
Prevalence, Risk Factors, Antimicrobial Resistance and Molecular Characterization of Salmonella in Northeast Tunisia Broiler Flocks

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
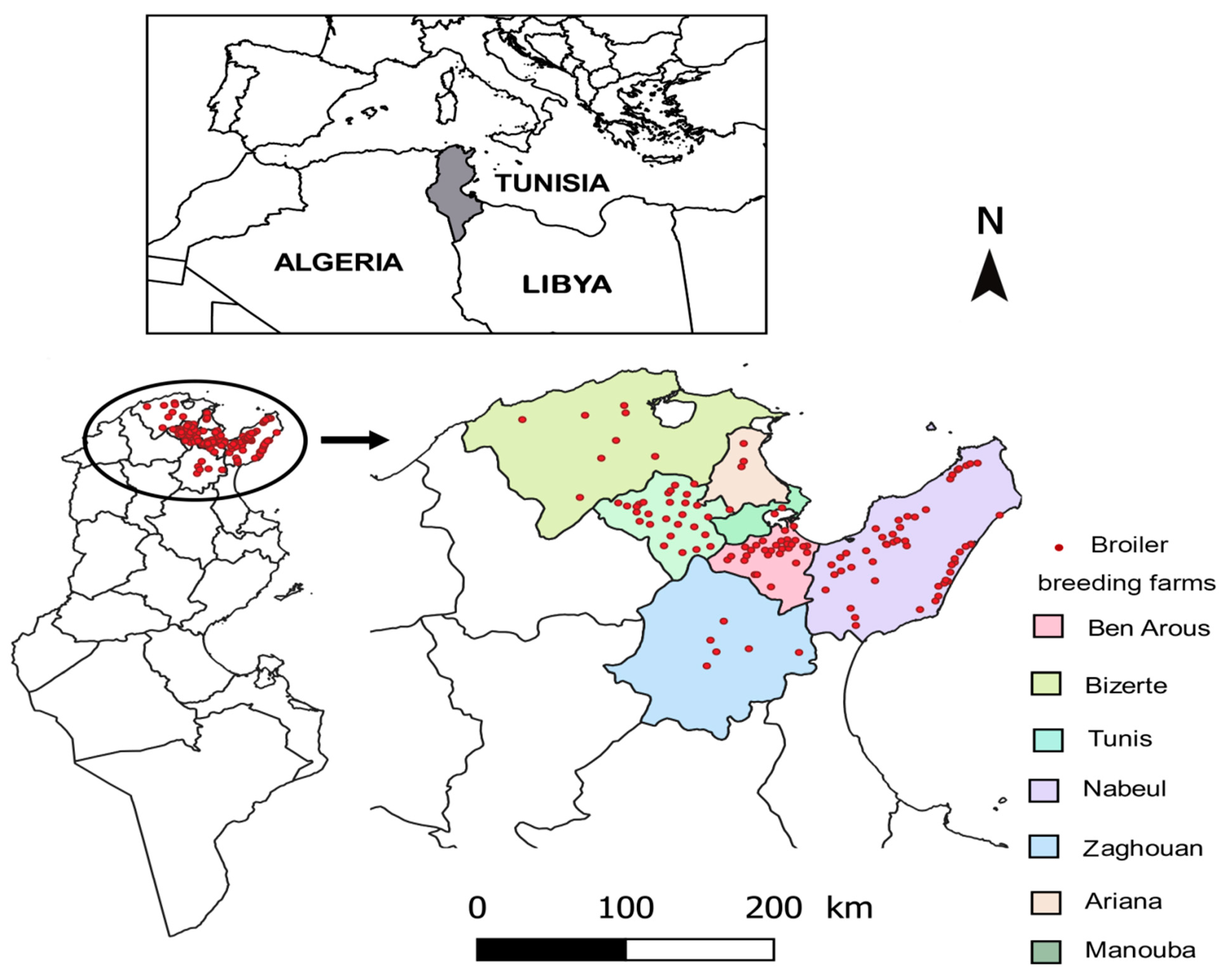
2.1. Choice of Breeding Sites
2.2. Sampling
2.3. Salmonella spp. Screening
2.4. Salmonella Strains Serotyping
2.5. Molecular Study
2.6. Antimicrobial Susceptibility Testing and Identification of Antimicrobial Resistance Genes
2.7. Statistical Analysis
3. Results
3.1. Prevalence of Salmonella spp. in Broiler
3.2. Risk Factors Associated with the Presence of Salmonella in the Fecal Samples of Broiler Chicken
3.3. Prevalence of Virulence Genes
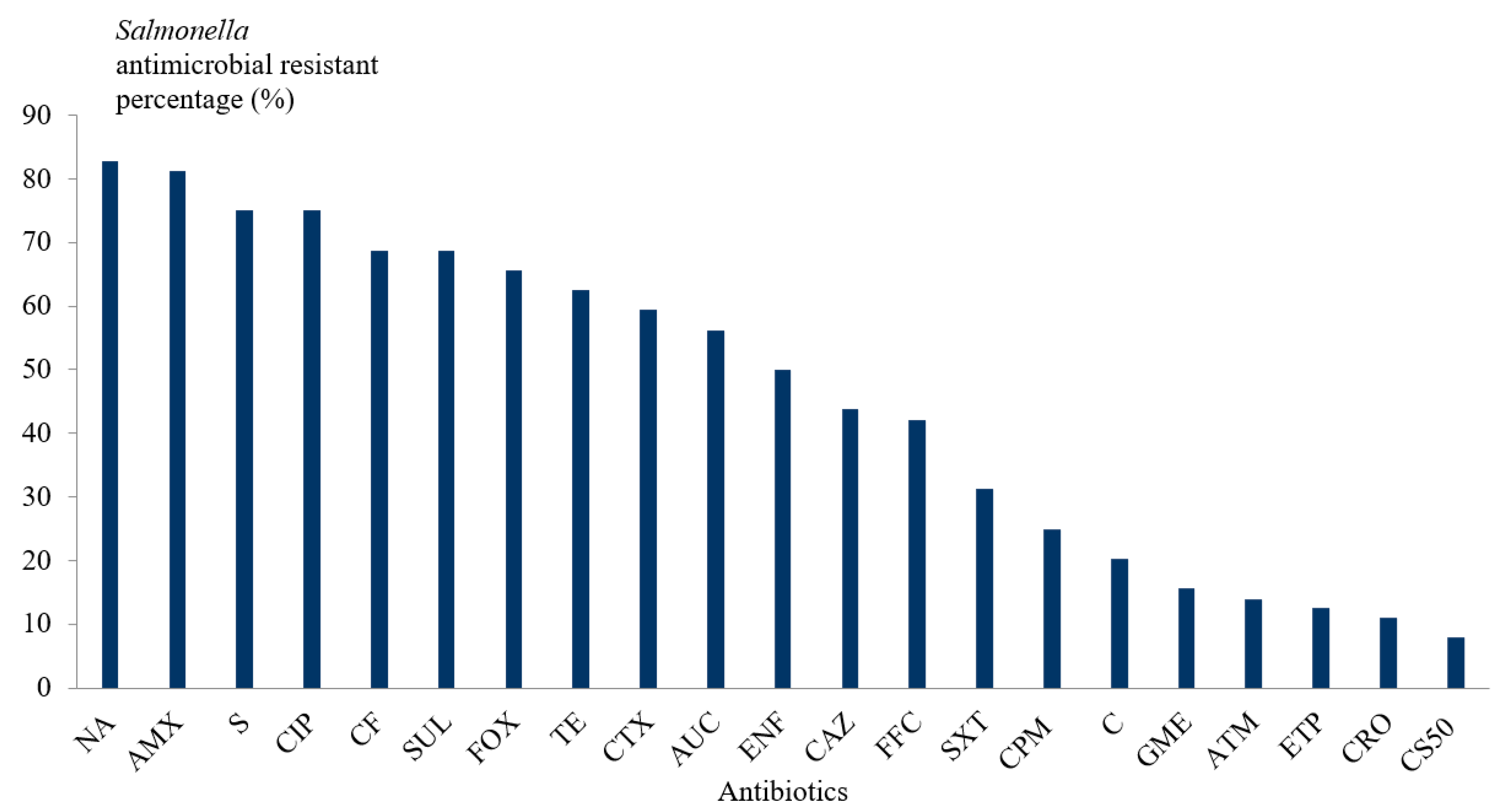
3.4. Antimicrobial Susceptibility Testing
3.5. Prevalence of Antimicrobial Resistance Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gordon, M.A.; Graham, S.M.; Walsh, A.L.; Wilson, L.; Phiri, A.; Molyneux, E.; Zijlstra, E.E.; Heyderman, R.S.; Hart, C.A.; Molyneux, M.E. Epidemics of invasive Salmonella enterica serovar enteritidis and S. enterica serovar typhimurium infection associated with multidrug resistance among adults and children in Malawi. Clin. Infect. Dis. 2008, 46, 963–969. [Google Scholar] [CrossRef] [Green Version]
- García, V.; Mandomando, I.; Ruiz, J.; Herrera-León, S.; Alonso, P.L.; Rodicio, M.R. Salmonella enterica serovars typhimurium and enteritidis causing mixed infections in febrile children in Mozambique. Infect. Drug Resist. 2018, 11, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breurec, S.; Reynaud, Y.; Frank, T.; Farra, A.; Costilhes, G.; Weill, F.O.-X.; Le Hello, S. Serotype distribution and antimicrobial resistance of human Salmonella enterica in Bangui, Central African Republic, from 2004 to 2013. PLoS Negl. Trop. Dis. 2019, 13, e0007917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention Salmonella Homepage CDC. Available online: https://www.cdc.gov/salmonella/index.html%0A (accessed on 30 November 2021).
- Cosby, D.E.; Cox, N.A.; Harrison, M.A.; Wilson, J.L.; Jeff Buhr, R.; Fedorka-Cray, P.J. Salmonella and antimicrobial resistance in broilers: A review. J. Appl. Poult. Res. 2015, 24, 408–426. [Google Scholar] [CrossRef]
