
Morphological and Phylogenetic Analyses Reveal Three Novel Species of Sanguinoderma (Ganodermataceae, Basidiomycota) from Yunnan Province, China


Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection
2.2. Morphological Observations
2.3. Growth Rate of Mycelium on Culture Media
2.4. DNA Extraction, PCR Amplification and Sequencing
2.5. Sequence Alignment and Phylogenetic Analysis
2.6. Phylogenetic Inference
3. Results
3.1. Phylogenetic Analyses
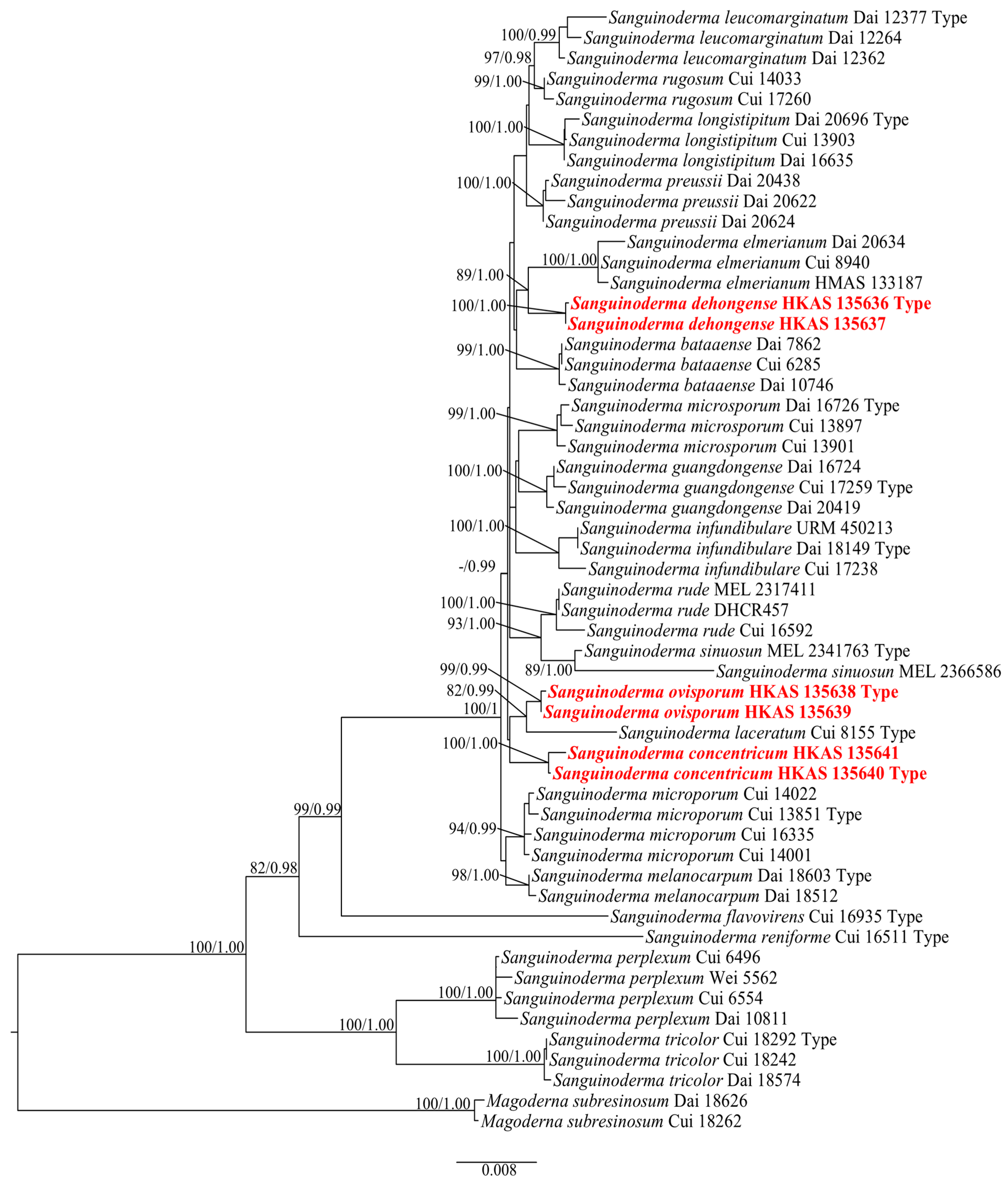
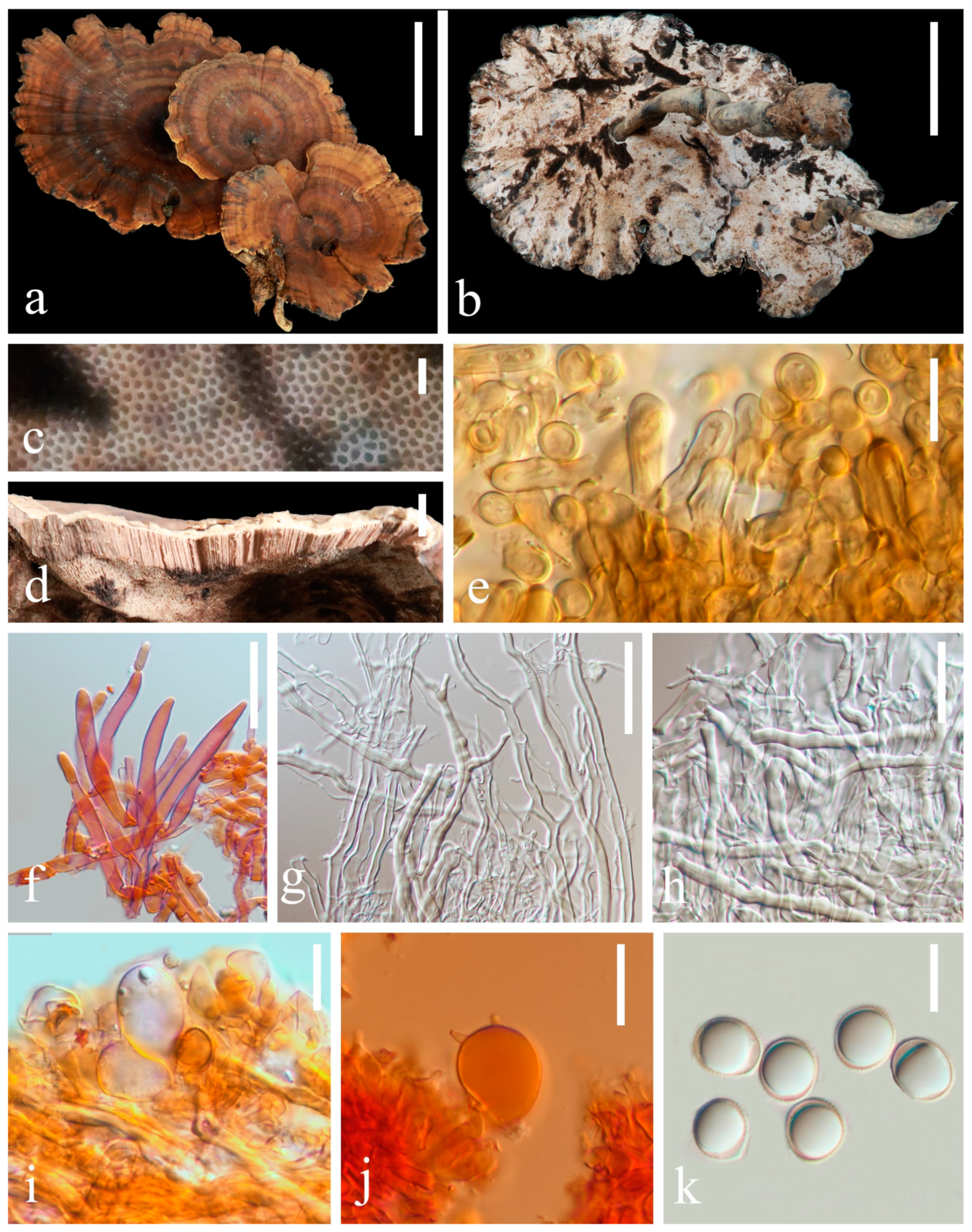
3.2. Taxonomy
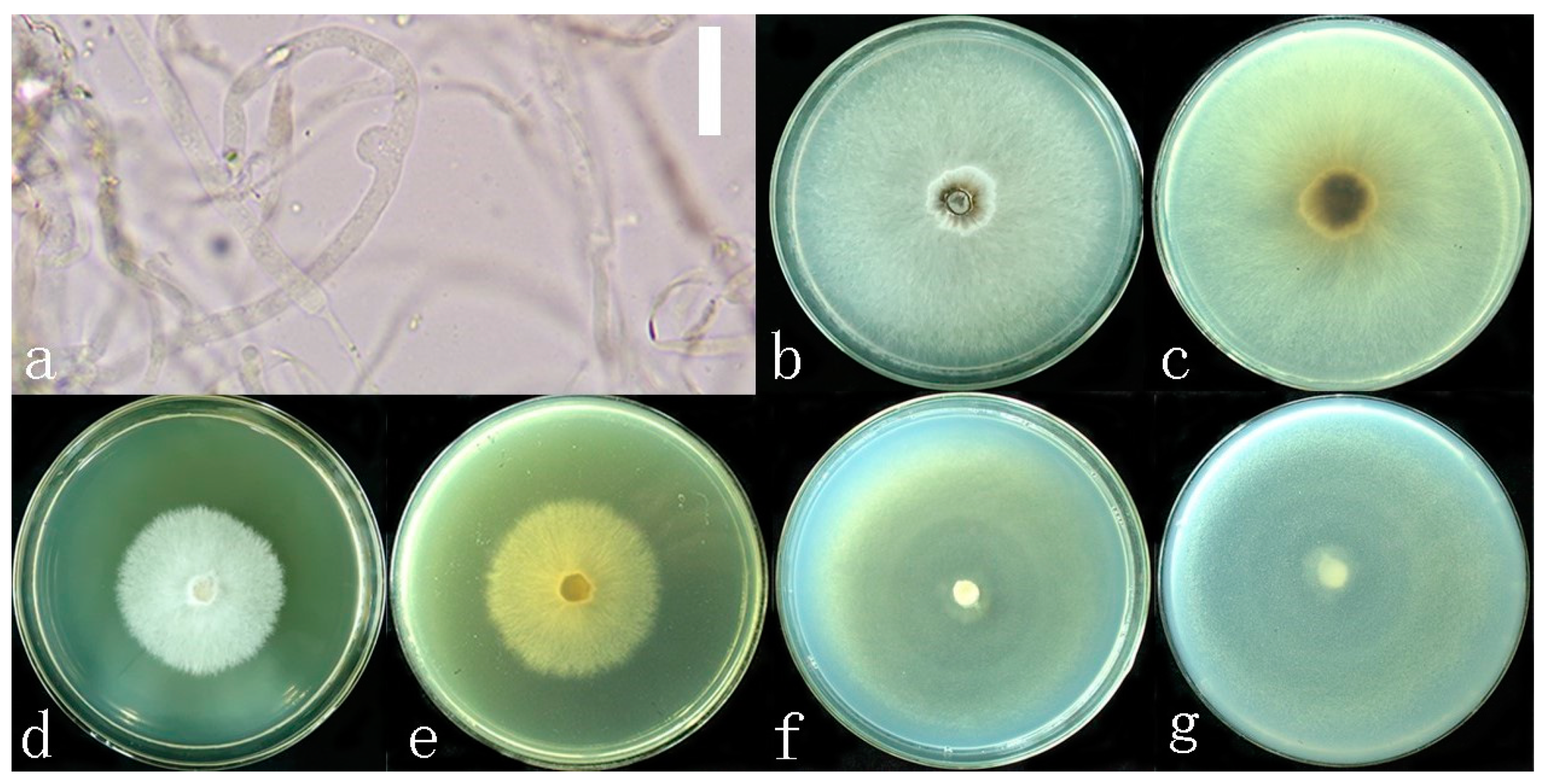
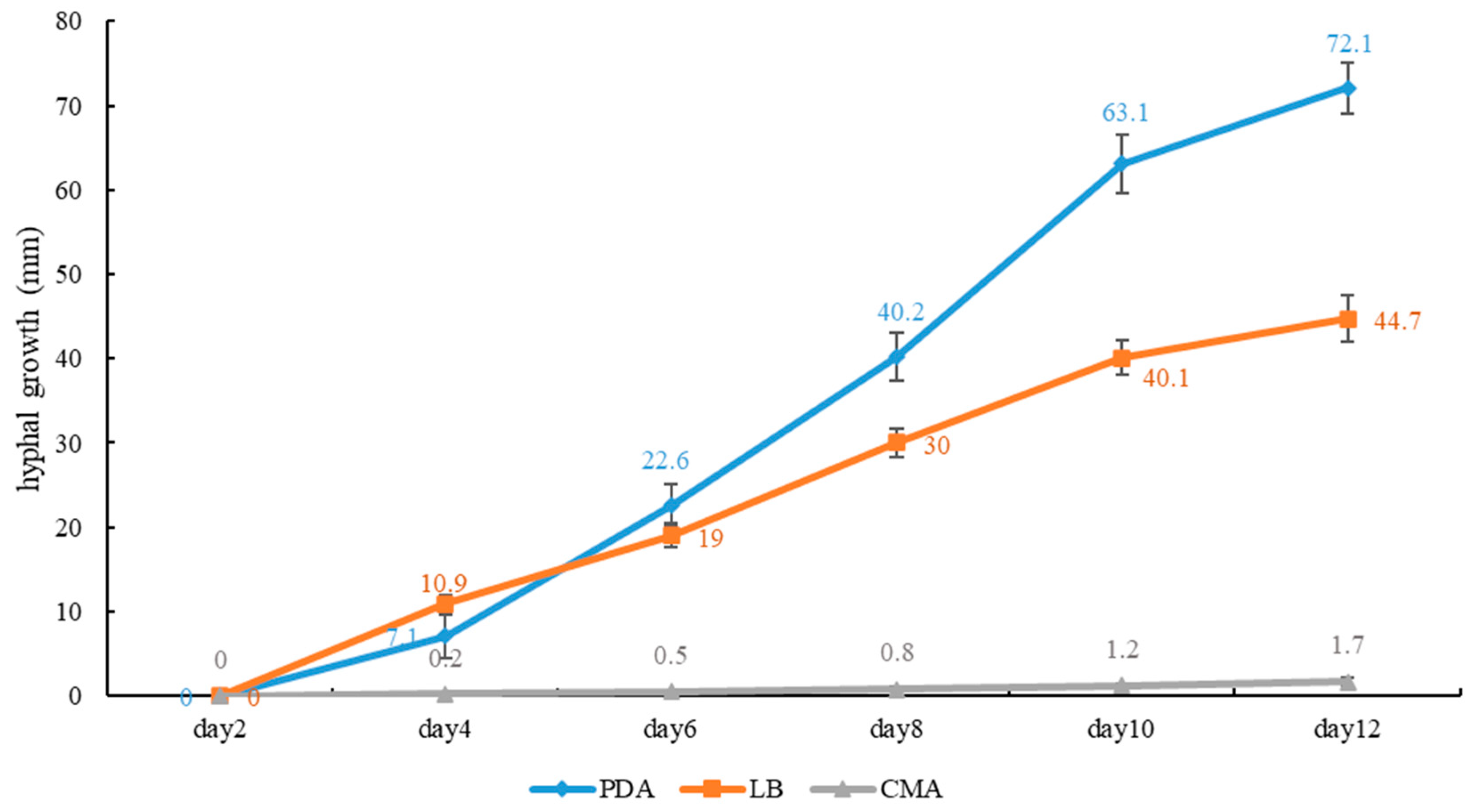
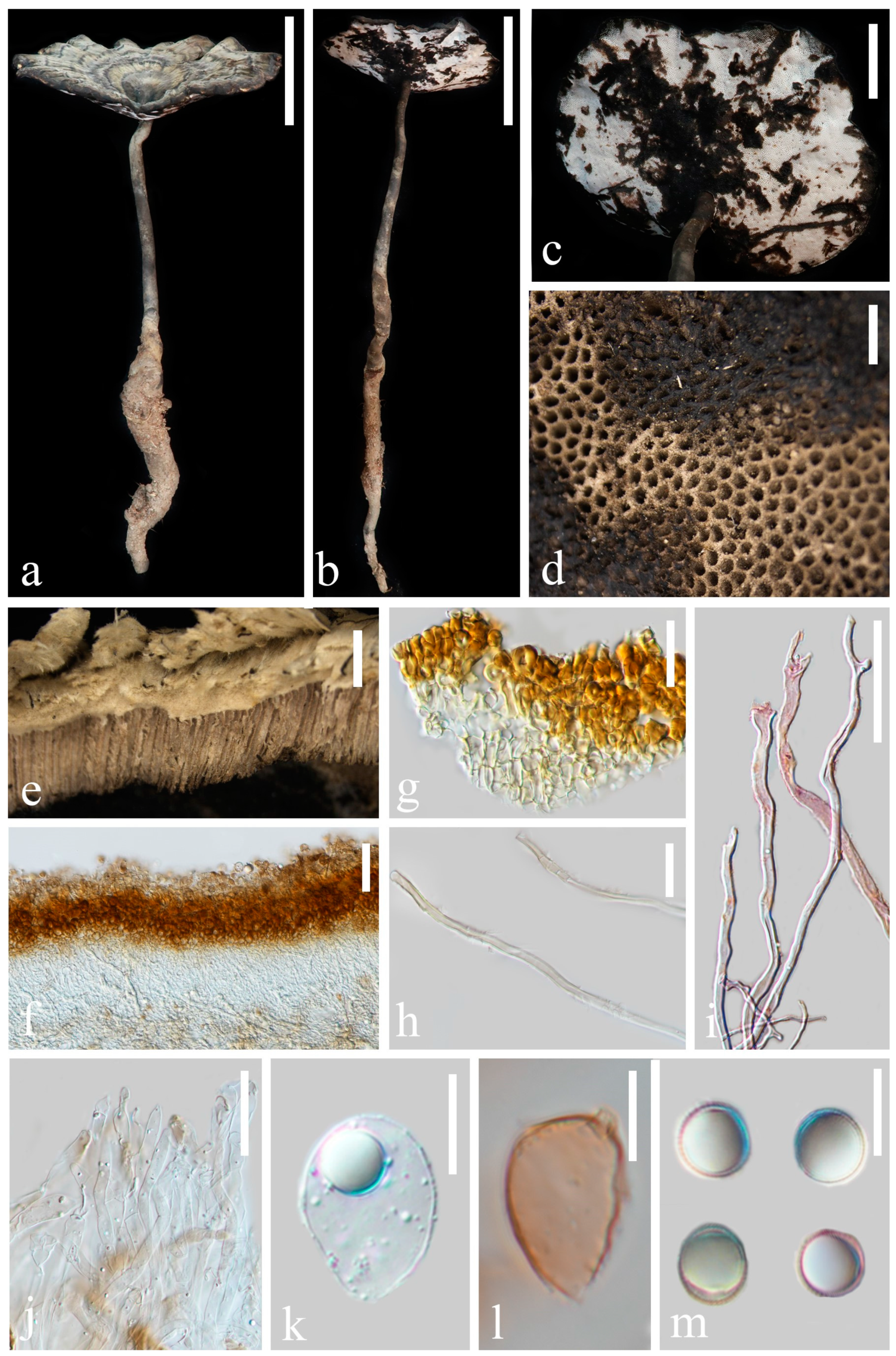
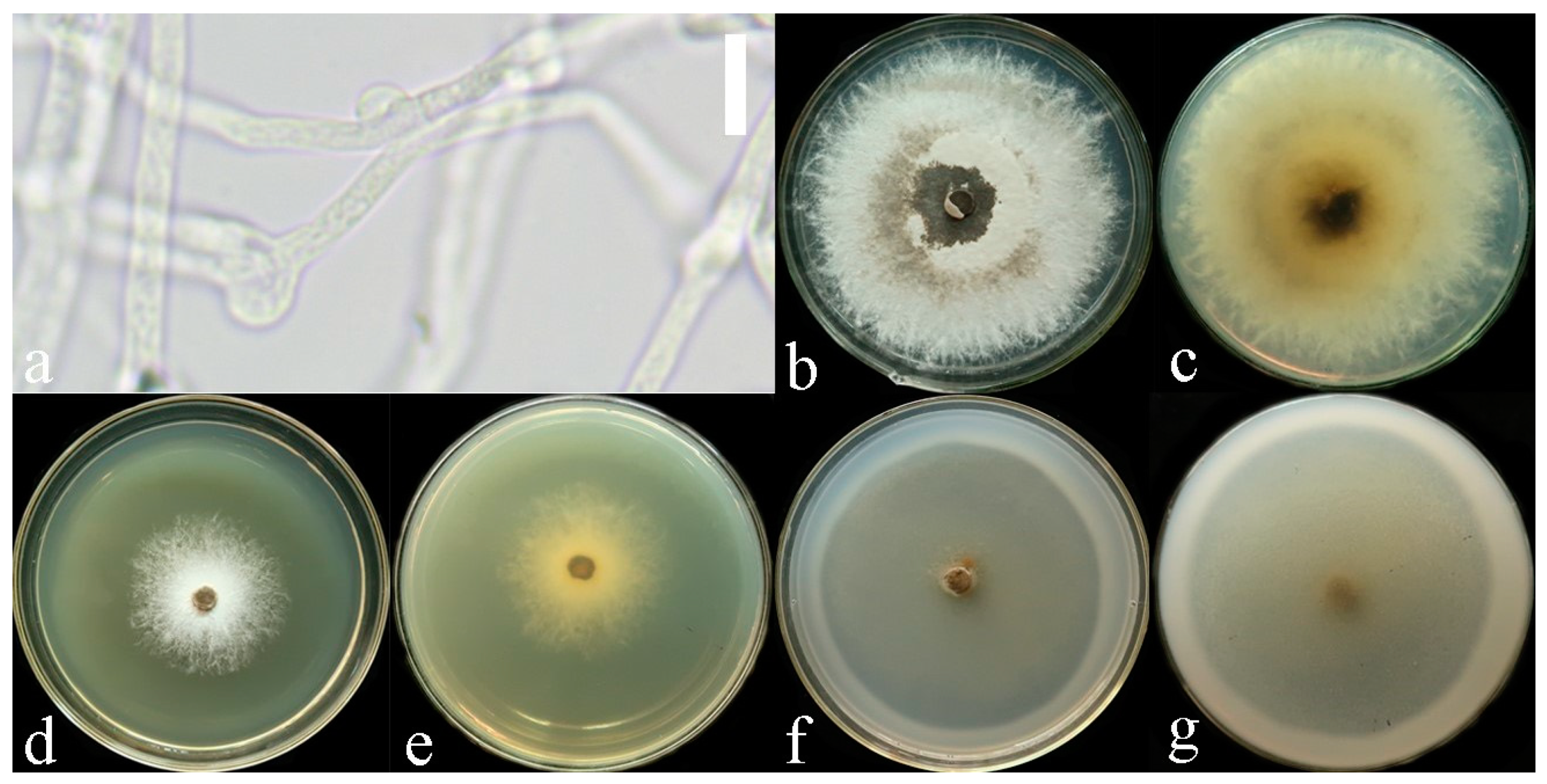
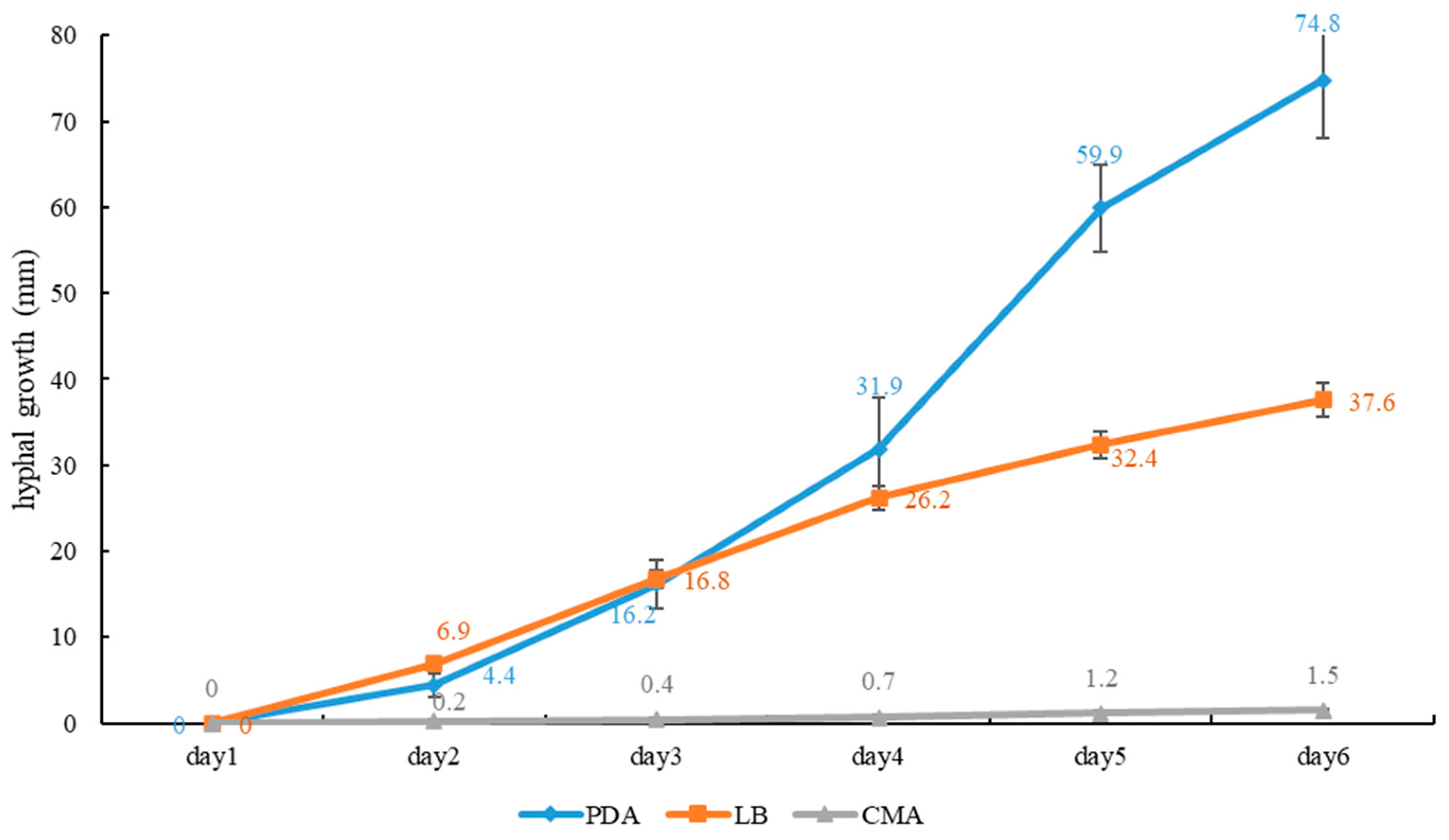
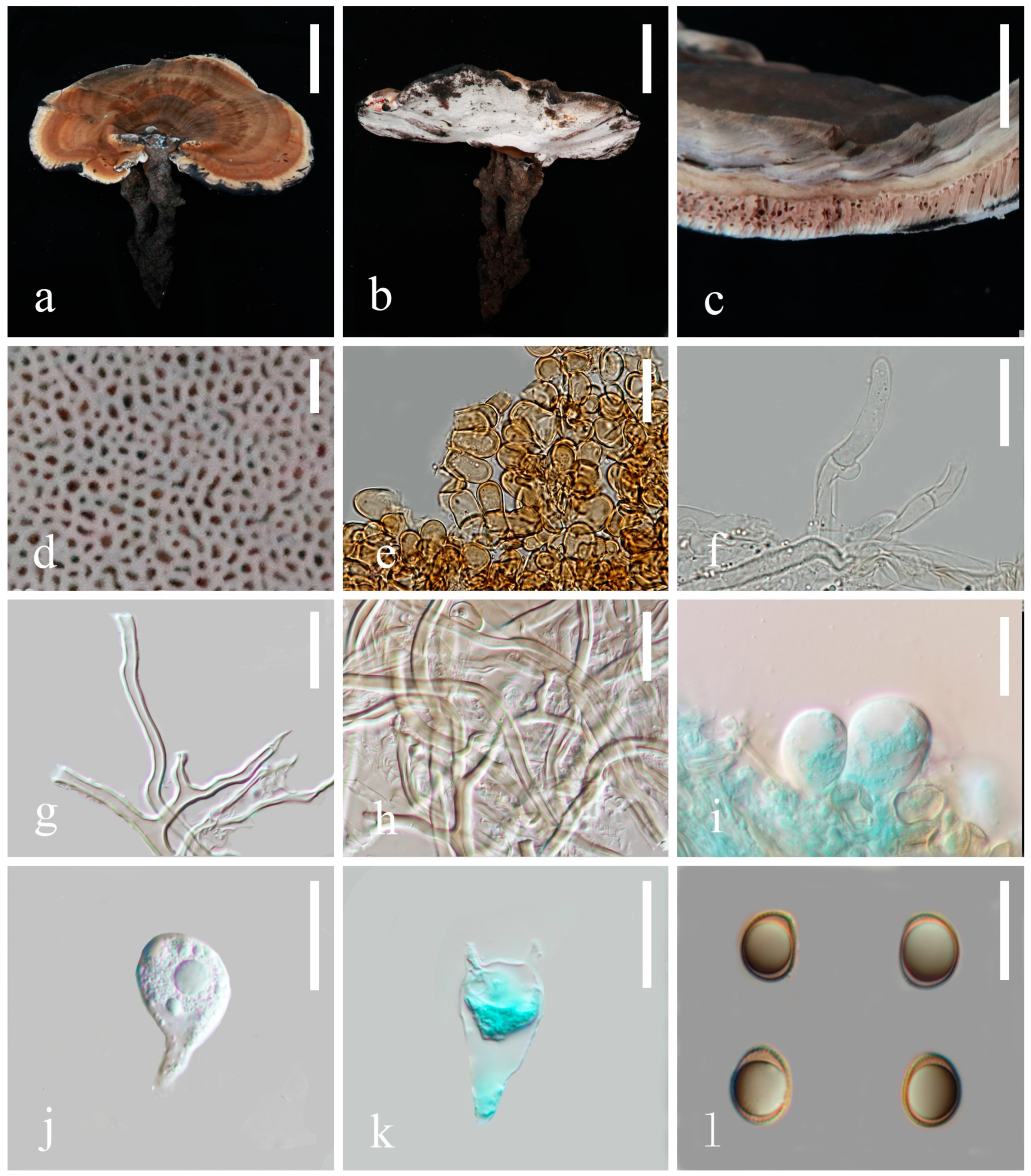
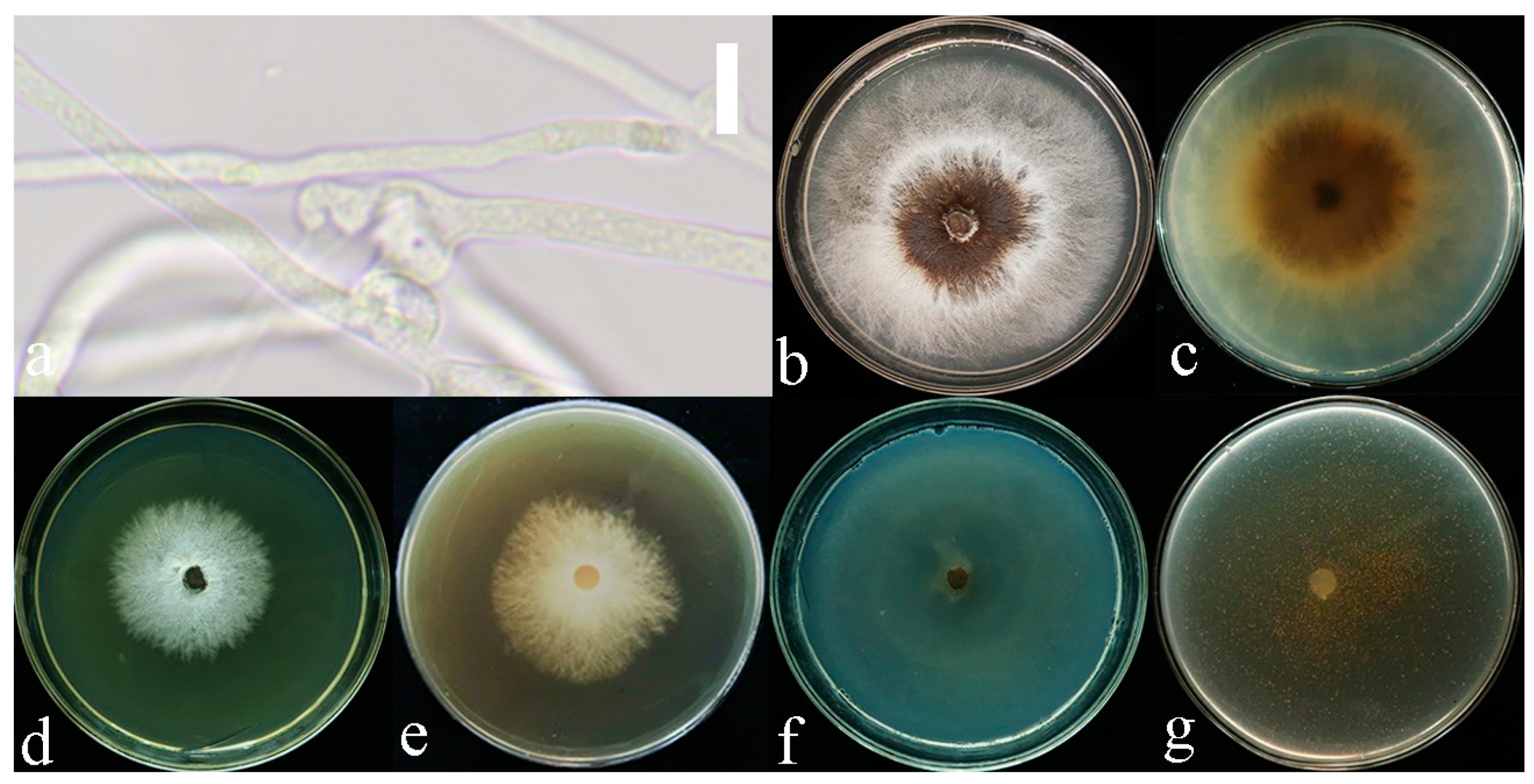
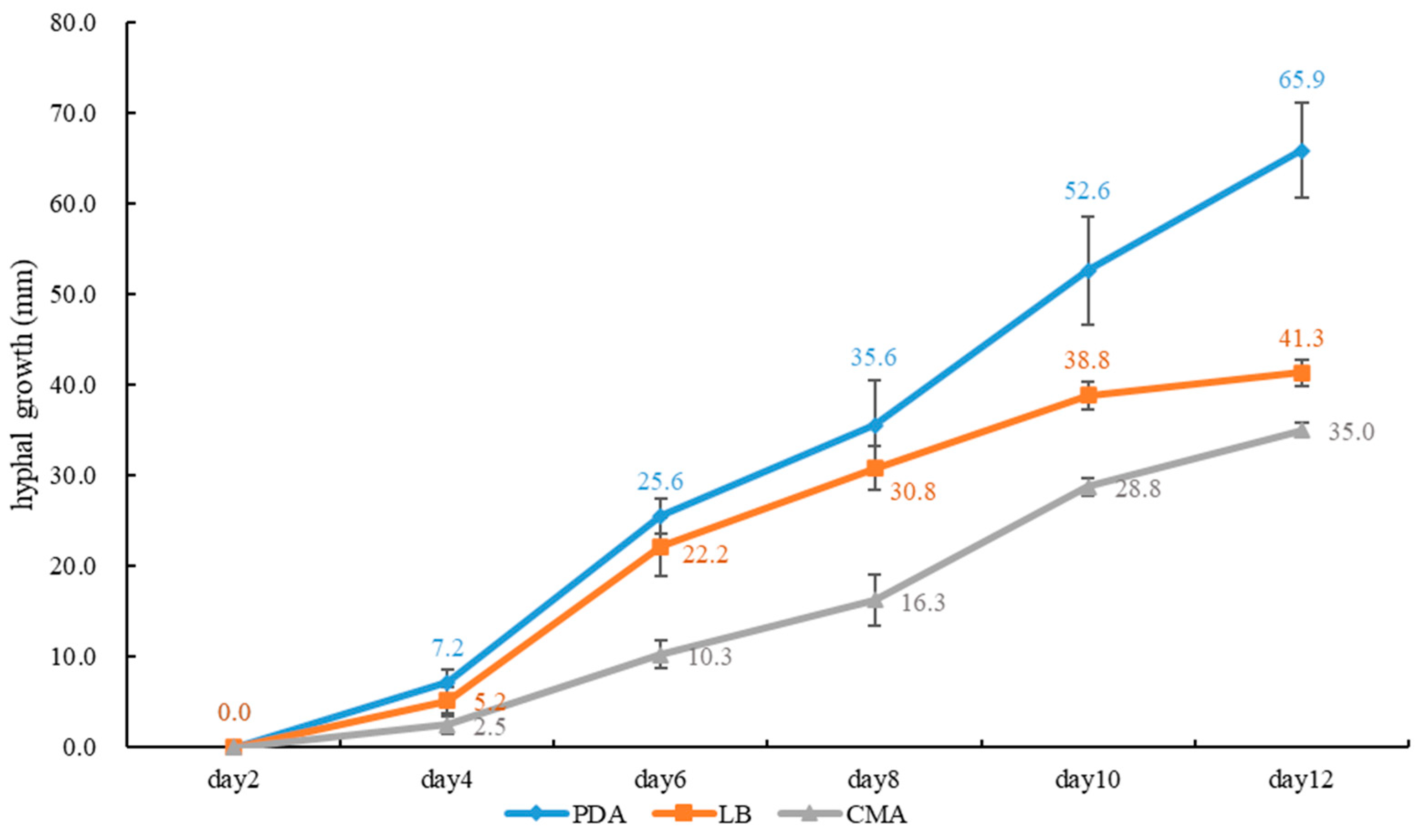

4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, Y.C.; Yang, Z.L.; Cui, B.K.; Yu, C.J.; Zhou, L.W. Species diversity and utilization of medicinal mushrooms and fungi in China. Int. J. Med. Mushrooms 2009, 11, 287–302. [Google Scholar] [CrossRef]
- Chan, K.S. Anti–Hyperlipidemic Activities of Amauroderma rugosum on Human Hepatocellular Carcinoma Cells. Ph.D. Thesis, University of Malaya, Kuala Lumpur, Malaysia, 2018. [Google Scholar]
- Chan, P.M.; Tan, Y.S.; Chua, K.H.; Sabaratnam, V.; Kuppusamy, U.R. Attenuation of inflammatory mediators (TNF-α and nitric oxide) and up–regulation of IL–10 by wild and domesticated Basidiocarps of Amauroderma rugosum (Blume and T. Nees) Torrend in LPS–stimulated RAW264.7 cells. PLoS ONE 2015, 10, e0139593. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.-M.; Kanagasabapathy, G.; Tan, Y.