1. Introduction
Just-hatched broiler chickens can experience a prolonged delay in access to nutrition for up to 72 h (delayed nutrition; DN), especially in the case of long post-hatch transport (for review, see [
1]). Broilers that received early nutrition (EN) and thus immediate provision of nutrition, post hatch onwards, showed enhanced immune activation and maturation compared to DN broilers; reviewed by [
2]. This is likely caused by early exposure of the immune system to antigens derived from commensal microbiota or ingested feed [
3,
4]. Thus, broilers receiving EN compared to DN, are expected to be exposed to a higher and more diverse load of antigens, derived from ingested feed and subsequent microbial colonization [
5,
6,
7]. In chicken, antibody responses towards antigens were reduced when these antigens were fed within the first 72 h after hatch (“window of opportunity”), which is known as oral tolerance [
8]. Interestingly, this window of opportunity parallels with the application of different feeding strategies (EN versus DN), during the first 72 h after hatch. Hence, it could be postulated that increased antigenic exposure as a result of EN, compared to DN, might affect the development of oral tolerance, resulting in altered later life antibody responses.
Indeed, EN versus DN broilers that were housed under relatively high antigenic pressure (floor housing) had lower IgY in blood plasma and growth depression, after immunization with LPS and HuSA, as compared to broilers housed under relatively low antigenic pressure (cage housing) [
9]. These authors suggested that effects of EN compared with DN on antibody responses might be affected by antigenic pressure. Studies comparing the later life immune system between EN and DN broilers under lower antigenic pressure, observed no differences among treatments [
9,
10]. This suggests that later life immune responses might be regulated by EN, but only when kept in a high antigenic pressure environment. Hence, we hypothesized that better regulation of antibody responses in EN compared to DN broilers, would be beneficial when the broilers are housed under high antigenic pressure. To test this hypothesis, we executed two consecutive experiments to understand the effects of feeding strategy (EN versus DN) and its interaction with antigenic pressure on later life antibody responses. The aim of experiment I was to test, under low antigenic pressure, whether EN might modulate later life antibody responses. In experiment II, we modeled contrasts in antigenic pressure by creating low (LSC) and high sanitary conditions (HSC). This allowed us to test whether the effects of feeding strategy on regulation of antibody responses and growth performance, depend on environmental antigenic pressure.
2. Materials and Methods
2.1. Experimental Designs
The experiments and respective procedures were ethically approved, according to the Dutch law under application number AVD104002016441, and were performed in climate respiration chambers (CRC) at the experimental facility. In both experiments, Ross 308 males were collected from a commercial hatchery within 1 h after hatch, to minimize age differences, and were immediately transported to the experimental facility, within 20 min, in cardboard chick boxes kept at 20 °C. All ages are expressed as biological age [
11]. After arrival, the broilers received an ID neck tag, and were distributed within 2 h over the floor pens (1.1 × 1.8 m) that contained SoftCell (Agromed GmbH, Kremsmünster, Austria) as bedding material, covered with chicken paper during the first 3 d of age, to prevent litter uptake. In both experiments, light and climate settings were identical. From placement onwards, a 16-h light:8-h dark schedule was applied. Ambient temperature was set at 36 °C (55% relative humidity) at 0 d of age, and gradually reduced to 29 °C until 7 d, and then further gradually reduced to 18 °C (75% relative humidity) at 35 d. Relative humidity was set at 55% at the start of the experiment and gradually increased to 75% at 42 d. Levels of CO2 were maintained ≤2500 ppm and that of NH3 ≤ 20 ppm. In both experiments, broilers had ad libitum access to water and a commercial broiler diet, except for the DN chickens, which had no access to feed and water, during the first 72 h from placement onwards.
2.1.1. Experiment I
The experiment was conducted in a single CRC, and designed as a 2 × 2 factorial approach, with antigen feeding (BSA feeding, PBS feeding) and early life feeding strategy (EN, DN) as factors. The 4 treatments were distributed over 12 floor pens, each containing 4 broilers. Three surplus broilers were distributed over the treatments, resulting in a total of 13 broilers for all treatments, except for the EN–BSA-fed broilers (n = 12). Body weight (BW) of the individual broilers was measured weekly. Broilers were not vaccinated in the hatchery or during the study. Commercial pelletized broiler starter (0–14 d; digestible energy (DE): 2950 kcal/kg; total lysine: 12.2 g/kg) and finisher diets (14–42 d; DE: 3000 kcal/kg; total lysine: 11.3 g/kg) were fed.
