
Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases

 ,
,  , , , ,
, , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bee Sample Processing
2.2. Bacterial Isolation
2.3. Genotypic Analysis
2.4. Screening of Antimicrobial Activity
2.5. Antimicrobial Activity Test
2.6. Biochemical Characterization
2.7. Cell Surface Properties
2.7.1. Bacterial Cultures
2.7.2. Auto-Aggregation
2.7.3. Hydrophobicity
2.7.4. Statistical Analysis
3. Results
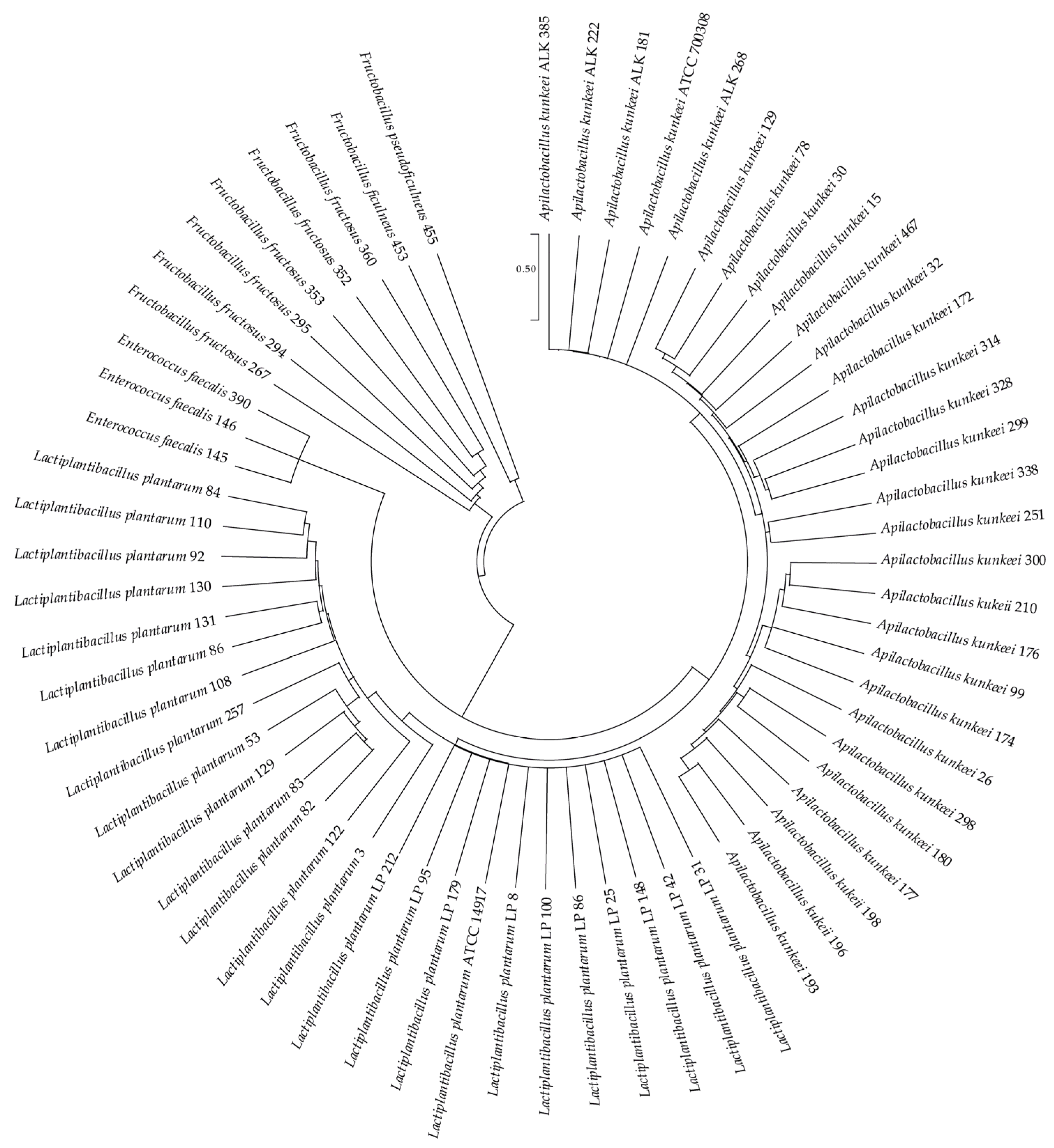
3.1. LAB Species Diversity
3.2. Antimicrobial Activity
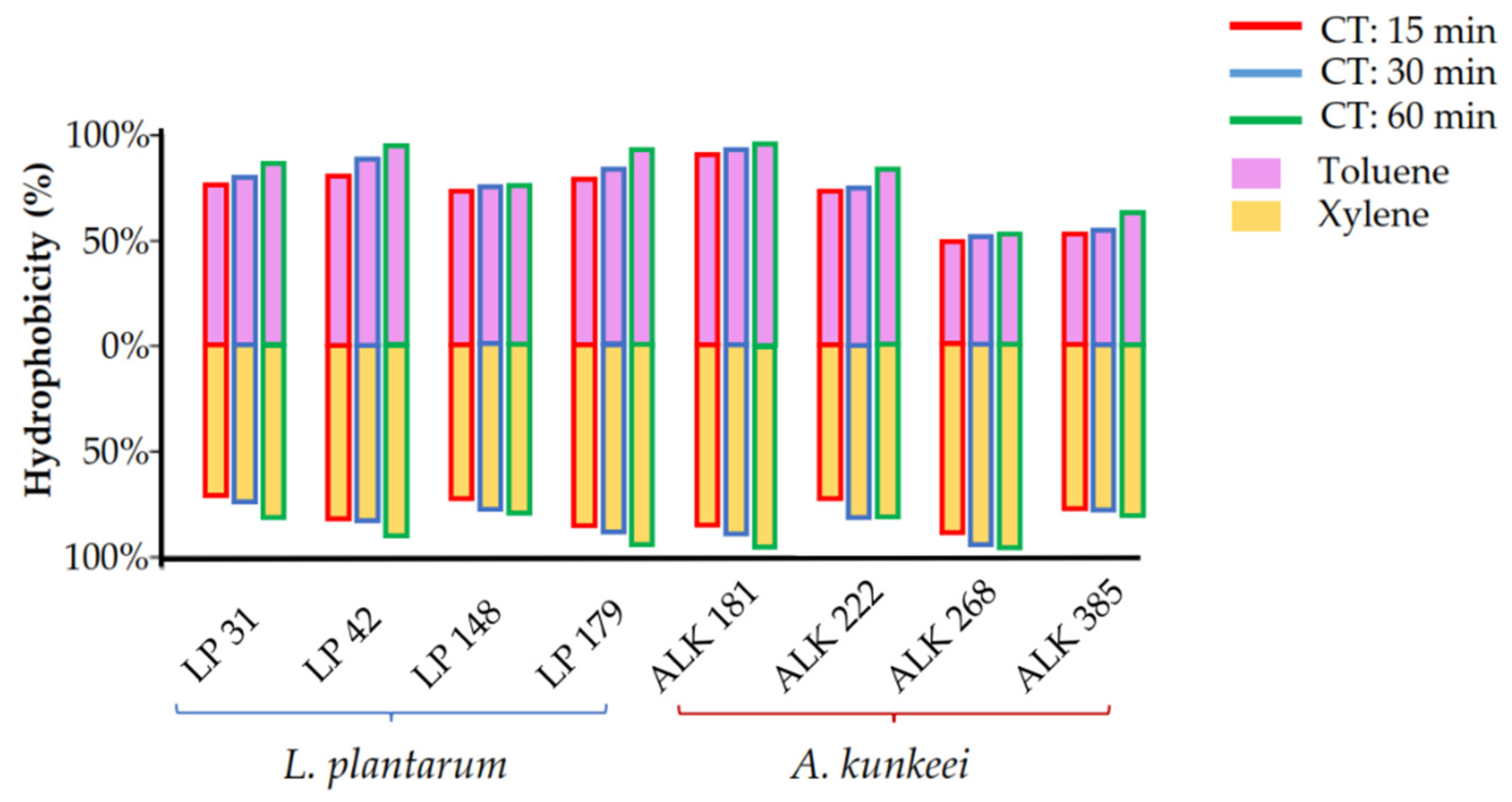
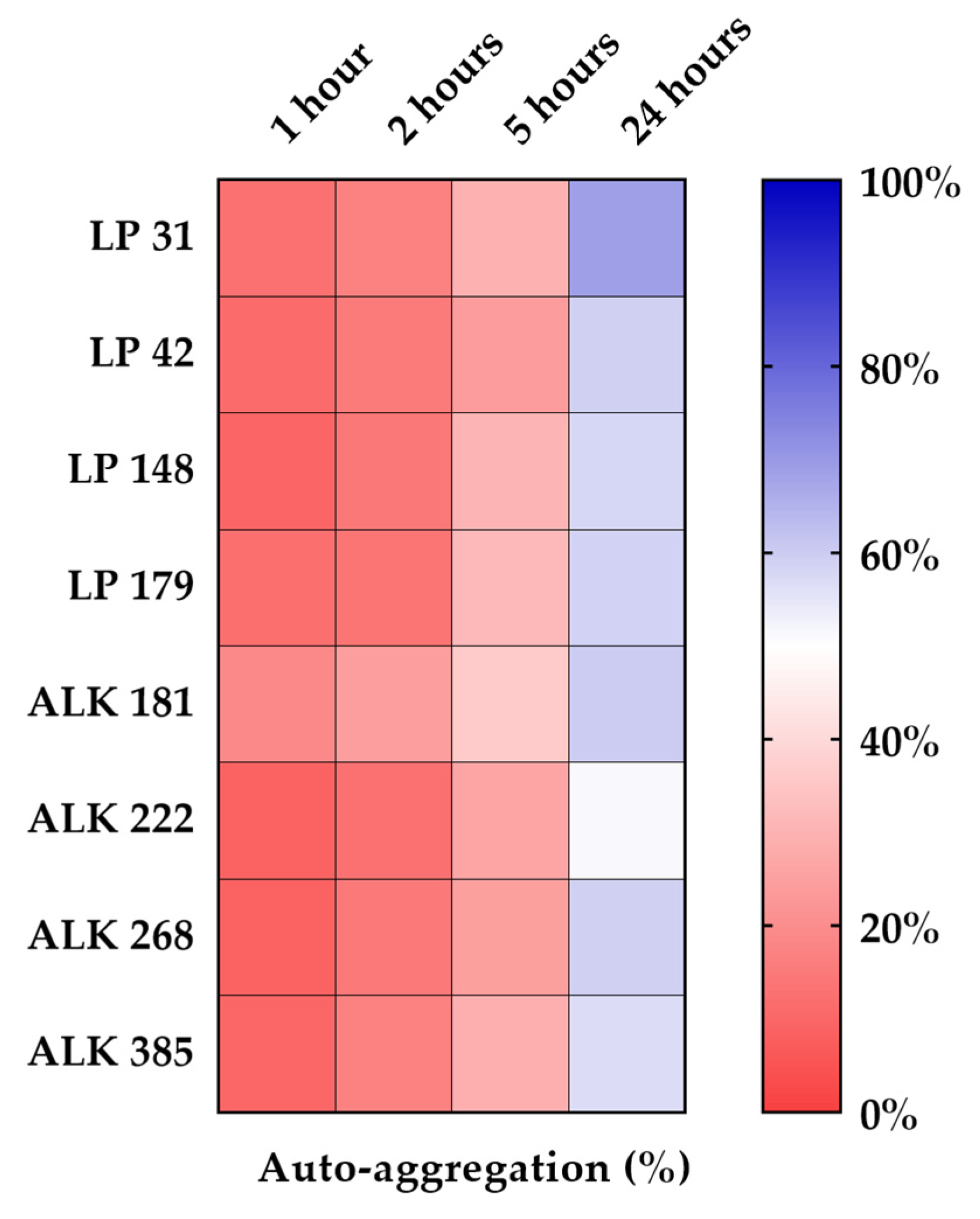
3.3. Hydrophobicity and Auto-Aggregation
3.4. Biochemical Characterization
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EFSA Panel on Plant Protection Products and their Residues (PPR). Scientific Opinion on the Science behind the Development of a Risk Assessment of Plant Protection Products on Bees (Apis mellifera, Bombus Spp. and Solitary Bees). EFSA J. 2012, 10, 2668. [Google Scholar] [CrossRef]
- Moritz, R.F.; De Miranda, J.; Fries, I.; Le Conte, Y.; Neumann, P.; Paxton, R.J. Research Strategies to Improve Honeybee Health in Europe. Apidologie 2010, 41, 227–242. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D.; Schwarz, R.S. Bees Brought to Their Knees: Microbes Affecting Honey Bee Health. Trends Microbiol. 2011, 19, 614–620. [Google Scholar] [CrossRef]
- Smith, K.M.; Loh, E.H.; Rostal, M.K.; Zambrana-Torrelio, C.M.; Mendiola, L.; Daszak, P. Pathogens, Pests, and Economics: Drivers of Honey Bee Colony Declines and Losses. EcoHealth 2013, 10, 434–445. [Google Scholar] [CrossRef]
- Gray, A.; Brodschneider, R.; Adjlane, N.; Ballis, A.; Brusbardis, V.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Cornelissen, B.; Amaro da Costa, C. Loss Rates of Honey Bee Colonies during Winter 2017/18 in 36 Countries Participating in the COLOSS Survey, Including Effects of Forage Sources. J. Apic. Res. 2019, 58, 479–485. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, T.; Kojima, Y.; Toki, T.; Komeda, Y.; Yoshiyama, M.; Kimura, K.; Nirasawa, K.; Kadowaki, T. The Habitat Disruption Induces Immune-Suppression and Oxidative Stress in Honey Bees. Ecol. Evol. 2011, 1, 201–217. [Google Scholar] [CrossRef]
- Bartlett, L.J.; Rozins, C.; Brosi, B.J.; Delaplane, K.S.; de Roode, J.C.; White, A.; Wilfert, L.; Boots, M. Industrial Bees: The Impact of Apicultural Intensification on Local Disease Prevalence. J. Appl. Ecol. 2019, 56, 2195–2205. [Google Scholar] [CrossRef] [Green Version]
- Genersch, E.; Evans, J.D.; Fries, I. Honey Bee Disease Overview. J. Invertebr. Pathol. 2010, 103, S2–S4. [Google Scholar] [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.; vanEngelsdorp, D. Drivers of Colony Losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- Raymann, K.; Moran, N.A. The Role of the Gut Microbiome in Health and Disease of Adult Honey Bee Workers. Curr. Opin. Insect Sci. 2018, 26, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A Metagenomic Survey of Microbes in Honey Bee Colony Collapse Disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.E.; Ricigliano, V.A. Honey Bee Gut Dysbiosis: A Novel Context of Disease Ecology. Vectors Med. Vet. Entomol. Soc. Insects 2017, 22, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Evison, S.E.; Jensen, A.B. The Biology and Prevalence of Fungal Diseases in Managed and Wild Bees. Curr. Opin. Insect Sci. 2018, 26, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Bailey, L.; Collins, M. Reclassification of ‘Streptococcus pluton’(White) in a New Genus Melissococcus, as Melissococcus pluton Nom. Rev. Comb. Nov. J. Appl. Bacteriol. 1982, 53, 215–217. [Google Scholar] [CrossRef]