-S.; Sabaratnam, V.; Kuppusamy, U.R. Amauroderma rugosum (Blume and T. Nees) Torrend: Nutritional composition and antioxidant and potential anti–inflammatory properties. Evid. Based Complement. Altern. Med. 2013, 304713. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Lee, S.S. Utilisation of macrofungi species in Malaysia. Fungal Divers. 2013, 1, 15–22. [Google Scholar]
- Liew, G.M.; Khong, H.Y.; Kutoi, C.J. Phytochemical screening, antimicrobial and antioxidant activities of selected fungi from mount Singai, Sarawak. Malays. Int. J. Res. Stud. Biosci. 2015, 3, 191–197. [Google Scholar]
- Zheng, C.W.; Cheung, T.M.Y.; Leung, G.P.H. A review of the phytochemical and pharmacological properties of Amauroderma rugosum. Kaohsiung J. Med. Sci. 2022, 38, 509–516. [Google Scholar] [CrossRef]
- Sun, Y.-F.; Costa-Rezende, D.H.; Xing, J.-H.; Zhou, J.-L.; Zhang, B.; Gibertoni, T.B.; Gates, G.; Glen, M.; Dai, Y.-C.; Cui, B.-K. Multi-gene phylogeny and taxonomy of Amauroderma s. lat. (Ganodermataceae). Persoonia Mol. Phylogeny Evol. Fungi 2020, 44, 206–239. [Google Scholar] [CrossRef]
- Murrill, W.A. Tomophagus for Dendrophagus. Torreya 1905, 5, 197. [Google Scholar]
- Costa-Rezende, D.H.; Gugliotta, A.M.; Goes-Neto, A.; Reck, M.A.; Robledo, G.L.; Drechsler-Santos, E.R. Amauroderma calcitum sp. nov. and notes on taxonomy and distribution of Amauroderma species Ganodermataceae. Phytotaxa 2016, 244, 101–124. [Google Scholar]
- Costa-Rezende, D.H.; Robledo, G.L.; Góes-Neto, A.; Reck, M.A.; Crespo, E.; Drechsler-Santos, E.R. Morphological reassessment and molecular phylogenetic analyses of Amauroderma s. lat. raised new perspectives in the generic classification of the Ganodermataceae family. Persoonia Mol. Phylogeny Evol. Fungi 2017, 39, 254–269. [Google Scholar] [CrossRef]