2.1.2. Experiment II
The experiment was designed as a 2 × 2 factorial approach with early life feeding strategy (EN, DN) and sanitary conditions (LSC, HSC) as factors. Low sanitary conditions were induced by the introduction of used litter from commercial broiler farms (see
Section 2.2). The experiment was executed in 3 consecutive batches, to account for variations in antigenic pressure due to differences in health status of litter-donating farms, as previously demonstrated in a pig model (Van der Meer et al., 2016). Broilers (parent stock age: batch 1: 31 w, batch 2: 33 w, batch 3: 48 w) were housed in either an LSC or an HSC CRC, each containing 8 floor pens. Both CRC were completely identical in their set-up and were controlled for identical climate conditions (temperature, humidity, CO2, NH3) (Van der Meer, 2017). Body weight and feed intake were measured weekly until 33 d of age to calculate the average daily gain (ADG), feed intake (ADFI), and feed conversion ratio (FCR). Broilers were vaccinated against Newcastle disease at 3 d of age, but this was accidentally omitted in batch 1. Broilers received no other vaccinations in the hatchery or during the study. Commercial pelletized broiler starter (0–7 d; DE: 2850 kcal/kg; total lysine: 11.8 g/kg), grower (7–28 d; DE: 2900 kcal/kg; total lysine: 11.2 g/kg), and finisher diets (28–35 d; DE: 2950 kcal/kg; total lysine: 10.7 g/kg) were fed. The grower diet contained decoquinate (0.05 g/kg; Deccox 6%, Zoetis, Capelle aan den IJssel, The Netherlands).
2.2. Sanitary Conditions
In experiment I, the broilers were kept under HSC from placement onwards, as follows. The HSC chamber was cleaned with water and disinfected (Halamid, Veip Disinfectants, Wijk bij Duurstede, The Netherlands), following the manufacturer’s instructions, and over-pressurized (100 ± 5 Pa) to prevent the influx of external pathogens. A strict hygiene protocol (consisting of showering, cleaning and disinfection boots, wearing gloves and hairnet, and minimal pen entrance) was maintained. All procedures, except for dissection, were performed inside the chamber, to prevent introduction of novel antigens. In experiment II, all broilers were kept under HSC (similar to experiment I) until 3 d of age, after which the contrasts in sanitary conditions were made as follows. The HSC broilers were kept under HSC until the end of the experiment. Low sanitary conditions were induced as follows—the LSC chamber was under-pressurized (−65 ± 5 Pa) and no hygiene protocol was maintained. Coveralls and boots were not cleaned and disinfected. Seven days before onset of each respective batch, litter was obtained from 3 commercial broiler farms with flock age of at least 35 d. The litter was collected all at once, during the cleaning of the broiler houses, and after arrival at the experimental facility, the litter was pooled by weight and stored in 8–10 kg portions at 4 °C. From 3 d of age onwards, one portion (8–10 kg) of homogenized used litter was distributed in each pen, every 4 d.
2.3. Induction of Oral Tolerance and Immunizations
From placement until 3 d of age, broilers were orally fed with either BSA to induce tolerance (V = 0.25 mL; 100 mg/mL; Sigma Aldrich CO, St. Louis, MO, USA) or phosphate buffered saline (PBS) as a control, every 12 h. In experiment I, BSA or PBS was administered by pipetting the volume in the beak of each broiler. In experiment II, BSA was administered by oral gavage in the esophagus via a blunted needle. An intratracheal dose (V = 0.5 mL) containing 0.5 mg BSA and 0.5 mg RGG (rabbit γ-globulin, Sigma) was given at 21 and 22 d of age (experiment I; all broilers) or 24 d of age (experiment II; 2 broilers per pen), via a blunted needle, as a secondary immunization.
2.4. Sample Collection
In experiment I, blood was collected at 21, 24, 28, 35, and 42 d (0, 3, 7, 14, and 14 d post immunization (p.i.)) from the wing vein of all broilers in heparinized tubes, and subsequently centrifuged (12,000× g, 5 min) to obtain plasma. In experiment II, blood was collected at 14, 24, 29, and 33 d (−10, 0, 5, and 9 d p.i.) from 1 broiler per pen in tubes, incubated for 2 h at 4 °C, and subsequently centrifuged (12,000× g, 5 min) to obtain serum. Bile was collected from all broilers either at 42 d of age (experiment I) or 33 and 34 d of age (experiment II), as follows. After euthanizing the broiler by a percussive blow on the head, the bile bladder was located, and the contents were collected with a syringe and needle, and placed in tubes. The spleens were located, collected, and weighed. Plasma, sera, and bile were stored at −20 °C for further analyses. In experiment II, after euthanizing the broiler by decapitation, the whole right cecum with contents was collected within 6 h after hatch and at 1, 2, 3, and 7 d of age, from one broiler per pen. The cecum was weighed, placed in sterile 0.9% NaCl, and homogenized with a tissue homogenizer (IKA T50, IKA-Werke, Staufen, Germany). Afterwards, the homogenate was diluted 1:1 in sterile 15% glycerol, and stored at −80 °C, pending further analyses.
