Abstract
Entoloma is a relatively large genus in Agaricales, with a rich diversity of species and a wide distribution. In this study, seven new species of Entoloma belonging to the subgenus Cubospora have been identified based on morphological and phylogenetic evidence from subtropical regions of China. Morphologically, E. excavatum is characterized by the yellow, depressed, estriate pileus and medium-sized basidiospores; E. lacticolor is recognized by the white and papillate pileus, adnexed lamellae, and presence of clamp connections; E. phlebophyllum is identified by the pink-to-maroon and estriate pileus, and lamellae with lateral veins; E. rufomarginatum differs from other cuboid-spored species by the lamellae edge which is red-brown-underlined; E. subcycneum is characterized by the white pileus and carneogriseum-type cheilocystidia; E. submurrayi is recognized by the pileus margin exceeding the lamellae, 2-layered pileipellis with hyphae of different widths, and the presence of clamp connections; E. tomentosum is identified by the tomentose pileus, heterogeneous lamella edge, and versiform cheilocystidia with brown-yellow contents. Their distinct taxonomic status is confirmed by the positions of the seven new species in both the ITS + LSU and 3-locus (LSU, tef-1α, rpb2) phylogenetic trees. Detailed descriptions, color photos, and a key to related species are presented.
1. Introduction
Entoloma (Fr.) P. Kumm., with E. sinuatum (Bull.) P. Kumm designated as the type species, was established by Kummer in 1871 [1]. It is characterized by the pink spore prints and angular basidiospores viewed from any side [2]. The species of Entoloma are worldwide, encompassing habitats from the frigid zone to the tropics, alpine to basins, with the majority being saprobic on shady and humid ground or mosses in the forests [3,4]. So far, approximately 1800 species of Entoloma have been reported in the world [5,6] (https://www.speciesfungorum.org (accessed on 27 May 2024)).
In the Entoloma sensu lato, some species with cuboid basidiospores (with six, more or less equal, quadrangular faces, and a dihedral base) are notable due to the unique basidiospores morphology. More than 170 such species are reported worldwide, mainly occurring in the tropical and subtropical regions [7]. Traditionally, these species were classified in the subgenus Nolanea section Staurospori [4] and subgenus Inocephalus section Staurospora [8]. To clarify the taxonomic position of cuboid-spored species within Entoloma, analyses of phylogeny and basidiospore morphology were performed by Arstedt et al. [9]. It was found that these species formed two well-supported branches and fall into two separate subgenera: Cubospora Karstedt, Capelari, Largent, T.J. Baroni and Bergemann with type species E. luteolamellatum (Largent and Aime) Blanco-Dios and Cuboeccilia Karstedt, Capelari, and Largent with type species E. omphalinoides (Largent) Blanco-Dios [9].
In China, approximately 150 species of the Entoloma have been reported, with about 20 of these featuring cuboid spores, and 8 of these have been published as new species [7]. During our investigation for Entoloma in subtropical regions of China, some species of Entoloma with cuboid basidiospores were found, but do not match the known species. Based on morphological comparisons and phylogenetic analysis, some unknown-to-science cuboid-spored species were further confirmed, with seven new taxa recorded in this paper.
2. Materials and Methods
2.1. Morphological Studies
Specimens were collected from Fujian, Jiangxi, and Zhejiang provinces of China between 2020 and 2022, and were deposited in the Herbarium of Fungi, Jiangxi Agricultural University (HFJAU). Macroscopic characteristics were recorded from fresh specimens. The color codes referred to the Methuen Handbook of Colour [10]. Micromorphological structures were observed and measured under an Olympus BX53 microscope (Olympus corporation, Tokyo, Japan) by making squash preparations of sections of dried specimens that were placed in 5% KOH solution or H2O, and 1% Congo red was used as the staining agent when observing colorless tissues. Melzer’s reagent was selected for determining whether the spores were amyloid or not [11]. At least 20 basidiospores, basidia, and cystidia were measured for each collection. The range of spore size is expressed as the form (a) b–c (d), in which “a” and “d” representing the minimum and maximum values, and 90% of the spores falling within the range ‘b–c’. The meanings of the other spore characteristics are as follows: “Q” stands for the ratio of length and width; “av” symbolizes average value; “n” means number; and “Qm” indicates average “Q” ± standard deviation [12]. The morphological description is based on the work of Noordeloos et al. [13].