- Trüper, H.G.; de’Clari, L. Taxonomic Note: Erratum and Correction of Further Specific Epithets Formed as Substantives (Nouns)‘in Apposition’. Int. J. Syst. Evol. Microbiol. 1998, 48, 615. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.D.; Munn, P.A. The Worldwide Health Status of Honey Bees. Bee World 2005, 86, 88–101. [Google Scholar] [CrossRef]
- Forsgren, E. European Foulbrood in Honey Bees. J. Invertebr. Pathol. 2010, 103, S5–S9. [Google Scholar] [CrossRef]
- Genersch, E.; Forsgren, E.; Pentikäinen, J.; Ashiralieva, A.; Rauch, S.; Kilwinski, J.; Fries, I. Reclassification of Paenibacillus larvae Subsp. pulvifaciens and Paenibacillus larvae Subsp. larvae as Paenibacillus larvae without Subspecies Differentiation. Int. J. Syst. Evol. Microbiol. 2006, 56, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Genersch, E. American Foulbrood in Honeybees and Its Causative Agent, Paenibacillus larvae. J. Invertebr. Pathol. 2010, 103, S10–S19. [Google Scholar] [CrossRef]
- Ebeling, J.; Knispel, H.; Hertlein, G.; Fünfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a Deadly Pathogen of Honey Bee Larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef] [PubMed]
- Genersch, E. Honey Bee Pathology: Current Threats to Honey Bees and Beekeeping. Appl. Microbiol. Biotechnol. 2010, 87, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Alippi, A. Bacterial Diseases of Honey Bees. Bee Health Vet. World Organ. Anim. Health 2014, 117–124. [Google Scholar]
- Amiri, E.; Strand, M.K.; Rueppell, O.; Tarpy, D.R. Queen Quality and the Impact of Honey Bee Diseases on Queen Health: Potential for Interactions between Two Major Threats to Colony Health. Insects 2017, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Alippi, A.M.; Leon, I.E.; López, A.C. Tetracycline-Resistance Encoding Plasmids from Paenibacillus larvae, the Causal Agent of American Foulbrood Disease, Isolated from Commercial Honeys. Int. Microbiol. 2014, 17, 49–61. [Google Scholar] [PubMed]
- Krongdang, S.; Evans, J.D.; Pettis, J.S.; Chantawannakul, P. Multilocus Sequence Typing, Biochemical and Antibiotic Resistance Characterizations Reveal Diversity of North American Strains of the Honey Bee Pathogen Paenibacillus larvae. PLoS ONE 2017, 12, e0176831. [Google Scholar] [CrossRef]
- Evans, J.D. Diverse Origins of Tetracycline Resistance in the Honey Bee Bacterial Pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Miyagi, T.; Peng, C.Y.S.; Chuang, R.Y.; Mussen, E.C.; Spivak, M.S.; Doi, R.H. Verification of Oxytetracycline-Resistant American Foulbrood Pathogen Paenibacillus larvae in the United States. J. Invertebr. Pathol. 2000, 75, 95–96. [Google Scholar] [CrossRef]
- Bargańska, Ż.; Namieśnik, J.; Ślebioda, M. Determination of Antibiotic Residues in Honey. Biog. Volatile Org. Compd. SI 2011, 30, 1035–1041. [Google Scholar] [CrossRef]
- Hammel, Y.-A.; Mohamed, R.; Gremaud, E.; LeBreton, M.-H.; Guy, P.A. Multi-Screening Approach to Monitor and Quantify 42 Antibiotic Residues in Honey by Liquid Chromatography–Tandem Mass Spectrometry. J. Chromatogr. A 2008, 1177, 58–76. [Google Scholar] [CrossRef]
- Lopez, M.I.; Pettis, J.S.; Smith, I.B.; Chu, P.-S. Multiclass Determination and Confirmation of Antibiotic Residues in Honey Using LC-MS/MS. J. Agric. Food Chem. 2008, 56, 1553–1559. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Wu, Y.; Chen, G.; Wang, S.; Hu, F.; Zheng, H. The Pass-on Effect of Tetracycline-Induced Honey Bee (Apis mellifera) Gut Community Dysbiosis. Front. Microbiol. 2022, 12, 781746. [Google Scholar] [CrossRef] [PubMed]
- Raymann, K.; Shaffer, Z.; Moran, N.A. Antibiotic Exposure Perturbs the Gut Microbiota and Elevates Mortality in Honeybees. PLoS Biol. 2017, 15, e2001861. [Google Scholar] [CrossRef]
- Mutinelli, F. Practical Application of Antibacterial Drugs for the Control of Honey Bee Diseases. APIACTA 2003, 38, 149–155. [Google Scholar]