2.5. Bacterial Load in Cecum
All procedures were performed under aseptic conditions. The specific culture media MacConkey agar (MAC; CM0007), De Man, Rogosa, and Sharpe agar (MRS; CM0359), and blood agar (BA; OXOIPB5039A) were prepared according to the instructions of the supplier (Oxoid Microbiology Products, Hampshire, United Kingdom). After pouring the specific culture media, the homogenates were thawed at 20 °C and singular, 10-fold serial dilutions in sterile PBS were spread on the agar plates (100 µL/plate). Plates were incubated upside down at 37 °C in an incubator, for 36 h (BA, MAC) or 48 h (MRS). After incubation, the number of colony forming units (CFU) were counted and expressed per gram of collected cecal tissue.
2.6. Antibody Levels
Two-step, indirect, enzyme-linked immunosorbent assays (ELISA) were performed to measure IgM and IgY levels in plasma or serum, and IgA levels in bile, binding either BSA or rabbit γ-globulin (RGG). Flat-bottomed, 96-well medium binding plates (Greiner Bio-One) were coated with either 2.5 µg/mL BSA or 2.5 µg/mL RGG in 100-µl coating buffer (5.3 g/L Na2CO3, 4.2 g/L NaHCO3; pH 9.6), and incubated overnight at 4 °C. After washing with tap water containing 0.05% Tween® 20, plates were filled with 100 µl dilution buffer (PBS (10.26 g/L Na2HPO4·2H2O, 2.36 g/L KH2PO4, 4.5 g/L NaCl; pH 7.2) containing 0.5% normal horse serum and 0.05% Tween® 20). Prediluted samples (1:10 in dilution buffer) were further diluted to 1:40, 1:160, 1:640, and 1:2560, and a standard positive control was included, in duplicates, from the pooled plasma, serum, or bile, for each day. Plates were incubated for 1.5 h at 20 °C. After washing, the plates were incubated with 1:20.000-diluted goat-anti-chicken IgA, goat-anti-chicken IgM, or goat-anti-chicken IgY, all labelled with horse radish peroxidase (Bethyl Laboratories Inc, Montgomery, AL, USA), for 1.5 h at 20 °C. After washing, 100 µL substrate buffer (containing reverse osmosis purified water, 10% tetramethylbenzidine buffer (15.0 g/L sodium acetate, 1.43 g/L urea hydrogen peroxide; pH 5.5) and 1% tetramethylbenzidine (8 g/L DMSO) were added. After 15 min of incubation at 20 °C, the reaction was stopped with 50 µL of 1.25 M H2SO4 solution. The extinctions were measured with a Multiskan GO (Thermo scientific, Breda, The Netherlands) at 450 nm. Titers were expressed as log2 values of the dilutions that gave an extinction closest to 50% of Emax, where Emax represents the highest mean extinction of the standard positive.
2.7. Statistical Analyses
Data were processed, analyzed, and presented using R version 3.6.1 [
12]. Linear mixed models were established with the nlme package version 3.1–140 [
13]. Model residuals of the linear models were tested to verify the assumptions of normality and homogeneity by QQ-plots and residual plots. Differences among means with
p ≤ 0.05 were considered to be statistically significant, and
p ≤ 0.10 were considered to be tendencies. All data are presented as (back-transformed) estimated marginal means with standard errors, unless specified otherwise.
2.7.1. Experiment I
General linear models were established to estimate the effects of BSA feeding (PBS fed, BSA fed), feeding (DN, EN), and their interaction on antibody levels, binding either BSA or RGG in plasma (IgM, IgY) on 0, 14, 21 d p.i., fold change (ratio between IgM or IgY levels at 7 versus 0 d p.i.), IgA levels in bile (21 d p.i.), BW (0, 21, 24, 42 d), and ADG (0–24, 21–24, 24–42, 0–42) with individual broiler within the pen as the experimental unit, and pen as the random effect. Logarithmic transformation was applied to normalize the residuals on all titer and performance data, except for the IgA binding BSA and RGG.