- Guerra, B.; Stoicescu, A.-V.; Mulligan, K.; Nagy, K.; Cioacata, G.; Thomas, D. The European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar] [CrossRef]
- Turki, Y.; Ouzari, H.; Mehri, I.; Ben Aissa, R.; Hassen, A. Biofilm formation, virulence gene and multi-drug resistance in Salmonella Kentucky isolated in Tunisia. Food Res. Int. 2012, 45, 940–946. [Google Scholar] [CrossRef]
- Oueslati, W.; Ridha Rjeibi, M.; Ettriqui, A.; Zrelli, S. Serotypes, Virulence and Antibiotic Susceptibility of Salmonella Spp. Strains, Isolated from Poultry Meat Cutting Parts in Greater Tunis (Tunisia). J. Food Nutr. Disord. 2017, 6, 1–6. [Google Scholar] [CrossRef]
- Chaiba, A.; Rhazi Filali, F. Prévalence de la contamination par Salmonella des élevages de poulet de chair au Maroc. Cah. Agric. 2016, 25, 35007. [Google Scholar] [CrossRef] [Green Version]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease Control and Prevention Centers For Disease Control and PreventionGonorrhea. Available online: http://www.cdc.gov/std/gonorrhea/stdfact-gonorrhea.htm (accessed on 20 March 2021).
- Kagambèga, A.; Thibodeau, A.; Trinetta, V.; Soro, D.K.; Sama, F.N.; Bako, É.; Bouda, C.S.; Wereme N’Diaye, A.; Fravalo, P.; Barro, N. Salmonella spp. and Campylobacter spp. in poultry feces and carcasses in Ouagadougou, Burkina Faso. Food Sci. Nutr. 2018, 6, 1601–1606. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhou, X.; Jiang, Z.; Qi, Y.; Ed-dra, A.; Yue, M. Epidemiological Investigation and Antimicrobial Resistance Profiles of Salmonella Isolated From Breeder Chicken Hatcheries in Henan, China. Front. Cell. Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Bichiou, S. Prévalence des Salmonelles Zoonotiques Dans Les Troupeaux de Reproducteurs de L’espèce Gallus Gallus et Dans Les Couvoirs du Gouvernorat de Nabeul. Ph.D. Thesis, National School of Veterinary Medicine of Sidi Thabet, University Manouba, Manouba, Tunisia, 2010. [Google Scholar]
- GIPAC Portail de L’aviculture en Tunisie: DATA-GIPAC. Available online: http://www.dataportal.gipac.tn/GipaWeb/FreeProduction.aspx (accessed on 20 March 2021).