- Hamdi, C.; Balloi, A.; Essanaa, J.; Crotti, E.; Gonella, E.; Raddadi, N.; Ricci, I.; Boudabous, A.; Borin, S.; Manino, A. Gut Mi-crobiome Dysbiosis and Honeybee Health. J. Appl. Entomol. 2011, 135, 524–533. [Google Scholar] [CrossRef] [Green Version]
- Alberoni, D.; Gaggìa, F.; Baffoni, L.; Di Gioia, D. Beneficial Microorganisms for Honey Bees: Problems and Progresses. Appl. Microbiol. Biotechnol. 2016, 100, 9469–9482. [Google Scholar] [CrossRef] [PubMed]
- Buford, T.W. (Dis) Trust Your Gut: The Gut Microbiome in Age-Related Inflammation, Health, and Disease. Microbiome 2017, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- Erban, T.; Ledvinka, O.; Kamler, M.; Nesvorna, M.; Hortova, B.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. Honeybee (Apis mellifera)-Associated Bacterial Community Affected by American Foulbrood: Detection of Paenibacillus Larvae via Micro-biome Analysis. Sci. Rep. 2017, 7, 5084. [Google Scholar] [CrossRef]
- Sopko, B.; Zitek, J.; Nesvorna, M.; Markovic, M.; Kamler, M.; Titera, D.; Erban, T.; Hubert, J. Detection and Quantification of Melissococcus plutonius in Honey Bee Workers Exposed to European Foulbrood in Czechia through Conventional PCR, QPCR, and Barcode Sequencing. J. Apic. Res. 2020, 59, 503–514. [Google Scholar] [CrossRef]
- Floyd, A.S.; Mott, B.M.; Maes, P.; Copeland, D.C.; McFREDERICK, Q.S.; Anderson, K.E. Microbial Ecology of European Foul Brood Disease in the Honey Bee (Apis mellifera): Towards a Microbiome Understanding of Disease Susceptibility. Insects 2020, 11, 555. [Google Scholar] [CrossRef]
- Sagona, S.; Turchi, B.; Fratini, F.; Giusti, M.; Torracca, B.; Nuvoloni, R.; Cerri, D.; Felicioli, A. Preliminary Evaluation of Glucose Oxidase and Its Products in Vitro Antimicrobial Activities on Paenibacillus larvae ATCC9545 Vegetative Form. Bull Insectol 2015, 68, 233–237. [Google Scholar]
- Alvarado, I.; Margotta, J.W.; Aoki, M.M.; Flores, F.; Agudelo, F.; Michel, G.; Elekonich, M.M.; Abel-Santos, E. Inhibitory Effect of Indole Analogs against Paenibacillus larvae, the Causal Agent of American Foulbrood Disease. J. Insect Sci. Online 2017, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Tsourkas, P.K. Paenibacillus larvae Bacteriophages: Obscure Past, Promising Future. Microb. Genom. 2020, 6, e000329. [Google Scholar] [CrossRef] [PubMed]
- Spivak, M.; Reuter, G.S. Resistance to American Foulbrood Disease by Honey Bee Colonies Apis mellifera Bred for Hygienic Behavior. Apidologie 2001, 32, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Grady, E.N.; Macdonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current Knowledge and Perspectives of Paenibacillus: A Review. Microb. Cell Factories 2016, 15, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Salces, R.M.; Cugnata, N.M.; Guaspari, E.; Pellegrini, M.C.; Aubone, I.; De Piano, F.G.; Antunez, K.; Fuselli, S.R. Natural Strategies for the Control of Paenibacillus larvae, the Causative Agent of American Foulbrood in Honey Bees: A Review. Apidologie 2017, 48, 387–400. [Google Scholar] [CrossRef]
- Flesar, J.; Havlik, J.; Kloucek, P.; Rada, V.; Titera, D.; Bednar, M.; Stropnicky, M.; Kokoska, L. In Vitro Growth-Inhibitory Effect of Plant-Derived Extracts and Compounds against Paenibacillus larvae and Their Acute Oral Toxicity to Adult Honey Bees. Vet. Microbiol. 2010, 145, 129–133. [Google Scholar] [CrossRef]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential Oils from Aromatic Herbs as Antimicrobial Agents. Food Biotechnol.-Plant Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef]
- Chaimanee, V.; Thongtue, U.; Sornmai, N.; Songsri, S.; Pettis, J. Antimicrobial Activity of Plant Extracts against the Honeybee Pathogens, Paenibacillus larvae and Ascosphaera apis and Their Topical Toxicity to Apis mellifera Adults. J. Appl. Microbiol. 2017, 123, 1160–1167. [Google Scholar] [CrossRef]