2.7.2. Experiment II
General linear models were established to estimate the effects of sanitary condition (LSC, HSC), feeding (DN, EN), and their interaction on antibody levels, binding either BSA or RGG in serum (IgM, IgY) on −10, 0, 7 d p.i., and fold change (ratio 5:0 levels d p.i.; IgM, IgY), with pen as the experimental unit. Batch (1, 2, 3) and its interaction with treatment effects was used as the blocking factor. Comparable models were used to analyze the levels of bile and spleen weights among treatments. As bile and spleens were collected on 2 consecutive days (10, 11 d p.i.), sampling day was added as a blocking factor. Day was found to not be significant and therefore was eliminated from the models. Effects of treatments on BW (3, 33 d), ADG (3–14; 14–28; 28–33 d), and FCR (3–14; 14–28; 28–33 d) were estimated with general linear models, using sanitary condition, feeding, and their respective interaction as fixed effects, with pen as the experimental unit. Fixed effects of treatments (EN, DN) on bacterial colonization in ceca were analyzed using the non-parametric Kruskal-Wallis test, separately for each day (0, 1, 2, 3, 7). For day 7, effects of sanitary condition (HSC, LSC) were also analyzed. Logarithmic transformation was applied to normalize the residuals of all titer and performance data. Interactions between batch and other main effects (feeding, sanitary condition, feeding * sanitary condition) were tested but excluded from the model if p > 0.10.
4. Discussion
We studied whether antibody responses of the broilers are influenced (either increased or decreased) by early life feeding strategy (EN versus DN) and sanitary conditions (LSC versus HSC), in two consecutive experiments. First, we studied whether feeding BSA during the first 3 d of life (“window of opportunity”) affects later life antibody responses and its interaction with feeding strategy, through the development of oral tolerance. Second, we tested the effects of EN on (regulation of) antibody