
Copper Sulfate Combined with Photodynamic Therapy Enhances Antifungal Effect by Downregulating AIF1


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains, Growth Conditions, and Agents
2.2. Measurement of Inhibition of C. albicans by HLA-ES
2.3. Inhibition of C. albicans by Copper Sulfate
2.4. Evaluation of the Inhibitory Effect of HE-PDT Combined with Copper Sulfate on C. albicans
2.5. Measurement of Intracellular ROS, H2O2, O2−, and ATP
2.6. Flow Cytometry for the Detection of ROS, Apoptosis, and Mitochondrial Membrane Potential (MMP) Changes
2.7. q-PCR to Verify AIF1 Expression
2.8. Data Analysis
3. Results
3.1. Copper Sulfate Increased the Suppressive Effect of HE-PDT on C. albicans
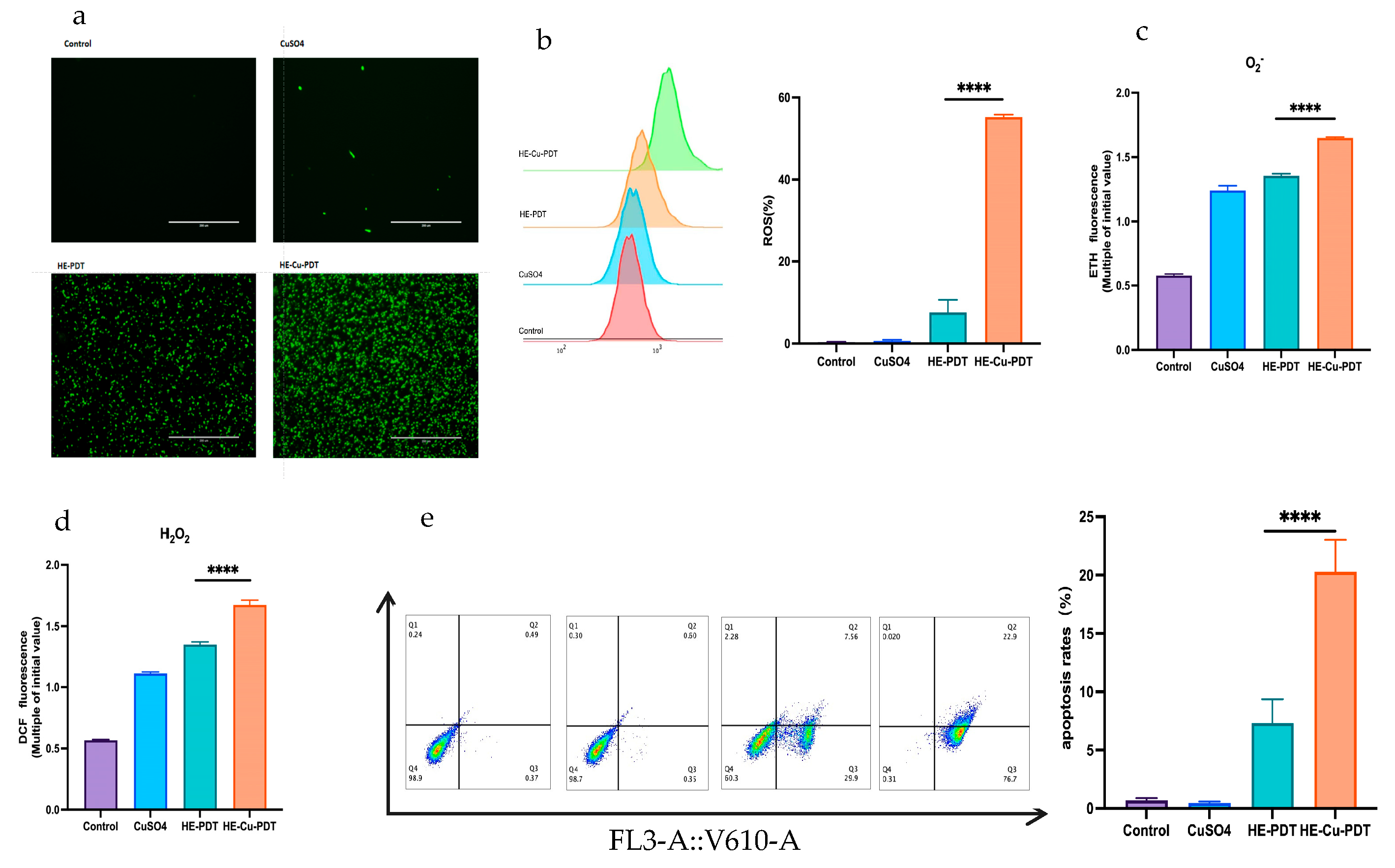
3.2. Copper Sulfate Enhances Oxidative Stress and Apoptosis
3.3. Effect of Copper Sulfate on Mitochondria
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action. WHO. Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 6 February 2024).