- Wachtel-Galor, S.; Yuen, J.; Buswell, J.A.; Benzie, I.F. Ganoderma lucidum (Lingzhi or Reishi): A Medicinal Mushroom. Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; CRC Press/Taylor and Francis: Boca Raton, FL, USA, 2011. [Google Scholar]
- Azliza, M.A.; Ong, H.C.; Vikineswary, S.; Noorlidah, A.; Haron, N.W. Ethno-medicinal resources used by the Temuan in Ulu Kuang Village. Stud. Ethno-Med. 2012, 6, 17–22. [Google Scholar] [CrossRef]
- Sun, Y.F.; Fang, Y.X.; Cui, B.K. Taxonomy and phylogeny of Sanguinoderma rugosum complex with descriptions of a new species and a new combination. Front. Microbiol. 2022, 13, 1087212. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Luo, Z.L.; Tang, S.M.; Li, Y.J.; Li, S.H.; Su, H.Y. Phylogenetic analyses and morphological characters reveal two new species of Ganoderma from Yunnan Province, China. MycoKeys 2021, 84, 141. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee SJ, W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press, Inc.: Cambridge, MA, USA, 1990; Volume 38, pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus specie. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorph. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Sun, Y.-F.; Xing, J.-H.; He, X.-L.; Wu, D.-M.; Song, C.-G.; Liu, S.; Vlasák, J.; Gates, G.; Gibertoni, T.B.; Cui, B.-K. Species diversity, systematic revision and molecular phylogeny of Ganodermataceae (Polyporales, Basidiomycota) with an emphasis on Chinese collections. Stud. Mycol. 2022, 101, 287–415. [Google Scholar] [CrossRef]
- Li, M.J.; Yuan, H.S. Type studies on Amauroderma species described by JD Zhao et al. and the phylogeny of species in China. Mycotaxon 2015, 130, 79–89. [Google Scholar] [CrossRef]
