

Research on the Isolation of Endophytic Fungi from Papaya and the Prevention of Colletotrichum gloeosporioides
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.1.1. Plant Materials and Pathogens
2.1.2. Drugs and Chemicals
2.2. Methods
2.2.1. Isolation, Purification, and Identification of Strains
2.2.2. Strain Fermentation and Sample Preparation
2.2.3. Screening of the Strains with Antioxidant Activity and Determination of the Half-Maximal Inhibitory Concentration (IC50)
2.3. Selection of Strains of Endophytic Fungi from Papaya with Antifungal Activity
2.4. Detection of the Control Effect of Endophytic Fungi from Papaya on Colletotrichum gloeosporioides
2.5. Determination of Defense Enzyme Activity and Related Indexes in the Fruit after Treatment with Endophytic Fungi
2.6. Construction of a Phylogenetic Tree of the Active Strains
2.7. Statistical Analysis
3. Results
3.1. Isolation of Endophytes
3.2. Identification
3.3. Screening of Antioxidant Strains
3.3.1. Radical Scavenging Assays
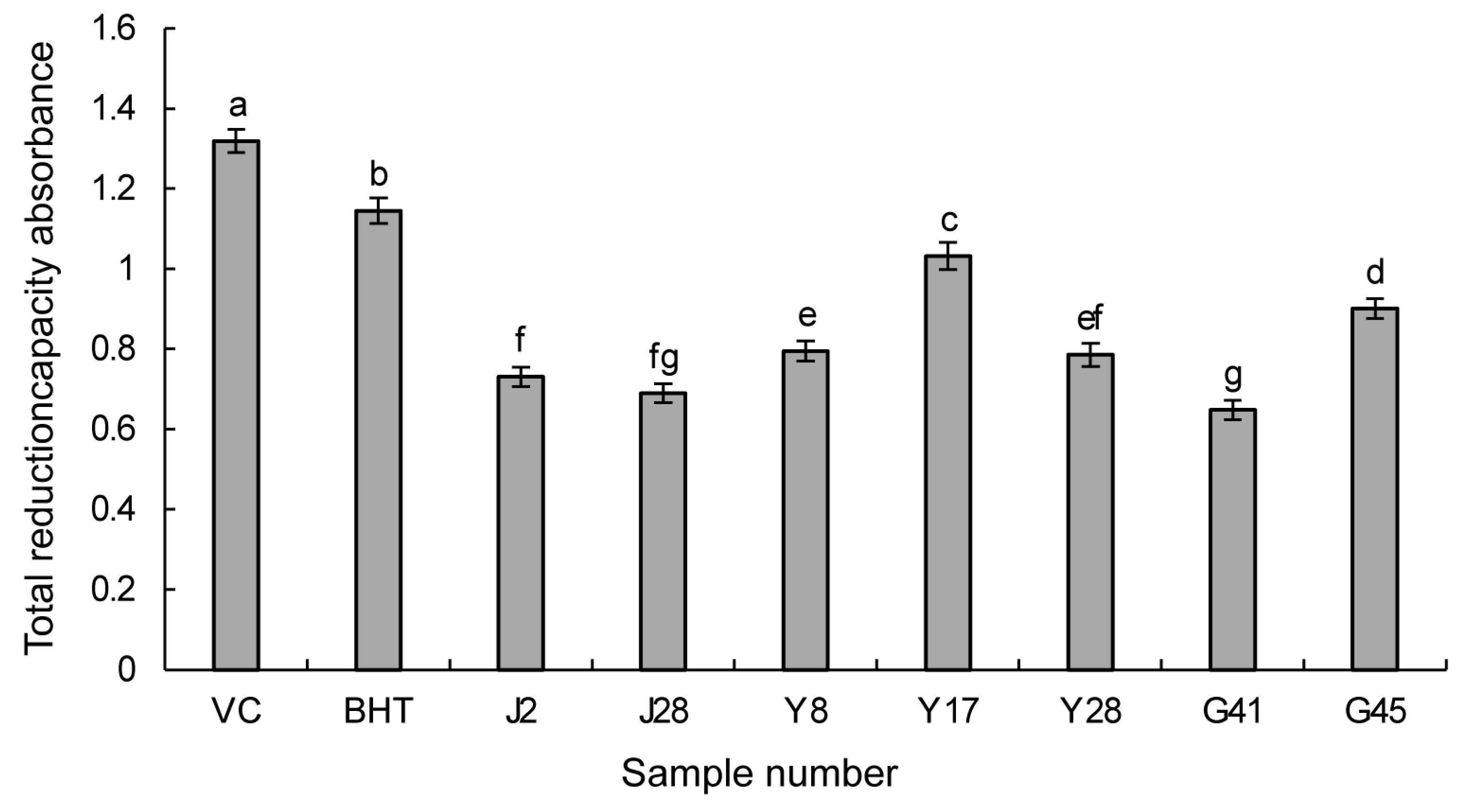
3.3.2. Reducing Power Assay
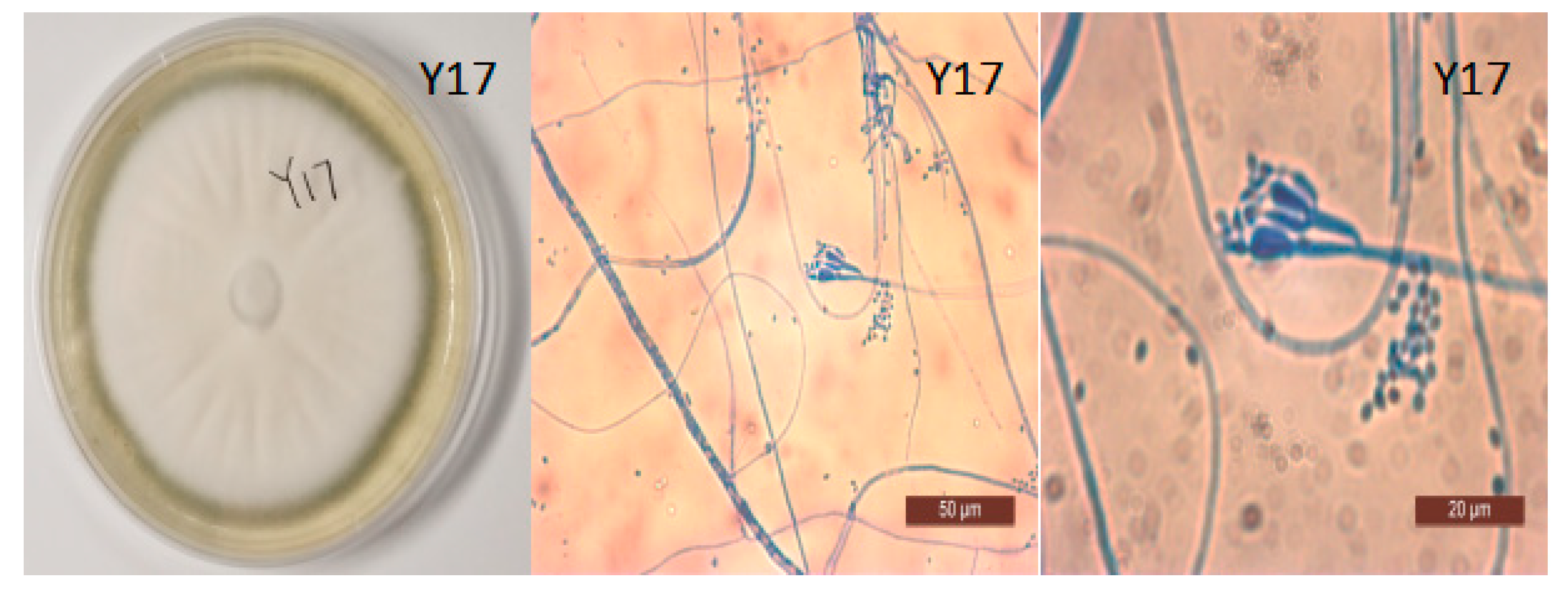
3.4. Morphological Identification of Y17 and the Construction of a Phylogenetic Tree
3.5. Screening of Strains with Antifungal Activity
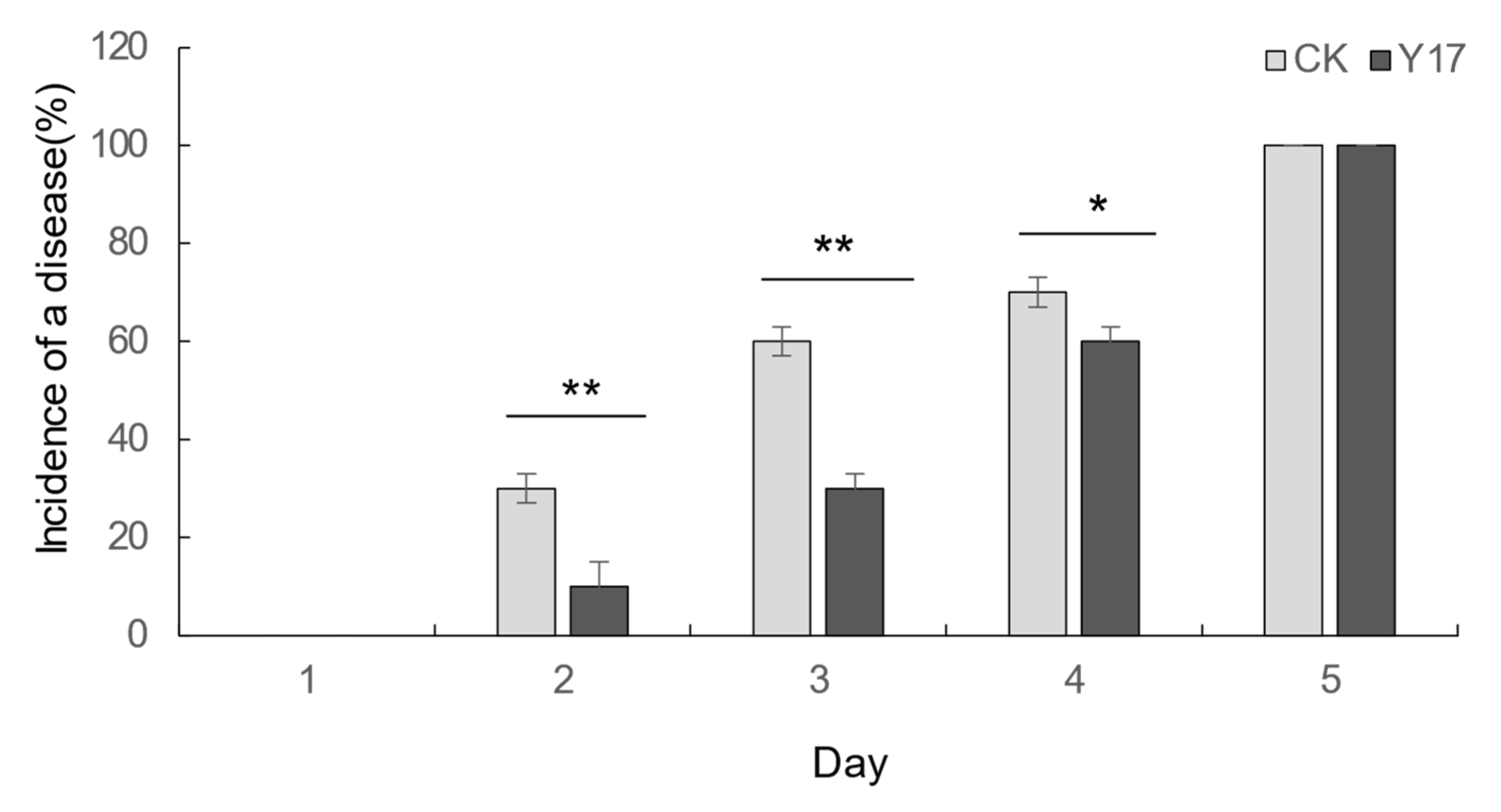
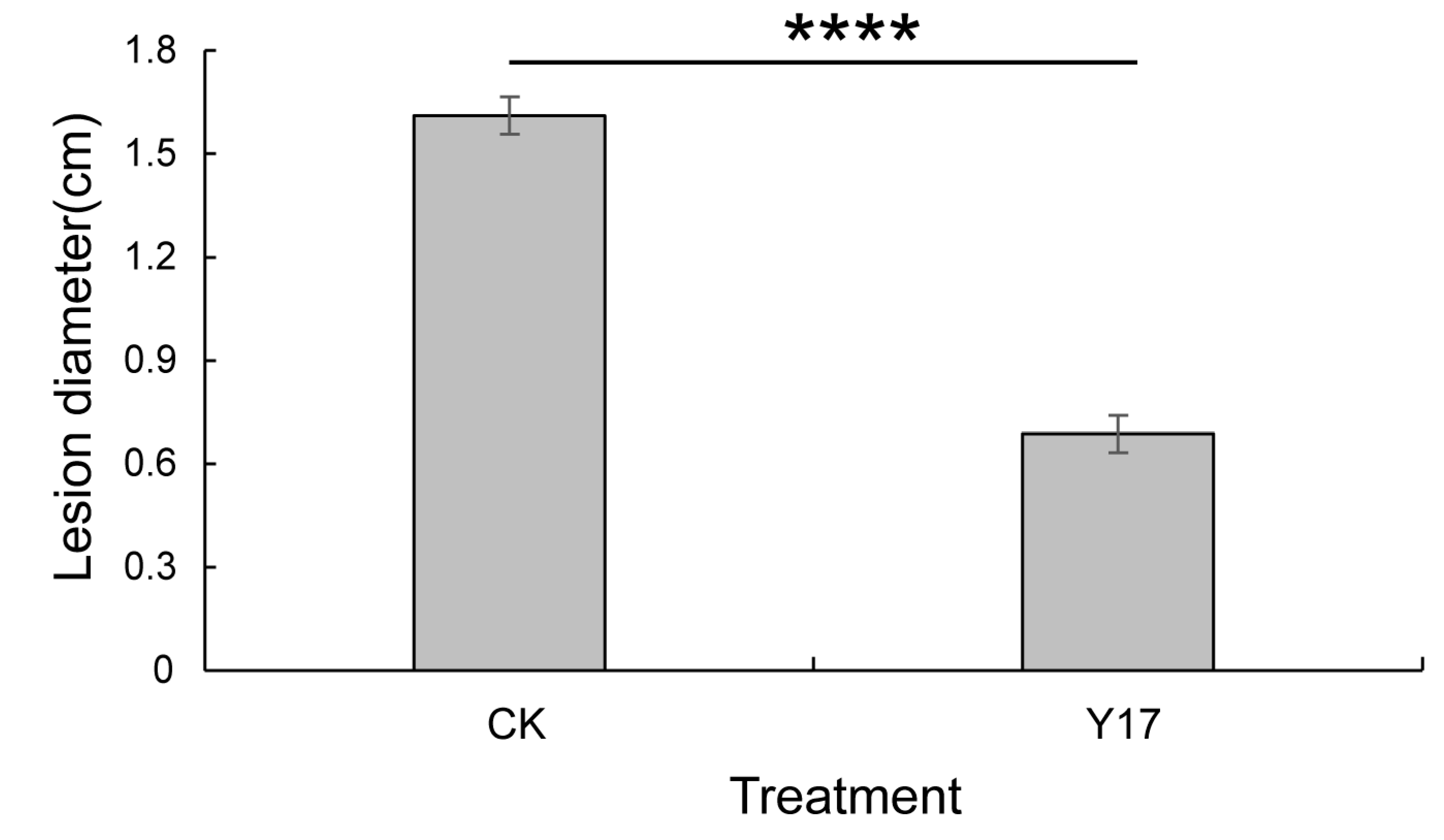
3.6. Detection and Control Effects of the Active Antifungal Strain Y17 on Colletotrichum gloeosporioides
3.7. Effect of the Active Antifungal Strain Y17 on the Antioxidant System of Papaya Fruit
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kataki, M.S.; Kakoti, B.B.; Bhuyan, B.; Rajkumari, A.; Rajak, P. Garden rue inhibits the arachidonic acid pathway, scavenges free radicals, and elevates FRAP: Role in inflammation. Chin. J. Nat. Med. 2014, 12, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Wang, L.H. Research progress on the diversity of endophytic bacteria and their secondary metabolites. Chem. Biol. Eng. 2012, 29, 3. [Google Scholar] [CrossRef]
- Tan, X.M.; Zhou, Y.Q.; Chen, J.; Gou, S.X. Advances in Research on Diversity of Endophytic Fungi from Medicinal Plants. Chin. Pharm. J. 2015, 50, 1563–1580. [Google Scholar] [CrossRef]
- Leneveu-Jenvrin, C.; Charles, F.; Barba, F.J.; Remize, F. Role of biological control agents and physical treatments in maintaining the quality of fresh and minimally-processed fruit and vegetables. Crit. Rev. Food Sci. Nutr. 2020, 60, 2837–2855. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Luo, H.C.; Zhang, X.S. Identification of Endophytic Funji in Bupleurum and Screening of Antimicrobial Activities. Biotechnol. Bull. 2015, 31, 165–170. [Google Scholar] [CrossRef]
