
Sperm Global DNA Methylation (SGDM) in Semen of Healthy Dogs

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Evaluation of Semen Quality
2.2. Head Area Analysis
2.3. Leukocytes Depletion and Spermatozoa Purification
2.4. DNA Extraction
2.5. 5-mC DNA ELISA
2.6. Determination of DNA Integrity
2.7. Statistical Analysis
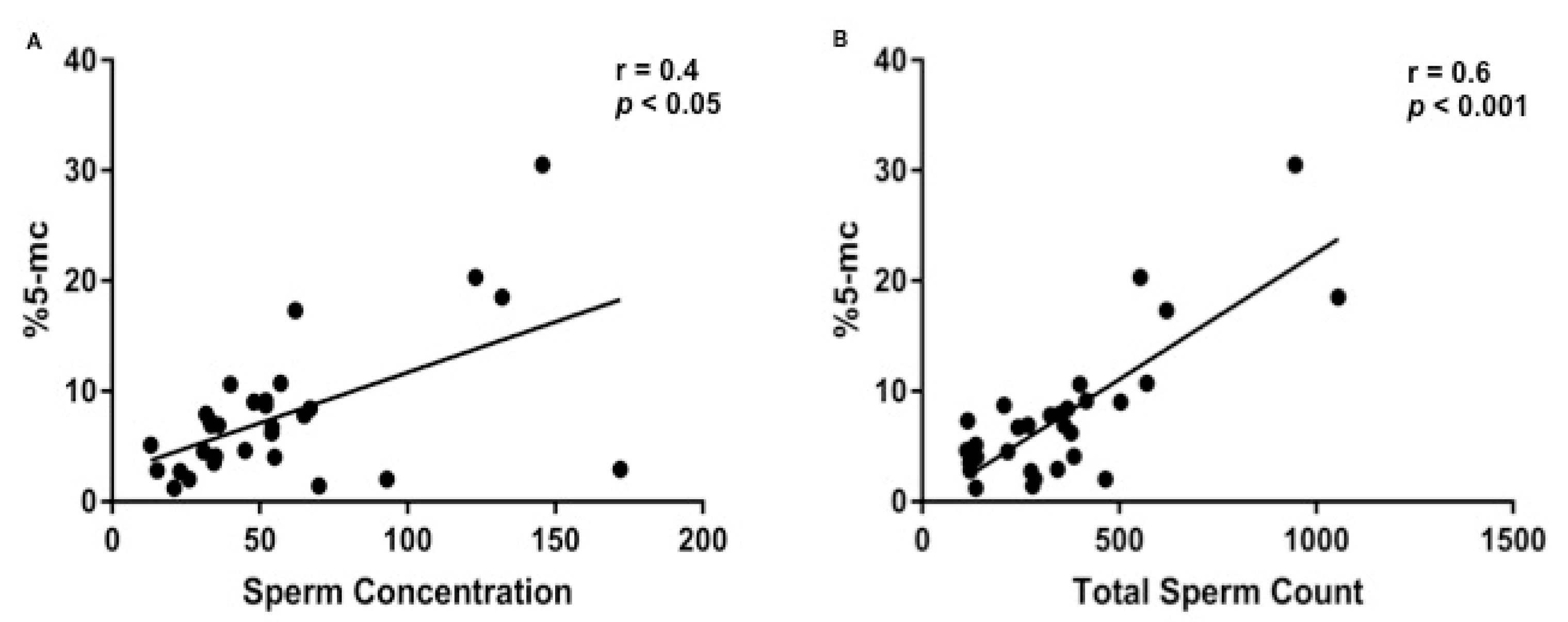
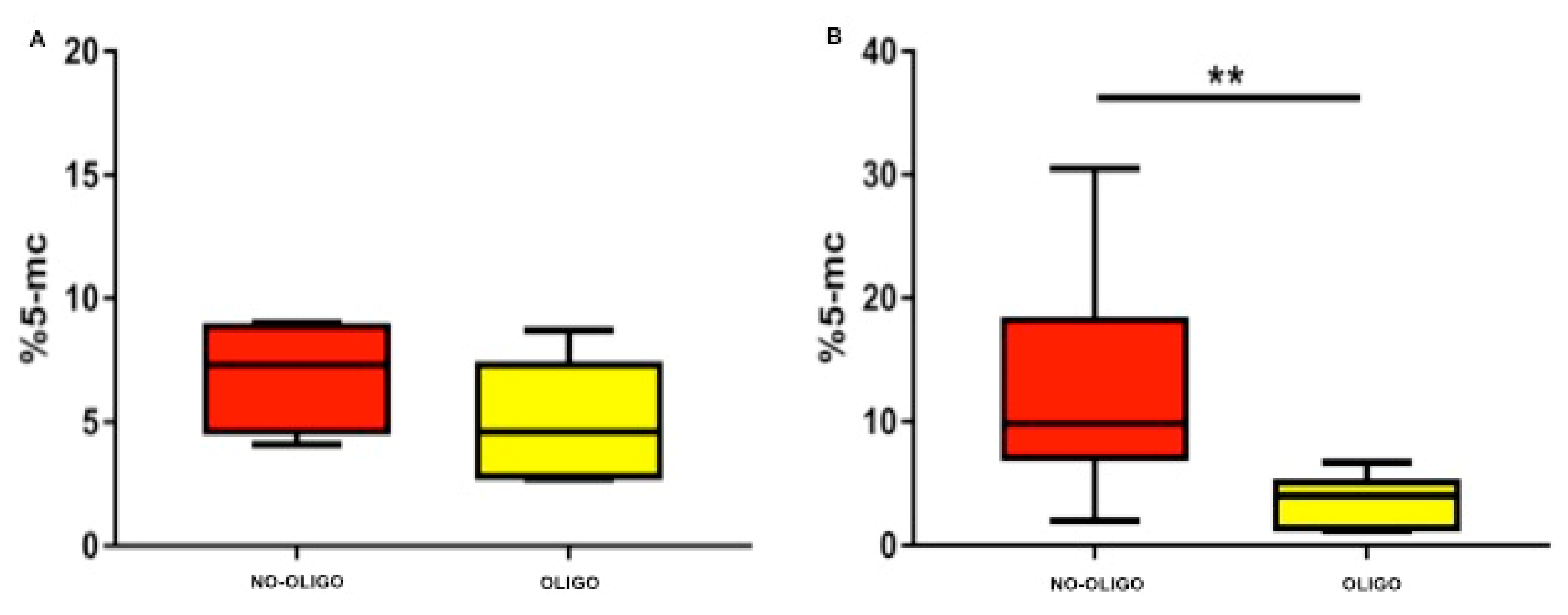
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Albarella, S.; D’Anza, E.; Galdiero, G.; Esposito, L.; De Biase, D.; Paciello, O.; Ciotola, F.; Peretti, V. Cytogenetic Analyses in Ewes with Congenital Abnormalities of the Genital Apparatus. Animals 2019, 9, 776. [Google Scholar] [CrossRef] [Green Version]
- Albarella, S.; Lorenzi, L.; Rossi, E.; Prisco, F.; Riccardi, M.G.; Restucci, B.; Ciotola, F.; Parma, P. Analysis of XX SRY-Negative Sex Reversal Dogs. Animals 2020, 10, 1667. [Google Scholar] [CrossRef]
- Albarella, S.; Ciotola, F.; D’Anza, E.; Coletta, A.; Zicarelli, L.; Peretti, V. Congenital Malformations in River Buffalo (Bubalus bubalis). Animals 2017, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Krzemińska, P.; D’Anza, E.; Ciotola, F.; Paciello, O.; Restucci, B.; Peretti, V.; Albarella, S.; Switonski, M. Polymorphisms of MAMLD1, SRD5A2, and AR Candidate Genes in Seven Dogs (78,XY; SRY-Positive) Affected by Hypospadias or Cryptorchidism. Sex Dev. 2019, 13, 92–98. [Google Scholar] [CrossRef]
- Szczerbal, I.; Nowacka-Woszuk, J.; Albarella, S.; Switonski, M. Technical note: Droplet digital PCR as a new molecular method for a simple and reliable diagnosis of freemartinism in cattle. J. Dairy Sci. 2019, 102, 10100–10104. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Stachowiak, M.; Nowacka-Woszuk, J.; Dzimira, S.; Szczepanska, K.; Switonski, M. Disorder of sex development in a cat with chromosome mosaicism 37,X/38,X,r(Y). Reprod. Domest. Anim. 2017, 52, 914–917. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Derar, D.R.; AlShahed, M. Management strategies, reproductive performance and causes of infertility in sheep flocks in the central region of Saudi Arabia. Trop. Anim. Health Prod. 2019, 52, 1691–1697. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.; Chakravarty, A.K.; Singh, A.; Upadhyay, A.; Singh, M.; Yousuf, S. Effect of heat stress on reproductive performances of dairy cattle and buffaloes: A review. Vet. World 2016, 9, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuente-Lara, A.; Hesser, A.; Christensen, B.; Gonzales, K.; Meyers, S. Effects from aging on semen quality of fresh and cryopreserved semen in Labrador Retrievers. Theriogenology 2019, 132, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Hollinshead, F.K.; Hanlon, D.W. Factors affecting the reproductive performance of bitches: A prospective cohort study involving 1203 inseminations with fresh and frozen semen. Theriogenology 2017, 101, 62–72. [Google Scholar] [CrossRef]
