
Protective and Therapeutic Effects of Orlistat on Metabolic Syndrome and Oxidative Stress in High-Fat Diet-Induced Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Rats: Role on Nrf2 Activation

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Treatment
- Normal control (N) group: Received normal pellet food and distilled water for 12 weeks;
- High-fat diet (HFD) group: Received HFD for 12 weeks;
- HFD + orlistat (HFD + O) group: Received HFD and orlistat (10 mg/kg of body weight/day) concomitantly for 12 weeks as a protective model;
- Obese + orlistat (OB + O) group: Received HFD for 6 weeks to induce obesity and received both HFD and orlistat (10 mg/kg of body weight/day) during the last 6 weeks as a therapeutic model.
2.2. Oral Glucose Tolerance Test (OGTT)
2.3. Measurements of Serum Glucose, Insulin and HOMA-IR
2.4. Determination of Lipid and Liver Function Profiles
2.5. Liver Lipid Content
2.6. Analysis of Liver Oxidative Stress Markers
2.7. Analysis of Liver Antioxidant Enzyme Activities
2.8. Analysis of Liver Glutathione and Total Antioxidant Capacity
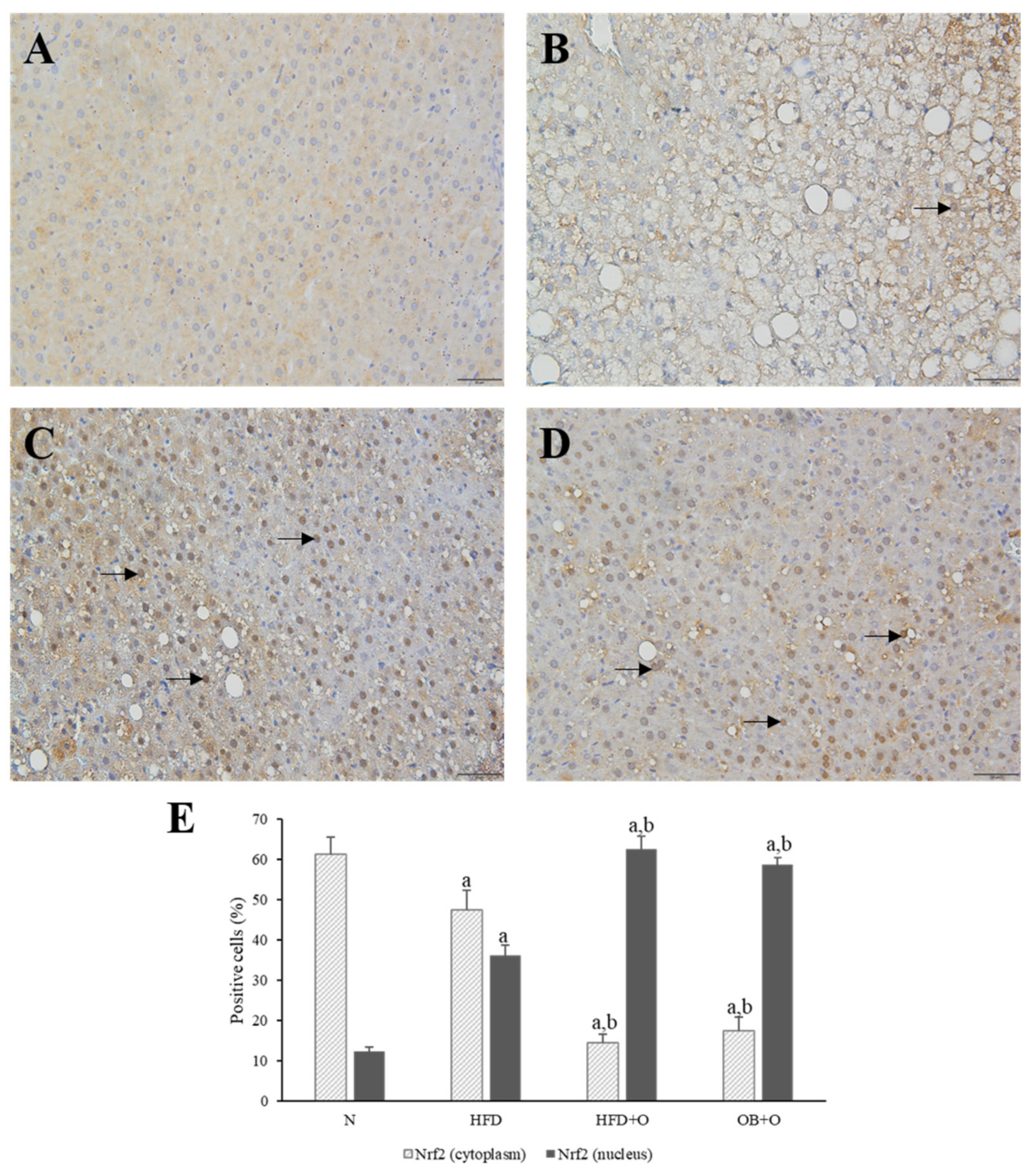
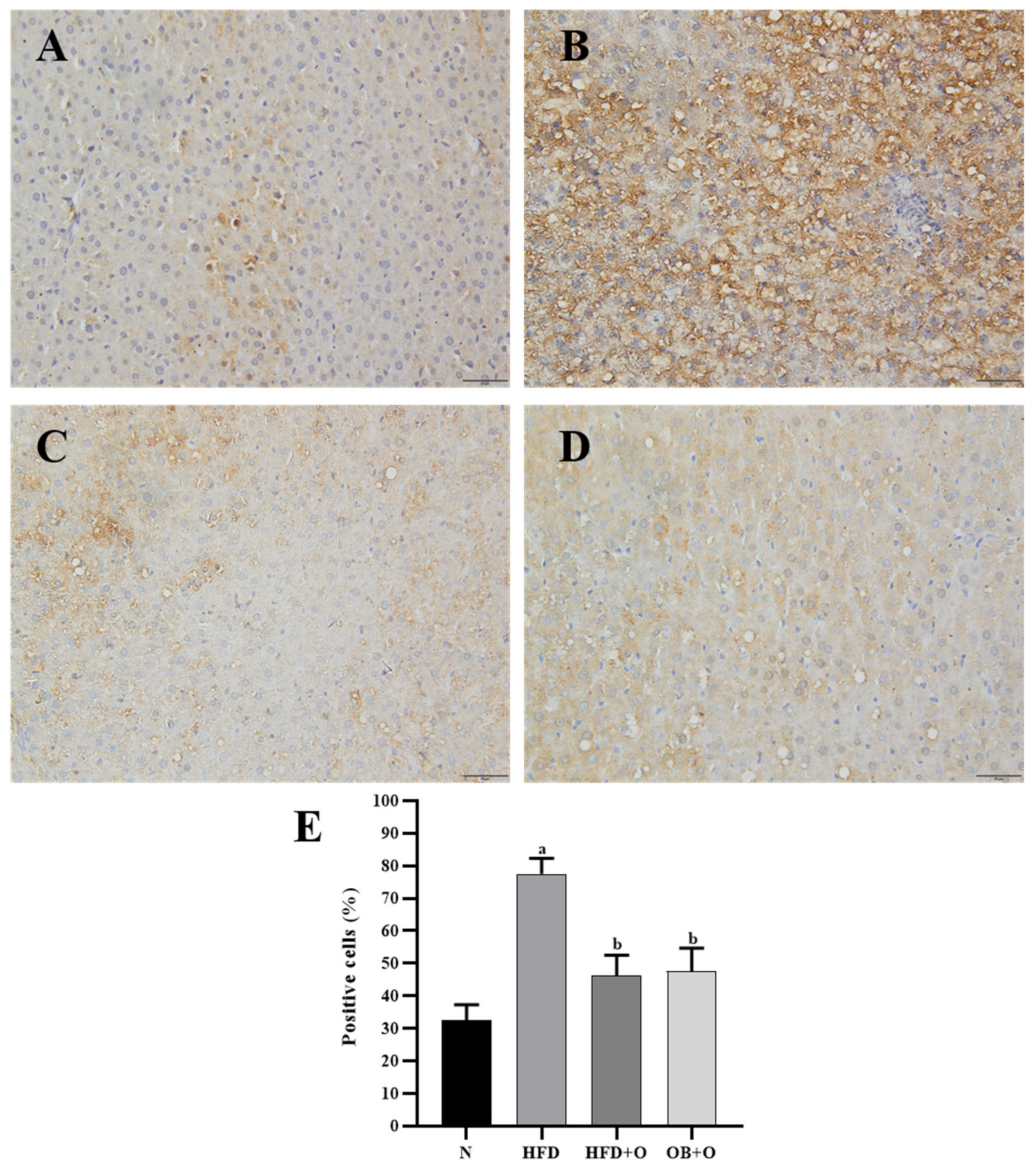
2.9. Immunohistochemical Detections of Nrf2 and Keap1 Expressions
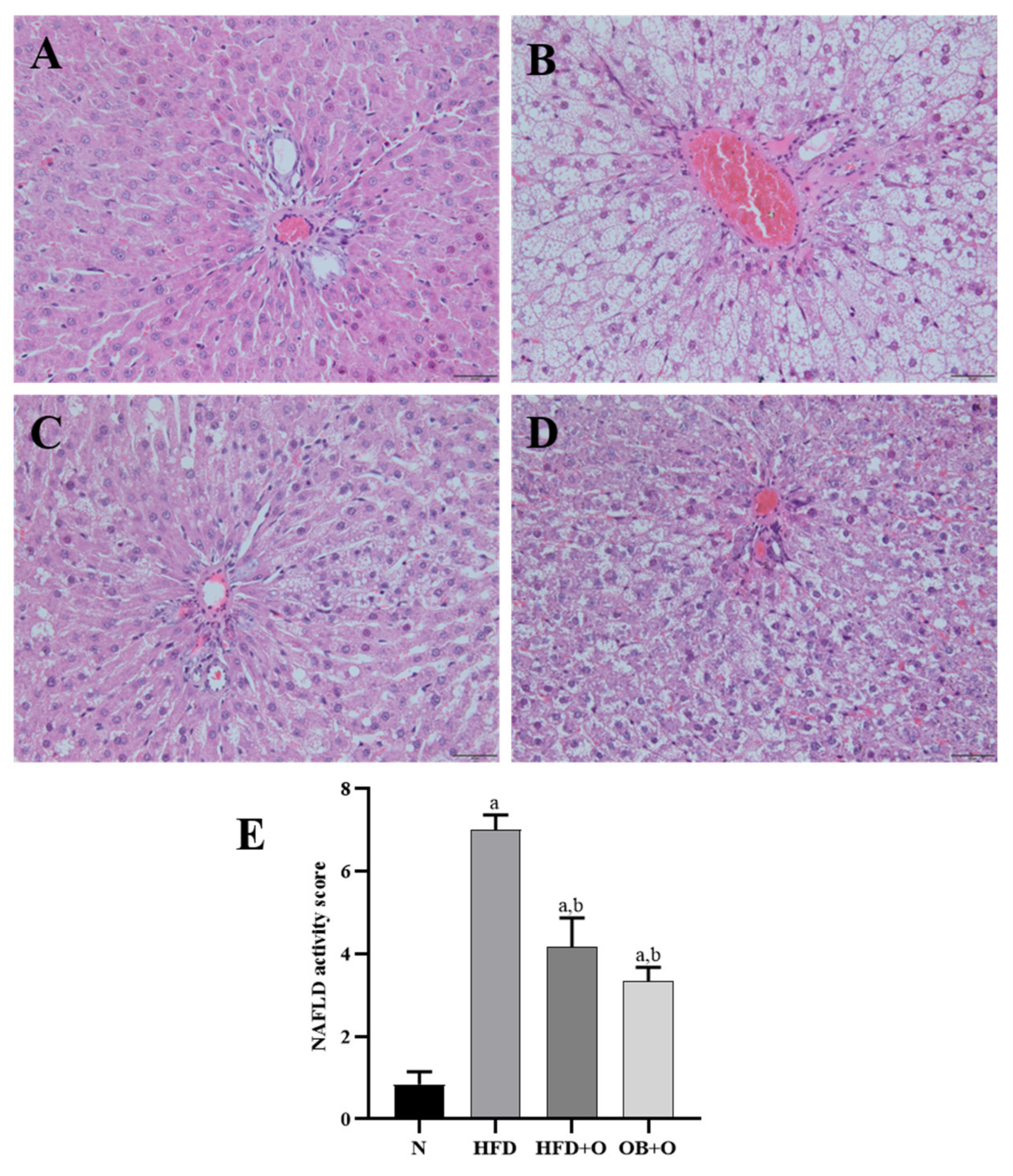
2.10. Histopathological Analysis
2.11. Statistical Analysis
3. Results
3.1. Lee Obesity Index, Body Weight Gain, Food Intake, and Calorie Intake
3.2. Effects of Orlistat on Liver and Adipose Tissue Weights
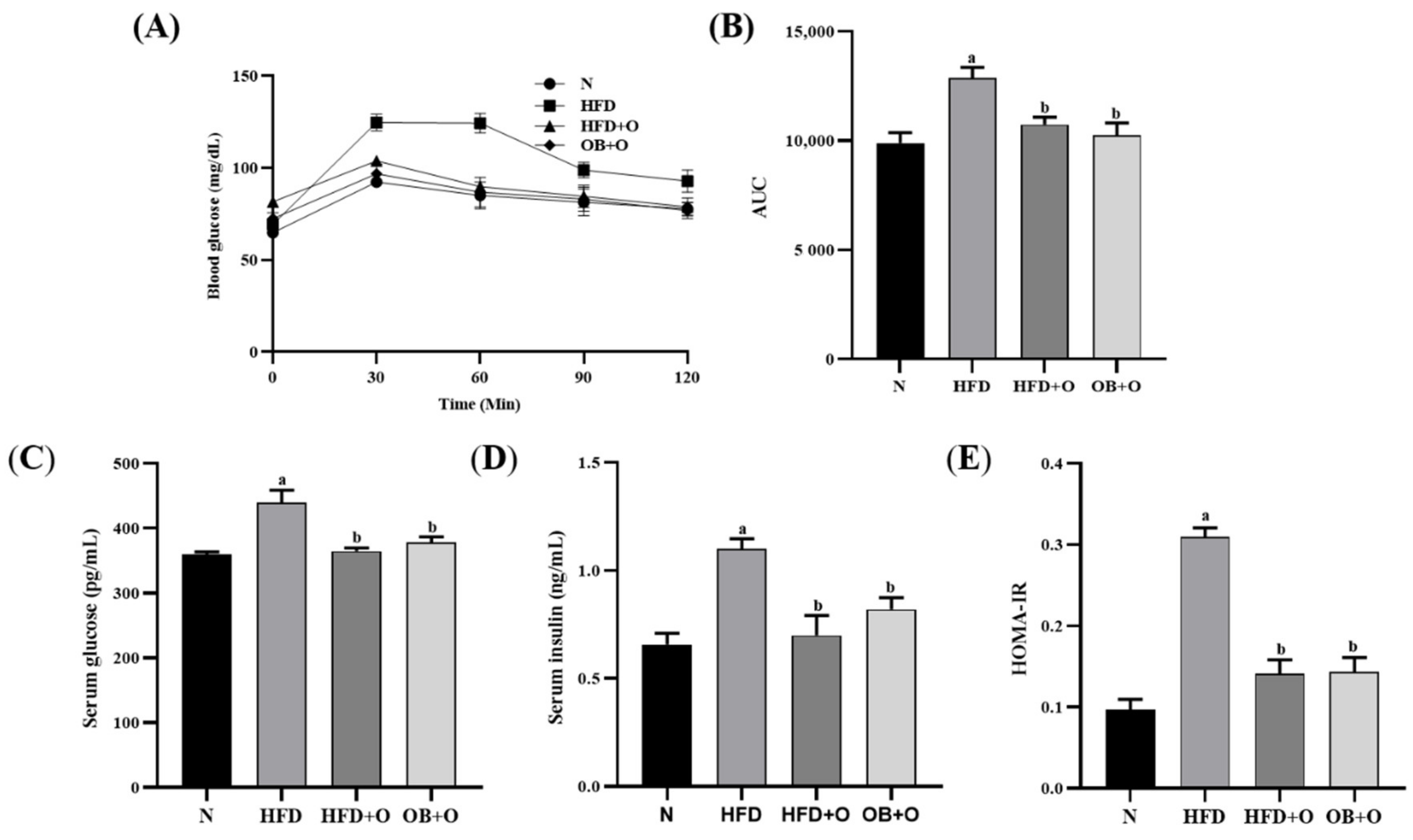
3.3. Effects of Orlistat on Oral Glucose Tolerance Test (OGTT)
3.4. Effects of Orlistat on Insulin Sensitivity
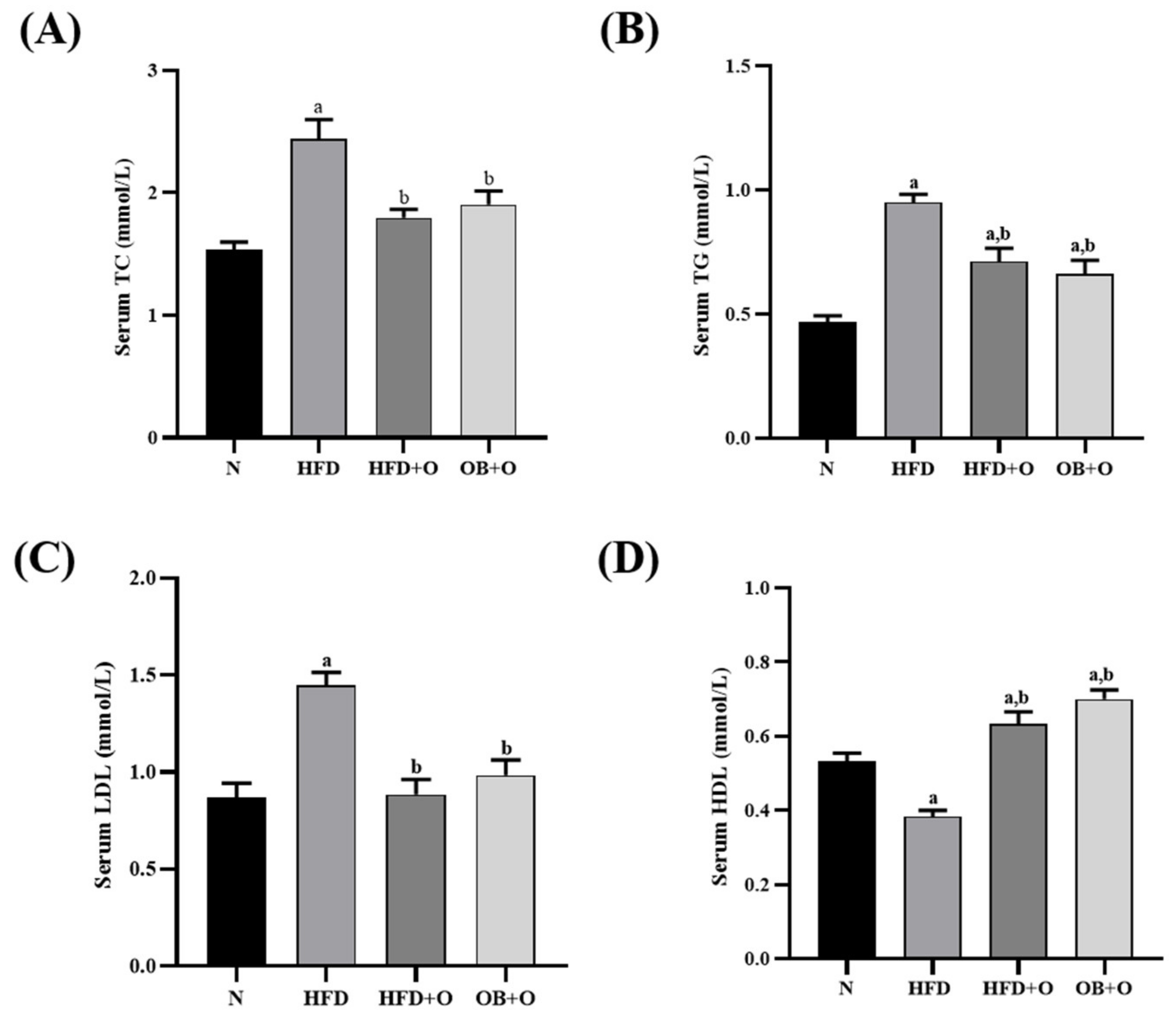
3.5. Effects of Orlistat on Lipid Profiles
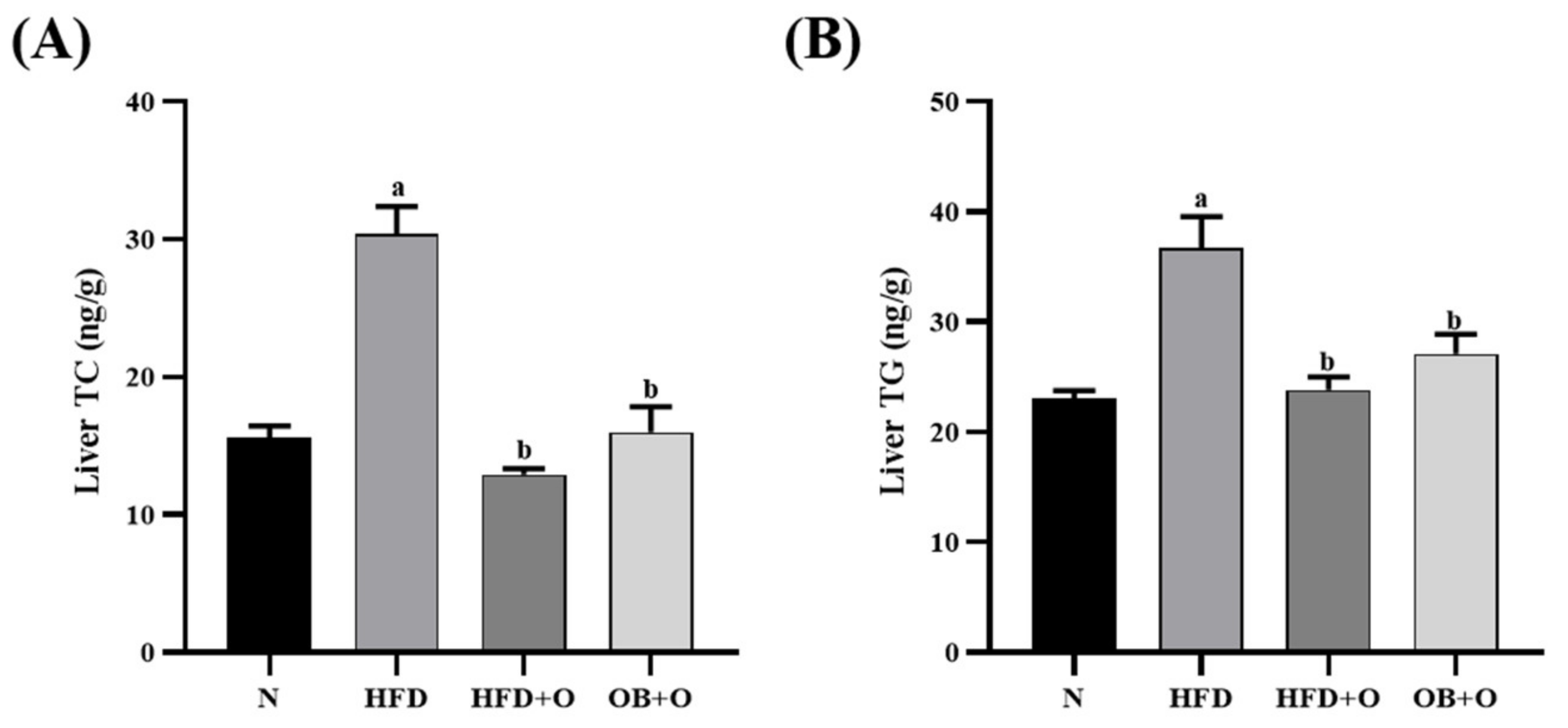
3.6. Effects of Orlistat on Liver Lipid Content
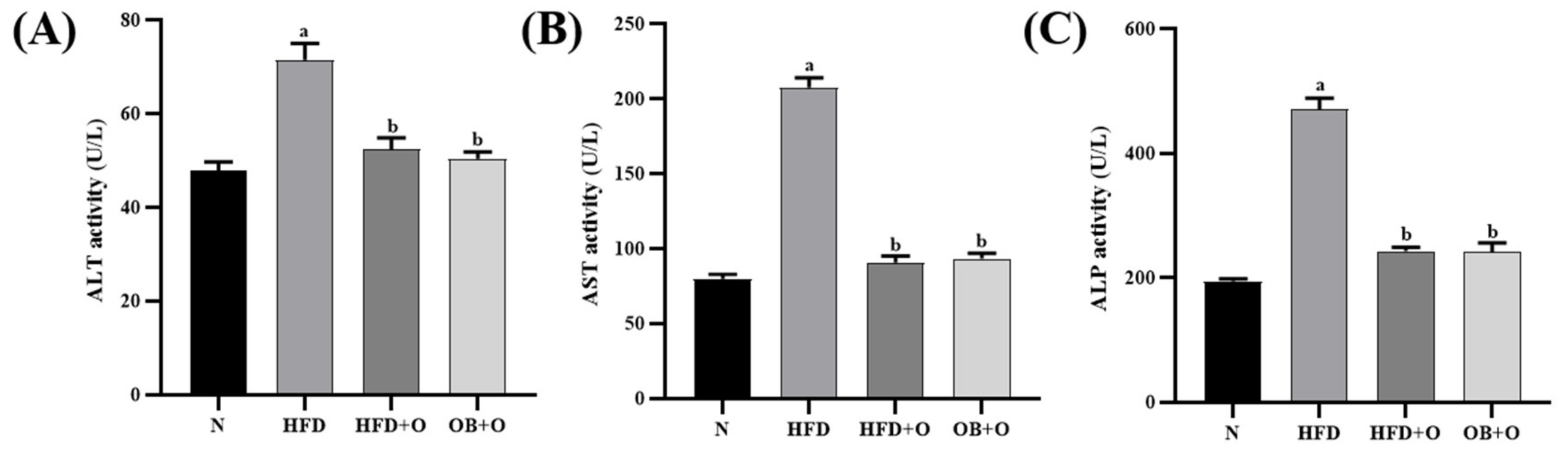
3.7. Effects of Orlistat on Liver Functions
3.8. Effects of Orlistat on Liver Oxidative Stress Markers
3.9. Effects of Orlistat on Liver Antioxidant Enzyme Activities
3.10. Effects of Orlistat on Glutathione and Total Antioxidant Capacity
3.11. Effects of Orlistat on Immunohistochemical Analysis of Nrf2 and Keap1 Expressions
3.12. Effects of Orlistat on MAFLD and Analysis of NAS
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rich, N.E.; Oji, S.; Mufti, A.R.; Browning, J.D.; Parikh, N.D.; Odewole, M.; Mayo, H.; Singal, A.G. Racial and ethnic disparities in non-alcoholic fatty liver disease prevalence, severity, and outcomes in the United States: A systematic review and meta-analysis. Clin. Gastroenterol. Hepatol. 2018, 16, 198–210.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schreuder, T.C.H.A.; Verwer, B.J.; van Nieuwkerk, C.M.J.; Mulder, C.J.J. Nonalcoholic fatty liver disease: An overview of current insights in pathogenesis, diagnosis and treatment. World J. Gastroenterol. 2008, 14, 2474–2486. [Google Scholar] [CrossRef]
- Adams, L.A.; Lymp, J.F.; Sauver, J.S.; Sanderson, S.O.; Lindor, K.D.; Feldstein, A.; Angulo, P. The natural history of nonalcoholic fatty liver disease: A population-based cohort study. Gastroenterology 2005, 129, 113–121. [Google Scholar] [CrossRef]
- Matteoni, C.A.; Younossi, Z.M.; Gramlich, T.; Boparai, N.; Liu, Y.C.; McCullough, A.J. Nonalcoholic fatty liver disease: A spectrum of clinical and pathological severity. Gastroenterology 1999, 116, 1413–1419. [Google Scholar] [CrossRef]
- Lasker, S.; Rahman, M.M.; Parvez, F.; Zamila, M.; Miah, P.; Nahar, K.; Kabir, F.; Sharmin, S.B.; Subhan, N.; Ahsan, G.U.; et al. High-fat diet-induced metabolic syndrome and oxidative stress in obese rats are ameliorated by yogurt supplementation. Sci. Rep. 2019, 9, 1–15. [Google Scholar] [CrossRef]
