The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next?



{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Staphylococcus pseudintermedius: A Pathogenic Bacterium of Veterinary Concern
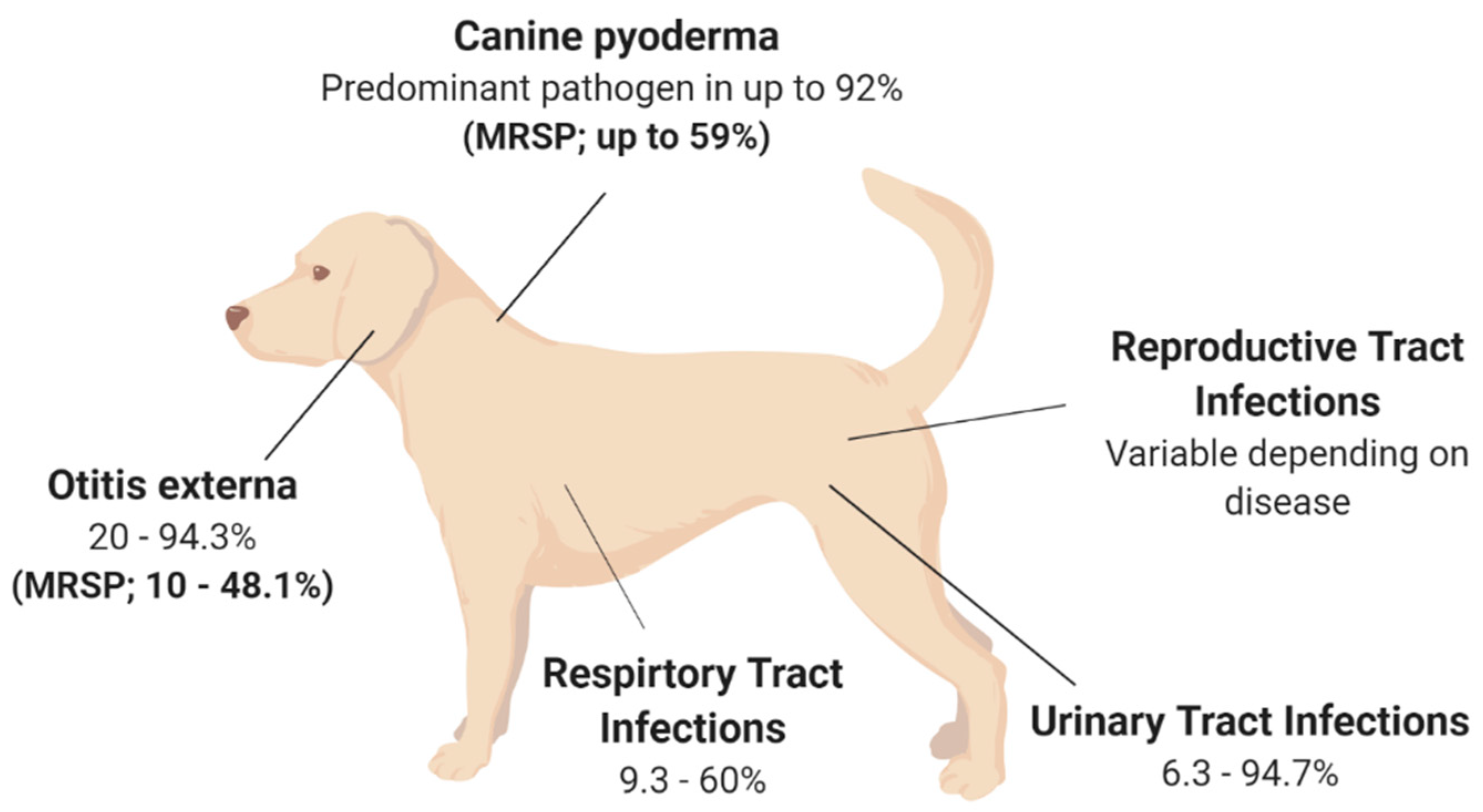
2.1. Canine Pyoderma
2.2. Otitis Externa
2.3. Urinary Tract Infections (UTI)
2.4. Respiratory Infections
2.5. Reproductive Tract Infections

3. Current and Future Treatment Options for Staphylococcus pseudintermedius in Canines
3.1. Human Antibiotics
3.2. Topical Therapies
3.3. Vaccines
3.3.1. Bacterins
3.3.2. Subunit
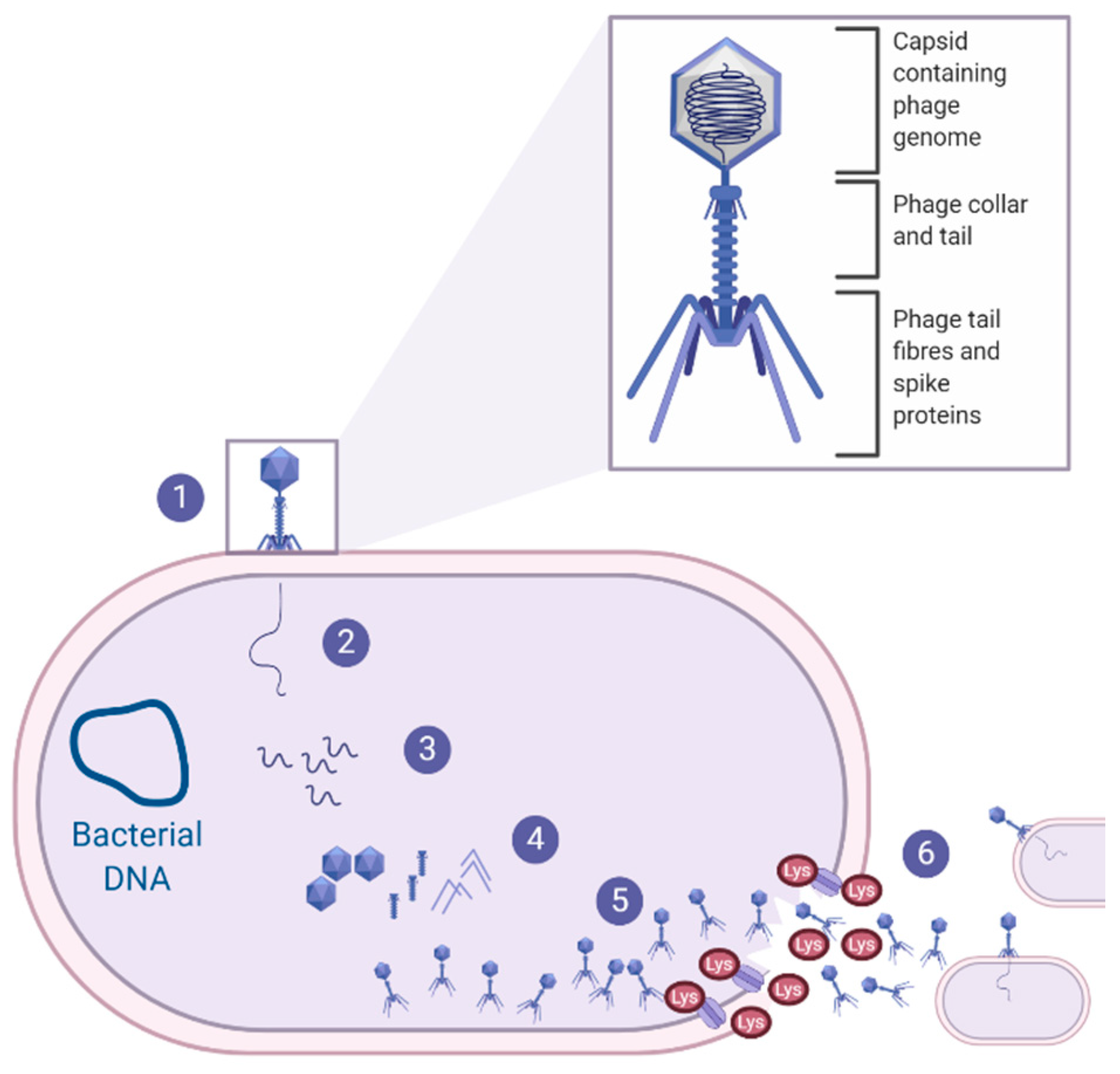
3.4. Phage Therapy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253–266,e51-2. [Google Scholar] [CrossRef] [PubMed]
- Ruscher, C.; Lübke-Becker, A.; Wleklinski, C.-G.; Soba, A.; Wieler, L.H.; Walther, B. Prevalence of Methicillin-resistant Staphylococcus pseudintermedius isolated from clinical samples of companion animals and equidaes. Vet. Microbiol. 2009, 136, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Wegener, A.; Broens, E.M.; Zomer, A.; Spaninks, M.; Wagenaar, J.A.; Duim, B. Comparative genomics of phenotypic antimicrobial resistances in methicillin-resistant Staphylococcus pseudintermedius of canine origin. Vet. Microbiol. 2018, 225, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Worthing, K.A.; Abraham, S.; Coombs, G.W.; Pang, S.; Saputra, S.; Jordan, D.; Trott, D.J.; Norris, J.M. Clonal diversity and geographic distribution of methicillin-resistant Staphylococcus pseudintermedius from Australian animals: Discovery of novel sequence types. Vet. Microbiol. 2018, 213, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Hajek, V. Staphylococcus intermedius, a new species isolated from animals. Int. J. Syst. Evol. Microbiol. 1976, 26, 401–408. [Google Scholar] [CrossRef]
- Devriese, L.A.; Vancanneyt, M.; Baele, M.; Vaneechoutte, M.; De Graef, E.; Snauwaert, C.; Cleenwerck, I.; Dawyndt, P.; Swings, J.; Decostere, A. Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. Int. J. Syst. Evol. Microbiol. 2005, 55, 1569–1573. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Kikuchi, K.; Tanaka, Y.; Takahashi, N.; Kamata, S.; Hiramatsu, K. Reclassification of phenotypically identified Staphylococcus intermedius strains. J. Clin. Microbiol. 2007, 45, 2770–2778. [Google Scholar] [CrossRef] [Green Version]
- Mališová, L.; Šafránková, R.; Kekláková, J.; Petráš, P.; Žemličková, H.; Jakubů, V. Correct species identification (reclassification in CNCTC) of strains of Staphylococcus intermedius-group can improve an insight into their evolutionary history. Folia Microbiol. 2019, 64, 231–236. [Google Scholar] [CrossRef]
- Bannoehr, J.; Franco, A.; Iurescia, M.; Battisti, A.; Fitzgerald, J.R. Molecular diagnostic identification of Staphylococcus pseudintermedius. J. Clin. Microbiol. 2009, 47, 469–471. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Kawakami, T.; Fukata, T.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive Staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Chrobak, D.; Kizerwetter-Świda, M.; Rzewuska, M.; Moodley, A.; Guardabassi, L.; Binek, M. Molecular characterization of Staphylococcus pseudintermedius strains isolated from clinical samples of animal origin. Folia Microbiol. 2011, 56, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.C.; Worthing, K.A.; Ward, M.P.; Norris, J.M. Commensal Staphylococci Including Methicillin-Resistant Staphylococcus aureus from Dogs and Cats in Remote New South Wales, Australia. Microb. Ecol. 2020, 79, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.C.; Bärgman, S.C.; Moodley, A.; Nielsen, S.S.; Guardabassi, L. Staphylococcus pseudintermedius colonization patterns and strain diversity in healthy dogs: A cross-sectional and longitudinal study. Vet. Microbiol. 2012, 160, 420–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iverson, S.A.; Brazil, A.M.; Ferguson, J.M.; Nelson, K.; Lautenbach, E.; Rankin, S.C.; Morris, D.O.; Davis, M.F. Anatomical patterns of colonization of pets with Staphylococcal species in homes of people with methicillin-resistant Staphylococcus aureus (MRSA) skin or soft tissue infection (SSTI). Vet. Microbiol. 2015, 176, 202–208. [Google Scholar] [CrossRef]