immune responses and growth performance, under either HSC or LSC. We observed no effects of the feeding strategy on antibody responses, irrespective of sanitary conditions. Broilers kept under LSC, as compared to HSC, showed higher levels of natural antibodies (NAb) and a smaller fold-change of specific antibodies (SpAb) after BSA and RGG immunization, and a reduced growth performance.
4.1. Aerobic Bacterial Colonization after Early Nutrition
Accelerated maturation of the immune system after EN as compared to DN, was suggested to be caused by enhanced bacterial colonization of the intestinal tract [
5,
6,
7]. Short-term effects of EN compared to DN, were found on the intestinal bacterial load [
16]. Data on the effects of EN compared to DN, on intestinal bacterial composition, suggest temporal differences in the ileal bacterial composition, up to 9 d of age [
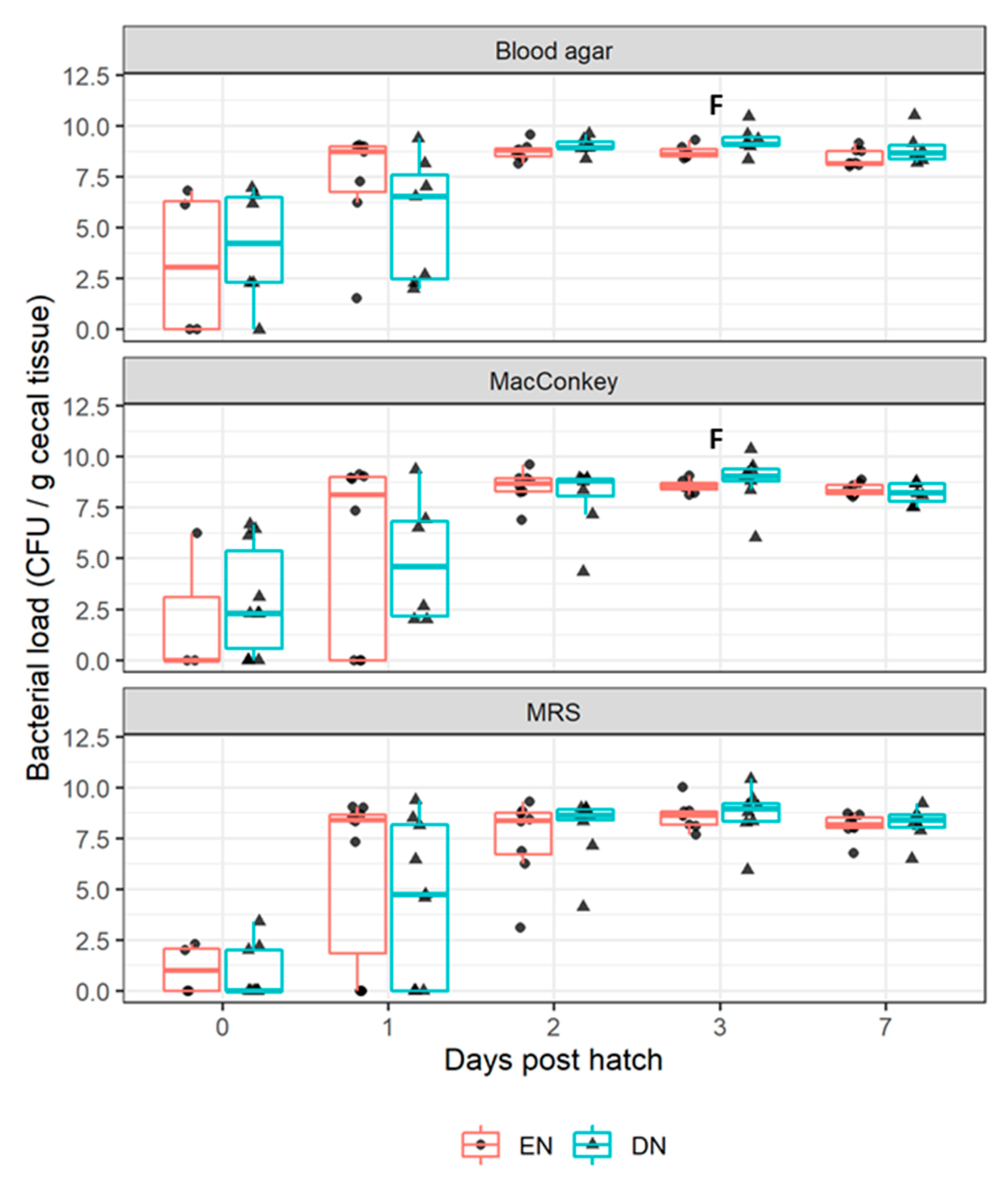
16]. To relate these effects to antibody responses, we analyzed the colonization dynamics of culturable aerobic bacteria in ceca, between EN and DN broilers, in the first week post-hatch (experiment II).
The intestinal tract of broilers was rapidly colonized by bacteria, within the first days after hatch and the total number of colonizing bacteria stabilized around 3 d of age, which was in accordance with previous research reviewed by [
17]. Relatively high within-treatment group variation of up to 2 (BA, MAC) or 3 (MRS) d of age, might suggest divergent colonization patterns among individuals. We observed no significant effects of feeding strategy (EN versus DN) up to 3 d of age on bacterial colonization, suggesting that EN, compared to DN, minimally affected aerobic bacterial load. In EN, compared to DN broilers, the aerobic bacterial load tended to be lower at 3 d, which was also previously observed by others [
5]. Speculatively, this might reflect earlier replacement of aerobes (
E. coli,
Enterococcus spp.) by obligate anaerobic species, after EN, as suggested before [
5].