- Song, J.; Xing, J.H.; Decock, C.; He, X.L.; Cui, B.K. Molecular phylogeny and morphology reveal a new species of Amauroderma (Basidiomycota) from China. Phytotaxa 2016, 260, 47–56. [Google Scholar] [CrossRef]
- Costa-Rezende, D.H.; Robledo, G.L.; Drechsler-Santos, E.R.; Glen, M.; Gates, G.; de Madrignac Bonzi, B.R.; Popoff, O.F.; Crespo, E. Taxonomy and phylogeny of polypores with ganodermatoid basidiospores (Ganodermataceae). Mycol. Prog. 2020, 19, 725–741. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user–friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Glez-Pen, D.; Gomez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program-oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post–analysis of large phylogenie. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi–gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A. MrModeltest 2.0. Program. Distributed; Department Systematic Zoology, EBC, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Luangharn, T.; Karunarathna, S.C.; Dutta, A.K.; Paloi, S.; Promputtha, I.; Hyde, K.D.; Xu, J.; Mortimer, P.E. Ganoderma (Ganodermataceae, Basidiomycota) species from the greater Mekong subregion. J. Fungi 2021, 7, 819. [Google Scholar] [CrossRef] [PubMed]
- Luangharn, T.; Karunarathna, S.C.; Mortimer, P.E.; Hyde, K.D.; Thongklang, N.; Xu, J. A new record of Ganodermatropicum (Basidiomycota, Polyporales) for Thailand and first assessment of optimum conditions for mycelia production. MycoKeys 2019, 51, 65. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | ITS | LSU | rpb2 | tef1-α | mtSSU | nSSU | References |
|---|---|---|---|---|---|---|---|---|---|
| Sanguinoderma bataaense | Dai 10746 | Hainan, China | MK119832 | MK119911 | MK121511 | MK121581 | MZ352801 | MZ355267 | [8,20] |