- Thrusfield, M. Veterinary Epidemiology, 4th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2018. [Google Scholar]
- Code of Practice for the Welfare of Meat Chickens and Meat Breeding Chickens; Department of Environment Food and Rural Affairs: London, UK, 2018.
- ISO NF EN ISO 6579. Microbiologie de la chaîne Alimentaire—Méthode horizontale Pour la Recherche, le Dénombrement et le Sérotypage des Salmonella—Partie 1: Recherche des Salmonella spp.—Microbiologie de la chaîne Alimentaire; AFNOR: Paris, France, 2017. [Google Scholar]
- Popoff, M.Y.; Bockemühl, J.; Gheesling, L.L. Supplement 2001 (no. 45) to the Kauffmann-White scheme. Res. Microbiol. 2003, 154, 173–174. [Google Scholar] [CrossRef]
- Karraouan, B.; Fassouane, A.; El Ossmani, H.; Cohen, N.; Charafeddine, O.; Bouchrif, B. Prevalence and virulence genes of Salmonella in raw minced meat from turkey in Casablanca, Morocco. Rev. Med. Vet. (Toulouse) 2010, 161, 127–132. [Google Scholar]
- Raffatellu, M.; Wilson, R.P.; Chessa, D.; Andrews-Polymenis, H.; Tran, Q.T.; Lawhon, S.; Khare, S.; Adams, L.G.; Bäumler, A.J. SipA, SopA, SopB, SopD, and SopE2 contribute to Salmonella enterica serotype typhimurium invasion of epithelial cells. Infect. Immun. 2005, 73, 146–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Z.; Carter, B.; Núñez-García, J.; AbuOun, M.; Fookes, M.; Ivens, A.; Woodward, M.J.; Anjum, M.F. Identification of genetic and phenotypic differences associated with prevalent and non-prevalent Salmonella Enteritidis phage types: Analysis of variation in amino acid transport. Microbiology 2009, 155, 3200–3213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, D.H.; Zhou, X.; Addwebi, T.; Davis, M.A.; Orfe, L.; Call, D.R.; Guard, J.; Besser, T.E. Cell invasion of poultry-associated salmonella enterica serovar enteritidis isolates is associated with pathogenicity, motility and proteins secreted by the type III secretion system. Microbiology 2011, 157, 1428–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oueslati, W.; Rjeibi, M.R.; Mhadhbi, M.; Jbeli, M.; Zrelli, S.; Ettriqui, A. Prevalence, virulence and antibiotic susceptibility of Salmonella spp. strains, isolated from beef in Greater Tunis (Tunisia). Meat Sci. 2016, 119, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Huehn, S.; La Ragione, R.M.; Anjum, M.; Saunders, M.; Woodward, M.J.; Bunge, C.; Helmuth, R.; Hauser, E.; Guerra, B.; Beutlich, J.; et al. Virulotyping and antimicrobial resistance typing of salmonella enterica serovars relevant to human health in Europe. Foodborne Pathog. Dis. 2010, 7, 523–535. [Google Scholar] [CrossRef]
- Malorny, B.; Hoorfar, J.; Bunge, C.; Helmuth, R. Multicenter validation of the analytical accuracy of salmonella PCR: Towards an international standard. Appl. Environ. Microbiol. 2003, 69, 290–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouzeed, Y.M.; Hariharan, H.; Poppe, C.; Kibenge, F.S.B. Characterization of Salmonella isolates from beef cattle, broiler chickens and human sources on prince Edward Island. Comp. Immunol. Microbiol. Infect. Dis. 2000, 23, 253–266. [Google Scholar] [CrossRef] [Green Version]
- CASFM/EUCAST 2019-Antibiogram Committee of the French Society for Microbiology—Veterinary Antibiograms: Recommandations 2019; Société Française de Microbiologie: Paris, France, 2019.
- Jouy, E.; Haenni, M.; Le Devendec, L.; Le Roux, A.; Châtre, P.; Madec, J.Y.; Kempf, I. Improvement in routine detection of colistin resistance in E. coli isolated in veterinary diagnostic laboratories. J. Microbiol. Methods 2017, 132, 125–127. [Google Scholar] [CrossRef]
- Colom, K.; Pérez, J.; Alonso, R.; Fernández-Aranguiz, A.; Lariño, E.; Cisterna, R. Simple and reliable multiplex PCR assay for detection of blaTEM, blaSHV and blaOXA-1 genes in Enterobacteriaceae. FEMS Microbiol. Lett. 2003, 223, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Mulvey, M.R.; Soule, G.; Boyd, D.; Demczuk, W.; Ahmed, R. Characterization of the first extended-spectrum beta-lactamase-producing Salmonella isolate identified in Canada. J. Clin. Microbiol. 2003, 41, 460–462. [Google Scholar] [CrossRef] [Green Version]