- Yang, M.; Jiang, Z.H.; Li, C.G.; Zhu, Y.J.; Li, Z.; Tang, Y.Z.; Ni, C.L. Apigenin prevents metabolic syndrome in high-fructose diet-fed mice by Keap1-Nrf2 pathway. Biomed. Pharmacother. 2018, 105, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Golabi, P.; Paik, J.M.; Arshad, T.; Younossi, Y.; Mishra, A.; Younossi, Z.M. Mortality of NAFLD according to the body composition and presence of metabolic abnormalities. Hepatol. Commun. 2020, 4, 1136–1148. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Younossi, Z.M.; Golabi, P.; de Avila, L.; Paik, J.M.; Srishord, M.; Fukui, N.; Qiu, Y.; Burns, L.; Afendy, A.; Nader, F. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: A systematic review and meta-analysis. J. Hepatol. 2019, 71, 793–801. [Google Scholar] [CrossRef]
- Ipsen, D.H.; Tveden-Nyborg, P.; Lykkesfeldt, J. Normal weight dyslipidemia: Is it all about the liver? Obesity 2016, 24, 556–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza, M.R.A.; Diniz, M.F.F.M.; Medeiros-Filho, J.E.M.; Araújo, M.S.T. Metabolic syndrome and risk factors for non-alcoholic fatty liver disease. Arq. Gastroenterol. 2012, 49, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.L.; Lin, J.S.; Li, Y.H.; Liu, M.; Deng, Y.Y.; Wang, C.Y.; Chen, Y.M. Adherence to the dietary approaches to stop hypertension (DASH) diet is associated with lower presence of non-alcoholic fatty liver disease in middle-aged and elderly adults. Public Health Nutr. 2020, 23, 674–682. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Zhao, G.J.; Chen, Z.; She, Z.G.; Cai, J.; Li, H. Nonalcoholic fatty liver disease: An emerging driver of hypertension. Hypertension 2020, 75, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Tanase, D.M.; Gosav, E.M.; Costea, C.F.; Ciocoiu, M.; Lacatusu, C.M.; Maranduca, M.A.; Ouatu, A.; Floria, M. The intricate relationship between type 2 diabetes mellitus (T2DM), insulin resistance (IR), and nonalcoholic fatty liver disease (NAFLD). J. Diabetes Res. 2020, 2020, 16. [Google Scholar] [CrossRef]
- Coccia, F.; Testa, M.; Guarisco, G.; Di Cristofano, C.; Silecchia, G.; Leonetti, F.; Gastaldelli, A.; Capoccia, D. Insulin resistance, but not insulin response, during oral glucose tolerance test (OGTT) is associated to worse histological outcome in obese NAFLD. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Eslam, M.; Newsome, P.N.; Sarin, S.K.; Anstee, Q.M.; Targher, G.; Romer-Gomez, M.; Zelber-Sagi, S.; Wong, V.W.-S.; Dufor, J.F.; Schattenberg, J.M.; et al. A new definition for metabolic dysfunction-associated fatty liver disease: An international consensus statement. J. Hepatol. 2020, 73, 202–209. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J. MAFLD: A consensus-driven proposed nomenclature for metabolic associated fatty liver disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Marchesini, G.; Brizi, M.; Bianchi, G.; Tomassetti, S.; Bugianesi, E.; Lenzi, M.; McCullough, A.J.; Natale, S.; Forlani, G.; Melchionda, N. Nonalcoholic fatty liver disease: A feature of the metabolic syndrome. Diabetes 2001, 50, 1844–1850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.M.; Alam, M.N.; Ulla, A.; Sumi, F.A.; Subhan, N.; Khan, T.; Sikder, B.; Hossain, H.; Reza, H.M.; Alam, M.A. Cardamom powder supplementation prevents obesity, improves glucose intolerance, inflammation and oxidative stress in liver of high carbohydrate high fat diet induced obese rats. Lipids Health. Dis. 2017, 16, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poznyak, A.V.; Grechko, A.V.; Orekhova, V.A.; Chegodaev, Y.S.; Wu, W.-K.; Orekhov, A.N. Oxidative stress and antioxidants in atherosclerosis development and treatment. Biology 2020, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Chaudhary, P.; Pandey, A.; Azad, C.S.; Tia, N.; Singh, M.; Gambhir, I.S. Association of oxidative stress and endothelial dysfunction in hypertension. Anal. Biochem. 2020, 590, 113535. [Google Scholar] [CrossRef]
- Mistry, K.N.; Dabhi, B.K.; Joshi, B.B. Evaluation of oxidative stress biomarkers and inflammation in pathogenesis of diabetes and diabetic nephropathy. Indian J. Biochem. Biophys. 2020, 57, 45–50. [Google Scholar]
