
Relationship among Serum Progestagens, Cortisol, and Prolactin in Pregnant and Cycling Asian Elephants in Thailand

 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Hormonal Analysis
2.3. Statistical Analysis
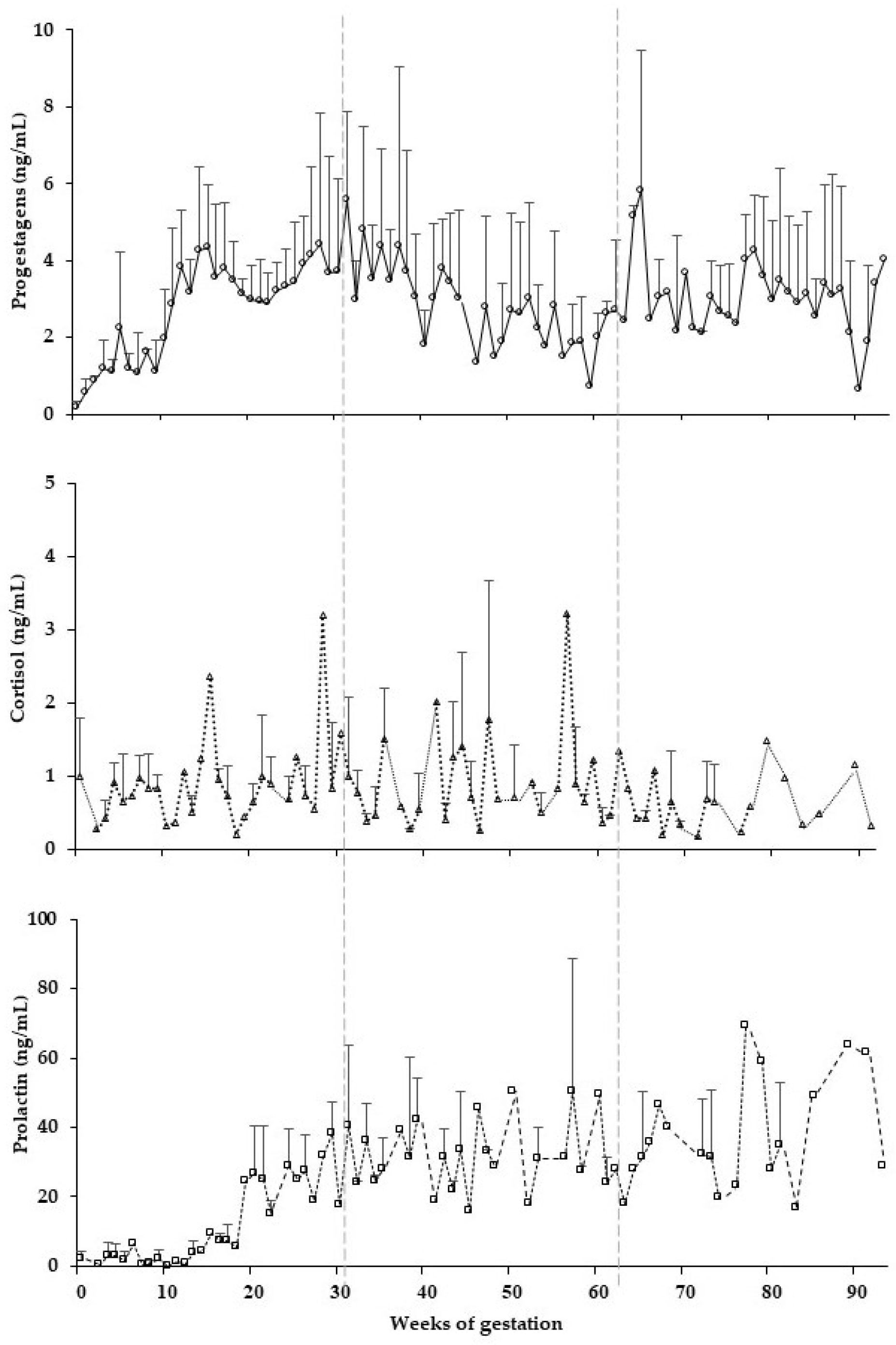
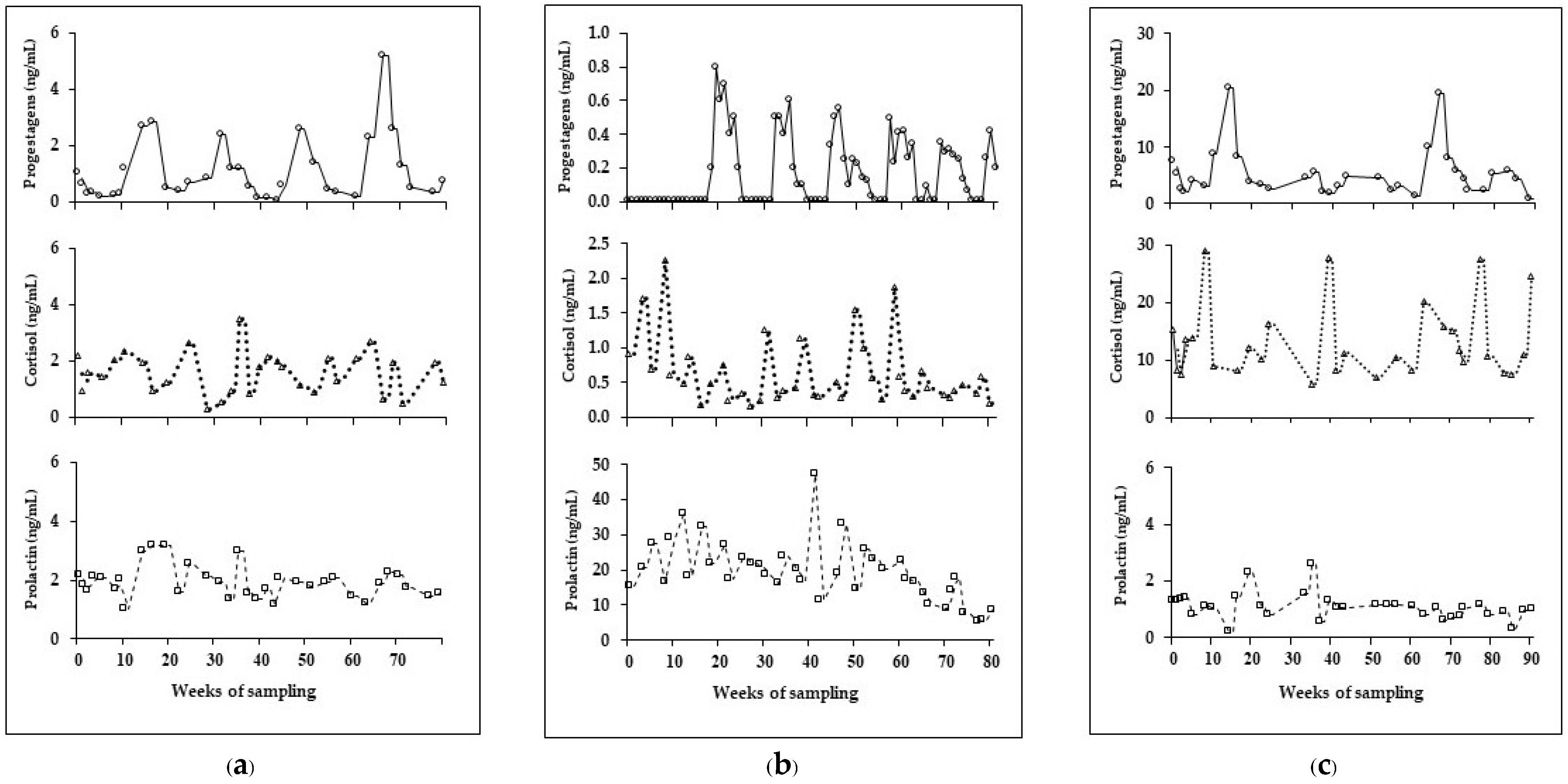
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Menon, V.; Tiwari, S.K. Population Status of Asian Elephants Elephas maximus and Key Threats. Int. Zoo Yearb. 2019, 53, 17–30. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Brown, J.L.; Thitaram, C.; Punyapornwithaya, V.; Nganvongpanit, K. Elephant Tourism in Thailand: A Review of Animal Welfare Practices and Needs. J. Appl. Anim. Welf. Sci. 2020, 23, 164–177. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L. Comparative Reproductive Biology of Elephants. In Reproductive Sciences in Animal Conservation, Advance in Experimental Medicine and Biology; Al, W.V.H., Ed.; Springer: New York, NY, USA, 2014; Volume 753, pp. 135–169. ISBN 9781493908196. [Google Scholar]
- Thitaram, C. Breeding Management of Captive Asian Elephant (Elephas maximus) in Range Countries and Zoos. Jpn. J. Zoo Wildl. Med. 2012, 17, 91–96. [Google Scholar] [CrossRef]
- Brown, J.L.; Lehnhardt, J. Serum and Urinary Hormones during Pregnancy and the Peri- and Postpartum Period in an Asian Elephant (Elephas maximus). Zoo Biol. 1995, 14, 555–564. [Google Scholar] [CrossRef]
- Meyer, J.M.; Walker, S.L.; Freeman, E.W.; Steinetz, B.G.; Brown, J.L. Species and Fetal Gender Effects on the Endocrinology of Pregnancy in Elephants. Gen. Comp. Endocrinol. 2004, 138, 263–270. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamamoto, T.; Yuto, N.; Shiina, O.; Mouri, Y.; Sugimura, K.; Nagaoka, K.; Watanabe, G.; Taya, K. The Secretary Pattern and Physiological Role of Prolactin in Pregnant African (Loxodonta africana) and Asian (Elephas maximus) Elephants. In Proceedings of the International Conference on Elephant and Wildlife Health Management in Asia, Bangkok, Thailand, 5 September 2011; pp. 113–118. [Google Scholar]
- Yamamoto, Y.; Yamamoto, T.; Taya, K.; Watanabe, G.; Stansfield, F.J.; Allen, W.R. Placentation in the African Elephant (Loxodonta africana). V: The Trophoblast Secretes Placental Lactogen. Placenta 2011, 32, 506–510. [Google Scholar] [CrossRef]
- Forsyth, I.A.; Wallis, M. Growth Hormone and Prolactin–Molecular and Functional Evolution. J. Mammary Gland. Biol. Neoplasia 2002, 7, 291–312. [Google Scholar] [CrossRef]