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic therapy—Mechanisms, photosensitizers and combinations. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef]
- Morrow, D.I.; McCarron, P.A.; Woolfson, A.D.; Juzenas, P.; Juzeniene, A.; Iani, V.; Moan, J.; Donnelly, R.F. Hexyl aminolaevulinate is a more effective topical photosensitiser precursor than methyl aminolaevulinate and 5-aminolaevulinic acids when applied in equimolar doses. J. Pharm. Sci. 2010, 99, 3486–3498. [Google Scholar] [CrossRef]
- Yang, L.; Wu, L.; Wu, D.; Shi, D.; Wang, T.; Zhu, X. Mechanism of transdermal permeation promotion of lipophilic drugs by ethosomes. Int. J. Nanomed. 2017, 12, 3357–3364. [Google Scholar] [CrossRef]
- Wang, T.; Wu, L.; Wang, Y.; Song, J.; Zhang, F.; Zhu, X. Hexyl-aminolevulinate ethosome–mediated photodynamic therapy against acne: In vitro and in vivo analyses. Drug Deliv. Transl. Res. 2022, 12, 325–332. [Google Scholar] [CrossRef]
- Wang, Y.; Song, J.; Zhang, F.; Zeng, K.; Zhu, X. Antifungal Photodynamic Activity of Hexyl-Aminolevulinate Ethosomes Against Candida albicans Biofilm. Front. Microbiol. 2020, 11, 2052. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Long, W.; Zhang, F.; Zhang, M.; Zeng, K.; Zhu, X. Hexyl-Aminolevulinate Ethosomes: A Novel Antibiofilm Agent Targeting Zinc Homeostasis in Candida albicans. Microbiol. Spectr. 2022, 10, e0243822. [Google Scholar] [CrossRef] [PubMed]
- Nunes, I.P.; Crugeira, P.J.; Sampaio, F.J.; de Oliveira, S.C.; Azevedo, J.M.; Santos, C.L.; Soares, L.G.; Samuel, I.D.; Persheyev, S.; de Ameida, P.F.; et al. Evaluation of dual application of photodynamic therapy—PDT in Candida albicans. Photodiagn. Photodyn. Ther. 2023, 42, 103327. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Wu, H.; Wang, H.; Tang, B.; Xu, J.F.; Zhang, X. A Self-Degradable Supramolecular Photosensitizer with High Photodynamic Therapeutic Efficiency and Improved Safety. Angew. Chem. Int. Ed. Engl. 2021, 60, 706–710. [Google Scholar] [CrossRef]
- Grass, G.; Rensing, C.; Solioz, M. Metallic copper as an antimicrobial surface. Appl. Environ. Microbiol. 2011, 77, 1541–1547. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, L.; Qin, Y.; Zhang, L.; Jiang, Y.; Cao, Y. Regulation and control of transcription factor Cup2 on Cu2+ metabolism and oxidative stress in Candida albicans. J. Pharm. Pract. 2017, 35, 224–228. Available online: https://kns.cnki.net/kcms2/article/abstract?v=NR7yonmY8oO2DgRBy2u02VjNKaY4Q8W6JGN0GnE0eDQSat74lKZycbXA81dvafBr-2lmIjpK321KsfvSR1dpGd3OZDapRx73R-PXEz78f6sQElwZcx24sXDiiCu9Rq66hV_yMA9Q84vXebZkIYyL1g==&uniplatform=NZKPT&language=CHS (accessed on 6 February 2024).
