T.; Shimada, A.; Yoshida, A.; Okajima, M.; Morita, T.; Inoue, K.; Takano, H. Pathological study of acute pulmonary toxicity induced by intratracheally instilled Asian sand dust (kosa). Toxicol. Pathol. 2010, 38, 1099–1110. [Google Scholar] [CrossRef]
- Khalil, N.; Bereznay, O.; Sporn, M.; Greenberg, A.H. Macrophage production of transforming growth factor beta and fibroblast collagen synthesis in chronic pulmonary inflammation. J. Exp. Med. 1989, 170, 727–737. [Google Scholar] [CrossRef] [PubMed]
- Kapanci, Y.; Desmouliere, A.; Pache, J.C.; Redard, M.; Gabbiani, G. Cytoskeletal protein modulation in pulmonary alveolar myofibroblasts during idiopathic pulmonary fibrosis. Possible role of transforming growth factor beta and tumor necrosis factor alpha. Am. J. Respir. Crit. Care Med. 1995, 152, 2163–2169. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.O.; Flanders, K.C.; Saffiotti, U. Immunohistochemical localization of transforming growth factor-beta 1 in rats with experimental silicosis, alveolar type II hyperplasia, and lung cancer. Am. J. Pathol. 1993, 142, 1831–1840. [Google Scholar] [PubMed]
- Yamada, M.; Kuwano, K.; Maeyama, T.; Hamada, N.; Yoshimi, M.; Nakanishi, Y.; Kasper, M. Dual-immunohistochemistry provides little evidence for epithelial-mesenchymal transition in pulmonary fibrosis. Histochem. Cell Biol. 2008, 129, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, F.; Sun, Y.; Yuan, Y.; Cheng, H.; Wei, Z.; Li, S.; Cheng, T.; Brann, D.; Wang, R. A new antifibrotic target of Ac-SDKP: Inhibition of myofibroblast differentiation in rat lung with silicosis. PLoS ONE 2012, 7, e40301. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Kim, T.J.; Peng, D.H.; Duan, D.; Gibbons, D.L.; Yamauchi, M.; Jackson, J.R.; Le Saux, C.J.; Calhoun, C.; Peters, J.; et al. Fibroblast-specific inhibition of TGF-β1 signaling attenuates lung and tumor fibrosis. J. Clin. Investig. 2017, 127, 3675–3688. [Google Scholar] [CrossRef]
- Rong, Y.; Shen, Y.; Zhang, Z.; Cui, X.; Xiao, L.; Liu, Y.; Luo, X.; Chen, W. Blocking TGF-β expression inhibits silica particle-induced epithelial-mesenchymal transition in human lung epithelial cells. Environ. Toxicol. Pharmacol. 2015, 40, 861–869. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komai, M.; Mihira, K.; Shimada, A.; Miyamoto, I.; Ogihara, K.; Naya, Y.; Morita, T.; Inoue, K.; Takano, H. Pathological Study on Epithelial-Mesenchymal Transition in Silicotic Lung Lesions in Rat. Vet. Sci. 2019, 6, 70. https://doi.org/10.3390/vetsci6030070
Komai M, Mihira K, Shimada A, Miyamoto I, Ogihara K, Naya Y, Morita T, Inoue K, Takano H. Pathological Study on Epithelial-Mesenchymal Transition in Silicotic Lung Lesions in Rat. Veterinary Sciences. 2019; 6(3):70. https://doi.org/10.3390/vetsci6030070
Chicago/Turabian StyleKomai, Mao, Karin Mihira, Akinori Shimada, Ikumi Miyamoto, Kikumi Ogihara, Yuko Naya, Takehito Morita, Kenichiro Inoue, and Hirohisa Takano. 2019. "Pathological Study on Epithelial-Mesenchymal Transition in Silicotic Lung Lesions in Rat" Veterinary Sciences 6, no. 3: 70. https://doi.org/10.3390/vetsci6030070
APA StyleKomai, M., Mihira, K., Shimada, A., Miyamoto, I., Ogihara, K., Naya, Y., Morita, T., Inoue, K., & Takano, H. (2019). Pathological Study on Epithelial-Mesenchymal Transition in Silicotic Lung Lesions in Rat. Veterinary Sciences, 6(3), 70. https://doi.org/10.3390/vetsci6030070