- Freeman, M.E.; Kanyicska, B.; Lerant, A.; Nagy, G. Prolactin: Structure, Function, and Regulation of Secretion. Physiol. Rev. 2000, 80, 1523–1631. [Google Scholar] [CrossRef]
- Takahashi, T. Biology of the Prolactin Family in Bovine Placenta. I. Bovine Placental Lactogen: Expression, Structure and Proposed Roles. Anim. Sci. J. 2006, 77, 10–17. [Google Scholar] [CrossRef]
- Allen, W.R.; Mathias, S.S.; Wooding, F.B.; Skidmore, J.A.; van Aarde, R.J. Placentation in the African Elephant, Loxodonta africana. I. Endocrinological Aspects. Reprod. Suppl. 2002, 60, 105–116. [Google Scholar]
- Brown, J.L. Update on Comparative Biology of Elephants: Factors Affecting Reproduction, Health and Welfare. Adv. Exp. Med. Biol. 2019, 1200, 243–273. [Google Scholar] [CrossRef] [PubMed]
- Kajaysri, J.; Nokkaew, W. Assessment of Pregnancy Status of Asian Elephants (Elephas) by Measurement of Progestagen and Glucocorticoid and Their Concentrations in Serum and Feces, Using Enzyme Immunoassay (EIA). J. Vet. Med. Sci. 2014, 76, 363. [Google Scholar] [CrossRef] [PubMed]
- Fanson, K.V.; Keeley, T.; Fanson, B.G. Cyclic Changes in Cortisol across the Estrous Cycle in Parous and Nulliparous Asian Elephants. Endocr. Connect. 2014, 3, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Bechert, U.S.; Swanson, L.; Wasser, S.K.; Hess, D.L.; Stormshak, F. Serum Prolactin Concentrations in the Captive Female African Elephant (Loxodonta Africana): Potential Effects of Season and Steroid Hormone Interactions. Gen. Comp. Endocrinol. 1999, 114, 269–278. [Google Scholar] [CrossRef]
- Brown, J.L.; Lehnhardt, J. Secretory Patterns of Serum Prolactin in Asian (Elephas maximus) and African (Loxodonta Africana) Elephants during Different Reproductive States: Comparison with Concentrations in a Noncycling African Elephant. Zoo Biol. 1997, 16, 149–159. [Google Scholar] [CrossRef][Green Version]
- Prado-Oviedo, N.A.; Malloy, E.J.; Deng, X.; Brown, J.L. Hyperprolactinemia Is Not Associated with Hyperestrogenism in Noncycling African Elephants (Loxodonta africana). Gen. Comp. Endocrinol. 2013, 189, 7–14. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Yamamoto, T.; Watanabe, G.; Yuto, N.; Keio, M.; Narushima, E.; Katayanagi, M.; Nakao, R.; Morikubo, S.; Sakurai, Y.; et al. Prolactin Secretion and Ovarian Function in Cycling and Non-Cycling African Female Elephants (Loxodonta africana). J. Vet. Med. Sci. 2010, 72, 845–852. [Google Scholar] [CrossRef]