4.2. Early-Life BSA Feeding Lowers Later-Life Antibody Responses Towards BSA under High Sanitary Conditions
In experiment I, we first tested whether early-life oral exposure towards antigens affected later-life antibody responses. Therefore, BSA or PBS (negative control) was orally administered during the first 3 d of age. At 21 d of age, broilers received an i.t. BSA immunization, following procedures derived from the oral tolerance model in laying hens [
8]. Although broilers are known to be immunologically different from laying hens [
18,
19,
20], our study demonstrated that the mechanism of oral tolerance also exist in broilers. Thus, our findings confirmed that after immunization at 21 d in broilers, antibody responses are lowered towards antigens that were fed in the first 3 d after hatch [
8,
21,
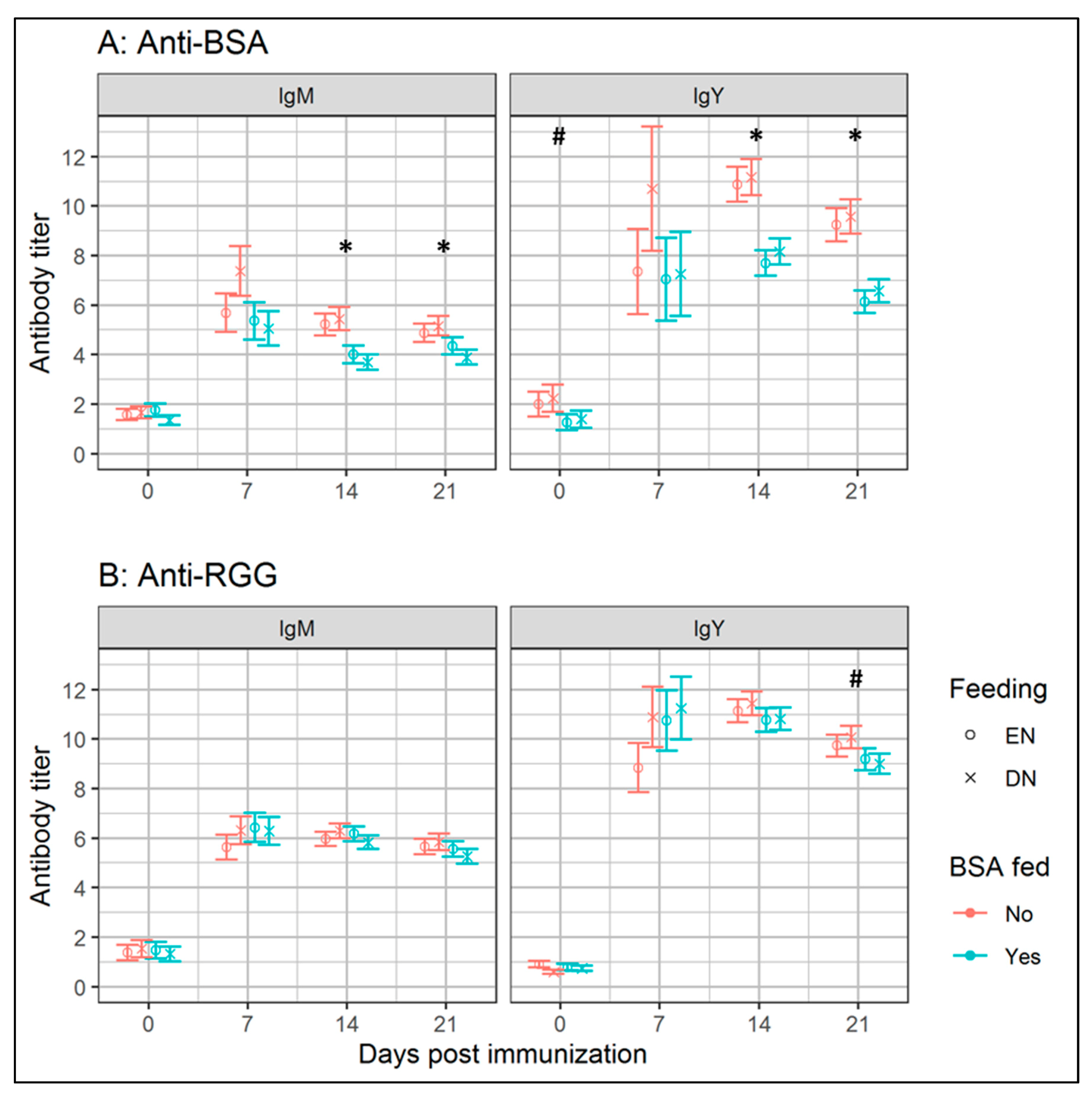
22]. We also investigated whether BSA feeding in early-life (0–3 d of age) affect SpAb responses to a non-BSA related antigen. Therefore, broilers were immunized i.t. with RGG at 21 d of age, to test whether antibody responses differed between the BSA and PBS-fed groups. We observed that systemic IgM and IgY (
Figure 1B) and biliary IgA (
Table 1) antibodies binding RGG, were unaffected (
p ≥ 0.05) by BSA feeding. Thus, early-life oral antigen exposure affected antibody responses in a specific fashion towards the specifically fed antigen, and not to novel, unrelated antigens. Effects of BSA feeding on IgA responses in bile were studied at 21 d p.i., as an indicator for immune regulation—IgA is present at the mucosal surfaces and contributes to immune homeostasis by binding antigens in the intestinal lumen and coating intestinal bacteria [
23,
24,
25]. Our finding that BSA feeding results in higher levels of IgA binding BSA, suggests that early-life antigen exposure also affects IgA responses. In summary, we demonstrated that the specific IgA response is increased, and specific IgM and IgY responses are lowered, by oral exposure to antigen (BSA), during the first 3 d of age.
