
Effect of Vaccination against Glässer’s Disease in a Farm Suffering from Polyserositis in Weaned Pigs

 ,
,  ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
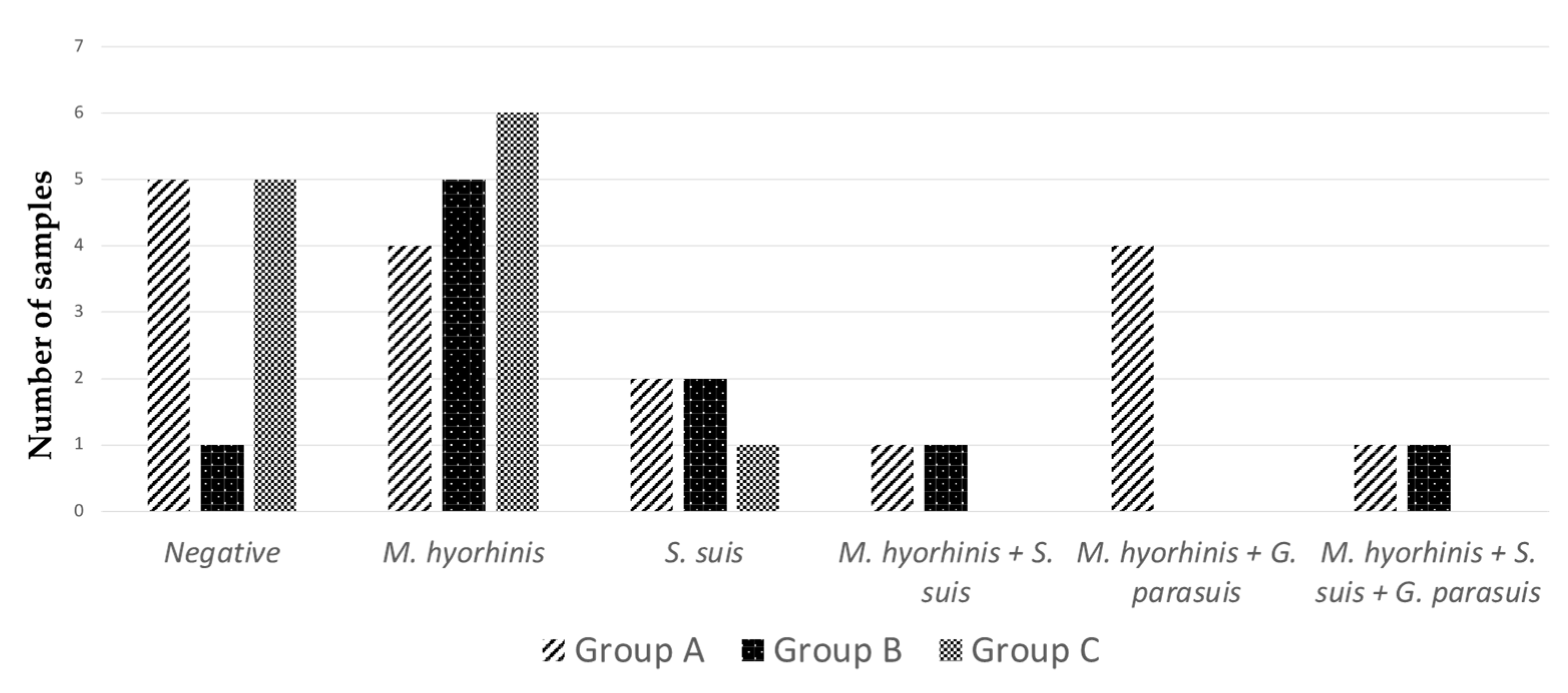
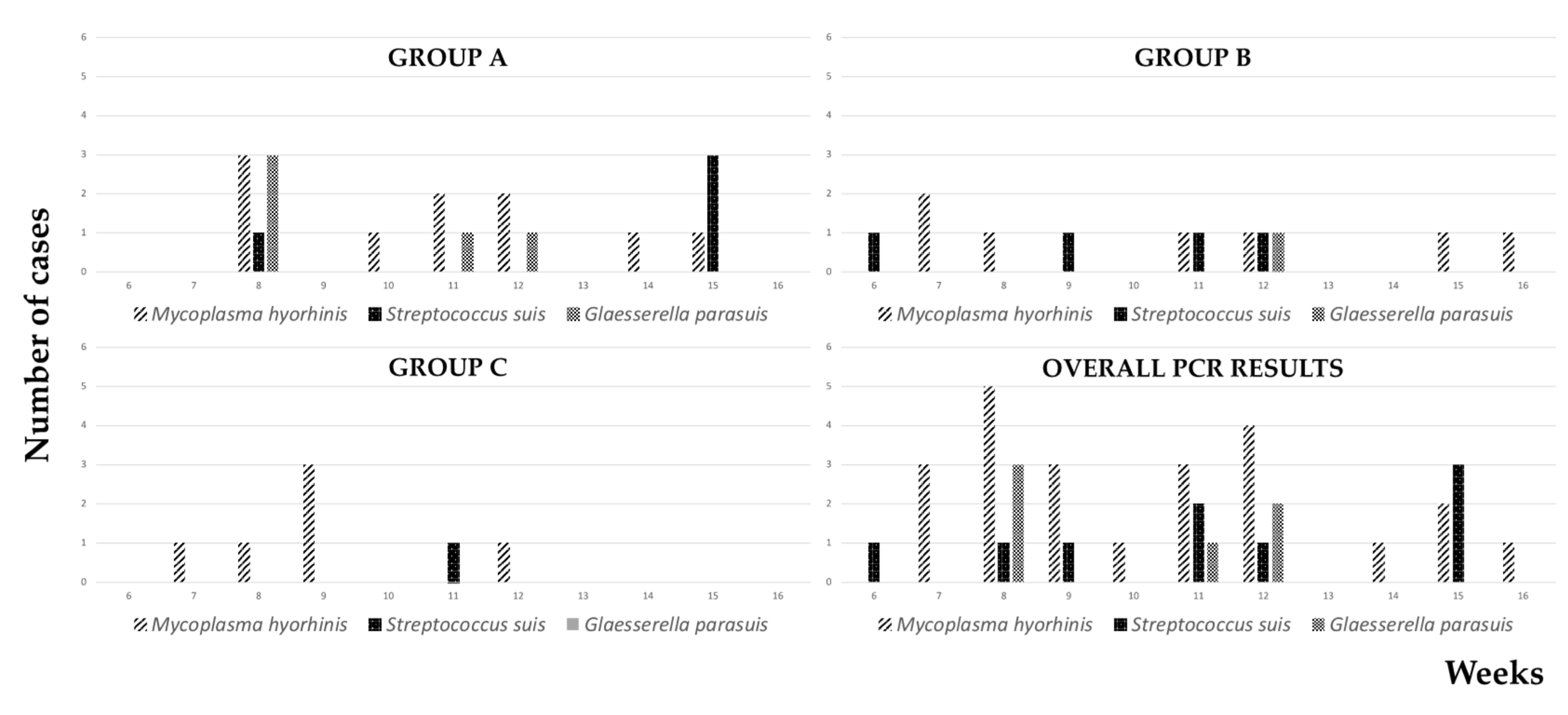
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zachary, J.F.; McGavin, M.D. Pathologic Basis of Veterinary Disease, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 536–537. [Google Scholar]
- Oliveira, S.; Pijoan, C. Haemophilus parasuis: New trends on diagnosis, epidemiology and control. Vet. Microbiol. 2004, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Thacker, H.L.; Bowersock, T.L. Streptococcus suis infection in swine: A retrospective study of 256 cases. Part II. Clinical signs, gross and microscopic lesions, and coexisting microorganisms. J. Vet. Diagn. Investig. 1994, 6, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Kobisch, M.; Friis, N.F. Swine mycoplasmoses. Rev. Sci. Tech. 1996, 15, 1569–1605. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, N.C.; Bille, N.; Riising, H.J.; Dam, A. Polyserositis in pigs due to generalized Escherichia coli infection. Can. J. Comp. Med. 1975, 39, 421–426. [Google Scholar] [PubMed]
- Oliveira Filho, J.X.; Morés, M.A.Z.; Rebellato, R.; Kich, J.D.; Cantão, M.E.; Klein, C.S.; Carvalho Guedes, R.M.; Coldebella, A.; Santos Neves de Barcellos, D.E.; Morés, N. Pathogenic variability among Pasteurella multocida type A isolates from Brazilian pig farms. BMC Vet. Res. 2018, 14, 244. [Google Scholar] [CrossRef] [PubMed]
- Hoeltig, D.; Rohde, J.; Frase, R.; Nietfeld, F.; Waldmann, K.H.; Valentin-Weigand, P.; Means, J. Multi-organ spreading of Actinobacillus pleuropneumoniae serovar 7 in weaned pigs during the first week after experimental infection. Vet. Res. 2018, 49, 97. [Google Scholar] [CrossRef]