- Robicsek, A.; Strahilevitz, J.; Jacoby, G.A.; Macielag, M.; Abbanat, D.; Chi, H.P.; Bush, K.; Hooper, D.C. Fluoroquinolone-modifying enzyme: A new adaptation of a common aminoglycoside acetyltransferase. Nat. Med. 2006, 12, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, Y.; Wilharm, G.; Zander, E.; Wichelhaus, T.A.; Göttig, S.; Hunfeld, K.-P.; Seifert, H.; Witte, W.; Higgins, P.G. Molecular characterization of blaNDM-1 in an Acinetobacter baumannii strain isolated in Germany in 2007. J. Antimicrob. Chemother. 2011, 66, 1998–2001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, L.K.; Martin, I.; Alfa, M.; Mulvey, M. Multiplex PCR for the detection of tetracycline resistant genes. Mol. Cell. Probes 2001, 15, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Torkan, S. Bahadoranian, M.A.; Khamesipour, F.; Anyanwu, M.U. Detection of Virulence and Antimicrobial Resistance Genes in Escherichia coli Isolates from Diarrhoiec Dogs in Iran Deteccion de Virulencia Y Genes de Resistencia Antimicrobiana en Aislados de Escherichia Coli Provenientes de Perros en Irán. Arch. Méd. Vétérinaire 2016, 48, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Rebelo, A.R.; Bortolaia, V.; Kjeldgaard, J.S.; Pedersen, S.K.; Leekitcharoenphon, P.; Hansen, I.M.; Guerra, B.; Malorny, B.; Borowiak, M.; Hammerl, J.A.; et al. Multiplex PCR for detection of plasmid-mediated colistin resistance determinants, mcr-1, mcr-2, mcr-3, mcr-4 and mcr-5 for surveillance purposes. Eurosurveillance 2018, 23, 17–00672. [Google Scholar] [CrossRef] [PubMed]
- Mazel, D.; Dychinco, B.; Webb, V.A.; Davies, J. Antibiotic resistance in the ECOR collection: Integrons and identification of a novel aad gene. Antimicrob. Agents Chemother. 2000, 44, 1568–1574. [Google Scholar] [CrossRef] [Green Version]
- Hosmer, D.W.; Lemeshow, S. Goodness of fit tests for the multiple logistic regression model. Commun. Stat. Theory Methods 1980, 9, 1043–1069. [Google Scholar] [CrossRef]
- Vodovar, D.; Marcadé, G.; Raskine, L.; Malissin, I.; Mégarbane, B. Entérobactéries productrices de bêta-lactamases à spectre élargi: Épidémiologie, facteurs de risque et mesures de prévention. Rev. Med. Interne 2013, 34, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Ghafourian, S.; Sadeghifard, N.; Soheili, S.; Sekawi, Z. Extended spectrum beta-lactamases: Definition, classification and epidemiology. Curr. Issues Mol. Biol. 2014, 17, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Drieux, L.; Brossier, F.; Sougakoff, W.; Jarlier, V. Phenotypic detection of extended-spectrum β-lactamase production in Enterobacteriaceae: Review and bench guide. Clin. Microbiol. Infect. 2008, 14, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Cui, M.; Xie, M.; Qu, Z.; Zhao, S.; Wang, J.; Wang, Y.; He, T.; Wang, H.; Zuo, Z.; Wu, C. Prevalence and antimicrobial resistance of Salmonella isolated from an integrated broiler chicken supply chain in Qingdao, China. Food Control 2016, 62, 270–276. [Google Scholar] [CrossRef]
- Sivaramalingam, T.; McEwen, S.A.; Pearl, D.L.; Ojkic, D.; Guerin, M.T. A temporal study of Salmonella serovars from environmental samples from poultry breeder flocks in Ontario between 1998 and 2008. Can. J. Vet. Res. 2013, 77, 1–11. [Google Scholar] [PubMed]
- Choi, S.W.; Ha, J.S.; Kim, B.Y.; Lee, D.H.; Park, J.K.; Youn, H.N.; Hong, Y.H.; Lee, S.B.; Lee, J.B.; Park, S.Y.; et al. Prevalence and characterization of Salmonella species in entire steps of a single integrated broiler supply chain in Korea. Poult. Sci. 2014, 93, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Lay, K.S.; Vuthy, Y.; Song, P.; Phol, K.; Sarthou, J.L. Prevalence, numbers and antimicrobial susceptibilities of Salmonella serovars and Campylobacter spp. in retail poultry in Phnom Penh, Cambodia. J. Vet. Med. Sci. 2011, 73, 325–329. [Google Scholar] [CrossRef] [Green Version]
- Thai, T.H.; Yamaguchi, R. Molecular characterization of antibiotic-resistant Salmonella isolates from retail meat from markets in Northern Vietnam. J. Food Prot. 2012, 75, 1709–1714. [Google Scholar] [CrossRef]