- Dow, T.L.; Brown, J.L. Evidence That Hyperprolactinaemia Is Associated with Ovarian Acyclicity in Female Zoo African Elephants. Reprod. Fertil. Dev. 2012, 24, 1019–1027. [Google Scholar] [CrossRef][Green Version]
- Menargues, A.; Urios, V.; Limiñana, R. Seasonal Rhythms of Salivary Cortisol Secretion in Captive Asian Elephants (Elephas maximus). Gen. Comp. Endocrinol. 2012, 176, 259–264. [Google Scholar] [CrossRef]
- Menargues, A.; Urios, V.; Limiñana, R.; Mauri, M. Circadian Rhythm of Salivary Cortisol in Asian Elephants (Elephas maximus) A Factor to Consider during Welfare Assessment. J. Appl. Anim. Welf. Sci. 2012, 15, 383–390. [Google Scholar] [CrossRef]
- Brown, J.L.; Wielebnowski, N.; Cheeran, J.V. Pain, Stress, and Suffering in Elephants: What Is the Evidence and How Can We Measure It? In Elephants and Ethics: Toward a Morality of Coexistence; Wemmer, C.M., Christen, C.A., Eds.; Johns Hopkins University Press: Baltimore, MD, USA, 2008; pp. 121–145. [Google Scholar]
- Dallman, M.F. Chronic (Repeated) Stress: Consequences, Adaptations. Encycl. Neurosci. 2009, 1, 879–885. [Google Scholar] [CrossRef]
- Buckingham, J.C. Glucocorticoids: Exemplars of Multi-Tasking. Br. J. Pharmacol. 2006, 147, S258–S268. [Google Scholar] [CrossRef] [PubMed]
- Möstl, E.; Palme, R. Hormones as Indicators of Stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Thitaram, C.; Brown, J.L.; Pongsopawijit, P.; Chansitthiwet, S.; Wongkalasin, W.; Daram, P.; Roongsri, R.; Kalmapijit, A.; Mahasawangkul, S.; Rojansthien, S.; et al. Seasonal Effects on the Endocrine Pattern of Semi-Captive Female Asian Elephants (Elephas maximus): Timing of the Anovulatory Luteinizing Hormone Surge Determines the Length of the Estrous Cycle. Theriogenology 2008, 69, 237–244. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Hunt, K.E.; Martin, M.S.; Finnegan, M.; Brown, J.L. Investigation of Individual and Group Variability in Estrous Cycle Characteristics in Female Asian Elephants (Elephas maximus) at the Oregon Zoo. Theriogenology 2012, 78, 285–296. [Google Scholar] [CrossRef]
- Carden, M.; Schmitt, D.; Tomasi, T.; Bradford, J.; Moll, D.; Brown, J. Utility of Serum Progesterone and Prolactin Analysis for Assessing Reproductive Status in the Asian Elephant (Elephas maximus). Anim. Reprod. Sci. 1998, 53, 133–142. [Google Scholar] [CrossRef]