- Rahman, M.S.; Kwon, W.S.; Pang, M.G. Prediction of male fertility using capacitation-associated proteins in spermatozoa. Mol. Reprod. Dev. 2017, 84, 749–759. [Google Scholar] [CrossRef] [Green Version]
- D’Anza, E.; Albarella, S.; Galdiero, G.; Tafuri, S.; Del Prete, C.; Cocchia, N.; Ciani, F.; Mastellone, V.; Pasolini, M.P.; Carotenuto, D.; et al. DNA fragmentation and morphometric studies in sperm of stallions supplemented with maca (Lepidium meyenii). Zygote 2021, 1–6. [Google Scholar] [CrossRef]
- Albarella, S.; Ciotola, F.; Coletta, A.; Genualdo, V.; Iannuzzi, L.; Peretti, V. A new translocation t(1p;18) in an Italian Mediterranean river buffalo (Bubalus bubalis, 2n = 50) bull: Cytogenetic, fertility and inheritance studies. Cytogenet. Genome Res. 2013, 139, 17–21. [Google Scholar] [CrossRef]
- Del Prete, C.; Tafuri, S.; Ciani, F.; Pasolini, M.P.; Ciotola, F.; Albarella, S.; Carotenuto, D.; Peretti, V.; Cocchia, N. Influences of dietary supplementation withLepidium meyenii(Maca) on stallion sperm production and on preservation of sperm quality during storage at 5 °C. Andrology 2018, 6, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Kolster, K.A. Evaluation of Canine Sperm and Management of Semen Disorders. Vet. Clin. N. Am. Small Anim. Pract. 2018, 48, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Urdinguio, R.G.; Bayón, G.F.; Dmitrijeva, M.; Toraño, E.G.; Bravo, C.; Fraga, M.F.; Bassas, L.; Larriba, S.; Fernández, A.F. Aberrant DNA methylation patterns of spermatozoa in men with unexplained infertility. Hum. Reprod. 2015, 30, 1014–1028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egger, G.; Liang, G.; Aparicio, A.; Jones, P.A. Epigenetics in human disease and prospects for epigenetic therapy. Nature 2004, 429, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Doerksen, T.; Trasler, J.M. Developmental exposure of male germ cells to 5-azacytidine results in abnormal preimplantation development in rats. Biol. Reprod. 1996, 55, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Holliday, R. The inheritance of epigenetic defects. Science 1987, 238, 163–170. [Google Scholar] [CrossRef]
- Mussa, A.; Russo, S.; de Crescenzo, A.; Freschi, A.; Calzari, L.; Maitz, S.; Macchiaiolo, M.; Molinatto, C.; Baldassarre, G.; Mariani, M.; et al. Fetal growth patterns in Beckwith-Wiedemann syndrome. Clin. Genet. 2016, 90, 21–27. [Google Scholar] [CrossRef]
- de Angelis, C.; Galdiero, M.; Pivonello, C.; Salzano, C.; Gianfrilli, D.; Piscitelli, P.; Lenzi, A.; Colao, A.; Pivonello, R. The environment and male reproduction: The effect of cadmium exposure on reproductive function and its implication in fertility. Reprod. Toxicol. 2017, 73, 105–127. [Google Scholar] [CrossRef]