2.2. DNA Extraction, PCR Amplification, and Sequencing
Genomic DNA was extracted from dried specimens with the NuClean Plant Genomic DNA kit (CWBIO, China) [14]. The ITS, LSU, tef-1α, and rpb2 regions were amplified respectively using the primer pairs of ITS1F/ITS4, LR0R/LR5 [15], EF983F/EF1953R [2], and rpb2-i6f-RhoF1/rpb2-RhoR1 [16].
PCR amplification was conducted with a 25 μL reaction system as follows: 1 µL DNA, 2 µL primers, 9.5 µL ddH2O, and 12.5 µL 2 × Taq Master Mix (Dye Plus). For ITS, PCR was carried out using a touchdown amplification procedure: initial 95 °C for 5 min, and then 14 cycles of denaturing at 95 °C for 30 s, annealing at 65 °C for 45 s (−1 °C per cycle), extension at 72 °C for 1 min, and then 30 cycles of denaturing at 95 °C for 30 s, annealing at 52 °C for 30 s, and extension at 72 °C for 1 min, with the final extension at 72 °C for 10 min [17]. For others, the procedure was initial 98 °C for 5 min, and then 8 cycles of denaturing at 98 °C for 5 s, annealing at 61 °C for 40 s (−1 °C per cycle), extension at 72 °C for 2 min, and then 35 cycles of denaturing at 98 °C for 5 s, annealing at 54 °C for 1.5 min, extension at 72 °C for 2 min, with the final extension at 72 °C for 10 min. The PCR products were sequenced by Qing Ke Biotechnology Co. Ltd. (Wuhan City, China).
2.3. Alignment and Phylogenetic Analyses
Since the ITS data are absent for many species of E. subgenus Cubospora, two phylogenetic trees based on ITS + LSU and LSU + tef-1α + rpb2, were constructed by Bayesian inference (BI) and Maximum likelihood (ML), respectively, according to the previous studies of Karstedt et al. [9] and Morozova and Pham [18]. Some species of E. subgenus Nolanea are designated as outgroups. Information on specimens and GenBank accession numbers are listed in Table 1. ITS, LSU, tef-1α, and rpb2 sequence datasets were separately aligned on the MAFFT online server [19]. BI and ML phylogenetic analyses of the processed sequences were run using Mrbayes v.3.2.7a and IQtree v.2.1.2, respectively [20]. The best-fit models of ML and BI were determined by PartitionFinder [21] complying with Corrected Akaike information criterion (AICc). For the ML analysis, 1000 replicates are performed based on the ultrafast bootstrap option of ML that allowed partitions from different seeds. For the BI analysis, the genes chains were run for 2,000,000 generations. The first 25% of trees were discarded as burn-in. The branches of Bayesian posterior probability (BI-PP) ≥ 0.95 and ML bootstrap support (ML-BP) ≥ 75% are considered as statistical supports. The identifying criteria for new species are, according to the viewpoints proposed by Dettman et al., exhibiting 1 to 2 stable morphological differences from similar species and forming separated and stable clades in the phylogenetic tree [22].

Table 1.
Details of sequences used in the phylogenetic analyses. Newly generated sequences are in bold.
3. Results
3.1. Phylogenetic Analysis
In total, 243 sequences of 96 samples were used for phylogenetic analyses. For the IL (ITS + LSU) tree, a total of 2135 characters were used in the analyses, of which 1494 were constant, 496 were parsimony-informative, and 145 were singleton. The best-fit models of both ML and BI were the same: GTR + F + I + G4 for ITS and HKY + F + I + G4 for LSU. For the LTR (LSU + tef-1α + rpb2) tree, a total of 3033 characters were used in the analyses, of which 1998 were constant, 809 were parsimony-informative, and 226 were singleton. The best-fit models of both ML and BI were the same: GTR + F + I + G4 for LSU and tef-1α, SYM + I + G4 for rpb2. For Bayes analysis, the average standard deviation of split frequencies is less than 0.01 after 650,000 generations.