- Hodges, J.K.; Mcneilly, A.S.; Hess, D.L. Circulating Hormones during Pregnancy in the Asian and African Elephants Elephas maximus and Loxodonta africana a Diagnostic Test Based on the Measurement of Prolactin. Int. Zoo Yearb. 1987, 26, 285–289. [Google Scholar] [CrossRef]
- McNeilly, A.S.; Martin, R.D.; Hodges, J.K.; Smuts, G.L. Blood Concentrations of Gonadotrophins, Prolactin and Gonadal Steroids in Males and in Non-Pregnant and Pregnant Female African Elephants (Loxodonta africana). J. Reprod. Fertil. 1983, 67, 113–120. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; LaPensee, C.R.; LaPensee, E.W. What Can We Learn from Rodents about Prolactin in Humans? Endocr. Rev. 2008, 29, 1–41. [Google Scholar] [CrossRef]
- Chen, J.C.; Lin, J.-H.; Wu, L.-S.; Tsai, Y.-F.; Su, T.H.; Chen, C.J.; Chen, T.J. Luteotropic Roles of Prolactin in Early Pregnant Hamsters. Biol. Reprod. 2002, 67, 8–13. [Google Scholar] [CrossRef][Green Version]
- Yamamoto, Y.; Yamamoto, T.; Yuto, N.; Hildebrandt, T.B.; Lueders, I.; Wibbelt, G.; Shiina, O.; Mouri, Y.; Sugimura, K.; Sakamoto, S.; et al. The Secretory Pattern and Source of Immunoreactive Prolactin in Pregnant African (Loxodonta africana) and Asian (Elephas maximus) Elephants. J. Reprod. Dev. 2012, 58, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Walker, S.L.; Moeller, T. Comparative Endocrinology of Cycling and Non-Cycling Asian (Elephas maximus) and African (Loxodonta africana) Elephants. Gen. Comp. Endocrinol. 2004, 136, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Pushpakumara, A.; Thitaram, C.; Brown, J.L. Elephants. In Veterinary Reproduction and Obstetrics; Noakes, D., Parkinson, T., England, G., Eds.; W.B. Saunders: Philadelphia, PA, USA, 2018; ISBN 9780702072338. [Google Scholar]
- Lueders, I.; Niemuller, C.; Steinmetz, H.W.; Bouts, T.; Gray, C.; Knauf-Witzens, T.; Taya, K.; Watanabe, G.; Yamamoto, Y.; Oerke, A.K. Prolonged Luteal Lifespan and Pseudopregnancy in Asian Elephants (Elephas maximus). Anim. Reprod. Sci. 2018, 197, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.A.; Felippe, E.C.G.; Chelini, M.O.M. Serum Cortisol and Progestin Concentrations in Pregnant and Non-Pregnant Asian Elephants (Elephas maximus). Res. Vet. Sci. 2008, 84, 361–363. [Google Scholar] [CrossRef]