- Chambel, S.S.; Santos-Gonçalves, A.; Duarte, T.L. The dual role of Nrf2 in nonalcoholic fatty liver disease: Regulation of antioxidant defenses and hepatic lipid metabolism. Biomed. Res. Int. 2015, 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell survival responses to environmental stresses via the Keap1-Nrf2-ARE pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cui, Y.; Li, C.; Zhang, Y.; Xu, S.; Li, X.; Li, H.; Zhang, X. Nrf2 deletion causes “benign” simple steatosis to develop into nonalcoholic steatohepatitis in mice fed a high-fat diet. Lipids Health Dis. 2013, 12, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugimoto, H.; Okada, K.; Shoda, J.; Warabi, E.; Ishige, K.; Ueda, T.; Taguchi, K.; Yanagawa, T.; Nakahara, A.; Hyodo, I.; et al. Deletion of nuclear factor-E2-related factor-2 leads to rapid onset and progression of nutritional steatohepatitis in mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G283–G294. [Google Scholar] [CrossRef]
- Hosseini, H.; Teimouri, M.; Shabani, M.; Koushki, M.; Khorzoughi, R.B.; Namvarjah, F.; Izadi, P.; Meshkani, R. Resveratrol alleviates non-alcoholic fatty liver disease through epigenetic modification of the Nrf2 signaling pathway. Int. J. Biochem. Cell Biol. 2020, 119, 105667. [Google Scholar] [CrossRef]
- Abbate, J.M.; Macrì, F.; Capparucci, F.; Iaria, C.; Briguglio, G.; Cicero, L.; Salvo, A.; Arfuso, F.; Ieni, A.; Piccione, G.; et al. Administration of protein hydrolysates from anchovy (Engraulis encrasicolus) waste for twelve weeks decreases metabolic dysfunction-associated fatty liver disease severity in ApoE−/− mice. Animals 2020, 10, 2303. [Google Scholar] [CrossRef]
- Abbate, J.M.; Macrì, F.; Arfuso, F.; Iaria, C.; Capparucci, F.; Anfuso, C.; Ieni, A.; Cicero, L.; Briguglio, G.; Lanteri, G. Anti-atherogenic effect of 10% supplementation of anchovy (Engraulis encrasicolus) waste protein hydrolysates in ApoE-deficient mice. Nutrients 2021, 13, 2137. [Google Scholar] [CrossRef]
- Ye, J.; Wu, Y.; Li, F.; Wu, T.; Shao, C.; Lin, Y.; Wang, W.; Fenf, S.; Zhong, B. Effect of orlistat on liver fat content in patients with nonalcoholic fatty liver disease with obesity: Assessment using magnetic resonance imaging-derived proton density fat fraction. Ther. Adv. Gastroenterol. 2019, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Simental-Mendía, L.E.; Kovanen, P.T.; Pedone, C.; Simental-Mendía, M.; Cicero, A.F.G. Effects of orlistat on blood pressure: A systematic review and meta-analysis of 27 randomized controlled clinical trials. J. Am. Soc. Hypertens. 2018, 12, 80–96. [Google Scholar] [CrossRef]
- Khan, R.A.; Kapur, P.; Jain, A.; Farah, F.; Bhandari, U. Effect of orlistat on periostin, adiponectin, inflammatory markers and ultrasound grades of fatty liver in obese NAFLD patients. Ther. Clin. Risk Manag. 2017, 13, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Othman, Z.A.; Wan Ghazali, W.S.; Noordin, L.; Omar, N.; Yusof, N.A.; Mohamed, M. Protective effects of orlistat on lipid profile, cardiac oxidative stress biomarkers and histology in high-fat diet-induced obese. IIUM Med. J. Malaysia 2019, 18, 23–28. [Google Scholar] [CrossRef]
- Suleiman, J.B.; Nna, V.U.; Zakaria, Z.; Othman, Z.A.; Abu Bakar, A.B.; Mohamed, M. Obesity-induced testicular oxidative stress, inflammation and apoptosis: Protective and therapeutic effects of orlistat. Reprod. Toxicol. 2020, 95, 113–122. [Google Scholar] [CrossRef]
- Suleiman, J.B.; Nna, V.U.; Zakaria, Z.; Othman, Z.A.; Eleazu, C.O.; Abu Bakar, A.B.; Ahmad, A.; Usman, U.Z.; Rahman, F.W.A.; Mohamed, M. Protective effects of bee bread on testicular oxidative stress, NF-κB-mediated inflammation, apoptosis and lactate transport decline in obese male rats. Biomed. Pharmacother. 2020, 131, 110781. [Google Scholar] [CrossRef] [PubMed]
- Bellinger, L.L.; Bernardis, L.L. Effect of dorsomedial hypothalamic nuclei knife cuts on ingestive behavior. Am. J. Physiol. Regul. Interg. Comp. Physiol. 1999, 276, R1772–R1779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malafaia, A.B.; Nassif, P.A.N.; Ribas, C.A.P.M.; Ariede, B.L.; Sue, K.N.; Cruz, M.A. Obesity induction with high fat sucrose in rats. ABCD Arq. Bras. Cir. Dig. 2013, 26, 17–21. [Google Scholar] [CrossRef]