- Chen, J.; Xu, Z.; Zhang, X.; Li, Y.P.; Liu, C.S. Diversity and isolation parameters of endophytes from Glycyrrhiza uralensis of different habitats. Acta Pharm. Sin. 2019, 54, 373–379. [Google Scholar] [CrossRef]
- Khuzhaev, V.U.; Aripova, S.F. Pseudocarpaine from Carica Papaya. Chem. Nat. Compd. 2000, 36, 418. [Google Scholar] [CrossRef]
- Romasi, E.F.; Karina, J.K.; Parhusip, A.J.N. Antibacterial activity of papaya leaf extracts against pathogenic bacteria. MAKARA Technol. Ser. 2012, 15, 2. [Google Scholar] [CrossRef]
- Krishna, K.L.; Paridhavi, M.; Patel, J.A. Review on nutritional, medicinal and pharmacological properties of papaya (Carica papaya Linn.). Nat. Prod. Radiance 2008, 7, 364–373. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870. [Google Scholar] [CrossRef] [PubMed]
- Ge, F.; Tang, Y.; Gong, Q.; Ma, Q.S.; Yang, L.J. Comparison on antimicrobial activity of 40 endophytic fungi strains from Ginkgo biloba and preliminary analysis on ingredients with antimicrobial activity. Chin. Tradit. Herb. Drugs 2016, 47, 1554–1559. [Google Scholar] [CrossRef]
- Wang, X.J.; Min, C.L.; Han, P.L.; Qian, J. Identification and Fermentation Optimization of an Endophytic Fungus BLH34 with Antibacterial Activity from Macleaya cordata. J. Chin. Med. Mater. 2016, 39, 1728–1733. [Google Scholar] [CrossRef]
- Chen, X.X.; Fan, M.M.; Zhang, H.; Luo, X.L.; Zhao, C.L.; Wu, J.R.; Li, J.; Zhao, P. Identification and Antifungal Activity of An Endophytic Fungus from Vaccinium dunalianum. J. Southwest For. Univ. (Nat. Sci.) 2018, 38, 187–192. [Google Scholar]
- Farhat, H.; Urooj, F.; Tariq, A.; Sultana, V.; Ansari, M.; Ahmad, V.U.; Ehteshamul-Haque, S. Evaluation of antimicrobial potential of endophytic fungi associated with healthy plants and characterization of compounds produced by endophytic Cephalosporium and Fusarium solani. Biocatal. Agric. Biotechnol. 2019, 18, 101043. [Google Scholar] [CrossRef]
- Ren, A.Z.; Gao, Y.B. Endophytic fungi—A kind of resource microorganisms with broad application prospects. Microbiol. China 2001, 06, 90–93. [Google Scholar] [CrossRef]