- Bansiddhi, P.; Brown, J.L.; Khonmee, J.; Norkaew, T.; Nganvongpanit, K.; Punyapornwithaya, V.; Angkawanish, T.; Somgird, C.; Thitaram, C. Management Factors Affecting Adrenal Glucocorticoid Activity of Tourist Camp Elephants in Thailand and Implications for Elephant Welfare. PLoS ONE 2019, 14, e0221537. [Google Scholar] [CrossRef]
- Ferin, M. Stress and the Reproductive Cycle. J. Clin. Endocrinol. Metab. 1999, 84, 1768–1774. [Google Scholar] [CrossRef]
- Fernandez-Novo, A.; Pérez-Garnelo, S.S.; Villagrá, A.; Pérez-Villalobos, N.; Astiz, S. The Effect of Stress on Reproduction and Reproductive Technologies in Beef Cattle-A Review. Animals 2020, 10, 2096. [Google Scholar] [CrossRef]
- Proctor, C.M.; Freeman, E.W.; Brown, J.L. Influence of Dominance Status on Adrenal Activity and Ovarian Cyclicity Status in Captive African Elephants. Zoo Biol. 2010, 29, 168–178. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Elephant | Age | Sampling Date | Parturition Date | Camp | Work Type | Reproductive State | Parity | BCS |
|---|---|---|---|---|---|---|---|---|
| E1 | 24 | June 2005 to March 2007 | 7 March 2007 | A | Saddle riding | Pregnant | 2 | 4 |
| E2 | 24 | August 2007 to May 2009 | 31 May 2009 | A | Saddle riding | Pregnant | 2 | 4.5 |
| E3 | 21 | December 2003 to Sep 2005 | 30 Sep 2005 | B | Saddle riding | Pregnant | 1 | 3.5 |
| E4 | 28 | June 2004 to August 2005 | 14 March 2006 | B | Saddle riding | Pregnant | 2 | 3.5 |
| E5 | 23 | August 2004 to March 2006 | 6 May 2004 | A | Saddle riding | Postpartum and cycling | 1 | 4.5 |
| E6 | 20 | November 2014 to August 2016 | - | C | Bareback riding | Cycling | 2 | 4 |
| E7 | 16 | November 2014 to August 2016 | - | C | Bareback riding | Cycling | Nulliparous | 3 |
| E8 | 8 | November 2014 to August 2016 | - | C | Bareback riding | Cycling | No data | 4 |
| E9 | 34 | November 2014 to August 2016 | - | C | Bareback riding | Cycling | Nulliparous | 4.5 |
| Status | Period | Progestagens (ng/mL) | Cortisol (ng/mL) | Prolactin (ng/mL) |
|---|---|---|---|---|
| Pregnant | 1st | 2.74 ± 0.24 a | 0.76 ± 0.08 a | 8.59 ± 1.76 a |
| 2nd | 3.40 ± 0.22 a | 0.91 ± 0.11 a | 31.15 ± 2.53 b | |
| 3rd | 3.09 ± 0.21 a | 0.62 ± 0.08 a | 36.47 ± 3.34 b | |
| Overall mean | 2.92 ± 0.11 | 0.81 ± 0.06 | 24.82 ± 1.64 | |
| Cycling | Follicular phase | 0.23 ± 0.05 a | 2.80 ± 0.77 a | 2.06 ± 0.15 a,* |
| Luteal phase | 2.54 ± 0.29 b | 6.52 ± 1.00 b | 1.54 ± 0.24 a,* | |
| Overall mean | 1.68 ± 0.19 | 3.65 ± 0.44 | 1.86 ± 0.08 * |