- Kim, J.; Park, S.; Shin, Y.-K.; Kang, H.; Kim, K.-Y. In Vitro Antibacterial Activity of Macelignan and Corosolic Acid against the Bacterial Bee Pathogens Paenibacillus larvae and Melissococcus plutonius. Acta Vet. Brno. 2018, 87, 277–284. [Google Scholar] [CrossRef]
- Evans, J.D.; Armstrong, T.-N. Antagonistic Interactions between Honey Bee Bacterial Symbionts and Implications for Disease. BMC Ecol. 2006, 6, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.E.; Sheehan, T.; Eckholm, B.; Mott, B.; DeGrandi-Hoffman, G. An Emerging Paradigm of Colony Health: Microbial Balance of the Honey Bee and Hive (Apis mellifera). Insectes Sociaux 2011, 58, 431–444. [Google Scholar] [CrossRef]
- Vásquez, A.; Forsgren, E.; Fries, I.; Paxton, R.J.; Flaberg, E.; Szekely, L.; Olofsson, T.C. Symbionts as Major Modulators of Insect Health: Lactic Acid Bacteria and Honeybees. PLoS ONE 2012, 7, e33188. [Google Scholar] [CrossRef]
- Cariveau, D.P.; Elijah Powell, J.; Koch, H.; Winfree, R.; Moran, N.A. Variation in Gut Microbial Communities and Its Associ-ation with Pathogen Infection in Wild Bumble Bees (Bombus). ISME J. 2014, 8, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Forfert, N.; Natsopoulou, M.E.; Frey, E.; Rosenkranz, P.; Paxton, R.J.; Moritz, R.F. Parasites and Pathogens of the Honeybee (Apis mellifera) and Their Influence on Inter-Colonial Transmission. PLoS ONE 2015, 10, e0140337. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Baffoni, L.; Alberoni, D. Probiotics for Honeybees’ Health. In Probiotics and Prebiotics in Animal Health and Food Safety; Springer: Cham, Switzerland, 2018; pp. 219–245. [Google Scholar]
- Alberoni, D.; Baffoni, L.; Gaggìa, F.; Ryan, P.; Murphy, K.; Ross, P.; Stanton, C.; Di Gioia, D. Impact of Beneficial Bacteria Supplementation on the Gut Microbiota, Colony Development and Productivity of Apis mellifera L. Benef. Microbes 2018, 9, 269–278. [Google Scholar] [CrossRef]
- Zheng, H.; Steele, M.I.; Leonard, S.P.; Motta, E.V.S.; Moran, N.A. Honey Bees as Models for Gut Microbiota Research. Lab. Anim. 2018, 47, 317–325. [Google Scholar] [CrossRef]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial Ecology of the Hive and Pollination Landscape: Bacterial Associates from Floral Nectar, the Alimentary Tract and Stored Food of Honey Bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Pannella, G.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Succi, M.; Sorrentino, E.; Petrarca, S.; De Cristofaro, A.; Coppola, R.; et al. Inter- and Intra-Species Diversity of Lactic Acid Bacteria in Apis mellifera ligustica Colonies. Microorganisms 2020, 8, 1578. [Google Scholar] [CrossRef]
- Gaggìa, F.; Baffoni, L.; Stenico, V.; Alberoni, D.; Buglione, E.; Lilli, A.; Di Gioia, D.; Porrini, C. Microbial Investigation on Honey Bee Larvae Showing Atypical Symptoms of European Foulbrood. Bull. Insectology 2015, 68, 321–327. [Google Scholar]
- Olofsson, T.C.; Vásquez, A. Detection and Identification of a Novel Lactic Acid Bacterial Flora within the Honey Stomach of the Honeybee Apis mellifera. Curr. Microbiol. 2008, 57, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Cilia, G.; Fratini, F.; Tafi, E.; Turchi, B.; Mancini, S.; Sagona, S.; Nanetti, A.; Cerri, D.; Felicioli, A. Microbial Profile of the Ventriculum of Honey Bee (Apis mellifera ligustica Spinola, 1806) Fed with Veterinary Drugs, Dietary Supplements and Non-Protein Amino Acids. Vet. Sci. 2020, 7, 76. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; Ganassi, S.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A. Antimicrobial Activity against Paenibacillus larvae and Functional Properties of Lactiplantibacillus plantarum Strains: Po-tential Benefits for Honeybee Health. Antibiotics 2020, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Gaggìa, F.; Di Gioia, D.; Baffoni, L.; Biavati, B. The Role of Protective and Probiotic Cultures in Food and Feed and Their Impact in Food Safety. Pathog.-Unique Achiev. Fight. Pathog. 2011, 22, S58–S66. [Google Scholar] [CrossRef]