- Du, Y.X.; Feng, Y.; Zhang, Q.L.; Tian, S.H.; Liu, K.Q. Biological characteristics of Colletorichum gloeosprioides Penz.causing anthracnose of Carica papaya L. J. Zhongkai Univ. Agric. Technol. 2006, 19, 4–7. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebensm. Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ginting, R.C.B.; Sukarno, N.; Widyastuti, U.; Darusman, L.K.; Kanaya, S. Diversity of Endophytic Fungi from Red Ginger (Zingiber officinale Rosc.) Plant and Their Inhibitory Effect to Fusarium oxysporum Plant Pathogenic Fungi. HAYATI J. Biosci. 2013, 20, 127–137. [Google Scholar] [CrossRef]
- Gunde, M.C.; Amnerkar, N.D. Nutritional, medicinal and pharmacological propertiesofpapaya (Carica papaya·Linn.): A review. J. Innov. Pharm. Biol. Sci. 2016, 3, 162–169. [Google Scholar]
- Huang, W.Y.; Cai, Y.Z.; Hyde, K.D.; Corke, H.; Sun, M. Biodiversity of endophytic fungi associated with 29 traditional. Chinese medicinal plants. Fungal Divers. 2008, 33, 61–75. [Google Scholar]
- Huang, W.Y.; Cai, Y.Z.; Xing, J.; Corke, H.; Sun, M. A potential antioxidant resource: Endophytic fungi from medicinalplants. Econ. Bot. 2007, 61, 14–30. [Google Scholar] [CrossRef]
- Liu, X.J.; Yang, Y.; Huang, S.M. The species and distribution of latent fungi of papaya fruit in Hainan province. Acta Phytopathol. Sin. 1994, 24, 313–317. [Google Scholar] [CrossRef]
- Shi, J.Y.; Liu, A.Y.; Feng, S.J.; Li, X.P.; Chen, W.X. Identification and biocontrol effect of endophytic bacterial strain MG-Y2 isolated from papaya. J. Fruit Sci. 2007, 24, 810–814. [Google Scholar] [CrossRef]
- Shi, J.Y.; Liu, A.Y.; Li, X.P.; Chen, W.X. Identification and antagonistic activities of an endophytic fungal MGP3 isolated from papaya fruit. Acta Microbiol. Sin. 2011, 51, 1240–1247. [Google Scholar] [CrossRef]
- Fang, Z.D. Research Methods of Plant Diseases, 3rd ed.; China Agriculture Press: Beijing, China, 1998; pp. 124–125. ISBN 9787109052550. [Google Scholar]
- Venugopalan, A.; Srivastava, S. Endophytes as in vitro production platforms of high value plant secondary metabolites. Biotechnol. Adv. 2015, 33, 873–887. [Google Scholar] [CrossRef]
- Wu, X.H.; Li, W.C.; Qin, L.P. Composition and Diversity analysis of Endophytic Fungi from Different Parts of Pepper in Tianmu Mountain. J. Plant Resour. Environ. 2012, 21, 107–113. [Google Scholar] [CrossRef]
- Landeweert, R.; Leeflang, P.; Kuyper, T.W.; Hoffland, E.; Rosling, A.; Wernars, K.; Smit, E. Molecular identification of ectomycorrhizal mycelium in soil horizons. Appl. Environ. Microbiol. 2003, 69, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Boffa, L.; Binello, A.; Boscaro, V.; Gallicchio, M.; Amisano, G.; Fornasero, S.; Cravotto, G. Commiphora myrrha (Nees) Engl. extracts: Evaluation of antioxidant and antiproliferative activity and their ability to reduce microbial growth on fresh-cut salad. Int. J. Food Sci. Technol. 2016, 51, 625–632. [Google Scholar] [CrossRef]