- El Allaoui, A.; Rhazi Filali, F.; Ameur, N.; Bouchrif, B. Contamination of broiler turkey farms by Salmonella spp. in Morocco: Prevalence, antimicrobial resistance and associated risk factors. Rev. Sci. Tech. 2017, 36, 935–946. [Google Scholar] [CrossRef]
- Heyndrickx, M.; Vandekerchove, D.; Herman, L.; Rollier, I.; Grijspeerdt, K.; De Zutter, L. Routes for salmonella contamination of poultry meat: Epidemiological study from hatchery to slaughterhouse. Epidemiol. Infect. 2002, 129, 253–265. [Google Scholar] [CrossRef]
- Line, J.E. Campylobacter and Salmonella populations associated with chickens raised on acidified litter. Poult. Sci. 2002, 81, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Cardinale, E.; Tall, F.; Guèye, E.F.; Cisse, M.; Salvat, G. Risk factors for Salmonella enterica subsp. enterica infection in senegalese broiler-chicken flocks. Prev. Vet. Med. 2004, 63, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Aragaw, K.; Molla, B.; Muckle, A.; Cole, L.; Wilkie, E.; Poppe, C.; Kleer, J.; Hildebrandt, G. The characterization of Salmonella serovars isolated from apparently healthy slaughtered pigs at Addis Ababa abattoir, Ethiopia. Prev. Vet. Med. 2007, 82, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Collard, J.-M.; Place, S.; Denis, O.; Rodriguez-Villalobos, H.; Vrints, M.; Weill, F.-X.; Baucheron, S.; Cloeckaert, A.; Struelens, M.; Bertrand, S. Travel-acquired salmonellosis due to Salmonella Kentucky resistant to ciprofloxacin, ceftriaxone and co-trimoxazole and associated with treatment failure. J. Antimicrob. Chemother. 2007, 60, 190–192. [Google Scholar] [CrossRef] [Green Version]
- Molla, B.; Berhanu, A.; Muckle, A.; Cole, L.; Wilkie, E.; Kleer, J.; Hildebrandt, G. Multidrug resistance and distribution of Salmonella serovars in slaughtered pigs. J. Vet. Med. Ser. B Infect. Dis. Vet. Public Health 2006, 53, 28–33. [Google Scholar] [CrossRef]
- Lapierre, L.; Cornejo, J.; Zavala, S.; Galarce, N.; Sánchez, F.; Benavides, M.B.; Guzmán, M.; Sáenz, L. Phenotypic and genotypic characterization of virulence factors and susceptibility to antibiotics in salmonella infantis strains isolated from chicken meat: First findings in Chile. Animals 2020, 10, 1049. [Google Scholar] [CrossRef]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15. [Google Scholar] [CrossRef] [Green Version]
- Hassena, A.B.; Siala, M.; Guermazi, S.; Zormati, S.; Gdoura, R.; Sellami, H. Occurrence and phenotypic and molecular characterization of antimicrobial resistance of salmonella isolates from food in Tunisia. J. Food Prot. 2019, 82, 1166–1175. [Google Scholar] [CrossRef]
- WHO. Global Report on Surveillance 2014. In WHO 2014 AMR Report; WHO Press: Geneve, Switzerland, 2014; pp. 1–8. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
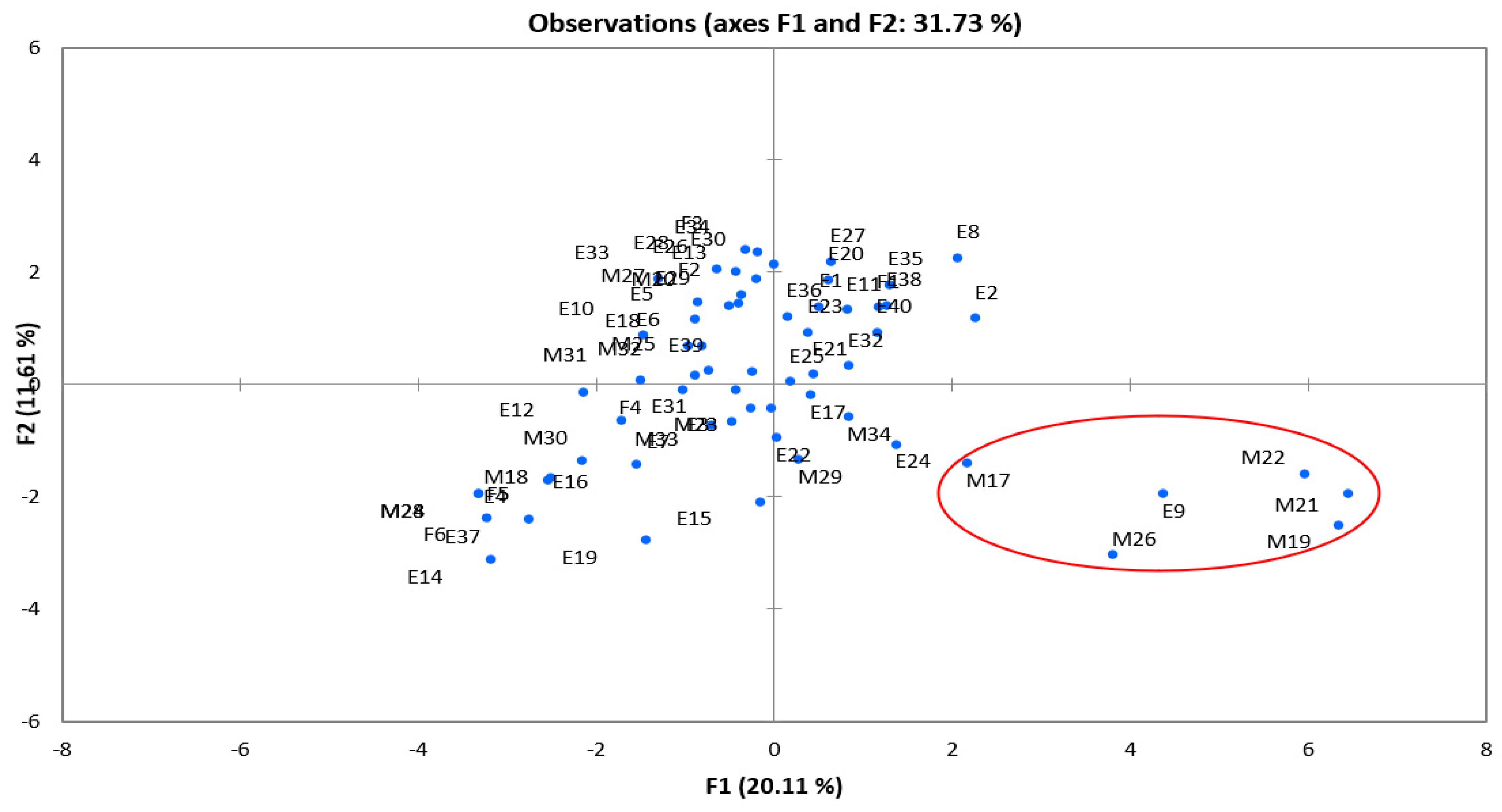{kind=link}
| Gene | Function | Primer Sequence (5′ to 3′) | Product Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|---|
| SEN1417 | Intracellular survival | F: GATCGCTGGCTGGTC | 670 | 58 | [22] |
| R: CTGACCGTAATGGCGA | |||||
| sipA | Host cell invasion | F: ATGGTTACAAGTGTAAGGACTCAG | 2055 | 53 | [23] |
| R: ACGCTGCATGTGCAAGCCATC | |||||
| sipD | Host cell invasion | F: ATGCTTAATATTCAAAATTATTCCG | 1029 | 53 | [23] |
| R: TCCTTGCAGGAAGCTTTTG | |||||
| sopD | Host cell invasion | F: GAGCTCACGACCATTTGCGGCG | 1291 | 59 | [21] |
| R: GAGCTCCGAGACACGCTTCTTCG | |||||
| gipA | Growth or survival in a Peyer’s patch | F: ACGACTGAGCAGGCTGAG | 518 | 58 | [25] |
| R: TTGGAAATGGTGACGGTAGAC | |||||
| mgtC | Intracellular survival | F: TGACTATCAATGCTCCAGTGAAT | 677 | 58 | [25] |
| R: ATTTACTGGCCGCTATGCTGTTG | |||||
| trhH | Code for the putative F pilus assembly protein | F: AACTGGTGCCGTTGTCATTG | 418 | 53 | [25] |
| R: GATGGTCTGTGCTTGCTGAG | |||||
| spvC | Multiplication in host cell | F: CTCCTTGCACAACCAAATGCG | 570 | 53 | [25] |
| R: TGTCTCTGCATTTCACCACCATC | |||||
| sirA | Control enteropathogenic virulence functions | F: TGCGCCTGGTGACAAAACTG | 313 | 55 | [25] |
| R: ACTGACTTCCCAGGCTACAGCA | |||||