- Costa-Hurtado, M.; Barba-Vidal, E.; Maldonado, J.; Aragon, V. Update on Glässer’s disease: How to control the disease under restrictive use of antimicrobials. Vet. Microbiol. 2020, 242, 108595. [Google Scholar] [CrossRef]
- Salogni, C.; Lazzaro, M.; Giovannini, S.; Vitale, N.; Boniotti, M.B.; Pozzi, P.; Pasquali, P.; Alborali, G.L. Causes of swine polyserositis in a high-density breeding area in Italy. J. Vet. Diagn. Investig. 2020, 32, 594–597. [Google Scholar] [CrossRef]
- Oliveira, S.; Galina, L.; Pijoan, C. Development of a PCR test to diagnose Haemophilus parasuis infections. J. Vet. Diagn. Investig. 2001, 13, 495–501. [Google Scholar] [CrossRef]
- Olvera, A.; Pina, S.; Macedo, N.; Oliveira, S.; Aragon, V.; Bensaid, A. Identification of potentially virulent strains of Haemophilus parasuis using a multiplex PCR for virulence-associated autotransporters (vtaA). Vet. J. 2012, 191, 213–218. [Google Scholar] [CrossRef]
- Fourour, S.; Fablet, C.; Tocqueville, V.; Dorenlor, V.; Eono, F.; Eveno, E.; Kempf, I.; Marois-Créhan, C. A new multiplex real-time TaqMan® PCR for quantification of Mycoplasma hyopneumoniae, M. hyorhinis and M. flocculare: Exploratory epidemiological investigations to research mycoplasmal association in enzootic pneumonia-like lesions in slaughtered pigs. J. Appl. Microbiol. 2018, 125, 345–355. [Google Scholar] [CrossRef]
- Okwumabua, O.; O’Connor, M.; Shull, E. A polymerase chain reaction (PCR) assay specific for Streptococcus suis based on the gene encoding the glutamate dehydrogenase. FEMS Microbiol. Lett. 2003, 218, 79–84. [Google Scholar] [CrossRef]
- Howell, K.J.; Peters, S.E.; Wang, J.; Hernandez-Garcia, J.; Weinert, L.A.; Luan, S.L.; Chaudhuri, R.R.; Angen, Ø.; Aragon, V.; Williamson, S.M.; et al. Development of a Multiplex PCR Assay for Rapid Molecular Serotyping of Haemophilus parasuis. J. Clin. Microbiol. 2015, 53, 3812–3821. [Google Scholar] [CrossRef]
- Lee, C.Y.; Ooi, P.T.; Zunita, Z.; Noordin, M.M. Molecular detection of Haemophilus parasuis serotypes 4, 5 or 12 and 13 in Peninsular Malaysia. Trop Biomed. 2019, 36, 482–487. [Google Scholar]