- Genesio, R.; Melis, D.; Gatto, S.; Izzo, A.; Ronga, V.; Cappuccio, G.; Lanzo, A.; Andria, G.; D’Esposito, M.; Matarazzo, M.R.; et al. Variegated silencing through epigenetic modifications of a large Xq region in a case of balanced X;2 translocation with Incontinentia Pigmenti-like phenotype. Epigenetics 2011, 6, 1242–1247. [Google Scholar] [CrossRef] [Green Version]
- Herceg, Z.; Vaissière, T. Epigenetic mechanisms and cancer: An interface between the environment and the genome. Epigenetics 2011, 6, 804–819. [Google Scholar] [CrossRef]
- Liu, L.; Rando, T.A. Manifestations and mechanisms of stem cell aging. J. Cell Biol. 2011, 193, 257–266. [Google Scholar] [CrossRef]
- Kelsey, G.; Feil, R. New insights into establishment and maintenance of DNA methylation imprints in mammals. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2013, 368, 20110336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seisenberger, S.; Peat, J.R.; Reik, W. Conceptual links between DNA methylation reprogramming in the early embryo and primordial germ cells. Curr. Opin. Cell Biol. 2013, 25, 281–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissonnas, C.C.; Abdalaoui, H.E.; Haelewyn, V.; Fauque, P.; Dupont, J.M.; Gut, I.; Vaiman, D.; Jouannet, P.; Tost, J.; Jammes, H. Specific epigenetic alterations of IGF2-H19 locus in spermatozoa from infertile men. Eur. J. Hum. Genet. 2010, 18, 73–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammoud, S.S.; Purwar, J.; Pflueger, C.; Cairns, B.R.; Carrell, D.T. Alterations in sperm DNA methylation patterns at imprinted loci in two classes of infertility. Fertil. Steril. 2010, 94, 1728–1733. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Sato, A.; Otsu, E.; Hiura, H.; Tomatsu, C.; Utsunomiya, T.; Sasaki, H.; Yaegashi, N.; Arima, T. Aberrant DNA methylation of imprinted loci in sperm from oligospermic patients. Hum. Mol. Genet. 2007, 16, 2542–2551. [Google Scholar] [CrossRef]
- Marques, C.J.; Costa, P.; Vaz, B.; Carvalho, F.; Fernandes, S.; Barros, A.; Sousa, M. Abnormal methylation of imprinted genes in human sperm is associated with oligozoospermia. Mol. Hum. Reprod. 2008, 14, 67–74. [Google Scholar] [CrossRef]
- Minor, A.; Chow, V.; Ma, S. Aberrant DNA methylation at imprinted genes in testicular sperm retrieved from men with obstructive azoospermia and undergoing vasectomy reversal. Reproduction 2011, 141, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poplinski, A.; Tüttelmann, F.; Kanber, D.; Horsthemke, B.; Gromoll, J. Idiopathic male infertility is strongly associated with aberrant methylation of MEST and IGF2/H19 ICR1. Int. J. Androl. 2010, 33, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Benchaib, M.; Braun, V.; Ressnikof, D.; Lornage, J.; Durand, P.; Niveleau, A.; Guérin, J. Influence of global sperm DNA methylation on IVF results. Hum. Reprod. 2005, 20, 768–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Hajj, N.; Zechner, U.; Schneider, E.; Tresch, A.; Gromoll, J.; Hahn, T.; Schorsch, M.; Haaf, T. Methylation status of imprinted genes and repetitive elements in sperm DNA from infertile males. Sex Dev. 2011, 5, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.J.; Carvalho, F.; Sousa, M.; Barros, A. Genomic imprinting in disruptive spermatogenesis. Lancet 2004, 363, 1700–1702. [Google Scholar] [CrossRef]