- Zaitone, S.A.; Essawy, S. Addition of a low dose of rimonabant to orlistat therapy decreases weight gain and reduces adiposity in dietary obese rats. Clin. Exp. Pharmacol. Physiol. 2012, 39, 551–559. [Google Scholar] [CrossRef]
- Othman, Z.A.; Ghazali, W.S.W.; Noordin, L.; Mohd Yusof, N.A.; Mohamed, M. Phenolic compounds and the anti-atherogenic effect of bee bread in high-fat diet-induced obese rats. Antioxidants 2020, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Roza, N.A.V.; Possignolo, L.F.; Palanch, A.C.; Gontijo, J.A.R. Effect of long-term high-fat diet intake on peripheral insulin sensibility, blood pressure, and renal function in female rats. Food Nutr. Res. 2016, 60, 28536. [Google Scholar] [CrossRef] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.K.; Cuzzocrea, S.; Brown, P.A.J.; Zacharowski, K.; Stewart, K.N.; Mota-Filipe, H.; Thiemermann, C. Tempol, a membrane-permeable radical scavenger, reduces oxidant stress-mediated renal dysfunction and injury in the rat. Kidney Int. 2000, 58, 658–673. [Google Scholar] [CrossRef] [Green Version]
- Evans, P.; Lyras, L.; Halliwell, B. Measurement of protein carbonyls in human brain tissue. Methods Enzymol. 1999, 300, 145–156. [Google Scholar] [CrossRef]
- Al Batran, R.; Al-Bayaty, F.; Al-Obaidi, M.M.J.; Abdualkader, A.M.; Hadi, H.A.; Ali, H.M.; Abdulla, M.A. In vivo antioxidant and antiulcer activity of Parkia speciosa ethanolic leaf extract against ethanol-induced gastric ulcer in rats. PLoS ONE 2013, 8, e64751. [Google Scholar] [CrossRef] [PubMed]
- Chandran, G.; Sirajudeen, K.N.S.; Yusoff, N.S.N.; Swamy, M.; Samarendra, M.S. Effect of the antihypertensive drug enalapril on oxidative stress markers and antioxidant enzymes in kidney of spontaneously hypertensive rat. Oxid. Med. Cell. Longev. 2014, 2014, 10. [Google Scholar] [CrossRef]
- Winiarska, K.; Focht, D.; Sierakowski, B.; Lewandowski, K.; Orlowska, M.; Usarek, M. NADPH oxidase inhibitor, apocynin, improves renal glutathione status in Zucker diabetic fatty rats: A comparison with melatonin. Chem. Biol. Interact. 2014, 218, 12–19. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Luchese, C.; Pinton, S.; Nogueira, C.W. Brain and lungs of rats are differently affected by cigarette smoke exposure: Antioxidant effect of an organoselenium compound. Pharmacol. Res. 2009, 59, 194–201. [Google Scholar] [CrossRef]
- Annuk, M.; Zilmer, M.; Lind, L.; Linde, T.; Fellström, B. Oxidative stress and endothelial function in chronic renal failure. J. Am. Soc. Nephrol. 2001, 12, 2747–2752. [Google Scholar] [CrossRef]
- Koracevic, D.; Koracevic, G.; Djordjevic, V.; Andrejevic, S.; Cosic, V. Method for the measurement of antioxidant activity in human fluids. J. Clin. Pathol. 2001, 54, 356–361. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.F.; Yao, J.; Gao, S.G.; Wang, X.S.; Peng, X.Q.; Yang, Y.T.; Feng, X.S. Nrf2 overexpression predicts prognosis and 5-FU resistance in gastric cancer. Asian Pac. J. Cancer Prev. 2013, 4, 5231–5235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.-C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Liang, W.; Menke, A.L.; Driessen, A.; Koek, G.H.; Lindeman, J.H.; Stoop, R.; Havekes, L.M.; Kleeman, R.; den Hoek, A.M. Establishment of a general NAFLD scoring system for rodent models and comparison to human liver pathology. PLoS ONE 2014, 9, e115922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, K.H.; Umashanker, D.; Igel, L.I.; Kumar, R.B.; Aronne, L.J. Obesity pharmacotherapy. Med. Clin. North. Am. 2018, 102, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Kujawska-Łuczak, M.; Musialik, K.; Szulińska, M.; Swora-Cwynar, E.; Kargulewicz, A.; Grzymisławska, M.; Pupek-Musialik, D.; Bogdański, P. The effect of orlistat versus metformin on body composition and insulin resistance in obese premenopausal women: 3-month randomized prospective open-label study. Arch. Med. Sci. 2017, 13, 725–731. [Google Scholar] [CrossRef] [Green Version]