- van Duijkeren, E.; Catry, B.; Greko, C.; Moreno, M.A.; Pomba, M.C.; Pyörälä, S.; Ruzauskas, M.; Sanders, P.; Threlfall, E.J.; Torren-Edo, J.; et al. Review on methicillin-resistant Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 2011, 66, 2705–2714. [Google Scholar] [CrossRef] [Green Version]
- Hartantyo, S.H.P.; Chau, M.L.; Fillon, L.; Ariff, A.Z.B.M.; Kang, J.S.L.; Aung, K.T.; Gutiérrez, R.A. Sick pets as potential reservoirs of antibiotic-resistant bacteria in Singapore. Antimicrob. Resist. Infect. Control 2018, 7, 106. [Google Scholar] [CrossRef]
- Laarhoven, L.M.; de Heus, P.; van Luijn, J.; Duim, B.; Wagenaar, J.A.; van Duijkeren, E. Longitudinal study on methicillin-resistant Staphylococcus pseudintermedius in households. PLoS ONE 2011, 6, e27788. [Google Scholar] [CrossRef]
- van Duijkeren, E.; Kamphuis, M.; van der Mije, I.C.; Laarhoven, L.M.; Duim, B.; Wagenaar, J.A.; Houwers, D.J. Transmission of methicillin-resistant Staphylococcus pseudintermedius between infected dogs and cats and contact pets, humans and the environment in households and veterinary clinics. Vet. Microbiol. 2011, 150, 338–343. [Google Scholar] [CrossRef]
- Windahl, U.; Gren, J.; Holst, B.S.; Börjesson, S. Colonization with methicillin-resistant Staphylococcus pseudintermedius in multi-dog households: A longitudinal study using whole genome sequencing. Vet. Microbiol. 2016, 189, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Feßler, A.T.; Schuenemann, R.; Kadlec, K.; Hensel, V.; Brombach, J.; Murugaiyan, J.; Oechtering, G.; Burgener, I.A.; Schwarz, S. Methicillin-resistant Staphylococcus aureus (MRSA) and methicillin-resistant Staphylococcus pseudintermedius (MRSP) among employees and in the environment of a small animal hospital. Vet. Microbiol. 2018, 221, 153–158. [Google Scholar] [CrossRef]
- Loeffler, A.; Lloyd, D.H. What has changed in canine pyoderma? A narrative review. Vet. J. 2018, 235, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffeth, G.C.; Morris, D.O.; Abraham, J.L.; Shofer, F.S.; Rankin, S.C. Screening for skin carriage of methicillin-resistant coagulase-positive Staphylococci and Staphylococcus schleiferi in dogs with healthy and inflamed skin. Vet. Dermatol. 2008, 19, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Bryan, J.; Frank, L.A.; Rohrbach, B.W.; Burgette, L.J.; Cain, C.L.; Bemis, D.A. Treatment outcome of dogs with meticillin-resistant and meticillin-susceptible Staphylococcus pseudintermedius pyoderma. Vet. Dermatol. 2012, 23, 361-e65. [Google Scholar] [CrossRef]
- Huerta, B.; Maldonado, A.; Ginel, P.J.; Tarradas, C.; Gómez-Gascón, L.; Astorga, R.J.; Luque, I. Risk factors associated with the antimicrobial resistance of Staphylococci in canine pyoderma. Vet. Microbiol. 2011, 150, 302–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.-H.; Yoon, J.W.; Lee, S.-Y.; Park, H.-M. High prevalence of Fluoroquinolone- and Methicillin-resistant Staphylococcus pseudintermedius isolates from canine pyoderma and otitis externa in veterinary teaching hospital. J. Microbiol. Biotechnol. 2010, 20, 798–802. [Google Scholar]
- Fazakerley, J.; Williams, N.; Carter, S.; McEwan, N.; Nuttall, T. Heterogeneity of Staphylococcus pseudintermedius isolates from atopic and healthy dogs. Vet. Dermatol. 2010, 21, 578–585. [Google Scholar] [CrossRef]
- Shimizu, A.; Berkhoff, H.A.; Kloos, W.E.; George, C.G.; Ballard, D.N. Genomic DNA fingerprinting, using pulsed-field gel electrophoresis, of Staphylococcus intermedius isolated from dogs. Am. J. Vet. Res. 1996, 57, 1458–1462. [Google Scholar]
- Hesselbarth, J.; Witte, W.; Cuny, C.; Rohde, R.; Amtsberg, G. Characterization of Staphylococcus intermedius from healthy dogs and cases of superficial pyoderma by DNA restriction endonuclease patterns. Vet. Microbiol. 1994, 41, 259–266. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, J.; Logue, C.M.; Liu, K.; Cao, X.; Zhang, W.; Shen, J.; Wu, C. Methicillin-resistant Staphylococcus pseudintermedius isolated from canine pyoderma in North China. J. Appl. Microbiol. 2012, 112, 623–630. [Google Scholar] [CrossRef]