| Status | Elephant | Progestagens (ng/mL) | Cortisol (ng/mL) | Prolactin (ng/mL) | |
|---|---|---|---|---|---|
| Pregnant | E1 | 3.06 ± 0.21 b | 1.08 ± 0.13 b | 16.61 ± 2.77 a | |
| E2 | 3.81 ± 0.30 c | 0.54 ± 0.06 a | 20.07 ± 3.15 a | ||
| E3 | 2.86 ± 0.22 b | 0.73 ± 0.13 ab | 27.64 ± 3.84 b | ||
| E4 | 1.53 ± 0.09 a | 1.05 ± 0.14 b | 18.41 ± 2.81 a | ||
| Cycling | Follicular phase | Luteal phase | |||
| E5 | 0.01 ± 0.01 a | 0.39 ± 0.04 a | 0.63 ± 0.09 a | 20.95 ± 1.42 d | |
| E6 | 1.91 ± 0.16 c | 6.39 ± 1.15 c | 13.82 ± 1.32 c | 1.09 ± 0.08 a | |
| E7 | 0.39 ± 0.02 b | 1.41 ± 0.22 b | 1.66 ± 0.17 b | 2.90 ± 0.16 c | |
| E8 | 0.34 ± 0.06 b | 3.11 ± 0.56 bc | 2.07 ± 0.28 b | 1.47 ± 0.11 a | |
| E9 | 0.16 ± 0.03 ab | 1.09 ± 0.19 b | 1.61 ± 0.15 b | 1.93 ± 0.09 b | |
| Hormone | Pregnant | Cycling | ||
|---|---|---|---|---|
| r | p | r | p | |
| Progestagens and cortisol | −0.114 | 0.224 | 0.386 | 0.000 |
| Cortisol and prolactin * | −0.092 | 0.330 | −0.030 | 0.781 |
| Progestagens and prolactin * | 0.177 | 0.058 | −0.046 | 0.593 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Towiboon, P.; Saenphet, K.; Tayapiwattana, C.; Tangyuenyong, S.; Watanabe, G.; Mahasawangkul, S.; Brown, J.L.; Thitaram, C. Relationship among Serum Progestagens, Cortisol, and Prolactin in Pregnant and Cycling Asian Elephants in Thailand. Vet. Sci. 2022, 9, 244. https://doi.org/10.3390/vetsci9050244
Towiboon P, Saenphet K, Tayapiwattana C, Tangyuenyong S, Watanabe G, Mahasawangkul S, Brown JL, Thitaram C. Relationship among Serum Progestagens, Cortisol, and Prolactin in Pregnant and Cycling Asian Elephants in Thailand. Veterinary Sciences. 2022; 9(5):244. https://doi.org/10.3390/vetsci9050244
Chicago/Turabian StyleTowiboon, Patcharapa, Kanokporn Saenphet, Chatchai Tayapiwattana, Siriwan Tangyuenyong, Gen Watanabe, Sittidet Mahasawangkul, Janine L. Brown, and Chatchote Thitaram. 2022. "Relationship among Serum Progestagens, Cortisol, and Prolactin in Pregnant and Cycling Asian Elephants in Thailand" Veterinary Sciences 9, no. 5: 244. https://doi.org/10.3390/vetsci9050244
APA StyleTowiboon, P., Saenphet, K., Tayapiwattana, C., Tangyuenyong, S., Watanabe, G., Mahasawangkul, S., Brown, J. L., & Thitaram, C. (2022). Relationship among Serum Progestagens, Cortisol, and Prolactin in Pregnant and Cycling Asian Elephants in Thailand. Veterinary Sciences, 9(5), 244. https://doi.org/10.3390/vetsci9050244