- Chudzik, B.; Tracz, I.B.; Czernel, G.; Fiołka, M.J.; Borsuk, G.; Gagoś, M. Amphotericin B–copper(II) complex as a potential agent with higher antifungal activity against Candida albicans. Eur. J. Pharm. Sci. 2013, 49, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Rigo, G.V.; Cardoso, F.G.; Devereux, M.; McCann, M.; Macedo, A.J.; Santos, A.L.S.; Tasca, T. Antimicrobial and Antibiofilm Activities of Copper(II)-1,10-phenanthroline-5,6-pione Against Commensal Bacteria and Fungi Responsible for Vaginal Microbiota Dysbiosis. Curr. Microbiol. 2023, 80, 383. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Li, Y.; Ding, C. The Role of Copper Homeostasis at the Host-Pathogen Axis: From Bacteria to Fungi. Int. J. Mol. Sci. 2019, 20, 175. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Kang, J.; Zheng, R. Oxidative stress is involved in inhibition of copper on histone acetylation in cells. Chem. Interact. 2005, 151, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.J., Jr.; Gutterman, D.D.; Rios, C.D.; Heistad, D.D.; Davidson, B.L. Superoxide production in vascular smooth muscle contributes to oxidative stress and impaired relaxation in atherosclerosis. Circ. Res. 1998, 82, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Somers, M.J.; Mavromatis, K.; Galis, Z.S.; Harrison, D.G. Vascular superoxide production and vasomotor function in hypertension induced by deoxycorticosterone acetate–salt. Circ. 2000, 101, 1722–1728. [Google Scholar] [CrossRef] [PubMed]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2′,7′-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef]
- Hamuro, J.; Yamashita, T.; Otsuki, Y.; Hiramoto, N.; Adachi, M.; Miyatani, T.; Tanaka, H.; Ueno, M.; Kinoshita, S.; Sotozono, C. Spatiotemporal Coordination of RPE Cell Quality by Extracellular Vesicle miR-494-3p Via Competitive Interplays with SIRT3 or PTEN. Investig. Opthalmol. Vis. Sci. 2023, 64, 9. [Google Scholar] [CrossRef]
- Kessel, D. Apoptosis, Paraptosis and Autophagy: Death and Survival Pathways Associated with Photodynamic Therapy. Photochem. Photobiol. 2019, 95, 119–125. [Google Scholar] [CrossRef]
- Gheewala, T.; Skwor, T.; Munirathinam, G. Photosensitizers in prostate cancer therapy. Oncotarget 2017, 8, 30524–30538. [Google Scholar] [CrossRef]
- Calixto, G.M.F.; Bernegossi, J.; De Freitas, L.M.; Fontana, C.R.; Chorilli, M.; Grumezescu, A.M. Nanotechnology-Based Drug Delivery Systems for Photodynamic Therapy of Cancer: A Review. Molecules 2016, 21, 342. [Google Scholar] [CrossRef]
- Yaqoob, M.D.; Xu, L.; Li, C.; Leong, M.M.L.; Xu, D.D. Targeting mitochondria for cancer photodynamic therapy. Photodiagn. Photodyn. Ther. 2022, 38, 102830. [Google Scholar] [CrossRef]
- Chen, Q. Rational application of antifungal drugs in the treatment of superficial cutaneous fungal diseases. China Med. Guide 2012, 10, 87–88. [Google Scholar] [CrossRef]
- d’Enfert, C.; Kaune, A.K.; Alaban, L.R.; Chakraborty, S.; Cole, N.; Delavy, M.; Kosmala, D.; Marsaux, B.; Frois-Martins, R.; Morelli, M.; et al. The impact of the Fungus-Host-Microbiota interplay upon Candida albicans infections: Current knowledge and new perspectives. FEMS Microbiol. Rev. 2021, 45, fuaa060. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef]