- Gorgojo-Martínez, J.J.; Basagoiti-Carreño, B.; Sanz-Velasco, A.; Serrano-Moreno, C.; Almodóvar-Ruiz, F. Effectiveness and tolerability of orlistat and liraglutide in patients with obesity in a real-world setting: The XENSOR Study. Int. J. Clin. Pract. 2019, 73, e13399. [Google Scholar] [CrossRef]
- Ke, J.; An, Y.; Cao, B.; Lang, J.; Wu, N.; Zhao, D. Orlistat-induced gut microbiota modification in obese mice. Evid. Based Complement. Alternat. Med. 2020, 2020, 9. [Google Scholar] [CrossRef] [Green Version]
- Sahebkar, A.; Simental-Mendía, L.E.; Reiner, Ž.; Kovanen, P.T.; Simental-Mendía, M.; Bianconi, V.; Pirro, M. Effect of orlistat on plasma lipids and body weight: A systematic review and meta-analysis of 33 randomized controlled trials. Pharmacol. Res. 2017, 122, 53–65. [Google Scholar] [CrossRef]
- Druce, M.; Bloom, S.R. The regulation of appetite. Arch. Dis. Child. 2006, 91, 183–187. [Google Scholar] [CrossRef]
- Azman, K.F.; Amom, Z.; Azlan, A.; Esa, N.M.; Ali, R.M.; Shah, Z.M.; Kadir, K.K.A. Antiobesity effect of Tamarindus indica L. pulp aqueous extract in high-fat diet-induced obese rats. J. Nat. Med. 2012, 66, 333–342. [Google Scholar] [CrossRef]
- Othman, Z.A.; Zakaria, Z.; Suleiman, J.B.; Ghazali, W.S.W.; Mohamed, M. Anti-atherogenic effects of orlistat on obesity-induced vascular oxidative stress rat model. Antioxidants 2021, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Pamuk, B.; Yilmaz, H.; Kebapçilar, L.; Kirbiyik, H.; Alacacioǧlu, A.; Bozkaya, G.; Pamuk, G.; Demirpence, M. The effect of orlistat and weight loss diet on plasma ghrelin and obestatin. J. Res. Med. Sci. 2018, 23, 95. [Google Scholar] [CrossRef] [PubMed]
- Ellrichmann, M.; Kapelle, M.; Ritter, P.R.; Holst, J.J.; Herzig, K.H.; Schmidt, W.E.; Schmitz, F.; Meier, J.J. Orlistat inhibition of intestinal lipase acutely increases appetite and attenuates postprandial glucagon-like peptide-1-(7-36)-amide-1, cholecystokinin, and peptide YY concentrations. J. Clin. Endocrinol. Metab. 2008, 93, 3995–3998. [Google Scholar] [CrossRef] [Green Version]
- Petersen, M.C.; Shulman, G.I. Mechanism of insulin action and insulin resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [Green Version]
- Meijer, R.I.; Barrett, E.J. The insulin receptor mediates insulin’s early plasma clearance by liver, muscle, and kidney. Biomedicines 2021, 9, 37. [Google Scholar] [CrossRef]
- Kuipers, E.N.; Held, N.M.; Panhuis, W.i.h.; Modder, M.; Ruppert, P.M.M.; Kersten, S.; Kooijman, S.; Guigas, B.; Houtkooper, R.H.; Rensen, P.C.N.; et al. A single day of high-fat diet feeding induces lipid accumulation and insulin resistance in brown adipose tissue in mice. Am. J. Physiol. Endocrinol. Metab. 2019, 317, E820–E830. [Google Scholar] [CrossRef]
- Kothari, V.; Luo, Y.; Tornabene, T.; O’Neill, A.M.; Greene, M.W.; Geetha, T.; Babu, J.R. High fat diet induces brain insulin resistance and cognitive impairment in mice. Biochim. Biophys. Acta Mol. Basis Dis. 2017, 1863, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Svedberg, J.; Strömblad, G.; Wirth, A.; Smith, U.; Björntorp, P. Fatty acids in the portal vein of the rat regulate hepatic insulin clearance. J. Clin. Investig. 1991, 88, 2054–2058. [Google Scholar] [CrossRef]
- Ter Horst, K.W.; Gilijamse, P.W.; Versteeg, R.I.; Ackermans, M.T.; Nederveen, A.J.; la Fleur, S.E.; Romijn, J.A.; Nieuwdorp, M.; Zhang, D.; Samuel, V.T.; et al. Hepatic diacylglycerol-associated protein kinase Cε translocation links hepatic steatosis to hepatic insulin resistance in humans. Cell Rep. 2017, 19, 1997–2004. [Google Scholar] [CrossRef] [Green Version]
- Antuna-Puente, B.; Feve, B.; Fellahi, S.; Bastard, J.P. Adipokines: The missing link between insulin resistance and obesity. Diabetes Metab. 2008, 34, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Kamada, Y.; Takehara, T.; Hayashi, N. Adipocytokines and liver disease. J. Gastroenterol. 2008, 43, 811–822. [Google Scholar] [CrossRef]