- Zeng, P.Y.; Wu, J.G.; Liao, L.M.; Chen, T.-Q.; Wu, J.Z.; Wong, K.H. In vitro antioxidant activities of endophytic fungi isolated from the liverwort Scapania verrucosa. Genet. Mol. Res. 2011, 10, 3169–3179. [Google Scholar] [CrossRef]
- Hichri, F.; Omri Hichri, A.; Maha, M.; Saad Mana Hossan, A.; Flamini, G.; Ben Jannet, H. Chemical Composition, Antibacterial, Antioxidant and in Vitro Antidiabetic Activities of Essential Oils from Eruca vesicaria. Chem. Biodivers. 2019, 16, e1900183. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Y.; Yao, X.H.; Duan, M.H.; Wei, F.Y.; Wu, G.H.; Li, L. Variat ion of essential oil content and antioxidant activity of Lonicera species in different sites of China. Ind. Crops Prod. 2015, 77, 772–779. [Google Scholar] [CrossRef]
- Zhang, X.G.; Tang, P.; Liu, Y.J.; Zhao, P.; Chen, X.Q. Antioxidant activities of endophytic extracts from six medicinal plants. Mod. Food Technol. 2016, 32, 66–74. [Google Scholar] [CrossRef]
- Capdeville, G.; Souza, M.T.; Santos, J.R.P.; Miranda, S.P.; Caetano, A.R.; Torres, F.A.G. Selection and testing of epiphytic yeasts to control anthacnose in post-harvest of papaya fruit. Sci. Hortic. 2006, 111, 179–185. [Google Scholar] [CrossRef]
- Li, Z.H.; Ling, J.F.; Zeng, H.C. Antifungal Activity of Six Soil Streptomyces Strains and Their Control Effects Against Banana Postharvest Anthracnose. Trop. Agric. Sci. 2006, 26, 35–37+55. [Google Scholar]
- Yu, C.L.; Meng, L.L.; Lian, S.; Li, B.H.; Liang, W.X.; Wang, C.X. Butylated hydroxytoluene control effect on apple fruit ring spot and the influence of defense enzyme activity. Plant Physiol. J. 2018, 54, 819–826. [Google Scholar] [CrossRef]
- Zhang, X.; Mo, Y.; Zhou, L.; Wang, Y.N.; Wang, Z.C.; Zhao, B.S. Induction of hsp70, hsp90, and catalase activity in planarian Dugesia japonica exposed to cadmium. Toxicol. Ind. Health 2016, 32, 1373–1380. [Google Scholar] [CrossRef]
- Peng, J.; Wang, D.Y.; Xu, C.M.; Chen, L.P.; Deng, F.; Zhang, F.X. Ammonium Molybdate Method for Detecting the Activities of Rice Catalase. Chin. Agric. Sci. Bull. 2009, 25, 61–64. [Google Scholar]