- Ostrander, E.A.; Giniger, E. Semper fidelis: What man’s best friend can teach us about human biology and disease. Am. J. Hum. Genet. 1997, 61, 475–480. [Google Scholar] [CrossRef] [Green Version]
- Tsai, K.L.; Clark, L.A.; Murphy, K.E. Understanding hereditary diseases using the dog and human as companion model systems. Mamm. Genome 2007, 18, 444–451. [Google Scholar] [CrossRef] [Green Version]
- Kutzler, M.A. Semen collection in the dog. Theriogenology 2005, 64, 747–754. [Google Scholar] [CrossRef]
- Björndahl, L.; Barratt, C.L.; Mortimer, D.; Jouannet, P. ‘How to count sperm properly’: Checklist for acceptability of studies based on human semen analysis. Hum. Reprod. 2015, 31, 227–232. [Google Scholar] [CrossRef] [Green Version]
- Root Kustritz, M.V. The value of canine semen evaluation for practitioners. Theriogenology 2007, 68, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Hirai, M.; Boersma, A.; Hoeflich, A.; Wolf, E.; Foll, J.; Aumüller, R.; Braun, J. Objectively measured sperm motility and sperm head morphometry in boars (Sus scrofa): Relation to fertility and seminal plasma growth factors. J. Androl. 2001, 22, 104–110. [Google Scholar]
- Ostermeier, G.; Sargeant, G.; Yandell, B.; Evenson, D.; Parrish, J. Relationship of bull fertility to sperm nuclear shape. J. Androl. 2001, 22, 595–603. [Google Scholar]
- Vicente-Fiel, S.; Palacín, I.; Santolaria, J.; Fantova, E.; Quintin-Casorran, F.; Sevilla-Mur, E. In vitro assessment of sperm quality from rams of high and low field fertility. Anim. Reprod. Sci. 2014, 146, 15–20. [Google Scholar] [CrossRef]
- Waheed, M.; Ghoneim, I.; Abdou, M. Morphometric characteristics of spermatozoa in the arabian horse with regard to season, age, sperm concentration, and fertility. J. Equine Vet. Sci. 2015, 35, 244–249. [Google Scholar] [CrossRef]
- Carrell, D.T.; Aston, K.I. (Eds.) Spermatogenesis: Methods and Protocols; Methods in Molecular Biology; Springer Science + Business Media LLC: Berlin/Heidelberg, Germany, 2013; Volume 927. [Google Scholar]
- Tesi, M.; Sabatini, C.; Vannozzi, I.; Di Petta, G.; Panzani, D.; Camillo, F.; Rota, A. Variables affecting semen quality and its relation to fertility in the dog: A retrospective study. Theriogenology 2018, 118, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Montjean, D.; Zini, A.; Ravel, C.; Belloc, S.; Dalleac, A.; Copin, H.; Boyer, P.; McElreavey, K.; Benkhalifa, M. Sperm global DNA methylation level: Association with semen parameters and genome integrity. Andrology 2015, 3, 235–240. [Google Scholar] [CrossRef]