- Pires Dos Santos, T.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic Review on Global Epidemiology of Methicillin-Resistant Staphylococcus pseudintermedius: Inference of Population Structure from Multilocus Sequence Typing Data. Front. Microbiol. 2016, 7, 1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hensel, N.; Zabel, S.; Hensel, P. Prior antibacterial drug exposure in dogs with meticillin-resistant Staphylococcus pseudintermedius (MRSP) pyoderma. Vet. Dermatol. 2016, 27, 72-e20. [Google Scholar] [CrossRef] [Green Version]
- Dziva, F.; Wint, C.; Auguste, T.; Heeraman, C.; Dacon, C.; Yu, P.; Koma, L.M. First identification of methicillin-resistant Staphylococcus pseudintermedius strains among coagulase-positive staphylococci isolated from dogs with otitis externa in Trinidad, West Indies. Infect. Ecol. Epidemiol. 2015, 5, 29170. [Google Scholar] [CrossRef] [PubMed]
- Lyskova, P.; Vydrzalova, M.; Mazurova, J. Identification and antimicrobial susceptibility of bacteria and yeasts isolated from healthy dogs and dogs with otitis externa. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2007, 54, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Bugden, D.L. Identification and antibiotic susceptibility of bacterial isolates from dogs with otitis externa in Australia. Aust. Vet. J. 2013, 91, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Paterson, S. Discovering the causes of otitis externa. Practice 2016, 38, 7–11. [Google Scholar] [CrossRef] [Green Version]
- Zur, G.; Lifshitz, B.; Bdolah-Abram, T. The association between the signalment, common causes of canine otitis externa and pathogens. J. Small Anim. Pract. 2011, 52, 254–258. [Google Scholar] [CrossRef]
- Saridomichelakis, M.N.; Farmaki, R.; Leontides, L.S.; Koutinas, A.F. Aetiology of canine otitis externa: A retrospective study of 100 cases. Vet. Dermatol. 2007, 18, 341–347. [Google Scholar] [CrossRef]
- Ngo, J.; Taminiau, B.; Fall, P.A.; Daube, G.; Fontaine, J. Ear canal microbiota—A comparison between healthy dogs and atopic dogs without clinical signs of otitis externa. Vet. Dermatol. 2018, 29, 425-e140. [Google Scholar] [CrossRef]
- Zur, G.; Gurevich, B.; Elad, D. Prior antimicrobial use as a risk factor for resistance in selected Staphylococcus pseudintermedius isolates from the skin and ears of dogs. Vet. Dermatol. 2016, 27, 468-e125. [Google Scholar] [CrossRef]
- Perry, L.R.; MacLennan, B.; Korven, R.; Rawlings, T.A. Epidemiological study of dogs with otitis externa in Cape Breton, Nova Scotia. Can. Vet. J. 2017, 58, 168–174. [Google Scholar]
- Chan, W.Y.; Hickey, E.E.; Khazandi, M.; Page, S.W.; Trott, D.J.; Hill, P.B. In vitro antimicrobial activity of narasin against common clinical isolates associated with canine otitis externa. Vet. Dermatol. 2018, 29, 149-e57. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.X.F.; Khazandi, M.; Chan, W.Y.; Trott, D.J.; Deo, P. Antimicrobial activity of thyme oil, oregano oil, thymol and carvacrol against sensitive and resistant microbial isolates from dogs with otitis externa. Vet. Dermatol. 2019, 30, 524-e159. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Zhelev, G.; Marutsov, P.; Koev, K.; Georgieva, S.; Toneva, I.; Urumova, V. Microbiological and antibacterial resistance profile in canine otitis externa—A comparative analysis. Bulg. J. Vet. Med. 2019, 22, 447–456. [Google Scholar] [CrossRef]
- Roberts, M.; White, J.; Lam, A. Prevalence of bacteria and changes in trends in antimicrobial resistance of Escherichia coli isolated from positive canine urinary samples from an Australian referral hospital over a 5-year period (2013–2017). Vet. Rec. Open 2019, 6, e000345. [Google Scholar] [CrossRef] [Green Version]
- Windahl, U.; Holst, B.S.; Nyman, A.; Grönlund, U.; Bengtsson, B. Characterisation of bacterial growth and antimicrobial susceptibility patterns in canine urinary tract infections. BMC Vet. Res. 2014, 10, 217. [Google Scholar] [CrossRef] [Green Version]
- Penna, B.; Varges, R.; Martins, R.; Martins, G.; Lilenbaum, W. In vitro antimicrobial resistance of Staphylococci isolated from canine urinary tract infection. Can. Vet. J. 2010, 51, 738–742. [Google Scholar]
- Ling, G.V.; Norris, C.R.; Franti, C.E.; Eisele, P.H.; Johnson, D.L.; Ruby, A.L.; Jang, S.S. Interrelations of organism prevalence, specimen collection method, and host age, sex, and breed among 8,354 canine urinary tract infections (1969–1995). J. Vet. Intern. Med. 2001, 15, 341–347. [Google Scholar]