- Gatselis, N.K.; Ntaios, G.; Makaritsis, K.; Dalekos, G.N. Adiponectin: A key playmaker adipocytokine in non-alcoholic fatty liver disease. Clin. Exp. Med. 2014, 14, 121–131. [Google Scholar] [CrossRef]
- Fjære, E.; Myrmel, L.S.; Lützhøft, D.O.; Andersen, H.; Holm, J.B.; Kiilerich, P.; Hannisdal, R.; Liaset, B.; Kristiansen, K.; Madsen, L. Effects of exercise and dietary protein sources on adiposity and insulin sensitivity in obese mice. J. Nutr. Biochem. 2019, 66, 98–109. [Google Scholar] [CrossRef]
- Silva, M.I.B.; Klein, M.R.S.T.; Cardoso, É.S.; Costa, M.S.; Martins, C.J.M.; Bregman, R. Synergistic effect of inflammatory cytokines and body adiposity on insulin resistance and endothelial markers in patients with stages 3–5 chronic kidney disease. J. Ren. Nutr. 2020, 30, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Boyko, E.J.; Fujimoto, W.Y.; Kahn, S.E.; Leonetti, D.L. Low plasma adiponectin concentrations predict increases in visceral adiposity and insulin resistance. J. Clin. Endocrinol. Metab. 2017, 102, 4626–4633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suleiman, J.B.; Nna, V.U.; Othman, Z.A.; Zakaria, Z.; Bakar, A.B.A.; Mohamed, M. Orlistat attenuates obesity-induced decline in steroidogenesis and spermatogenesis by up-regulating steroidogenic genes. Andrology 2020, 8, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Kujawska-Luczak, M.; Szulinska, M.; Skrypnik, D.; Musialik, K.; Swora-Cwynar, E.; Kregielska-Narozna, M.; Markuszewski, L.; Grzymislawska, M.; Bogdanski, P. The influence of orlistat, metformin and diet on serum levels of insulin-like growth factor-1 in obeses women with and without insulin resistance. J. Physiol. Pharmacol. 2018, 69, 737–745. [Google Scholar] [CrossRef]
- Annamalai, S.; Mohanam, L.; Raja, V.; Dev, A.; Prabhu, V. Antiobesity, antioxidant and hepatoprotective effects of Diallyl trisulphide (DATS) alone or in combination with Orlistat on HFD induced obese rats. Biomed. Pharmacother. 2017, 93, 81–87. [Google Scholar] [CrossRef]
- Evans, J.L.; Maddux, B.A.; Goldfine, I.D. The molecular basis for oxidative stress-induced insulin resistance. Antioxid. Redox Signal. 2005, 7, 1040–1052. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Żebrowska, E.; Zalewska, A.; Chabowski, A. Redox balance, antioxidant defense, and oxidative damage in the hypothalamus and cerebral cortex of rats with high fat diet-induced insulin resistance. Oxid. Med. Cell. Longev. 2018, 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Ferré, P.; Foufelle, F. Hepatic steatosis: A role for de novo lipogenesis and the transcription factor SREBP-1c. Diabetes Obes. Metab. 2010, 12, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Coniglio, R.I.; Meroño, T.; Montiel, H.; Malaspina, M.M.; Salgueiro, A.M.; Otero, J.C.; Ferraris, R.; Schreier, L.; Brites, F.; Rosso, L.G. HOMA-IR and non-HDL-C as predictors of high cholesteryl ester transfer protein activity in patients at risk for type 2 diabetes. Clin. Biochem. 2012, 45, 566–570. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, R.H.; Elnour, W.A. Comparative evaluation of the efficacy of ginger and orlistat on obesity management, pancreatic lipase and liver peroxisomal catalase enzyme in male albino rats. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 75–83. [Google Scholar] [PubMed]
- Jaradat, N.; Zaid, A.N.; Hussein, F.; Zaqzouq, M.; Aljammal, H.; Ayesh, O. Anti-lipase potential of the organic and aqueous extracts of ten traditional edible and medicinal plants in Palestine; a comparison study with orlistat. Medicines 2017, 4, 89. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, R.M.; Zarricueta, M.L.; Moreira, D.K.T.; de Morais, T.R.; Rizzardi, K.F.; Parisotto, T.M.; de Pádua Gandra, R.L.; Zuin, J.C.; e Paiva Caria, C.R.; Macedo, J.A.; et al. Structured lipid containing behenic acid versus orlistat for weight loss: An experimental study in mice. PharmaNutrition 2020, 14, 100213. [Google Scholar] [CrossRef]