| pagK | Biofilm formation | F: ACCATCTTCACTATATTCTGCTC | 151 | 60 | [25] |
| R: ACCTCTACACATTTTAAACCAATC | |||||
| invA | Host cell invasion | F: GTGAAATTATCGCCACGTTCGGGCAA | 284 | 64 | [26] |
| R: TCATCGCACCGTCAAAGGAACC | |||||
| Hli | Control of phase change and motility | F: AGCCTCGGCTACTGGTCTTG | 173 | 55 | [27] |
| R: CCGCAGCAAGAGTCACCTCA |
| Gene | Primer Sequence (5′ to 3′) | Product Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| blaTEM | F: ATCAGCAATAAACCAGC | 516 | 54 | [30] |
| R: CCCCGAAGAACGTTTTC | ||||
| blaCTX-M | F: ATGTGCAGYACCAGTAARGTKATGGC | 592 | 58 | [31] |
| R: TGGGTRAARTARGTSACCAGAAYSAGCGG | ||||
| blaNDM1 | F: CTGAGCACCGCATTAGCC | 621 | 52 | [33] |
| R: GGGCCGTATGAGTGATTGC | ||||
| tetA | F: GGTTCACTCGAACGACGTCA | 577 | 55 | [34] |
| R: CTGTCCGACAAGTTGCATGA | ||||
| tetB | F: CCTCAGCTTCTCAACGCGTG | 634 | 55 | [34] |
| R: GCACCTTGCTGATGACTCTT | ||||
| dfrA1 | F: GGAGTGCCAAAGGTGAACAGC | 367 | 55 | [35] |
| R: GAGGCGAAGTCTTGGGTAAAAAC | ||||
| qnrB | F: GATCGTGAAAGCCAGAAAGG | 469 | 53 | [32] |
| R: ACGATGCCTGGTAGTTGTCC | ||||
| mcr-1 | F: AGTCCGTTTGTTCTTGTGGC | 320 | 58 | [36] |
| R: AGATCCTTGGTCTCGGCTTG | ||||
| mcr-2 | F: CAAGTGTGTTGGTCGCAGTT | 715 | 58 | [36] |
| R: TCTAGCCCGACAAGCATACC | ||||
| mcr-3 | F: AAATAAAAATTGTTCCGCTTATG | 929 | 58 | [36] |
| R: AATGGAGATCCCCGTTTTT | ||||
| mcr-4 | F: TCACTTTCATCACTGCGTTG | 1116 | 58 | [36] |
| R: TTGGTCCATGACTACCAATG | ||||
| mcr-5 | F: ATGCGGTTGTCTGCATTTATC | 1644 | 58 | [36] |
| R: TCATTGTGGTTGTCCTTTTCTG | ||||
| int1 | F: GGGTCAAGGATCTGGATTTCG | 483 | 62 | [37] |
| R: ACATGGGTGTAAATCATCGTC | ||||
| int2 | F: CACGGATATGCGACAAAAAGGT | 233 | 62 | [37] |
| R: GTAGCAAACGAGTGACGAAATG |
| Risk Factor | Category | Prevalence in % (Positive/Tested) | OR [95% CI] | p-Value | Multivariate Logistic Regression OR [95% CI] |
|---|---|---|---|---|---|
| No cleaning and disinfection around the breeding unit | Yes | 29.4 (40/136) | 2.810 [1.600–4.950] | <0.001 | 8.642 [1.770–42.196] |
| No | 12.9 (24/186) | ||||
| Absence of treatment with an antimicrobial at the start | Yes | 33.3 (52/156) | 6.420 [3.270–12.610] | <0.001 | 4.675 [1.720–12.703] |
| No | 7.2 (12/166) | ||||
| Duration of crawl space < 15 days | Yes | 32.7 (53/162) | 6.590 [3.290–13.200] | <0.001 | 3.562 [1.436–8.835] |
| No | 6.9 (11/160) | ||||
| Wet litter | Yes | 27.8 (50/180) | 3.520 [1.850–6.680] | <0.001 | |
| No | 9.9 (14/142) | ||||
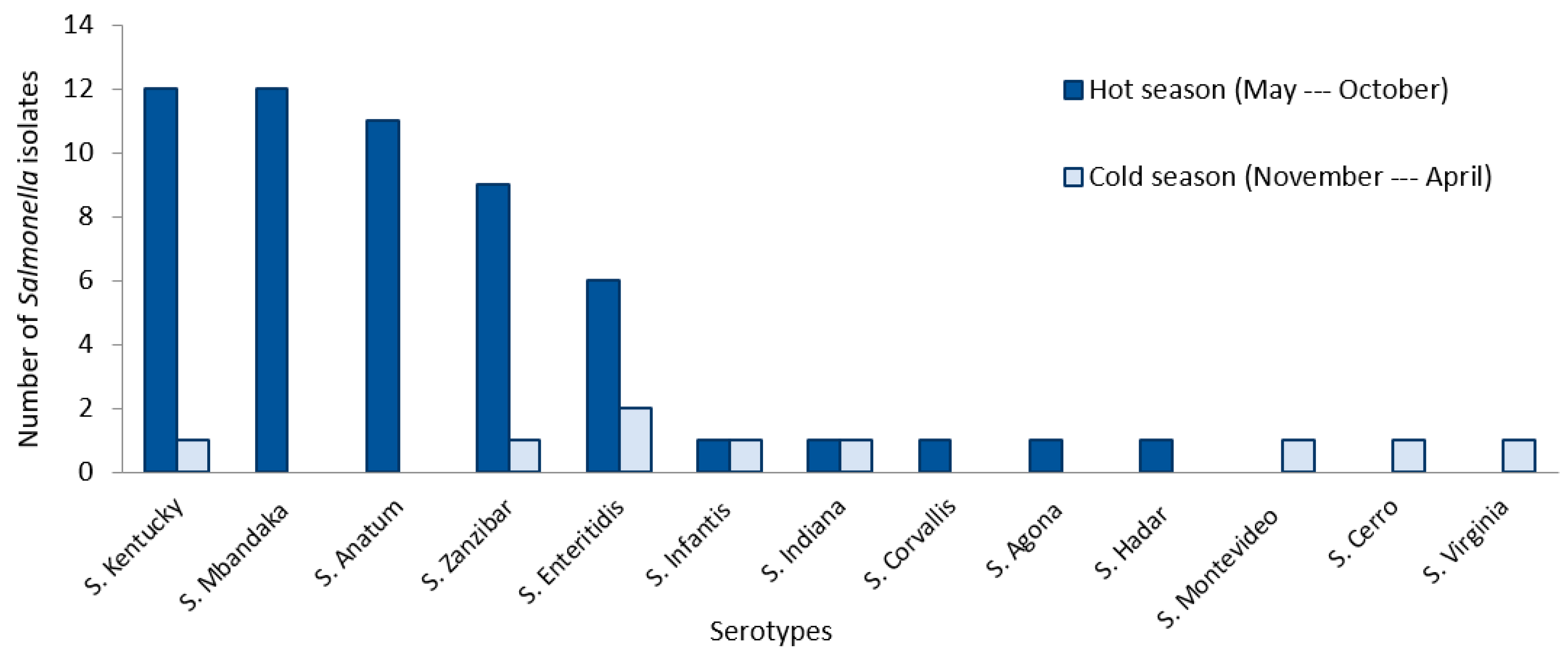
| Hot season (T ≥ 20 °C) * | Yes | 24.6 (55/224) | 3.218 [1.520–6.813] | 0.001 | |
| No | 9.2 (9/98) | ||||
| Number of chicks at setting in place > 25/m2 | Yes | 26.6 (47/177) | 2.720 [1.480–4.990] | 0.009 | |
| No | 11.7 (17/145) | ||||
| Absence of rodent control in the building | Yes | 17.6 (42/238) | 0.604 [0.335–1.089] | 0.092 | |
| No | 26.2 (22/84) | ||||
| Poor state of cleanliness of poultry | Yes | 18.7 (45/241) | 0.749 [0.408–1.375] | 0.351 | |
| No | 23.5 (19/81) |
| Serotypes Prevalence in % (Positive/Tested) | Strains | Virulence Genes (a) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| invA | spvC | hli | gipA | mgtC | trhH | sirA | pagK | sipA | sipD | sopD | SEN | ||
| S. Kentucky 20.3% (13/64) | E2 | + | − | − | − | + | − | + | + | − | − | − | − |
| E8 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E11 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E17 | + | − | − | − | + | − | + | + | − | − | − | − | |
| E22 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E24 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E25 | + | − | − | − | + | − | + | + | − | − | − | − | |
| E31 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E36 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E38 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E40 | + | − | + | − | + | − | + | + | − | − | − | − | |
| F1 | + | − | + | + | + | − | + | + | − | − | − | − | |
| F4 | + | − | + | − | + | − | + | + | − | − | − | − | |
| S. Mbandaka 18.7% (12/64) | E12 | + | − | − | + | + | − | + | + | − | − | − | − |