- Giese, E.C.; Gascon, J.; Anzelmo, G.; Barbosa, A.M.; da Cunha, M.A.; Dekker, R.F. Free-radical scavenging properties and antioxidant activities of botryosphaeran and some other β-D-glucans. Int. J. Biol. Macromol. 2015, 72, 125–130. [Google Scholar] [CrossRef]
- Chen, S.T.; Liou, S.Y.; Wu, H.C.; Tsai, F.J.; Tsai, C.H.; Huang, C.Y.; Chang, Y.L. New Analytical Method for Investigating the Antioxidant Power of Food Extracts on the Basis of Their Electron-Donating Ability: Comparison to the Ferric Reducing/Antioxidant Power (FRAP) Assay. J. Agric. Food Chem. 2010, 58, 8477–8480. [Google Scholar] [CrossRef]
- Zeng, W.; Qin, W.; Tian, W.; Xue, Y.; Wang, G.; Liu, S. Antioxidant activity in vitro of endophytic fungi from myricaria laxiflora, a riparian plant with strong tolerance ability of flooding. J. Pure. Appl. Microbio. 2015, 9, 87–95. [Google Scholar]
- Wang, Y.; Li, Y.; Li, S.; Li, Q.Y.; Fan, W.G.; Kiatoukosin, L. Extracellular polysaccharides of endophytic fungus Alternaria tenuissima F1 from Angelica sinensis: Production conditions, purification, and antioxidant properties. Int. J. Biol. Macromol. 2019, 133, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Nisa, H.; Kamili, A.N.; Nawchoo, I.A.; Shafi, S.; Shameem, N.; Bandh, S.A. Fungal endophytesas prolific source of phytochemicals and other bioactive natural products: A review. Microb. Pathog. 2015, 82, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.S.; Tian, H.L. Function and study of catalase. Food Sci. Technol. 2007, 32, 8–11. [Google Scholar]
- He, X.L. Protective enzyme system in plants. Mod. Agric. Sci. Technol. 2010, 10, 37–38. [Google Scholar] [CrossRef]
- Wang, W.L.; Wang, Z.; Wang, J.Y. Optimization of methods for determination of plant peroxidase activity. Lab. Res. Explor. 2010, 29, 21–23. [Google Scholar] [CrossRef]
- Qin, P.; Liu, F.H.; Liang, X.N. Superoxide Dismutase and Plant Resistance to the Environmental Stress. Heilongjiang Agric. Sci. 2002, 01, 31–34. [Google Scholar] [CrossRef]
- Sun, H.P.; Li, L.; Wang, X.; Wu, S.; Wang, X.F. Ascorbate–glutathione cycle of mitochondria in osmoprimed soybean cotyledons in response to imbibitional chilling injury. J. Plant Physiol. 2010, 168, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.L.; Song, S.Q.; Lan, Q.Y. Possible Involvement of Enzymatic and Non-enzymatic Antioxidant System in Acquisition of Desiccation Tolerance of Maize Embryos. Acta Bot. Yunnanica 2009, 31, 253–259. [Google Scholar] [CrossRef]