- Gatoria, I.S.; Saini, N.S.; Rai, T.S.; Dwivedi, P.N. Comparison of three techniques for the diagnosis of urinary tract infections in dogs with urolithiasis. J. Small Anim. Pract. 2006, 47, 727–732. [Google Scholar] [CrossRef]
- Ball, K.R.; Rubin, J.E.; Chirino-Trejo, M.; Dowling, P.M. Antimicrobial resistance and prevalence of canine uropathogens at the Western College of Veterinary Medicine Veterinary Teaching Hospital, 2002–2007. Can. Vet. J. 2008, 49, 985–990. [Google Scholar]
- Hoekstra, K.A.; Paulton, R.J.L. Clinical prevalence and antimicrobial susceptibility of Staphylococcus aureus and Staph. intermedius in dogs. J. Appl. Microbiol. 2002, 93, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Waki, M.F.; Kogika, M.M.; Wirthl, V.A.B.F.; Oyafuso, M.K.; Prosser, C.S.; Monteiro, P.R.; Coelho, B.P.; Simões, D.M.N.; Kanayama, K.K. Association of urinary tract infection with urolithiasis in dogs. Clín. Vet. 2009, 14, 130–131. [Google Scholar]
- Marques, C.; Belas, A.; Franco, A.; Aboim, C.; Gama, L.T.; Pomba, C. Increase in antimicrobial resistance and emergence of major international high-risk clonal lineages in dogs and cats with urinary tract infection: 16 year retrospective study. J. Antimicrob. Chemother. 2018, 73, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.; Gama, L.T.; Belas, A.; Bergström, K.; Beurlet, S.; Briend-Marchal, A.; Broens, E.M.; Costa, M.; Criel, D.; Damborg, P.; et al. European multicenter study on antimicrobial resistance in bacteria isolated from companion animal urinary tract infections. BMC Vet. Res. 2016, 12, 213. [Google Scholar] [CrossRef] [Green Version]
- Rubin, J.E.; Gaunt, M.C. Urinary tract infection caused by methicillin-resistant Staphylococcus pseudintermedius in a dog. Can. Vet. J. 2011, 52, 162. [Google Scholar]
- Hutchins, R.G.; Vaden, S.L.; Jacob, M.E.; Harris, T.L.; Bowles, K.D.; Wood, M.W.; Bailey, C.S. Vaginal microbiota of spayed dogs with or without recurrent urinary tract infections. J. Vet. Intern. Med. 2014, 28, 300–304. [Google Scholar] [CrossRef]
- Moyaert, H.; de Jong, A.; Simjee, S.; Rose, M.; Youala, M.; El Garch, F.; Vila, T.; Klein, U.; Rzewuska, M.; Morrissey, I. Survey of antimicrobial susceptibility of bacterial pathogens isolated from dogs and cats with respiratory tract infections in Europe: ComPath results. J. Appl. Microbiol. 2019, 127, 29–46. [Google Scholar] [CrossRef]
- Kalhoro, D.H.; Gao, S.; Kalhoro, M.S. Staphylococcus pseudintermedius isolation from canine, bacterial colonization and clinical picture in balb/c mouse model. JAPS J. Anim. Plant Sci. 2017, 27, 422–429. [Google Scholar]
- Proulx, A.; Hume, D.Z.; Drobatz, K.J.; Reineke, E.L. In vitro bacterial isolate susceptibility to empirically selected antimicrobials in 111 dogs with bacterial pneumonia. J. Vet. Emerg. Crit. Care 2014, 24, 194–200. [Google Scholar] [CrossRef]
- Vientós-Plotts, A.I.; Ericsson, A.C.; Rindt, H.; Reinero, C.R. Respiratory Dysbiosis in Canine Bacterial Pneumonia: Standard Culture vs. Microbiome Sequencing. Front. Vet. Sci. 2019, 6, 354. [Google Scholar] [CrossRef]
- Morrissey, I.; Moyaert, H.; de Jong, A.; El Garch, F.; Klein, U.; Ludwig, C.; Thiry, J.; Youala, M. Antimicrobial susceptibility monitoring of bacterial pathogens isolated from respiratory tract infections in dogs and cats across Europe: ComPath results. Vet. Microbiol. 2016, 191, 44–51. [Google Scholar] [CrossRef]
- Kalhoro, D.H.; Gao, S.; Xie, X.; Liang, S.; Luo, S.; Zhao, Y.; Liu, Y. Canine influenza virus coinfection with Staphylococcus pseudintermedius enhances bacterial colonization, virus load and clinical presentation in mice. BMC Vet. Res. 2016, 12, 87. [Google Scholar] [CrossRef] [Green Version]
- Qekwana, D.N.; Naidoo, V.; Oguttu, J.W.; Odoi, A. Occurrence and Predictors of Bacterial Respiratory Tract Infections and Antimicrobial Resistance Among Isolates From Dogs Presented With Lower Respiratory Tract Infections at a Referral Veterinary Hospital in South Africa. Front. Vet. Sci. 2020, 7, 304. [Google Scholar] [CrossRef]
- Rheinwald, M.; Hartmann, K.; Hähner, M.; Wolf, G.; Straubinger, R.K.; Schulz, B. Antibiotic susceptibility of bacterial isolates from 502 dogs with respiratory signs. Vet. Rec. 2015, 176, 357. [Google Scholar] [CrossRef]
- Dear, J.D. Bacterial pneumonia in dogs and cats. Vet. Clin. N. Am. Small Anim. Pract. 2014, 44, 143–159. [Google Scholar]