| E16 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E18 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E20 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E23 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E32 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E39 | + | − | − | + | + | − | + | + | − | − | − | − | |
| M23 | + | − | − | − | + | − | + | + | − | − | − | − | |
| M25 | + | − | − | + | + | − | + | + | − | − | − | − | |
| M29 | + | − | + | − | + | − | + | + | − | − | − | − | |
| M34 | + | − | + | + | + | − | + | + | − | − | − | − | |
| F6 | + | − | − | + | + | − | + | + | − | − | − | − | |
| S. Anatum 17.1% (11/64) | M17 | + | − | − | + | + | − | + | + | − | − | − | − |
| M19 | + | − | − | + | + | − | + | + | − | − | − | − | |
| M21 | + | − | − | + | + | − | + | + | − | − | − | − | |
| M22 | + | − | + | + | + | − | + | + | − | − | − | − | |
| M24 | + | − | + | + | + | − | + | + | − | − | − | − | |
| M26 | + | − | − | − | + | − | + | + | − | − | − | − | |
| M28 | + | − | + | + | + | − | + | + | − | − | − | − | |
| M30 | + | − | + | + | + | − | + | + | − | − | − | − | |
| M31 | + | − | − | + | + | − | + | + | − | − | − | − | |
| M32 | + | − | − | − | + | − | + | + | − | − | − | − | |
| M33 | + | − | − | − | + | − | + | + | − | − | − | − | |
| S. Zanzibar 15.6% (10/64) | E3 | + | − | − | + | + | − | + | + | − | − | − | − |
| E5 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E6 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E7 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E26 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E28 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E29 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E34 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E35 | + | − | − | + | + | − | + | + | − | − | − | − | |
| F3 | + | − | − | + | + | − | + | + | − | − | − | − | |
| S. Enteritidis 12.5% (8/64) | E1 | + | − | − | + | + | − | + | + | − | − | − | − |
| E13 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E14 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E15 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E21 | + | − | + | + | + | − | + | + | − | − | − | − | |
| E27 | + | − | − | − | + | − | + | + | − | − | − | − | |
| E30 | + | − | − | + | + | − | + | + | − | − | − | − | |
| E33 | + | − | − | + | + | − | + | + | − | − | − | − | |
| S. Infantis 3.1% (2/64) | M27 | + | − | − | + | + | − | + | + | − | − | − | − |
| F2 | + | − | − | + | + | − | + | + | − | − | − | − | |
| S. Indiana 3.1% (2/64) | E10 | + | − | − | + | + | − | + | + | − | − | − | − |
| M20 | + | − | + | + | + | − | + | + | − | − | − | − | |
| S. Corvallis 1.6% (1/64) | E9 | + | − | − | + | + | − | + | + | − | − | − | − |
| S. Agona 1.6% (1/64) | E4 | + | − | − | + | + | − | + | + | − | − | − | − |
| S. Hadar 1.6% (1/64) | E19 | + | − | + | + | + | − | + | + | − | − | − | − |
| S. Montevideo 1.6% (1/64) | M18 | + | − | − | − | + | − | + | + | − | − | − | − |
| S. Cerro 1.6% (1/64) | F5 | + | − | + | + | + | − | + | + | − | − | − | − |
| S. Virginia 1.6% (1/64) | E37 | + | − | − | + | + | − | + | + | − | − | − | − |
| Serotypes Prevalence in % (Positive/Tested) | Strains | Antimicrobial Resistance Profiles (b) | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMX | AUC | SF | FOX | CAZ | CTX | CRO | CPM | ATM | ETP | GME | S | CS50 | NA | ENF | CIP | FFC | C | TE | SUL | SXT | ||
| S. Kentucky 20.3% (13/64) | E2 (c) | |||||||||||||||||||||
| E8 (c) | ||||||||||||||||||||||
| E11 (c) | ||||||||||||||||||||||
| E17 (c) | ||||||||||||||||||||||
| E22 (c) | ||||||||||||||||||||||
| E24 (c) | ||||||||||||||||||||||
| E25 (c) | ||||||||||||||||||||||
| E31 (c) | ||||||||||||||||||||||
| E36 (c) | ||||||||||||||||||||||
| E38 (c) | ||||||||||||||||||||||
| E40 (c) | ||||||||||||||||||||||
| F1 (c) | ||||||||||||||||||||||
| F4 (c) | ||||||||||||||||||||||
| S. Mbandaka 18.7% (12/64) | E12 (c) | |||||||||||||||||||||
| E16 (c) | ||||||||||||||||||||||
| E18 (c) | ||||||||||||||||||||||
| E20 (c) | ||||||||||||||||||||||
| E23 (c) | ||||||||||||||||||||||
| E32 (c) | ||||||||||||||||||||||
| E39 (c) | ||||||||||||||||||||||
| M23 (c) | ||||||||||||||||||||||
| M25 (c) | ||||||||||||||||||||||