- Nakou, E.S.; Filippatos, T.D.; Kiortsis, D.N.; Derdemezis, C.S.; Tselepis, A.D.; Mikhailidis, D.P.; Elisaf, M.S. The effects of ezetimibe and orlistat, alone or in combination, on high-density lipoprotein (HDL) subclasses and HDL-associated enzyme activities in overweight and obese patients with hyperlipidaemia. Expert Opin. Pharmacother. 2008, 9, 3151–3158. [Google Scholar] [CrossRef]
- Mirza, M.S. Obesity, visceral fat, and NAFLD: Querying the role of adipokines in the progression of nonalcoholic fatty liver disease. ISRN Gastroenterol. 2011, 2011, 11. [Google Scholar] [CrossRef] [Green Version]
- Farrell, G.C.; Larter, C.Z. Nonalcoholic fatty liver disease: From steatosis to cirrhosis. Hepatology 2006, 43, S99–S112. [Google Scholar] [CrossRef]
- Chan, D.C.; Watts, G.F.; Ng, T.W.K.; Hua, J.; Song, S.; Barrett, P.H.R. Measurement of liver fat by magnetic resonance imaging: Relationships with body fat distribution, insulin sensitivity and plasma lipids in healthy men. Diabetes Obes. Metab. 2006, 8, 698–702. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, H.; Zheng, H.; Jiang, Y. Hepatic inflammation scores correlate with common carotid intima-media thickness in rats with NAFLD induced by a high-fat diet. BMC Vet. Res. 2014, 10, 162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuschwander-Tetri, B.A. Non-alcoholic fatty liver disease. BMC Med. 2017, 15, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paschos, P.; Paletas, K. Non alcoholic fatty liver disease and metabolic syndrome. Hippokratia 2009, 13, 9–19. [Google Scholar]
- Harrison, S.A.; Fincke, C.; Helinski, D.; Torgerson, S.; Hayashi, P. A pilot study of orlistat treatment in obese, non-alcoholic steatohepatitis patients. Aliment. Pharmacol. Ther. 2004, 20, 623–628. [Google Scholar] [CrossRef]
- Al Humayed, S. Protective and therapeutic effects of Crataegus aronia in non-alcoholic fatty liver disease. Arch. Physiol. Biochem. 2017, 123, 23–30. [Google Scholar] [CrossRef]
- Eliades, M.; Spyrou, E.; Agrawal, N.; Lazo, M.; Brancati, F.L.; Potter, J.J.; Koteish, A.A.; Clark, J.M.; Guallar, E.; Hernaez, R. Meta-analysis: Vitamin D and non-alcoholic fatty liver disease. Aliment. Pharmacol. Ther. 2013, 38, 246–254. [Google Scholar] [CrossRef]
- Van Hoof, V.O.; Deng, J.T.; De Broe, M.E. How do plasma membranes reach the circulation? Clin. Chim. Acta 1997, 266, 23–31. [Google Scholar] [CrossRef]
- Zelber-Sagi, S.; Kessler, A.; Brazowsky, E.; Webb, M.; Lurie, Y.; Santo, M.; Leshno, M.; Blendis, L.; Halpern, Z.; Oren, R. A double-blind randomized placebo-controlled trial of orlistat for the treatment of nonalcoholic fatty liver disease. Clin. Gastroenterol. Hepatol. 2006, 4, 639–644. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, S.; Jiang, X.; Wang, Y.-H.; Li, F.; Wang, Y.-G.; Zheng, Y.; Cai, L. The role of the Nrf2/Keap1 pathway in obesity and metabolic syndrome. Rev. Endocr. Metab. Disord. 2015, 16, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Castegna, A.; Drake, J.; Pocernich, C.; Butterfield, D.A. Protein carbonyl levels—An assessment of protein oxidation. In Methods in Biological Oxidative Stress; Hensley, K., Floyd, R.A., Eds.; Humana Press: Totowa, NJ, USA, 2003; pp. 161–168. [Google Scholar]
- Chen, X.L.; Gong, L.Z.; Xu, J.X. Antioxidative activity and protective effect of probiotics against high-fat diet-induced sperm damage in rats. Animal 2012, 7, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Bello, M.; Morales-González, J.A. Molecular recognition between potential natural inhibitors of the Keap1-Nrf2 complex. Int. J. Biol. Macromol. 2017, 105, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Bhakkiyalakshmi, E.; Dineshkumar, K.; Karthik, S.; Sireesh, D.; Hopper, W.; Paulmurugan, R.; Ramkumar, K.M. Pterostilbene-mediated Nrf2 activation: Mechanistic insights on Keap1:Nrf2 interface. Bioorg. Med. Chem. 2016, 24, 3378–3386. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Composition (g/100 g) | Energy (Kcal) | Fat | Carbohydrate | Protein | Ash |