- Lappin, M.R.; Blondeau, J.; Boothe, D.; Breitschwerdt, E.B.; Guardabassi, L.; Lloyd, D.H.; Papich, M.G.; Rankin, S.C.; Sykes, J.E.; Turnidge, J.; et al. Antimicrobial use Guidelines for Treatment of Respiratory Tract Disease in Dogs and Cats: Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases. J. Vet. Intern. Med. 2017, 31, 279–294. [Google Scholar] [CrossRef]
- Rota, A.; Milani, C.; Drigo, I.; Drigo, M.; Corrò, M. Isolation of methicillin-resistant Staphylococcus pseudintermedius from breeding dogs. Theriogenology 2011, 75, 115–121. [Google Scholar] [CrossRef]
- Zakošek Pipan, M.; Kajdič, L.; Kalin, A.; Plavec, T.; Zdovc, I. Do newborn puppies have their own microbiota at birth? Influence of type of birth on newborn puppy microbiota. Theriogenology 2020, 152, 18–28. [Google Scholar] [CrossRef]
- Saijonmaa-Koulumies, L.E.; Lloyd, D.H. Colonization of neonatal puppies by Staphylococcus intermedius. Vet. Dermatol. 2002, 13, 123–130. [Google Scholar] [CrossRef]
- Rota, A.; Corrò, M.; Drigo, I.; Bortolami, A.; Börjesson, S. Isolation of coagulase-positive Staphylococci from bitches’ colostrum and milk and genetic typing of methicillin-resistant Staphylococcus pseudintermedius strains. BMC Vet. Res. 2015, 11, 160. [Google Scholar] [CrossRef] [Green Version]
- Maluping, R.P.; Paul, N.C.; Moodley, A. Antimicrobial susceptibility of methicillin-resistant Staphylococcus pseudintermedius isolated from veterinary clinical cases in the UK. Br. J. Biomed. Sci. 2014, 71, 55–57. [Google Scholar] [CrossRef]
- Vasiu, I.; Dąbrowski, R.; Martinez-Subiela, S.; Ceron, J.J.; Wdowiak, A.; Pop, R.A.; Brudaşcă, F.G.; Pastor, J.; Tvarijonaviciute, A. Milk C-reactive protein in canine mastitis. Vet. Immunol. Immunopathol. 2017, 186, 41–44. [Google Scholar] [CrossRef]
- Ververidis, H.N.; Mavrogianni, V.S.; Fragkou, I.A.; Orfanou, D.C.; Gougoulis, D.A.; Tzivara, A.; Gouletsou, P.G.; Athanasiou, L.; Boscos, C.M.; Fthenakis, G.C. Experimental Staphylococcal mastitis in bitches: Clinical, bacteriological, cytological, haematological and pathological features. Vet. Microbiol. 2007, 124, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Ros, L.; Holst, B.S.; Hagman, R. A retrospective study of bitches with pyometra, medically treated with aglepristone. Theriogenology 2014, 82, 1281–1286. [Google Scholar] [CrossRef] [Green Version]
- Hagman, R. Pyometra in Small Animals. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 639–661. [Google Scholar]
- Robaj, A.; Sylejmani, I.; Hamidi, A. Occurrence and antimicrobial susceptibility of bacterial agents of canine pyometra. Indian J. Anim. Res. 2018, 52, 394–400. [Google Scholar]
- Pilla, R.; Bonura, C.; Malvisi, M.; Snel, G.G.M.; Piccinini, R. Methicillin-resistant Staphylococcus pseudintermedius as causative agent of dairy cow mastitis. Vet. Rec. 2013, 173, 19. [Google Scholar] [CrossRef]
- Paul, N.C.; Damborg, P.; Guardabassi, L. Dam-to-offspring transmission and persistence of Staphylococcus pseudintermedius clones within dog families. Vet. Dermatol. 2014, 25, 3-e2. [Google Scholar] [CrossRef]
- Saijonmaa-Koulumies, L.E.M.; Myllys, V.; Lloyd, D.H. Diversity and stability of the Staphylococcus intermedius flora in three bitches and their puppies. Epidemiol. Infect. 2003, 131, 931–937. [Google Scholar] [CrossRef]
- Latronico, F.; Moodley, A.; D’Abramo, M.; Greco, M.F.; Corrente, M.; Guardabassi, L. Outbreak of methicillin-resistant Staphylococcus pseudintermedius in a litter of puppies: Evidence of vertical perinatal and horizontal transmission. In Proceedings of the ASM Conference on Methicillin-Resistant Staphylococci in Animals, London, UK, 22–25 September 2009; Volume 76. [Google Scholar]
- Milani, C.; Corrò, M.; Drigo, M.; Rota, A. Antimicrobial resistance in bacteria from breeding dogs housed in kennels with differing neonatal mortality and use of antibiotics. Theriogenology 2012, 78, 1321–1328. [Google Scholar] [CrossRef]
- Meloni, T.; Martino, P.A.; Grieco, V.; Pisu, M.C.; Banco, B.; Rota, A.; Veronesi, M.C. A survey on bacterial involvement in neonatal mortality in dogs. Vet. Ital. 2014, 50, 293–299. [Google Scholar]