The main objective of experiment I was to test whether the feeding strategy (EN versus DN) affected later-life antibody responses towards BSA and RGG in BSA-fed broilers. We observed no effects of feeding strategy on systemic antibody responses towards BSA or RGG, indicating that immediate provision of nutrition after hatch (EN) did not interfere with antibody responses at later-life. Irrespective of BSA feeding, we observed higher levels of biliary IgA-binding BSA in EN broilers, suggesting that EN contributes to a more anti-inflammatory immune status in EN broilers. We would like to emphasize that experiment I was conducted under HSC. Limited antigenic pressure in experiment I might explain the lack of differences in IgM and IgY responses between EN and DN broilers, as less regulation of immune responses was required. Therefore, we designed a second experiment where the broilers were kept under LSC or HSC.
4.3. Introduction of Low Sanitary Conditions
To model contrasts in antigenic pressure in experiment II, we introduced either LSC and HSC from 3 d of age onwards, according to a pig model [
26,
27]. As litter from 3 different broiler farms per batch was used in LSC, we considered the obtained antigenic pressure to be illustrative for the range of antigens found in typical commercial broiler farms. Throughout the experiment, growth performance of broilers was depressed in LSC compared to the HSC groups. Relative spleen weights tended to be greater (
p = 0.09) at 33 d of age in LSC (0.26 ± 0.09), compared to the HSC broilers (0.11 ± 0.09), indicating greater activity of the immune system [
28]. From these data, we conclude that LSC was successfully induced. Batch and batch × treatment effects were present in this broiler model, which was in line with the pig model [
29]. We observed interactions (
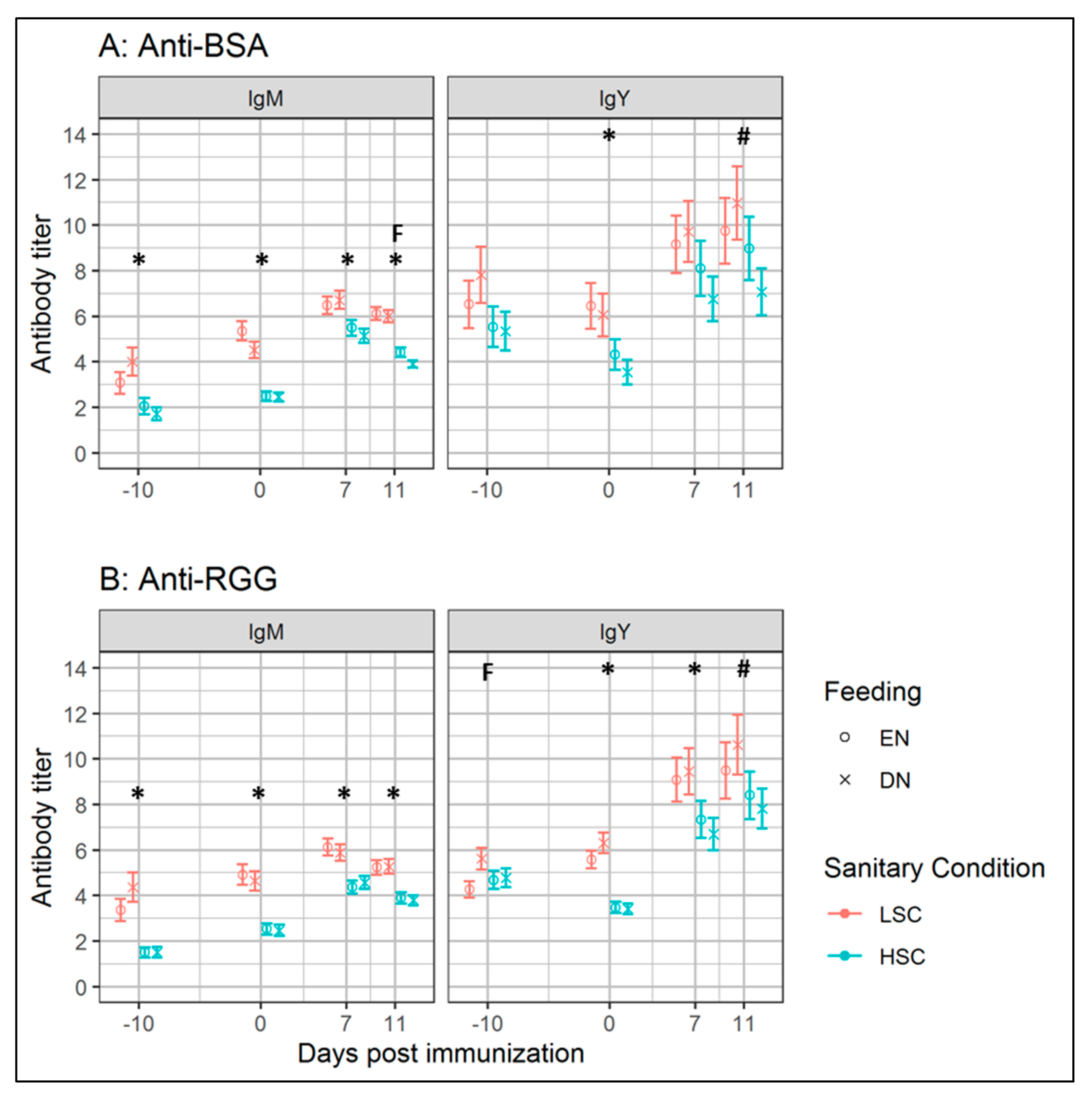
p ≤ 0.05) between batch and sanitary conditions for IgM binding BSA or RGG at 14 d/−10 d p.i., as well for BW (33 d of age), ADG (all phases), FCR (3–14 d), and standardized FCR. The observed batch effects did not change the direction, but only changed the magnitude of the observed effects. Hence, the batches were not analyzed separately.
4.4. Antibody Responses and Performance after Early Nutrition under Different Sanitary Conditions
In experiment II, we aimed to compare antibody responses and growth performance of the EN and DN broilers kept under either LSC or HSC. We could not confirm our hypothesis that EN versus DN broilers housed under LSC, differed in growth performance or antibody responses after BSA and RGG immunization. This was unexpected, as lower IgY responses and enhanced performance in EN compared to the DN broilers were reported in a previous study [