- Johnson, A.K.; Colpoys, J.D.; Edwards-Callaway, L.N.; Calvo-Lorenzo, M.; McGlone, J.J.; Millman, S.T.; Phillips, C.E.; Ritter, M.J.; Sutherland, M.A.; Tucker, A.L.; et al. Behavior and Welfare. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez., A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 17–41. [Google Scholar]
- Bruns, C.E.; Stalder, K.J. Genetics and health. In Diseases of Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 42–49. [Google Scholar]
- Knauer, M.T.; Hostetler, C.E. US swine industry productivity analysis, 2005 to 2010. J. Swine Health Prod. 2013, 21, 248–252. [Google Scholar]
- Day, M.J. Immune system ontogeny and neonatal immunology. In Veterinary Immunology. Principles and Practice, 2nd ed.; Day, M.J., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 213–220. [Google Scholar] [CrossRef]
- Muirhead, M.R.; Alexander, T.J.L. Managing health and disease. In Managing Pig Health, 2nd ed.; Carr, J., Ed.; 5m Publishing: Sheffield, UK, 2013; pp. 73–130. [Google Scholar]
- Pieters, M.G.; Maes, D. Mycoplasmosis. In Diseases of Swine Swine, 11th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Zhang, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 863–883. [Google Scholar] [CrossRef]
- Martinson, B.; Minion, F.C.; Kroll, J.; Hermann, J. Age susceptibility of caesarian derived colostrum deprived pigs to Mycoplasma hyorhinis challenge. Vet. Microbiol. 2017, 210, 147–152. [Google Scholar] [CrossRef]
- Martinson, B.; Zoghby, W.; Barrett, K.; Bryson, L.; Christmas, R.; Minion, F.C.; Kroll, J. Efficacy of an inactivated Mycoplasma hyorhinis vaccine in pigs. Vaccine 2018, 36, 408–412. [Google Scholar] [CrossRef]
- Klein, U.; Földi, D.; Belecz, N.; Hrivnák, V.; Somogyi, Z.; Gastaldelli, M.; Merenda, M.; Catania, S.; Dors, A.; Siesenop, U.; et al. Antimicrobial susceptibility profiles of Mycoplasma hyorhinis strains isolated from five European countries between 2019 and 2021. PLoS ONE 2022, 17, e0272903. [Google Scholar] [CrossRef]