- Zhao, X.; Hou, Z.B.; Dong, K.H.; Yang, W.D.; Zhu, H.S.; Liang, P.F. Effects of water stress and rehydration on enzymatic defense system of Branchia Dauri. Acta Agron. Sin. 2010, 18, 7. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Zhang, L.P.; Zhang, Y.; Wang, S.; Hu, D.N.; Zuo, J.L. The compositon and diversity of endophytic fungi in different parts of Camellia oleifera Abel. J. For. Eng. 2013, 27, 101–105. [Google Scholar] [CrossRef]
- Strobel, G.A. Endophytes as sources of bioactive products. Microbes Infect. 2003, 5, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Song, W.W. Plant endophytes and their control effects on plant diseases: A review. Jiangsu Agric. Sci. 2018, 46, 12–16. [Google Scholar] [CrossRef]
- Qin, S.; Chen, Y.W.; Xing, K.; Zhang, Q.; Wu, S.H. Isolation, Identification and Bioactivity Screening of Endophytic Fungus Associated with Opuntia ficusindica Mill. Microbiol. China 2006, 33, 95–99. [Google Scholar] [CrossRef]
- Fu, J.H.; Liu, X.; Li, J. Isolation and Antimicrobial Substances of Endophytic Fungi from Sorbus tianshanica Rupr. Acta Bot. Boreali-Occident. Sin. 2012, 32, 2537–2543. [Google Scholar] [CrossRef]
- Tan, Y.A.; Bai, L.X.; Xiao, L.B.; Wei, S.Y.; Zhao, H.X. Herbivore Stress by Lygus lucorum Inducing Protective Enzyme Activity and MDA Content on Different Cotton Varieties. Cotton Sci. 2010, 22, 479–485. [Google Scholar] [CrossRef]
- Shi, J.Y.; Liu, A.Y.; Li, X.P.; Feng, S.J.; Chen, W.X. Inhibitory mechanisms induced by the endophytic bacterium MGY2 in controlling anthracnose of papaya. Biol. Control 2011, 56, 2–8. [Google Scholar] [CrossRef]
- Zhu, X.M.; Shi, X.P.; Yong, D.J.; Zhang, Y.; Li, B.H.; Liang, W.X.; Wang, C.X. Resistance of apple to Anthrax leaf blight induced by endophytic actinomyces A-1. Chin. J. Plant Physiol. 2015, 51, 6. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stem | Leaf | Fruit | Summation | |
|---|---|---|---|---|
| Number of fungal strains | 31 | 45 | 55 | 131 |
| Isolation frequency, % | 12.76 | 18.52 | 22.63 | 18.66 |
| Endophytic Fungi Genera | Separation Tissue | Relative Separation Frequency, % | ||
|---|---|---|---|---|
| Stem, % | Leaf, % | Fruit, % | ||
| Aspergillus spp. | 8.40 | 16.03 | 22.90 | 47.33 |
| Colletotrichum spp. | 5.34 | 9.16 | 0 | 14.50 |
| Candida spp. | 1.53 | 2.29 | 4.58 | 8.40 |
| Cladosporium spp. | 2.29 | 0 | 3.82 | 6.11 |
| Penicillium spp. | 2.29 | 1.53 | 2.29 | 6.11 |
| Fusarium spp. | 1.53 | 1.53 | 0.76 | 3.82 |
| Phoma spp. | 0.76 | 0.76 | 0 | 1.53 |
| Phyllosticta spp. | 0.76 | 0.76 | 0 | 1.53 |
| Trichoderma spp. | 0 | 0.76 | 0.76 | 1.53 |