30]. In that study, however, broilers were immunized at one time-point, with a combination of the model antigens HuSA and LPS, at 28 d of age, while in our study, LSC broilers were continuously subjected to high antigenic pressure from 3 d of age onwards. In experiment II, we observed higher BSA-specific IgA levels in EN compared to DN broilers in batch 2, but not in other batches (batch × feeding strategy interaction;
p = 0.06). Levels of IgA-binding RGG were found to be higher in EN compared to the DN broilers, which was in contrast with experiment I, where we observed no effects of feeding strategy. As the titer differences (Δ = 0.3) between the EN and DN groups were relatively small in experiment I, it was unclear if the observed effects were biologically relevant. In summary, we concluded that EN compared to DN, minimally affected antibody responses, regardless of sanitary conditions, although the effects of feeding strategy on biliary IgA (both BSA and RGG) remained debatable.
4.5. Low Sanitary Conditions Increased Antibody Levels
Broilers housed under LSC from 3 d onwards, appeared to adapt towards the higher antigenic pressure from 14 d of age onwards. Levels of Nab-binding BSA or RGG before immunization with BSA and RGG, were higher in LSC, compared to the HSC broilers, at 14 d (−10 d p.i.; IgM) and 24 d (0 d p.i.; IgM, IgY) of age. Higher levels of natural IgM and the interaction between batch and sanitary condition on IgM levels, suggest that IgM NAb levels were affected by antigenic pressure from, at least, 14 d of age. Broilers kept under HSC, compared to LSC, showed a higher fold-change of IgM-binding BSA or RGG (
p ≤ 0.01), and IgY-binding RGG (
p = 0.07), after immunization. It is tempting to speculate that the higher levels of NAb and lower fold-change of SpAb after immunization in LSC broilers might indicate different defense strategies of the immune system. Whereas LSC broilers might depend on a less specific antibody (NAb) repertoire as a first line of defense, broilers under HSC need to generate higher specific immune responses (SpAb) after antigenic stimulation. Higher NAb levels, and lower fold-change of specific antibodies, were reported in layer hens that were continuously immunized with other antigens, compared to the non-immunized hens [
30]. The observed higher levels of biliary IgA-binding BSA or RGG (both
p < 0.001) in LSC, compared to HSC broilers, indicate increased protection of mucosal surfaces in LSC broilers [
23,
24]. This suggests that the immune system prevents innocuous responses to antigens during higher antigenic pressure through increasing IgA levels.

 ,
, _Lammers.jpeg)




{kind=link}
{kind=link}
{kind=link}