- Peacock, J.E.; Morris, A.J.; Tanner, D.C.; Nguyen, M.L.; Snydman, D.R.; Wagener, M.M.; Rinaldi, M.G.; Yu, V.L. The changing face of candidemia: Emergence of non-Candida albicans species and antifungal resistance. Am. J. Med. 1996, 100, 617–623. [Google Scholar] [CrossRef]
- Chowdhary, A.; Sharma, C.; Meis, J.F. Candida auris: A rapidly emerging cause of hospital-acquired multidrug-resistant fungal infections globally. PLoS Pathog. 2017, 13, e1006290. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Alastruey-Izquierdo, A.; Berman, J.; Bicanic, T.; Bignell, E.M.; Bowyer, P.; Bromley, M.; Brüggemann, R.; Garber, G.; Cornely, O.A.; et al. Tackling the emerging threat of antifungal resistance to human health. Nat. Rev. Microbiol. 2022, 20, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Smijs, T.G.; van der Haas, R.N.; Lugtenburg, J.; Liu, Y.; de Jong, R.L.; Schuitmaker, H.J. Photodynamic treatment of the dermatophyte Trichophyton rubrum and its microconidia with porphyrin photosensitizers. Photochem. Photobiol. 2004, 80, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Smijs, T.G.M.; Bouwstra, J.A.; Schuitmaker, H.J.; Talebi, M.; Pavel, S. A novel ex vivo skin model to study the susceptibility of the dermatophyte Trichophyton rubrum to photodynamic treatment in different growth phases. J. Antimicrob. Chemother. 2007, 59, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Cerdeira, C.; Martínez-Herrera, E.; Fabbrocini, G.; Sanchez-Blanco, B.; López-Barcenas, A.; El-Samahy, M.; Juárez-Durán, E.R.; González-Cespón, J.L. New Applications of Photodynamic Therapy in the Management of Candidiasis. J. Fungi 2021, 7, 1025. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Lu, L.; Chen, Y.; Yin, Q. Inhibitory effect of 5-aminolevulinic acid photodynamic therapy on Candida albicans. Hainan Med. 2013, 24, 1405–1407. Available online: https://kns.cnki.net/kcms2/article/abstract?v=NR7yonmY8oN-eHp6m1r25Ci45TJN13Qe6EV3POLHB4HINRDuks1kLUKwwOINUpgOyt44FswTfwtUHUE4XIxif5yslluGFhnjTxWddB-vAPmh4w7ypAy-wHaJmmi9XMgy0ZxMra_zOGk=&uniplatform=NZKPT&language=CHS (accessed on 6 February 2024).
- Shi, H.; Li, J.; Peng, C.; Xu, B.; Sun, H. The inhibitory activity of 5-aminolevulinic acid photodynamic therapy (ALA-PDT) on Candida albicans biofilms. Photodiagn. Photodyn. Ther. 2021, 34, 102271. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.-X.; Liu, C.-G.; Zhang, J.-T.; Zheng, X.; Yang, D.-Y.; Kankala, R.K.; Wang, S.-B.; Chen, A.-Z. Biodegradable Quantum Composites for Synergistic Photothermal Therapy and Copper-Enhanced Chemotherapy. ACS Appl. Mater. Interfaces 2020, 12, 47289–47298. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wu, H.; Lim, W.Q.; Phua, S.Z.F.; Xu, P.; Chen, Q.; Guo, Z.; Zhao, Y. A Mesoporous Nanoenzyme Derived from Metal–Organic Frameworks with Endogenous Oxygen Generation to Alleviate Tumor Hypoxia for Significantly Enhanced Photodynamic Therapy. Adv. Mater. 2019, 31, e1901893. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Chen, M.; Bhattarai, P.; Hameed, S.; Tang, Y.; Dai, Z. Complementing Cancer Photodynamic Therapy with Ferroptosis through Iron Oxide Loaded Porphyrin-Grafted Lipid Nanoparticles. ACS Nano 2021, 15, 20164–20180. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Chen, X.; Cheng, Y.; Lu, X.; Meng, J.; Chen, Z.; Li, M.; Lin, C.; Wang, Y.; Yang, J. CuWO4 Nanodots for NIR-Induced Photodynamic and Chemodynamic Synergistic Therapy. ACS Appl. Mater. Interfaces 2021, 13, 22150–22158. [Google Scholar] [CrossRef]