| Talaromyces spp. | 0 | 0.76 | 0.76 | 1.53 |
| Diaporthe spp. | 0 | 0.76 | 0 | 0.76 |
| Phialemonium spp. | 0 | 0 | 0.76 | 0.76 |
| Neofusicoccum spp. | 0 | 0 | 0.76 | 0.76 |
| Sporothrix spp. | 0 | 0 | 0.76 | 0.76 |
| Cerrena spp. | 0.76 | 0 | 0 | 0.76 |
| Wallemia spp. | 0 | 0 | 0.76 | 0.76 |
| Gibellulopsis spp. | 0 | 0 | 0.76 | 0.76 |
| Phlebia spp. | 0 | 0 | 0.76 | 0.76 |
| Alternaria spp. | 0 | 0 | 0.76 | 0.76 |
| Daldinia spp. | 0 | 0 | 0.76 | 0.76 |
| Sample Name | DPPH Scavenging Rate, %, at Different Concentrations of Samples | IC50 Value, mg/L | ||||
|---|---|---|---|---|---|---|
| 6.25 mg/L | 12.5 mg/L | 25 mg/L | 50 mg/L | 100 mg/L | ||
| VC | 30.37 ± 1.04 a | 43.43 ± 0.91 a | 70 ± 0.68 a | 96.63 ± 0.16 a | 97.85 ± 0.04 a | 12.55 ± 0.0004 e |
| BHT | 17.91 ± 0.81 b | 38.92 ± 0.35 b | 71.4 ± 0.27 a | 83.68 ± 0.60 b | 88.46 ± 0.26 b | 16.01 ± 0.0031 e |
| Y17 | 17.17 ± 0.38 b | 38.22 ± 0.29 b | 63 ± 0.45 b | 70.88 ± 0.84 c | 82.72 ± 0.68 c | 19.72 ± 0.0052 e |
| Y28 | 3.03 ± 0.48 d | 10.8 ± 0.73 d | 28.49 ± 0.39 c | 52.31 ± 0.24 d | 71.4 ± 0.13 d | 49.22 ± 0.0016 d |
| J28 | 9.58 ± 0.85 c | 19.35 ± 0.91 c | 29.63 ± 0.76 c | 43.88 ± 0.30 f | 69.03 ± 0.22 e | 54.32 ± 0.0027 d |
| G45 | 3.48 ± 0.27 d | 9.36 ± 0.46 d | 29.41 ± 0.23 c | 36.63 ± 0.48 g | 65.63 ± 1.00 e | 64.44 ± 0.0033 c |
| Y8 | 0.92 ± 0.74 d | 6.81 ± 0.43 e | 25.27 ± 0.75 d | 36.39 ± 0.95 e | 53.9 ± 0.1 g | 72.07 ± 0.0050 b |
| J2 | 1.44 ± 0.45 d | 10.06 ± 0.15 d | 20.09 ± 0.17 e | 32.93 ± 0.49 h | 61.64 ± 0.04 f | 74.84 ± 0.0018 b |
| G41 | 3.4 ± 0.20 d | 9.29 ± 0.61 d | 12.91 ± 0.32 f | 31.74 ± 0.40 h | 56.86 ± 0.13 g | 84.06 ± 0.0016 a |
| Sample Name | ABTS Scavenging Rate, %, at Different Concentrations of Samples | IC50 Value, mg/L | ||||
|---|---|---|---|---|---|---|
| 6.25 mg/L | 12.5 mg/L | 25 mg/L | 50 mg/L | 100 mg/L | ||
| VC | 45.79 ± 0.72 a | 73.17 ± 0.18 a | 84.22 ± 0.35 a | 98.89 ± 0.1 a | 99.6 ± 0.05 a | 6.91 ± 0.0012 d |
| BHT | 35.85 ± 0.85 b | 47.66 ± 0.53 b | 61.78 ± 0.93 b | 81.29 ± 0.53 b | 93.6 ± 0.20 b | 12.86 ± 0.0025 d |
| Y17 | 31.72 ± 0.13 c | 46.9 ± 0.68 b | 60.62 ± 0.46 b | 76.8 ± 0.13 c | 91.83 ± 0.38 c | 14.34 ± 0.0016 d |
| Y28 | 17.55 ± 0.17 f | 28.04 ± 0.36 e | 43.47 ± 0.36 c | 55.07 ± 0.35 d | 76.25 ± 0.09 d | 34.23 ± 0.0043 c |
| J2 | 28.34 ± 0.13 f | 33.99 ± 0.41 f | 40.34 ± 0.79 f | 52.75 ± 0.13 g | 69.64 ± 0.39 g | 36 ± 0.0021 c |
| G45 | 20.42 ± 0.43 e | 29.55 ± 0.63 d | 35.3 ± 0.10 e | 49.52 ± 0.68 f | 71.66 ± 0.31 e | 42.31 ± 0.0012 c |