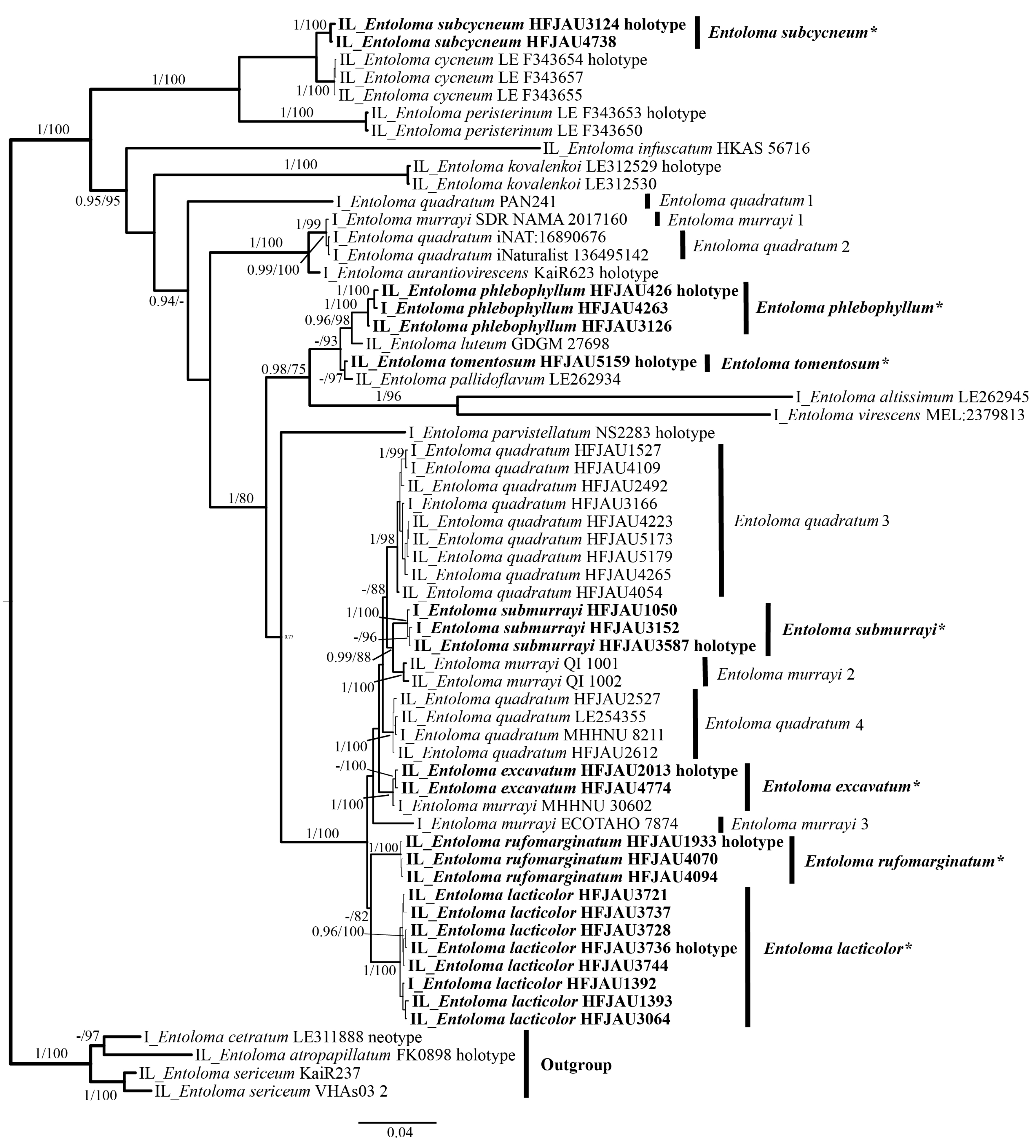
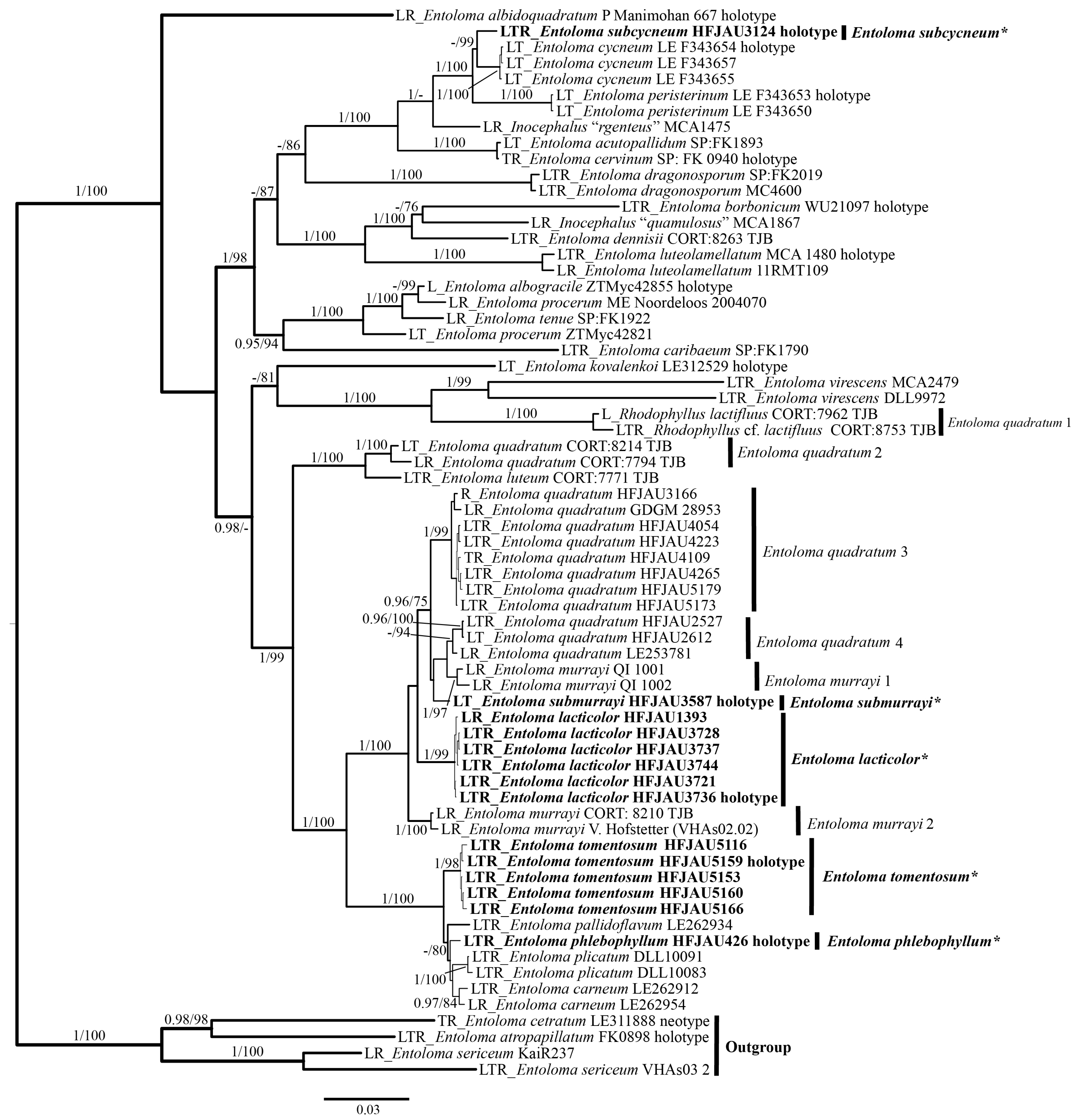
The results of the phylogenetic analysis are shown in the Figure 1 (IL tree) and Figure 2 (LTR tree). The overall frameworks were consistent with previous studies [9]. Seven new species were all clustered in the subgenus Cubospora clade and formed separated and stable branches, respectively. In the IL tree, E. excavatum, E. lacticolor, and E. rufomarginatum formed distinct lineages, separately (BI-PP = 1, ML-BP = 100%). E. submurrayi formed a sister lineage with E. murrayi (Berk. and M.A. Curtis) Sacc. and P. Syd. (BI-PP = 1, ML-BP = 100%). E. phlebophyllum formed a sister lineage with E. tomentosum (BI-PP = 1, ML-BP > 95%), and E. subcycneum formed a sister lineage with E. cycneum O.V. Morozova and T.H.G. Pham (BI-PP = 1, ML-BP = 100%). In the LTR tree, E. lacticolor, E. phlebophyllum, E. subcycneum, and E. tomentosum formed distinct lineages, separately (BI-PP > 0.95, ML-BP ≥ 75%). E. subcycneum formed a sister lineage with E. cycneum and E. peristerinum O.V. Morozova and T.H.G. Pham (BI-PP = 1, ML-BP = 100%).