|---|---|---|---|---|---|
| Normal diet | 318.8 | 12 | 64 | 24 | 6.9 |
| HFD | 516.5 | 31 | 46 | 12 | 3.8 |
| N | HFD | HFD + O | OB + O | |
|---|---|---|---|---|
| Lee obesity index | 304.60 ± 1.43 | 334.20 ± 2.66 a | 310.00 ± 3.77 b | 314.50 ± 2.91 b |
| Initial body weight (g) | 218.20 ± 6.26 | 233.50 ± 7.53 | 226.60 ± 6.23 | 216.80 ± 13.82 |
| Final body weight (g) | 352.60 ± 13.68 | 474.00 ± 12.12 a | 403.5 ± 14.44 b | 390.80 ± 15.03 b |
| Body weight gain (g) | 93.17 ± 8.97 | 213.20 ± 12.95 a | 159.6 ± 6.92 a,b | 152.8 ± 8.31 a,b |
| Food intake (g/day) | 20.79 ± 0.56 | 21.22 ± 0.82 | 18.58 ±0.71 | 19.28 ± 0.66 |
| Calorie intake (kJ/day) | 276.50 ± 6.81 | 458.70 ± 17.72 a | 401.40 ± 15.39 a | 409.30 ± 18.64 a |
| N | HFD | HFD + O | OB + O | |
|---|---|---|---|---|
| Absolute liver weight (g) | 8.73 ± 0.29 | 15.74 ± 0.56 a | 13.20 ± 0.61 a,b | 12.29 ± 0.41 a,b |
| Relative liver weight (g/body weight) | 2.44 ± 0.03 | 3.44 ± 0.09 a | 3.12 ± 0.11 a,b | 2.80 ±0.07 a,b |
| Epididymal adipose tissue weight (g) | 2.89 ± 0.14 | 12.20 ± 1.22 a | 5.52 ± 0.82 b | 5.20 ± 0.42 b |
| Peritoneal adipose tissue weight (g) | 2.66 ± 0.36 | 14.14 ± 2.45 a | 7.84 ± 1.67 b | 5.51 ± 0.32 b |
| Perirenal adipose weight tissue (g) | 0.34 ± 0.03 | 0.54 ± 0.03 a | 0.40 ± 0.02 b | 0.41 ± 0.02 b |
| Total adipose tissue weight (g) | 6.87 ± 0.47 | 26.78 ± 4.34 a | 13.11 ± 2.75 b | 10.08 ± 1.28 b |
| Parameters | N | HFD | HFD + O | OB + O |
|---|---|---|---|---|
| TBARS (nmol/mg protein) | 2.47 ± 0.07 | 6.25 ± 0.61 a | 2.07 ± 0.27 b | 1.82 ± 0.11 b |
| NO (µmol/g protein) | 0.84 ± 0.04 | 1.20 ± 0.07 a | 0.92 ± 0.03 b | 0.91 ± 0.06 b |
| PCO (mmol/mg protein) | 0.80 ± 0.05 | 1.80 ± 0.07 a | 0.86 ±0.06 b | 0.95 ± 0.11 b |
| SOD (unit/mg protein) | 5.65 ± 0.27 | 2.12 ± 0.51 a | 5.08 ± 0.37 b | 4.09 ± 0.50 b |
| CAT (unit/mg protein) | 28.28 ± 3.56 | 6.70 ± 1.14 a | 16.45 ± 0.80 b | 21.77 ± 3.03 b |
| GPx (unit/mg protein) | 2.45 ± 0.20 | 0.79 ± 0.16 a | 2.416 ± 0.18 b | 1.79 ± 0.11 a,b |
| GST (unit/mg protein) | 5.49 ± 0.64 | 1.47 ± 0.09 a | 4.26 ± 0.34 b | 4.62 ± 0.62 b |
| GSH (nmol/mg protein) | 3.29 ± 0.08 | 2.37 ± 0.10 a | 3.06 ± 0.06 b | 2.91 ± 0.19 b |
| GR (unit/mg protein) | 15.36 ± 0.46 | 11.40 ± 0.36 a | 15.55 ± 0.60 b | 14.61 ± 0.87 b |
| TAC (nmol/mg protein) | 83.11 ± 2.59 | 65.92 ± 0.56 a | 84.11 ± 3.04 b | 83.70 ± 6.51 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakaria, Z.; Othman, Z.A.; Bagi Suleiman, J.; Jalil, N.A.C.; Ghazali, W.S.W.; Mohamed, M. Protective and Therapeutic Effects of Orlistat on Metabolic Syndrome and Oxidative Stress in High-Fat Diet-Induced Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Rats: Role on Nrf2 Activation. Vet. Sci. 2021, 8, 274. https://doi.org/10.3390/vetsci8110274
Zakaria Z, Othman ZA, Bagi Suleiman J, Jalil NAC, Ghazali WSW, Mohamed M. Protective and Therapeutic Effects of Orlistat on Metabolic Syndrome and Oxidative Stress in High-Fat Diet-Induced Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Rats: Role on Nrf2 Activation. Veterinary Sciences. 2021; 8(11):274. https://doi.org/10.3390/vetsci8110274
Chicago/Turabian StyleZakaria, Zaida, Zaidatul Akmal Othman, Joseph Bagi Suleiman, Nur Asyilla Che Jalil, Wan Syaheedah Wan Ghazali, and Mahaneem Mohamed. 2021. "Protective and Therapeutic Effects of Orlistat on Metabolic Syndrome and Oxidative Stress in High-Fat Diet-Induced Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Rats: Role on Nrf2 Activation" Veterinary Sciences 8, no. 11: 274. https://doi.org/10.3390/vetsci8110274
APA StyleZakaria, Z., Othman, Z. A., Bagi Suleiman, J., Jalil, N. A. C., Ghazali, W. S. W., & Mohamed, M. (2021). Protective and Therapeutic Effects of Orlistat on Metabolic Syndrome and Oxidative Stress in High-Fat Diet-Induced Metabolic Dysfunction-Associated Fatty Liver Disease (MAFLD) in Rats: Role on Nrf2 Activation. Veterinary Sciences, 8(11), 274. https://doi.org/10.3390/vetsci8110274