- Pipan, M.Z.; Švara, T.; Zdovc, I.; Papić, B.; Avberšek, J.; Kušar, D.; Mrkun, J. Staphylococcus pseudintermedius septicemia in puppies after elective cesarean section: Confirmed transmission via dam’s milk. BMC Vet. Res. 2019, 15, 41. [Google Scholar]
- Ogbu, K.I.; Ochai, S.O.; Danladi, M.M.A.; Abdullateef, M.H.; Agwu, E.O.; Gyengdeng, J.G. A review of Neonatal mortality in Dogs. Int. J. Life Sci. 2016, 4, 451–460. [Google Scholar]
- Hillier, A.; Lloyd, D.H.; Scott Weese, J.; Blondeau, J.M.; Boothe, D.; Breitschwerdt, E.; Guardabassi, L.; Papich, M.G.; Rankin, S.; Turnidge, J.D.; et al. Guidelines for the diagnosis and antimicrobial therapy of canine superficial bacterial folliculitis (Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases). Vet. Dermatol. 2014, 25, 163-e43. [Google Scholar] [CrossRef]
- Summers, J.F.; Hendricks, A.; Brodbelt, D.C. Prescribing practices of primary-care veterinary practitioners in dogs diagnosed with bacterial pyoderma. BMC Vet. Res. 2014, 10, 240. [Google Scholar] [CrossRef] [Green Version]
- Ungemach, F.R.; Müller-Bahrdt, D.; Abraham, G. Guidelines for prudent use of antimicrobials and their implications on antibiotic usage in veterinary medicine. Int. J. Med. Microbiol. 2006, 296 (Suppl. 41), 33–38. [Google Scholar] [CrossRef]
- Stegemann, M.R.; Passmore, C.A.; Sherington, J.; Lindeman, C.J.; Papp, G.; Weigel, D.J.; Skogerboe, T.L. Antimicrobial activity and spectrum of cefovecin, a new extended-spectrum cephalosporin, against pathogens collected from dogs and cats in Europe and North America. Antimicrob. Agents Chemother. 2006, 50, 2286–2292. [Google Scholar] [CrossRef] [Green Version]
- Stegemann, M.R.; Coati, N.; Passmore, C.A.; Sherington, J. Clinical efficacy and safety of cefovecin in the treatment of canine pyoderma and wound infections. J. Small Anim. Pract. 2007, 48, 378–386. [Google Scholar] [CrossRef]
- Maaland, M.; Guardabassi, L. In vitro antimicrobial activity of nitrofurantoin against Escherichia coli and Staphylococcus pseudintermedius isolated from dogs and cats. Vet. Microbiol. 2011, 151, 396–399. [Google Scholar] [CrossRef] [Green Version]
- Şentürk, S.; Özel, E.; Şen, A. Clinical efficacy of rifampicin for treatment of canine pyoderma. Acta Vet. Brno 2005, 74, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Blondeau, J.M.; Fitch, S.D. In vitro killing of canine strains of Staphylococcus pseudintermedius and Escherichia coli by cefazolin, cefovecin, doxycycline and pradofloxacin over a range of bacterial densities. Vet. Dermatol. 2020, 31, 187. [Google Scholar] [CrossRef]
- Morris, D.O.; Loeffler, A.; Davis, M.F.; Guardabassi, L.; Weese, J.S. Recommendations for approaches to meticillin-resistant Staphylococcal infections of small animals: Diagnosis, therapeutic considerations and preventative measures. Clinical Consensus Guidelines of the World Association for Veterinary Dermatology. Vet. Dermatol. 2017, 28, 304-e69. [Google Scholar] [CrossRef]
- Mueller, R.S.; Bergvall, K.; Bensignor, E.; Bond, R. A review of topical therapy for skin infections with bacteria and yeast. Vet. Dermatol. 2012, 23, 330–341.e62. [Google Scholar] [CrossRef]
- Nuttall, T. Topical therapy in canine atopic dermatitis: New products. Companion Anim. 2020, 25, 76–82. [Google Scholar] [CrossRef]
- Fadok, V.A.; Irwin, K. Sodium Hypochlorite/Salicylic Acid Shampoo for Treatment of Canine Staphylococcal Pyoderma. J. Am. Anim. Hosp. Assoc. 2019, 55, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Loeffler, A.; Cobb, M.A.; Bond, R. Comparison of a chlorhexidine and a benzoyl peroxide shampoo as sole treatment in canine superficial pyoderma. Vet. Rec. 2011, 169, 249. [Google Scholar] [CrossRef] [PubMed]
- Murayama, N.; Nagata, M.; Terada, Y.; Shibata, S.; Fukata, T. Efficacy of a surgical scrub including 2% chlorhexidine acetate for canine superficial pyoderma. Vet. Dermatol. 2010, 21, 586–592. [Google Scholar] [CrossRef] [PubMed]
- Borio, S.; Colombo, S.; La Rosa, G.; De Lucia, M.; Damborg, P.; Guardabassi, L. Effectiveness of a combined (4% chlorhexidine digluconate shampoo and solution) protocol in MRS and non-MRS canine superficial pyoderma: A randomized, blinded, antibiotic-controlled study. Vet. Dermatol. 2015, 26, 339–344.e72. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.J.; Woodward, M.; Hoppers, S.M.; Liu, C.-C.; Pucheu-Haston, C.M.; Mitchell, M.S. Residual antibacterial activity of canine hair treated with five mousse products against Staphylococcus pseudintermedius in vitro. Vet. Dermatol. 2019, 30, 183-e57. [Google Scholar] [CrossRef]