- Iorizzo, M.; Lombardi, S.J.; Ganassi, S.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; Vergalito, F.; Cozzolino, A.; et al. Antagonistic Activity against Ascosphaera apis and Functional Properties of Lactobacillus kunkeei Strains. Antibiotics 2020, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Daisley, B.A.; Pitek, A.P.; Chmiel, J.A.; Al, K.F.; Chernyshova, A.M.; Faragalla, K.M.; Burton, J.P.; Thompson, G.J.; Reid, G. Novel Probiotic Approach to Counter Paenibacillus larvae Infection in Honey Bees. ISME J. 2020, 14, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Killer, J.; Dubná, S.; Sedláček, I.; Švec, P. Lactobacillus apis Sp. Nov., from the Stomach of Honeybees (Apis mellifera), Having an In Vitro Inhibitory Effect on the Causative Agents of American and European Foulbrood. Int. J. Syst. Evol. Microbiol. 2014, 64, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Maeno, S.; Tanizawa, Y.; Kneifel, W.; Arita, M.; Dicks, L.; Salminen, S. Fructophilic Lactic Acid Bacteria, a Unique Group of Fructose-Fermenting Microbes. Appl. Environ. Microbiol 2018, 84, e01290-18. [Google Scholar] [CrossRef] [Green Version]
- Endo, A.; Salminen, S. Honeybees and Beehives Are Rich Sources for Fructophilic Lactic Acid Bacteria. Syst. Appl. Microbiol. 2013, 36, 444–448. [Google Scholar] [CrossRef]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.; Rademaker, J.L.; Starrenburg, M.J.; Kleerebezem, M.; Molenaar, D.; van Hylckama Vlieg, J.E. Phenotypic and Genomic Diversity of Lactobacillus plantarum Strains Isolated from Various Environmental Niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms 2021, 9, 349. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ni, X.; Wang, Q.; Peng, Z.; Niu, L.; Wang, H.; Zhou, Y.; Sun, H.; Pan, K.; Jing, B. Lactobacillus plantarum BSGP201683 Isolated from Giant Panda Feces Attenuated Inflammation and Improved Gut Microflora in Mice Challenged with Enterotoxigenic Escherichia coli. Front. Microbiol. 2017, 8, 1885. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, J.K.; Kumar, A.; Duary, R.K.; Mohanty, A.K.; Grover, S.; Batish, V.K. Functional and Probiotic Attributes of an Indigenous Isolate of Lactobacillus plantarum. PLoS ONE 2009, 4, e8099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Jia, D.; Wang, J.; Li, H.; Yin, X.; Liu, J.; Wang, J.; Guan, G.; Luo, J.; Yin, H.; et al. Probiotics Isolated From Animals in Northwest China Improve the Intestinal Performance of Mice. Front. Vet. Sci. 2021, 8, 750895. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Lombardi, S.J.; Macciola, V.; Testa, B.; Lustrato, G.; Lopez, F.; De Leonardis, A. Technological Potential of Lactobacillus Strains Isolated from Fermented Green Olives: In Vitro Studies with Emphasis on Oleuropein-Degrading Capability. Sci. World J. 2016, 2016, 1917592. [Google Scholar] [CrossRef] [Green Version]
- Iorizzo, M.; Albanese, G.; Testa, B.; Ianiro, M.; Letizia, F.; Succi, M.; Tremonte, P.; D’Andrea, M.; Iaffaldano, N.; Coppola, R. Presence of Lactic Acid Bacteria in the Intestinal Tract of the Mediterranean Trout (Salmo macrostigma) in Its Natural Environment. Life 2021, 11, 667. [Google Scholar] [CrossRef]
- Iorizzo, M.; Testa, B.; Lombardi, S.J.; García-Ruiz, A.; Muñoz-González, C.; Bartolomé, B.; Moreno-Arribas, M.V. Selection and technological potential of Lactobacillus plantarum bacteria suitable for wine malolactic fermentation and grape aroma release. LWT 2016, 73, 557–566. [Google Scholar] [CrossRef] [Green Version]