| Cui 6285 | Hainan, China | MK119831 | MK119910 | MK121537 | MK121580 | MZ352793 | MZ355238 | [8,20] | |
| Dai 7862 | Hainan, China | KJ531658 | - | - | - | - | - | [21] | |
| S. concentricum | HKAS 135640 T | Yunnan, China | PP951682 | PP951731 | PP998313 | PP998318 | PP988462 | PP982317 | This study |
| HKAS 135641 | Yunnan, China | PP951683 | PP951732 | - | PP998319 | PP988463 | PP982318 | This study | |
| S. dehongense | HKAS 135636 T | Yunnan, China | PP947806 | PP951727 | PP998309 | PP998314 | PP988458 | PP982313 | This study |
| HKAS 135637 | Yunnan, China | PP951679 | PP951728 | PP998310 | PP998315 | PP988459 | PP982314 | This study | |
| S. elmerianum | HMAS 133187 | Yunnan, China | MK119834 | MK119913 | - | - | MZ352824 | MZ355234 | [8,20] |
| Dai 20634 | Yunnan, China | MZ354875 | MZ355082 | - | MZ221724 | MZ352821 | MZ355148 | [8,20] | |
| Cui 8940 | Guangdong, China | MK119833 | MK119912 | - | - | MZ352812 | MZ355305 | [8,20] | |
| S. flavovirens | Cui 16935 T | Zambia | - | MK119914 | MK121532 | MK121582 | MZ352811 | MZ355254 | [8,20] |
| S. guangdongense | Cui 17259 T | Guangdong, China | MZ354877 | MZ355123 | MZ358834 | MZ221726 | MZ352816 | MZ355139 | [20] |
| Dai 16724 | Thailand, China | MZ354876 | MZ355117 | MZ358833 | MZ221725 | MZ352815 | MZ355271 | [20] | |
| Dai 20419 | Yunnan, China | MZ354890 | MZ355083 | MZ358835 | MZ221727 | MZ352818 | MZ355155 | [20] | |
| S. infundibulare | Dai 18149 T | Guangdong, China | MK119847 | MK119926 | MK121529 | MK121597 | MZ352790 | MZ355239 | [8,20] |
| URM 450213 | Ecuador | MK119849 | MK119927 | - | - | MZ352792 | MZ355252 | [8,20] | |
| Cui 17238 | Guangdong, China | OM780277 | - | MZ358837 | MZ221729 | MZ352800 | MZ355149 | [20] | |