- Liu, H.; Xue, Q.; Zeng, Q.; Zhao, Z. Haemophilus parasuis vaccines. Vet. Immunol. Immunopathol. 2016, 180, 53–58. [Google Scholar] [CrossRef]
- Zhao, Z.; Liu, H.; Xue, Y.; Chen, K.; Liu, Z.; Xue, Q.; Wang, C. Analysis of efficacy obtained with a trivalent inactivated Haemophilus parasuis serovars 4, 5, and 12 vaccine and commercial vaccines against Glässer’s disease in piglets. Can. J. Vet. Res. 2017, 81, 22–27. [Google Scholar]
- Olvera, A.; Ballester, M.; Nofrarías, M.; Sibila, M.; Aragon, V. Differences in phagocytosis susceptibility in Haemophilus parasuis strains. Vet. Res. 2009, 40, 24. [Google Scholar] [CrossRef] [PubMed]
- Luppi, A.; Bonilauri, P.; Dottori, M.; Iodice, G.; Gherpelli, Y.; Merialdi, G.; Maioli, G.; Martelli, P. Haemophilus parasuis serovars isolated from pathological samples in Northern Italy. Transbound. Emerg. Dis. 2013, 60, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wu, J.; Zhang, Y.; Guo, L.; Cong, X.; Du, Y.; Li, J.; Sun, W.; Shi, J.; Peng, J.; et al. Concurrent highly pathogenic porcine reproductive and respiratory syndrome virus infection accelerates Haemophilus parasuis infection in conventional pigs. Vet. Microbiol. 2012, 158, 316–321. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Sows (Number) | Piglets (Number) | ||
|---|---|---|---|---|
| Vaccinated | Not vaccinated | Vaccinated | Not Vaccinated | |
| A | // | 19 | // | 172 |
| B | 12 | // | // | 99 |
| C | 15 | // | 116 | // |
| Primer Name | Primer Sequence (5′-3′) | Amplicon Size (bp) | Targeted Pathogen | Reference |
|---|---|---|---|---|
| Hps-f | GTGATGAGGAAGGGTGRTGT | 822 | Glaesserella parasuis | [10] |
| Hps-r | GGCTTCGTCRCCCTCTGT | |||
| YADAF 1 | TTTAGGTAAAGATAAGCAAGGAAATCC | 406 | Glaesserella parasuis; vtA domain group 1 | [11] |
| PADHR 1 | CCACACAAAACCTACCCCTCCTCC | |||