- Bielik, B.; Molnár, L.; Vrabec, V.; Andrášiová, R.; Maruščáková, I.C.; Nemcová, R.; Toporčák, J.; Mudroňová, D. Bio-film-Forming Lactic Acid Bacteria of Honey Bee Origin Intended for Potential Probiotic Use. Acta Vet. Hung. 2021, 68, 345–353. [Google Scholar] [CrossRef]
- Lamei, S.; Stephan, J.G.; Riesbeck, K.; Vasquez, A.; Olofsson, T.; Nilson, B.; de Miranda, J.R.; Forsgren, E. The Secretome of Honey Bee-Specific Lactic Acid Bacteria Inhibits Paenibacillus larvae Growth. J. Apic. Res. 2019, 58, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Andrearczyk, S.; Kadhim, M.J.; Knaga, S. Influence of a Probiotic on the Mortality, Sugar Syrup Ingestion and Infection of Honeybees with Nosema Spp. under Laboratory Assessment. Med. Weter 2014, 70, 762. [Google Scholar]
- Ptaszyńska, A.A.; Borsuk, G.; Zdybicka-Barabas, A.; Cytryńska, M.; Małek, W. Are Commercial Probiotics and Prebiotics Ef-fective in the Treatment and Prevention of Honeybee Nosemosis C? Parasitol. Res. 2016, 115, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A Greedy Algorithm for Aligning DNA Sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2018, 46, D8–D13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Iorizzo, M.; Albanese, G.; Letizia, F.; Testa, B.; Tremonte, P.; Vergalito, F.; Lombardi, S.J.; Succi, M.; Coppola, R.; Sorrentino, E. Probiotic Potentiality from Versatile Lactiplantibacillus plantarum Strains as Resource to Enhance Freshwater Fish Health. Microorganisms 2022, 10, 463. [Google Scholar] [CrossRef] [PubMed]
- Testa, B.; Lombardi, S.J.; Macciola, E.; Succi, M.; Tremonte, P.; Iorizzo, M. Efficacy of Olive Leaf Extract (Olea europaea L. Cv Gentile di Larino) in Marinated Anchovies (Engraulis Encrasicolus, L.) Process. Heliyon 2019, 5, e01727. [Google Scholar] [CrossRef] [Green Version]
- Cozzolino, A.; Vergalito, F.; Tremonte, P.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E.; Luongo, D.; Coppola, R.; Di Marco, R.; Succi, M. Preliminary Evaluation of the Safety and Probiotic Potential of Akkermansia muciniphila DSM 22959 in Comparison with Lactobacillus rhamnosus GG. Microorganisms 2020, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Endo, A. Fructophilic Lactic Acid Bacteria Inhabit Fructose-Rich Niches in Nature. Microb. Ecol. Health Dis. 2012, 23, 18563. [Google Scholar] [CrossRef]
- Corby-Harris, V.; Maes, P.; Anderson, K.E. The Bacterial Communities Associated with Honey Bee (Apis mellifera) Foragers. PLoS ONE 2014, 9, e95056. [Google Scholar] [CrossRef] [Green Version]
- Tajabadi, N.; Mardan, M.; Saari, N.; Mustafa, S.; Bahreini, R.; Manap, M.Y.A. Identification of Lactobacillus plantarum, Lactobacillus pentosus and Lactobacillus fermentum from Honey Stomach of Honeybee. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2014, 44, 717–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honey Chandran, C.; TR, K. Probiotic Potency of Lactobacillus plantarum KX519413 and KX519414 Isolated from Honey Bee Gut. FEMS Microbiol. Lett. 2018, 365, fnx285. [Google Scholar]
- Martino, M.E.; Bayjanov, J.R.; Caffrey, B.E.; Wels, M.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebezem, M.; van Hijum, S.A.; Leulier, F. Nomadic Lifestyle of Lactobacillus plantarum Revealed by Comparative Genomics of 54 Strains Isolated from Different Habitats. Environ. Microbiol. 2016, 18, 4974–4989. [Google Scholar] [CrossRef] [PubMed]
- Gustaw, K.; Michalak, M.; Polak-Berecka, M.; Waśko, A. Isolation and Characterization of a New Fructophilic Lactobacillus plantarum FPL Strain from Honeydew. Ann. Microbiol. 2018, 68, 459–470. [Google Scholar] [CrossRef]