- Kloos, I.; Straubinger, R.K.; Werckenthin, C.; Mueller, R.S. Residual antibacterial activity of dog hairs after therapy with antimicrobial shampoos. Vet. Dermatol. 2013, 24, 250-e54. [Google Scholar] [CrossRef]
- Jorge, S.; Dellagostin, O.A. The development of veterinary vaccines: A review of traditional methods and modern biotechnology approaches. Biotechnol. Res. Innov. 2017, 1, 6–13. [Google Scholar] [CrossRef]
- Chambers, E.D.; Severin, G.A. Staphylococcal bacterin for treatment of chronic Staphylococcal blepharitis in the dog. J. Am. Vet. Med. Assoc. 1984, 185, 422–425. [Google Scholar]
- DeBoer, D.J.; Moriello, K.A.; Thomas, C.B.; Schultz, K.T. Evaluation of a commercial Staphylococcal bacterin for management of idiopathic recurrent superficial pyoderma in dogs. Am. J. Vet. Res. 1990, 51, 636–639. [Google Scholar]
- Curtis, C.F.; Lamport, A.I.; Lloyd, D.H. Masked, controlled study to investigate the efficacy of a Staphylococcus intermedius autogenous bacterin for the control of canine idiopathic recurrent superficial pyoderma. Vet. Dermatol. 2006, 17, 163–168. [Google Scholar] [CrossRef]
- Borku, M.K.; Ozkanlar, Y.; Hanedan, B.; Duru, S.Y. Efficacy of Staphylococcal bacterin for treatment of canine re-current pyoderma: An open clinical trial. Revue de Médecine Vétérinaire 2007, 158, 234. [Google Scholar]
- Staphage Lysate (SPL). Available online: https://www.drugs.com/vet/staphage-lysate-spl.html (accessed on 4 January 2021).
- Wilson, A.; Allers, N.; Lloyd, D.H.; Bond, R.; Loeffler, A. Reduced antimicrobial prescribing during autogenous staphylococcal bacterin therapy: A retrospective study in dogs with pyoderma. Vet. Rec. 2019, 184, 739. [Google Scholar] [CrossRef]
- Thammavongsa, V.; Kim, H.K.; Missiakas, D.; Schneewind, O. Staphylococcal manipulation of host immune responses. Nat. Rev. Microbiol. 2015, 13, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Grandolfo, E. Looking through Staphylococcus pseudintermedius infections: Could SpA be considered a possible vaccine target? Virulence 2018, 9, 703–706. [Google Scholar] [CrossRef]
- Abouelkhair, M.A.; Bemis, D.A.; Kania, S.A. Characterization of recombinant wild-type and nontoxigenic protein A from Staphylococcus pseudintermedius. Virulence 2018, 9, 1050–1061. [Google Scholar] [CrossRef] [Green Version]
- Votintseva, A.A.; Fung, R.; Miller, R.R.; Knox, K.; Godwin, H.; Wyllie, D.H.; Bowden, R.; Crook, D.W.; Walker, A.S. Prevalence of Staphylococcus aureus protein A (spa) mutants in the community and hospitals in Oxfordshire. BMC Microbiol. 2014, 14, 63. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Cheng, A.G.; Kim, H.-Y.; Missiakas, D.M.; Schneewind, O. Nontoxigenic protein A vaccine for methicillin-resistant Staphylococcus aureus infections in mice. J. Exp. Med. 2010, 207, 1863–1870. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Falugi, F.; Thomer, L.; Missiakas, D.M.; Schneewind, O. Protein A suppresses immune responses during Staphylococcus aureus bloodstream infection in guinea pigs. MBio 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Balachandran, M.; Bemis, D.A.; Kania, S.A. Expression and function of protein A in Staphylococcus pseudintermedius. Virulence 2018, 9, 390–401. [Google Scholar] [CrossRef] [Green Version]
- Schiebel, J.; Chang, A.; Lu, H.; Baxter, M.V.; Tonge, P.J.; Kisker, C. Staphylococcus aureus FabI: Inhibition, substrate recognition, and potential implications for in vivo essentiality. Structure 2012, 20, 802–813. [Google Scholar] [CrossRef] [Green Version]
- El-Shibiny, A.; El-Sahhar, S. Bacteriophages: The possible solution to treat infections caused by pathogenic bacteria. Can. J. Microbiol. 2017, 63, 865–879. [Google Scholar] [CrossRef] [Green Version]
- Twort, F.W. An investigation on the nature of ultra-microscopic viruses. Lancet 1915, 186, 1241–1243. [Google Scholar] [CrossRef] [Green Version]
- D’Herelle, M.F. On an invisible microbe antagonistic to dysentery bacilli. Note by M. F. d’Herelle, presented by M. Roux. Comptes Rendus Academie des Sciences 1917; 165:373–5. Bacteriophage 2011, 1, 3–5. [Google Scholar]