| YADAF 2 | AGCTTAATATCTCAGCACAAGGTGC | 294 | Glaesserella parasuis; vtA domain group 2 | [11] |
| PADHR 1 | CCACACAAAACCTACCCCTCCTCC | |||
| YADAF 3 | AATGGTAGCCAGTTGTATAATGTTGC | 291 | Glaesserella parasuis; vtA domain group 3 | [11] |
| PADHR 1 | CCACACAAAACCTACCCCTCCTCC | |||
| Mhr_p37 f | TTCTATTTTCATCTATATTTTCGC | 101 | Mycoplasma hyorhinis | [12] |
| Mhr_p37 r | TCATTGACCTTGACTAACTG | |||
| JP4 | GCAGCGTATTCTGTCAAACG | 688 | Streptococcus suis | [13] |
| JP5 | CCATGGACAGATAAAGATGG |
| Group A | Group B | Group C | |
|---|---|---|---|
| Total mortality rate | 15.72 | 22.09 | 24.75 * |
| Polyserositis-specific mortality rate | 10.69 | 11.62 | 11.88 |
| Polyserositis-proportionate mortality | 68.00 | 52.63 | 48.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hattab, J.; Marruchella, G.; Trachtman, A.R.; Gabrielli, L.; Bernabò, N.; Mosca, F.; Tiscar, P.G. Effect of Vaccination against Glässer’s Disease in a Farm Suffering from Polyserositis in Weaned Pigs. Vet. Sci. 2022, 9, 691. https://doi.org/10.3390/vetsci9120691
Hattab J, Marruchella G, Trachtman AR, Gabrielli L, Bernabò N, Mosca F, Tiscar PG. Effect of Vaccination against Glässer’s Disease in a Farm Suffering from Polyserositis in Weaned Pigs. Veterinary Sciences. 2022; 9(12):691. https://doi.org/10.3390/vetsci9120691
Chicago/Turabian StyleHattab, Jasmine, Giuseppe Marruchella, Abigail Rose Trachtman, Luigino Gabrielli, Nicola Bernabò, Francesco Mosca, and Pietro Giorgio Tiscar. 2022. "Effect of Vaccination against Glässer’s Disease in a Farm Suffering from Polyserositis in Weaned Pigs" Veterinary Sciences 9, no. 12: 691. https://doi.org/10.3390/vetsci9120691
APA StyleHattab, J., Marruchella, G., Trachtman, A. R., Gabrielli, L., Bernabò, N., Mosca, F., & Tiscar, P. G. (2022). Effect of Vaccination against Glässer’s Disease in a Farm Suffering from Polyserositis in Weaned Pigs. Veterinary Sciences, 9(12), 691. https://doi.org/10.3390/vetsci9120691