- Houshdaran, S.; Cortessis, V.K.; Siegmund, K.; Yang, A.; Laird, P.W.; Sokol, R.Z. Widespread epigenetic abnormalities suggest a broad DNA methylation erasure defect in abnormal human sperm. PLoS ONE 2007, 2, e1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacheco, S.E.; Houseman, E.A.; Christensen, B.C.; Marsit, C.J.; Kelsey, K.T.; Sigman, M.; Boekelheide, K. Integrative DNA methylation and gene expression analyses identify DNA packaging and epigenetic regulatory genes associated with low motility sperm. PLoS ONE 2011, 6, e20280. [Google Scholar] [CrossRef]
- Jenkins, T.G.; Aston, K.I.; Cairns, B.R.; Carrell, D.T. Paternal aging and associated intraindividual alterations of global sperm 5-methylcytosine and 5-hydroxymethylcytosine levels. Fertil. Steril. 2013, 100, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Olar, T.T.; Amann, R.P.; Pickett, B.W. Relationships among testicular size, daily production and output of spermatozoa, and extragonadal spermatozoal reserves of the dog. Biol. Reprod. 1983, 29, 1114–1120. [Google Scholar] [CrossRef] [PubMed]
- Ciotola, F.; Albarella, S.; Pasolini, M.; Auletta, L.; Esposito, L.; Iannuzzi, L.; Peretti, V. Molecular and cytogenetic studies in a case of XX SRY-negative sex reversal in an Arabian horse. Sex Dev. 2012, 6, 104–107. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | N° of Animals | Age Range (Y) |
|---|---|---|
| Neapolitan Mastiff | 8 | 2–4 |
| German shepherd | 5 | 1–6 |
| English Bull dog | 4 | 2–5 |
| Dachshund | 2 | 2–9 |
| Beagle | 1 | 8 |
| Caucasian Shepherd dog | 1 | 4 |
| Maremmana Sheepdog | 1 | 5 |
| Pointer | 1 | 3 |
| Kangal | 1 | 2 |
| Labrador retriever | 1 | 2 |
| Half-breed | 5 | 2–8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galdiero, G.; D’Anza, E.; de Angelis, C.; Albarella, S.; Peretti, V.; Pivonello, R.; Ciotola, F. Sperm Global DNA Methylation (SGDM) in Semen of Healthy Dogs. Vet. Sci. 2021, 8, 50. https://doi.org/10.3390/vetsci8030050
Galdiero G, D’Anza E, de Angelis C, Albarella S, Peretti V, Pivonello R, Ciotola F. Sperm Global DNA Methylation (SGDM) in Semen of Healthy Dogs. Veterinary Sciences. 2021; 8(3):50. https://doi.org/10.3390/vetsci8030050
Chicago/Turabian StyleGaldiero, Giacomo, Emanuele D’Anza, Cristina de Angelis, Sara Albarella, Vincenzo Peretti, Rosario Pivonello, and Francesca Ciotola. 2021. "Sperm Global DNA Methylation (SGDM) in Semen of Healthy Dogs" Veterinary Sciences 8, no. 3: 50. https://doi.org/10.3390/vetsci8030050
APA StyleGaldiero, G., D’Anza, E., de Angelis, C., Albarella, S., Peretti, V., Pivonello, R., & Ciotola, F. (2021). Sperm Global DNA Methylation (SGDM) in Semen of Healthy Dogs. Veterinary Sciences, 8(3), 50. https://doi.org/10.3390/vetsci8030050