- Reis, J.A.; Paula, A.T.; Casarotti, S.N.; Penna, A.L.B. Lactic Acid Bacteria Antimicrobial Compounds: Characteristics and Applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Powell, J.E.; Martinson, V.G.; Urban-Mead, K.; Moran, N.A. Routes of Acquisition of the Gut Microbiota of the Honey Bee Apis mellifera. Appl. Environ. Microbiol. 2014, 80, 7378–7387. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Nishida, A.; Kwong, W.K.; Koch, H.; Engel, P.; Steele, M.I.; Moran, N.A. Metabolism of Toxic Sugars by Strains of the Bee Gut Symbiont Gilliamella apicola. MBio 2016, 7, e01326-16. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Perreau, J.; Powell, J.E.; Han, B.; Zhang, Z.; Kwong, W.K.; Tringe, S.G.; Moran, N.A. Division of Labor in Honey Bee Gut Microbiota for Plant Polysaccharide Digestion. Proc. Natl. Acad. Sci. USA 2019, 116, 25909–25916. [Google Scholar] [CrossRef]
- Stanley, D.; Rejzek, M.; Naested, H.; Smedley, M.; Otero, S.; Fahy, B.; Thorpe, F.; Nash, R.J.; Harwood, W.; Svensson, B.; et al. The Role of α-Glucosidase in Germinating Barley Grains. Plant. Physiol. 2011, 155, 932–943. [Google Scholar] [CrossRef] [Green Version]
- Lee, F.J.; Rusch, D.B.; Stewart, F.J.; Mattila, H.R.; Newton, I.L. Saccharide Breakdown and Fermentation by the Honey Bee Gut Microbiome. Environ. Microbiol. 2015, 17, 796–815. [Google Scholar] [CrossRef]
- Seeburger, V.C.; D’Alvise, P.; Shaaban, B.; Schweikert, K.; Lohaus, G.; Schroeder, A.; Hasselmann, M. The Trisaccharide Melezitose Impacts Honey Bees and Their Intestinal Microbiota. PLoS ONE 2020, 15, e0230871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.M. Honey Bee Toxicology. Annu. Rev. Entomol. 2015, 60, 415–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorizzo, M.; Letizia, F.; Ganassi, S.; Testa, B.; Petrarca, S.; Albanese, G.; Di Criscio, D.; De Cristofaro, A. Functional Properties and Antimicrobial Activity from Lactic Acid Bacteria as Resources to Improve the Health and Welfare of Honey Bees. Insects 2022, 13, 308. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Letizia, F.; Ganassi, S.; Testa, B.; Petrarca, S.; Albanese, G.; Di Criscio, D.; De Cristofaro, A. Recent Advances in the Biocontrol of Nosemosis in Honey Bees (Apis Mellifera L.). J. Fungi 2022, 8, 424. [Google Scholar] [CrossRef]
- Collado, M.C.; Meriluoto, J.; Salminen, S. Adhesion and Aggregation Properties of Probiotic and Pathogen Strains. Eur. Food Res. Technol. 2008, 226, 1065–1073. [Google Scholar] [CrossRef]
- Deng, Z.; Luo, X.M.; Liu, J.; Wang, H. Quorum Sensing, Biofilm, and Intestinal Mucosal Barrier: Involvement the Role of Probiotic. Front. Cell. Infect. Microbiol. 2020, 10, 538077. [Google Scholar] [CrossRef]
- Trunk, T.; Khalil, H.S.; Leo, J.C. Bacterial Autoaggregation. AIMS Microbiol. 2018, 4, 140–164. [Google Scholar] [CrossRef] [PubMed]
- Salas-Jara, M.J.; Ilabaca, A.; Vega, M.; García, A. Biofilm Forming Lactobacillus: New Challenges for the Development of Probiotics. Microorganisms 2016, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Kos, B.; Šušković, J.; Vuković, S.; Šimpraga, M.; Frece, J.; Matošić, S. Adhesion and Aggregation Ability of Probiotic Strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 2003, 94, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Gonzalez, E.; Genersch, E. Honey Bee Larval Peritrophic Matrix Degradation during Infection with Paenibacillus larvae, the Aetiological Agent of AmericanFoulbrood of Honey Bees, Is a Key Step in Pathogenesis. Environ. Microbiol. 2013, 15, 2894–2901. [Google Scholar]
- Grossar, D.; Kilchenmann, V.; Forsgren, E.; Charrière, J.-D.; Gauthier, L.; Chapuisat, M.; Dietemann, V. Putative Determinants of Virulence in Melissococcus plutonius, the Bacterial Agent Causing European Foulbrood in Honey Bees. Virulence 2020, 11, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ordóñez, G.; Romo-Chacón, A.; Rios-Velasco, C.; Sepúlveda, D.R.; de Jesús Ornelas-Paz, J.; Acosta-Muñiz, C.H. Viru-lence Variations between Clonal Complexes of Melisococcus plutonius and the Possible Causes. J. Invertebr. Pathol. 2021, 186, 107686. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Okumura, K.; Harada, M.; Okamoto, M.; Okura, M.; Takamatsu, D. Peritrophic Matrix-Degrading Proteins Are Dispensable Virulence Factors in a Virulent Melissococcus plutonius Strain. Sci. Rep. 2021, 11, 8798. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | ||||||||