| M29 (c) | ||||||||||||||||||||||
| M34 (c) | ||||||||||||||||||||||
| F6 (f) | ||||||||||||||||||||||
| S. Anatum 17.1% (11/64) | M17 (d) | |||||||||||||||||||||
| M19 (e) | ||||||||||||||||||||||
| M21 (e) | ||||||||||||||||||||||
| M22 (d) | ||||||||||||||||||||||
| M24 (f) | ||||||||||||||||||||||
| M26 (d) | ||||||||||||||||||||||
| M28 (f) | ||||||||||||||||||||||
| M30 (c) | ||||||||||||||||||||||
| M31 (c) | ||||||||||||||||||||||
| M32 (c) | ||||||||||||||||||||||
| M33 (c) | ||||||||||||||||||||||
| S. Zanzibar 15.6% (10/64) | E3 (c) | |||||||||||||||||||||
| E5 (c) | ||||||||||||||||||||||
| E6 (c) | ||||||||||||||||||||||
| E7 (c) | ||||||||||||||||||||||
| E26 (c) | ||||||||||||||||||||||
| E28 (c) | ||||||||||||||||||||||
| E29 (c) | ||||||||||||||||||||||
| E34 (c) | ||||||||||||||||||||||
| E35 (c) | ||||||||||||||||||||||
| F3 (c) | ||||||||||||||||||||||
| S. Enteritidis 12.5% (8/64) | E1 (c) | |||||||||||||||||||||
| E13 (c) | ||||||||||||||||||||||
| E14 (f) | ||||||||||||||||||||||
| E15 (c) | ||||||||||||||||||||||
| E21 (c) | ||||||||||||||||||||||
| E27 (c) | ||||||||||||||||||||||
| E30 (c) | ||||||||||||||||||||||
| E33 (c) | ||||||||||||||||||||||
| S. Infantis 3.1% (2/64) | M27 (c) | |||||||||||||||||||||
| F2 (c) | ||||||||||||||||||||||
| S. Indiana 3.1% (2/64) | E10 (c) | |||||||||||||||||||||
| M20 (c) | ||||||||||||||||||||||
| S. Corvallis 1.6% (1/64) | E9 (e) | |||||||||||||||||||||
| S. Agona 1.6% (1/64) | E4 (f) | |||||||||||||||||||||
| S. Hadar 1.6% (1/64) | E19 (c) | |||||||||||||||||||||
| S. Montevideo 1.6% (1/64) | M18 (f) | |||||||||||||||||||||
| S. Cerro 1.6% (1/64) | F5 (f) | |||||||||||||||||||||
| S. Virginia 1.6% (1/64) | E37 (f) | |||||||||||||||||||||
 (b) Sensitive;
(b) Sensitive;  Resistant. AMX: amoxicillin, AUC: amoxicillin + clavulanic acid, SF: cefalotin, FOX: cefoxitin, CAZ: ceftazidim, CTX: cefotaxim, CRO: ceftriaxon, CPM: cefepim, ATM: aztreonam, ETP: ertapenem, GME: gentamicin, S: streptomycin, CS50: colistin, NA: nalidixic acid, ENF: enrofloxacin, CIP: ciprofloxacin, FFC: florfenicol, C: chloramphenicol, TE: tetracycline, SUL: sulfamides, SXT: trimethoprim-sulfamethoxazole. (c) MDR+ Strain, (d) MDR+ & ESBL+ Strain, (e) MDR+ & AmpC+ Strain, MDR−, (f) ESBL− & AmpC− Strain.
Resistant. AMX: amoxicillin, AUC: amoxicillin + clavulanic acid, SF: cefalotin, FOX: cefoxitin, CAZ: ceftazidim, CTX: cefotaxim, CRO: ceftriaxon, CPM: cefepim, ATM: aztreonam, ETP: ertapenem, GME: gentamicin, S: streptomycin, CS50: colistin, NA: nalidixic acid, ENF: enrofloxacin, CIP: ciprofloxacin, FFC: florfenicol, C: chloramphenicol, TE: tetracycline, SUL: sulfamides, SXT: trimethoprim-sulfamethoxazole. (c) MDR+ Strain, (d) MDR+ & ESBL+ Strain, (e) MDR+ & AmpC+ Strain, MDR−, (f) ESBL− & AmpC− Strain.Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oueslati, W.; Rjeibi, M.R.; Benyedem, H.; Mamlouk, A.; Souissi, F.; Selmi, R.; Ettriqui, A. Prevalence, Risk Factors, Antimicrobial Resistance and Molecular Characterization of Salmonella in Northeast Tunisia Broiler Flocks. Vet. Sci. 2022, 9, 12. https://doi.org/10.3390/vetsci9010012
Oueslati W, Rjeibi MR, Benyedem H, Mamlouk A, Souissi F, Selmi R, Ettriqui A. Prevalence, Risk Factors, Antimicrobial Resistance and Molecular Characterization of Salmonella in Northeast Tunisia Broiler Flocks. Veterinary Sciences. 2022; 9(1):12. https://doi.org/10.3390/vetsci9010012
Chicago/Turabian StyleOueslati, Walid, Mohamed Ridha Rjeibi, Hayet Benyedem, Aymen Mamlouk, Fatma Souissi, Rachid Selmi, and Abdelfettah Ettriqui. 2022. "Prevalence, Risk Factors, Antimicrobial Resistance and Molecular Characterization of Salmonella in Northeast Tunisia Broiler Flocks" Veterinary Sciences 9, no. 1: 12. https://doi.org/10.3390/vetsci9010012
APA StyleOueslati, W., Rjeibi, M. R., Benyedem, H., Mamlouk, A., Souissi, F., Selmi, R., & Ettriqui, A. (2022). Prevalence, Risk Factors, Antimicrobial Resistance and Molecular Characterization of Salmonella in Northeast Tunisia Broiler Flocks. Veterinary Sciences, 9(1), 12. https://doi.org/10.3390/vetsci9010012