| S. laceratum | Cui 8155 T | Yunnan, China | NR174040 | MK119928 | - | - | MZ352810 | - | [8,20] |
| S. leucomarginatum | Dai 12264 | Yunnan, China | OP700311 | OP700344 | OP696845 | OP696857 | OP703259 | OP700325 | [14,20] |
| Dai 12377 T | Yunnan, China | OP700312 | OP700345 | OP696846 | OP696860 | OP703260 | OP700326 | [14] | |
| Dai 12362 | Yunnan, China | KU219986 | KU220009 | OP696847 | OP696858 | OP703261 | OP700327 | [22] | |
| S. longistipitum | Dai 20696 T | Yunnan, China | MZ354881 | MZ355084 | - | MZ221732 | MZ352822 | MZ355145 | [20] |
| Cui 13903 | Hainan, China | MZ354882 | MZ355114 | MZ358839 | MZ221733 | MZ352809 | MZ355301 | [20] | |
| Dai 16635 | Thailand, China | MZ354883 | MZ355120 | MZ358840 | MZ221734 | MZ352802 | MZ355260 | [20] | |
| S. melanocarpum | Dai 18512 | Malaysia | MZ354888 | MZ355118 | - | MZ221735 | MZ352794 | MZ355313 | [20] |
| Dai 18603 T | Malaysia | MZ354889 | MZ355113 | MZ358841 | MZ221736 | MZ352796 | MZ355281 | [20] | |
| S. microporum | Cui 13851 T | Hainan, China | MK119854 | MK119933 | MK121512 | MK121602 | MZ352797 | MZ355270 | [8,20] |
| Cui 14022 | Guangxi, China | MK119856 | MK119935 | MK121515 | MK121604 | MZ352798 | MZ355298 | [8,20] | |
| Cui 16335 | Guangxi, China | MK119857 | MK119936 | MK121514 | MK121605 | OP703262 | OP700328 | [14,20] | |
| Cui 14001 | Guangxi, China | MK119855 | MK119934 | MK121513 | MK121603 | OP703263 | OP700329 | [14,20] | |
| S. microsporum | Dai 16726 T | Thailand, China | - | MZ355119 | - | MZ221737 | MZ352795 | MZ355272 | [20] |
| Cui 13897 | Hainan, China | MZ354878 | MZ355127 | - | MZ221739 | MZ352804 | MZ355300 | [20] | |
| Cui 13901 | Hainan, China | MZ354879 | MZ355121 | - | MZ221738 | MZ352803 | MZ355299 | [20] | |