- Nain, A.; Tseng, Y.-T.; Wei, S.-C.; Periasamy, A.P.; Huang, C.-C.; Tseng, F.-G.; Chang, H.-T. Capping 1,3-propanedithiol to boost the antibacterial activity of protein-templated copper nanoclusters. J. Hazard. Mater. 2020, 389, 121821. [Google Scholar] [CrossRef]
- Lin, L.S.; Song, J.; Song, L.; Ke, K.; Liu, Y.; Zhou, Z.; Shen, Z.; Li, J.; Yang, Z.; Tang, W.; et al. Simultaneous Fenton-like Ion Delivery and Glutathione Depletion by MnO2—Based Nanoagent to Enhance Chemodynamic Therapy. Angew. Chem. Int. Ed. Engl. 2018, 57, 4902–4906. [Google Scholar] [CrossRef]
- Chen, Z.; Wu, Y.; Yao, Z.; Su, J.; Wang, Z.; Xia, H.; Liu, S. 2D Copper(II) Metalated Metal–Organic Framework Nanocomplexes for Dual-enhanced Photodynamic Therapy and Amplified Antitumor Immunity. ACS Appl. Mater. Interfaces 2022, 14, 44199–44210. [Google Scholar] [CrossRef]
- Gaetke, L.M.; Chow, C.K. Copper toxicity, oxidative stress, and antioxidant nutrients. Toxicology 2003, 189, 147–163. [Google Scholar] [CrossRef] [PubMed]
- Theophanides, T.; Anastassopoulou, J. Copper and carcinogenesis. Crit. Rev. Oncol. Hematol. 2002, 42, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Yuan, S.; Ma, S.; Jin, M.; Liu, D.; Cheng, D.; Liu, X.; Gan, Y.; Wang, Y. Production of Abundant Hydroxyl Radicals from Oxygenation of Subsurface Sediments. Environ. Sci. Technol. 2016, 50, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Pan, J.; Deng, J.; Zhang, Q.; Yan, Q. A Review of the Application of Advanced Oxidation Technology of Hydroxyl Radicals. J. Guangdong Univ. Technol. 2019, 36, 70–77+85. [Google Scholar] [CrossRef]
- Herrmann, J.M.; Riemer, J. Apoptosis inducing factor and mitochondrial NADH dehydrogenases: Redox-controlled gear boxes to switch between mitochondrial biogenesis and cell death. Biol. Chem. 2020, 402, 289–297. [Google Scholar] [CrossRef]
- Liang, M.; Huang, L.; Liu, Q.; Long, R.; Deng, Y. Potential ferroptosis pathway in pathogenic fungi: Reported functions and future perspectives. Microbiol. Bull. 2021, 48, 4387–4397. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; An, Q.; Wang, Y.; Ye, S.; Zhu, X. Copper Sulfate Combined with Photodynamic Therapy Enhances Antifungal Effect by Downregulating AIF1. J. Fungi 2024, 10, 213. https://doi.org/10.3390/jof10030213
Zhang M, An Q, Wang Y, Ye S, Zhu X. Copper Sulfate Combined with Photodynamic Therapy Enhances Antifungal Effect by Downregulating AIF1. Journal of Fungi. 2024; 10(3):213. https://doi.org/10.3390/jof10030213
Chicago/Turabian StyleZhang, Meimei, Qiyuan An, Yingzhe Wang, Shigan Ye, and Xiaoliang Zhu. 2024. "Copper Sulfate Combined with Photodynamic Therapy Enhances Antifungal Effect by Downregulating AIF1" Journal of Fungi 10, no. 3: 213. https://doi.org/10.3390/jof10030213
APA StyleZhang, M., An, Q., Wang, Y., Ye, S., & Zhu, X. (2024). Copper Sulfate Combined with Photodynamic Therapy Enhances Antifungal Effect by Downregulating AIF1. Journal of Fungi, 10(3), 213. https://doi.org/10.3390/jof10030213