|---|---|---|---|---|---|---|---|---|
| Matrices (CSF) | LP31 | LP42 | LP148 | LP179 | ALK181 | ALK222 | ALK268 | ALK385 |
| P. larvae | 14.9 ± 0.2 b | 15.1 ± 0.5 b | 15.8 ± 0.3 a | 13.9 ± 0.4 c | 15.5 ± 0.5 a | 16.3 ± 0.2 a | 15.0 ± 0.4 b | 13.7 ± 0.2 c |
| M. plutonius | 15.1 ± 0.3 a | 15.1 ± 0.3 a | 16.0 ± 0.1 a | 12.0 ± 0.2 d | 15.9 ± 0.4 a | 14.0 ± 0.3 b | 14.2 ± 0.3 b | 13.0 ± 0.5 c |
| Hydrophobicity (%) Xylene | ||||||||
| Time | LP31 | LP42 | LP148 | LP179 | ALK181 | ALK222 | ALK268 | ALK385 |
| 15 | 70.8 ± 1.3 Cc | 80.5 ± 2.6 Bb | 72.2 ± 1.8 Bc | 85.3 ± 1.1 Bb | 84.0 ± 1.7 Bb | 72.3 ± 2.7 Bc | 95.4 ± 1.5 Aa | 76.1 ± 2.6 Ac |
| 30 | 76.0 ± 2.2 Bc | 81.4 ± 1.9 Bc | 77.9 ± 1.7 Ac | 87.8 ± 2.9 Ab | 88.6 ± 1.8 Ab | 79.6 ± 2.6 Ac | 97.3 ± 1.5 Aa | 76.2 ± 1.0 Ac |
| 60 | 82.8 ± 3.9 Ac | 90.9 ± 2.3 Ab | 78.7 ± 1.2 Ac | 91.3 ± 1.4 Ab | 91.9 ± 1.9 Ab | 81.9 ± 3.3 Ac | 98.0 ± 0.5 Aa | 80.1 ± 2.3 Ac |
| Hydrophobicity (%) Toluene | ||||||||
| Time | LP31 | LP42 | LP148 | LP179 | ALK181 | ALK222 | ALK268 | ALK385 |
| 15 | 74.7 ± 2.0 Cc | 77.9 ± 0.5 Cb | 72.2 ± 1.7 Ac | 79.1 ± 0.8 Cb | 92.1 ± 1.6 Aa | 72.8 ± 0.4 Cc | 47.7 ± 1.0 Ae | 53.1 ± 0.4 Bd |
| 30 | 77.4 ± 2.5 Bd | 88.4 ± 0.4 Bb | 73.6 ± 1.2 Ae | 84.0 ± 0.5 Bc | 92.8 ± 0.3 Aa | 75.5 ± 1.2 Bd | 47.8 ± 1.9 Ag | 54.9 ± 2.3 Bf |
| 60 | 85.5 ± 2.4 Ab | 94.1 ± 0.9 Aa | 74.4 ± 0.4 Ac | 93.5 ± 0.7 Aa | 93.2 ± 0.3 Aa | 83.1 ± 1.7 Ab | 50.0 ± 0.3 Ae | 60.2 ± 1.2 Ad |
| Auto-Aggregation % | ||||||||
|---|---|---|---|---|---|---|---|---|
| Time (h) | LP31 | LP42 | LP148 | LP179 | ALK181 | ALK222 | ALK268 | ALK385 |
| 1 | 12.4 ± 0.3 Db | 11.2 ± 0.5 Cb | 9.5 ± 0.5 Db | 12.3 ± 0.9 Cb | 19.0 ± 0.9 Da | 8.93 ± 0.2 Cb | 9.1 ± 0.1 Db | 10.2 ± 0.4 Db |
| 2 | 17.0 ± 1.3 Cb | 15.1 ± 1.8 Cb | 14.5 ± 1.6 Cb | 13.8 ± 1.2 Cb | 24.4 ± 1.9 Ca | 12.4 ± 0.7 Cb | 14.7 ± 0.8 Cb | 17.0 ± 1.6 Cb |
| 5 | 29.4 ± 3.0 Bb | 24.0 ± 2.6 Bc | 30.2 ± 2.5 Bb | 31.5 ± 1.8 Ba | 36.1 ± 2.4 Ba | 26.3 ± 1.7 Bb | 24.8 ± 1.0 Bc | 28.8 ± 2.1 Bb |
| 24 | 68.8 ± 3.6 Aa | 59.2 ± 2.5 Ab | 57.9 ± 2.0 Ab | 58.7 ± 2.9 Ab | 60.0 ± 2.3 Ab | 51.1 ± 2.2 Ac | 59.2 ± 1.7 Ab | 56.8 ± 2.4 Ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iorizzo, M.; Ganassi, S.; Albanese, G.; Letizia, F.; Testa, B.; Tedino, C.; Petrarca, S.; Mutinelli, F.; Mazzeo, A.; De Cristofaro, A. Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases. Vet. Sci. 2022, 9, 236. https://doi.org/10.3390/vetsci9050236
Iorizzo M, Ganassi S, Albanese G, Letizia F, Testa B, Tedino C, Petrarca S, Mutinelli F, Mazzeo A, De Cristofaro A. Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases. Veterinary Sciences. 2022; 9(5):236. https://doi.org/10.3390/vetsci9050236
Chicago/Turabian StyleIorizzo, Massimo, Sonia Ganassi, Gianluca Albanese, Francesco Letizia, Bruno Testa, Cosimo Tedino, Sonia Petrarca, Franco Mutinelli, Alessandra Mazzeo, and Antonio De Cristofaro. 2022. "Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases" Veterinary Sciences 9, no. 5: 236. https://doi.org/10.3390/vetsci9050236
APA StyleIorizzo, M., Ganassi, S., Albanese, G., Letizia, F., Testa, B., Tedino, C., Petrarca, S., Mutinelli, F., Mazzeo, A., & De Cristofaro, A. (2022). Antimicrobial Activity from Putative Probiotic Lactic Acid Bacteria for the Biological Control of American and European Foulbrood Diseases. Veterinary Sciences, 9(5), 236. https://doi.org/10.3390/vetsci9050236