| S. ovisporum | HKAS 135638 T | Yunnan, China | PP951680 | PP951729 | PP998311 | PP998316 | PP988460 | PP982315 | This study |
| HKAS 135639 | Yunnan, China | PP951681 | PP951730 | PP998312 | PP998317 | PP988461 | PP982316 | This study | |
| S. perplexum | Cui 6496 | Hainan, China | KJ531650 | KU220001 | MK121538 | MK121583 | MZ352825 | MZ355263 | [20,21] |
| Cui 6554 | Hainan, China | MK119835 | MK119915 | MK121540 | MK121585 | MZ352826 | MZ355264 | [8,20] | |
| Dai 10811 | Hainan, China | KJ531651 | KU220002 | MK121539 | MK121584 | MZ352827 | MZ355302 | [20,21] | |
| Wei 5562 | Hainan, China | KJ531652 | - | - | - | - | - | [21] | |
| S. preussii | Dai 20438 | Yunnan, China | OP700314 | OP700347 | OP696848 | OP696869 | OP703265 | OP700331 | [14] |
| Dai 20622 | Yunnan, China | OP700315 | OP700348 | - | OP696862 | OP703266 | OP700332 | [14] | |
| Dai 20624 | Yunnan, China | OP700316 | OP700349 | - | OP696863 | OP703267 | OP700333 | [14] | |
| S. reniforme | Cui 16511 T | Zambia | NR174041 | MK119929 | MK121531 | MK121599 | - | MZ355322 | [8,20] |
| S. rude | MEL 2317411 | Australia | MK119842 | - | MK121524 | MK121592 | MZ352819 | MZ355306 | [8,20] |
| DHCR457 | Brazil | MN077517 | MN077551 | - | MN061693 | - | - | [23] | |
| Cui 16592 | Australia | MK119836 | MK119916 | MK121521 | MK121586 | MZ352924 | MZ355307 | [8,20] | |
| S. rugosum | Cui 17260 | Guangdong, China | OP700317 | OP700350 | OP696849 | OP696859 | OP703270 | OP700336 | [14] |
| Cui 14033 | Guangxi, China | OP700318 | OP700351 | OP696850 | OP696864 | OP703271 | OP700337 | [14] | |
| S. sinuosum | MEL 2341763 T | Australia | MK119853 | MK119931 | MK121525 | MK121601 | MZ352820 | MZ355291 | [8,20] |