- Gordillo Altamirano, F.L.; Barr, J.J. Phage Therapy in the Postantibiotic Era. Clin. Microbiol. Rev. 2019, 32. [Google Scholar] [CrossRef] [Green Version]
- Rakhuba, D.V.; Kolomiets, E.I.; Dey, E.S.; Novik, G.I. Bacteriophage receptors, mechanisms of phage adsorption and penetration into host cell. Pol. J. Microbiol. 2010, 59, 145–155. [Google Scholar] [CrossRef]
- Cieplak, T.; Soffer, N.; Sulakvelidze, A.; Nielsen, D.S. A bacteriophage cocktail targeting Escherichia coli reduces E. coli in simulated gut conditions, while preserving a non-targeted representative commensal normal microbiota. Gut Microbes 2018, 9, 391–399. [Google Scholar]
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [Green Version]
- Ryan, E.M.; Gorman, S.P.; Donnelly, R.F.; Gilmore, B.F. Recent advances in bacteriophage therapy: How delivery routes, formulation, concentration and timing influence the success of phage therapy. J. Pharm. Pharmacol. 2011, 63, 1253–1264. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, C.; Harper, D.; Burch, D.; Anggård, E.; Soothill, J. Topical treatment of Pseudomonas aeruginosa otitis of dogs with a bacteriophage mixture: A before/after clinical trial. Vet. Microbiol. 2010, 146, 309–313. [Google Scholar] [CrossRef] [PubMed]
- McCallin, S.; Sacher, J.C.; Zheng, J.; Chan, B.K. Current State of Compassionate Phage Therapy. Viruses 2019, 11, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeman, M.; Bárdy, P.; Vrbovská, V.; Roudnický, P.; Zdráhal, Z.; Růžičková, V.; Doškař, J.; Pantůček, R. New Genus Fibralongavirus in Siphoviridae Phages of Staphylococcus pseudintermedius. Viruses 2019, 11, 1143. [Google Scholar] [CrossRef] [Green Version]
- Moodley, A.; Kot, W.; Nälgård, S.; Jakociune, D.; Neve, H.; Hansen, L.H.; Guardabassi, L.; Vogensen, F.K. Isolation and characterization of bacteriophages active against methicillin-resistant Staphylococcus pseudintermedius. Res. Vet. Sci. 2019, 122, 81–85. [Google Scholar] [CrossRef]
- Azam, A.H.; Kadoi, K.; Miyanaga, K.; Usui, M.; Tamura, Y.; Cui, L.; Tanji, Y. Analysis host-recognition mechanism of staphylococcal kayvirus ɸSA039 reveals a novel strategy that protects Staphylococcus aureus against infection by Staphylococcus pseudintermedius Siphoviridae phages. Appl. Microbiol. Biotechnol. 2019, 103, 6809–6823. [Google Scholar] [CrossRef]
- Breteau, M. Study of Staphylococcus pseudintermedius Phages: Towards the Development of Phage Therapy. Ph.D. Thesis, University of Warwick, Coventry, UK, 2016. [Google Scholar]
- Nakamura, T.; Kitana, J.; Fujiki, J.; Takase, M.; Iyori, K.; Simoike, K.; Iwano, H. Lytic Activity of Polyvalent Staphylococcal Bacteriophage PhiSA012 and Its Endolysin Lys-PhiSA012 Against Antibiotic-Resistant Staphylococcal Clinical Isolates From Canine Skin Infection Sites. Front. Med. 2020, 7, 234. [Google Scholar] [CrossRef]
- Squires, R.A. Bacteriophage therapy for management of bacterial infections in veterinary practice: What was once old is new again. N. Z. Vet. J. 2018, 66, 229–235. [Google Scholar] [CrossRef]
- Schmelcher, M.; Donovan, D.M.; Loessner, M.J. Bacteriophage endolysins as novel antimicrobials. Future Microbiol. 2012, 7, 1147–1171. [Google Scholar] [CrossRef] [Green Version]
- Junjappa, R.P.; Desai, S.N.; Roy, P.; Narasimhaswamy, N.; Raj, J.R.M.; Durgaiah, M.; Vipra, A.; Bhat, U.R.; Satyanarayana, S.K.; Shankara, N.; et al. Efficacy of anti-staphylococcal protein P128 for the treatment of canine pyoderma: Potential applications. Vet. Res. Commun. 2013, 37, 217–228. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lynch, S.A.; Helbig, K.J. The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next? Vet. Sci. 2021, 8, 11. https://doi.org/10.3390/vetsci8010011
Lynch SA, Helbig KJ. The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next? Veterinary Sciences. 2021; 8(1):11. https://doi.org/10.3390/vetsci8010011
Chicago/Turabian StyleLynch, Stephanie A., and Karla J. Helbig. 2021. "The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next?" Veterinary Sciences 8, no. 1: 11. https://doi.org/10.3390/vetsci8010011
APA StyleLynch, S. A., & Helbig, K. J. (2021). The Complex Diseases of Staphylococcus pseudintermedius in Canines: Where to Next? Veterinary Sciences, 8(1), 11. https://doi.org/10.3390/vetsci8010011