| J28 | 18.91 ± 0.15 g | 25.92 ± 0.33 g | 31.16 ± 0.26 e | 44.68 ± 0.18 g | 66.46 ± 0.05 g | 54.1 ± 0.0022 b |
| Y8 | 11.9 ± 0.22 d | 17.35 ± 0.57 c | 33.23 ± 0.64 d | 45.89 ± 0.66 e | 66.92 ± 0.18 f | 54.36 ± 0.0032 b |
| G41 | 12.71 ± 0.23 g | 17.3 ± 0.22 g | 25.82 ± 0.18 g | 37.32 ± 0.10 h | 66.16 ± 0.22 g | 64.63 ± 0.0012 a |
| Fungal Genus | Strain Number | Inhibition Rate (%) | ||||
|---|---|---|---|---|---|---|
| Fusarium oxysporum | Neoscytalidium dimidiatum | Pestalotiopsis spp. | Botryodiplodia theobromae | Colletotrichum gloeosporioides penz. | ||
| Fusarium spp. | J8 | 44.16 ± 1.02 | - | 61.78 ± 1.49 | - | - |
| Phyllosticta spp. | J28 | - | - | 36.44 ± 0.21 | 48.52 ± 2.49 | 44.91 ± 4.55 |
| Penicillium spp. | Y17 | - | 53.29 ± 0.98 | 37.25 ± 1.27 | 40.62 ± 1.61 | 54.09 ± 1.45 |
| Y8 | 27.51 ± 1.42 | - | - | 45.86 ± 1.93 | ||
| Aspergillus spp. | J16 | 38.27 ± 0.75 | 68.44 ± 0.47 | - | - | 58.09 ± 2.80 |
| J17 | - | - | - | 46.62 ± 3.13 | 30.91 ± 2.32 | |
| Y23 | 23.82 ± 1.56 | 46.58 ± 1.14 | - | - | - | |
| Y28 | - | 50.81 ± 1.82 | 56.11 ± 1.01 | 48.52 ± 1.95 | - | |
| G5 | 29.83 ± 2.47 | 31.63 ± 1.65 | - | - | 43.00 ± 1.77 | |
| G16 | 24.62 ± 2.20 | - | 46.96 ± 1.69 | - | 53.82 ± 1.14 | |
| G45 | - | - | - | 45.04 ± 2.05 | 52.64 ± 2.56 | |
| Phoma spp. | Y9 | 41.97 ± 2.27 | - | 39.68 ± 0.57 | - | - |
| Candida spp. | G39 | 30.87 ± 4.05 | - | 66.40 ± 1.53 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, J.; Ke, S.; He, X.; Zhang, B.; Zheng, Z.; Chen, P. Research on the Isolation of Endophytic Fungi from Papaya and the Prevention of Colletotrichum gloeosporioides. J. Fungi 2024, 10, 550. https://doi.org/10.3390/jof10080550
Lv J, Ke S, He X, Zhang B, Zheng Z, Chen P. Research on the Isolation of Endophytic Fungi from Papaya and the Prevention of Colletotrichum gloeosporioides. Journal of Fungi. 2024; 10(8):550. https://doi.org/10.3390/jof10080550
Chicago/Turabian StyleLv, Jinhui, Shuwei Ke, Xinrui He, Baolong Zhang, Zhongbing Zheng, and Ping Chen. 2024. "Research on the Isolation of Endophytic Fungi from Papaya and the Prevention of Colletotrichum gloeosporioides" Journal of Fungi 10, no. 8: 550. https://doi.org/10.3390/jof10080550
APA StyleLv, J., Ke, S., He, X., Zhang, B., Zheng, Z., & Chen, P. (2024). Research on the Isolation of Endophytic Fungi from Papaya and the Prevention of Colletotrichum gloeosporioides. Journal of Fungi, 10(8), 550. https://doi.org/10.3390/jof10080550