| MEL 2366586 | Australia | MK119852 | MK119930 | MK121527 | MK121600 | MZ352920 | MZ355261 | [8,20] | |
| S. tricolor | Cui 18242 | Malaysia | MZ354992 | MZ355099 | MZ358843 | MZ221743 | MZ352829 | MZ355303 | [20] |
| Cui 18292 T | Malaysia | - | MZ355101 | - | MZ221742 | MZ352828 | MZ355273 | [20] | |
| Dai 18574 | Malaysia | MZ354993 | MZ355102 | MZ358844 | MZ221744 | MZ352830 | MZ355265 | [20] | |
| Magoderna subresinosum | Dai 18626 | Malaysia | MK119823 | MK119902 | MK121507 | MK121571 | MZ352831 | MZ355211 | [8,20] |
| Cui 18262 | Malaysia | MZ354871 | MZ355088 | - | - | MZ352832 | MZ355258 | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, K.-Y.; He, J.; Tang, S.-M.; Su, X.-J.; Luo, Z.-L. Morphological and Phylogenetic Analyses Reveal Three Novel Species of Sanguinoderma (Ganodermataceae, Basidiomycota) from Yunnan Province, China. J. Fungi 2024, 10, 589. https://doi.org/10.3390/jof10080589
Niu K-Y, He J, Tang S-M, Su X-J, Luo Z-L. Morphological and Phylogenetic Analyses Reveal Three Novel Species of Sanguinoderma (Ganodermataceae, Basidiomycota) from Yunnan Province, China. Journal of Fungi. 2024; 10(8):589. https://doi.org/10.3390/jof10080589
Chicago/Turabian StyleNiu, Kai-Yang, Jun He, Song-Ming Tang, Xi-Jun Su, and Zong-Long Luo. 2024. "Morphological and Phylogenetic Analyses Reveal Three Novel Species of Sanguinoderma (Ganodermataceae, Basidiomycota) from Yunnan Province, China" Journal of Fungi 10, no. 8: 589. https://doi.org/10.3390/jof10080589
APA StyleNiu, K.-Y., He, J., Tang, S.-M., Su, X.-J., & Luo, Z.-L. (2024). Morphological and Phylogenetic Analyses Reveal Three Novel Species of Sanguinoderma (Ganodermataceae, Basidiomycota) from Yunnan Province, China. Journal of Fungi, 10(8), 589. https://doi.org/10.3390/jof10080589