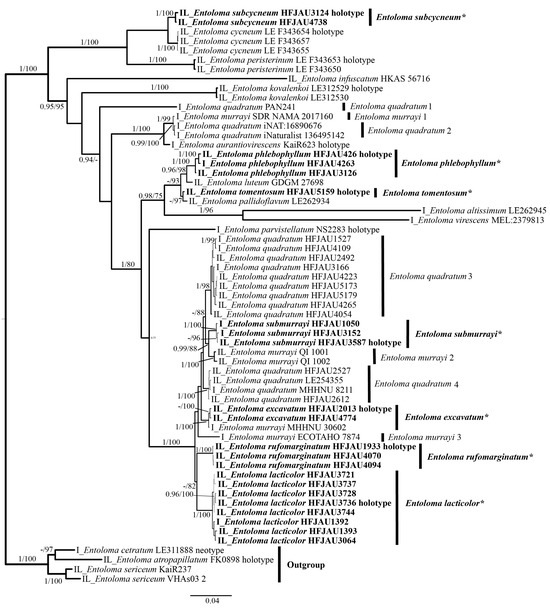
Figure 1.
Phylogram of Entoloma subgenus Cubospora spp. generated by Bayesian inference (BI) analysis based on ITS (I) and LSU (L), rooted with E. subgenus Nolanea spp. Bayesian inference (BI-PP) ≥ 0.95 and ML bootstrap proportions (ML-BP) ≥ 75 are indicated as PP/BP. The new taxa are marked in bold and *.
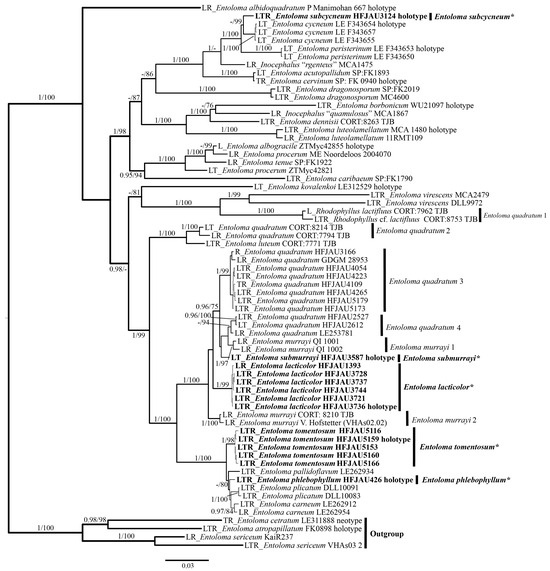
Figure 2.
Phylogram of Entoloma subgenus Cubospora spp. generated by Bayesian inference (BI) analysis based on LSU (L), tef-1α (T), and rpb2 (R), rooted with E. subgenus Nolanea spp. Bayesian inference (BI-PP) ≥ 0.95 and ML bootstrap proportions (ML-BP) ≥75 are indicated as PP/BP. The new taxa are marked in bold and *.
3.2. Taxonomy
MycoBank: MB854072

Figure 3.
Photos of basidiomata. (A) Entoloma excavatum: HFJAU2013, holotype; (B,C) Entoloma lacticolor: (B) HFJAU3064, (C) HFJAU3736, holotype; (D–F) Entoloma phlebophyllum: (D,E) HFJAU4261, holotype; (F) HFJAU4263; (G) Entoloma rufomarginatum: HFJAU1933, holotype. Scale bars: (A,D–G) 20 mm; (B,C) 30 mm.
Figure 3.
Photos of basidiomata. (A) Entoloma excavatum: HFJAU2013, holotype; (B,C) Entoloma lacticolor: (B) HFJAU3064, (C) HFJAU3736, holotype; (D–F) Entoloma phlebophyllum: (D,E) HFJAU4261, holotype; (F) HFJAU4263; (G) Entoloma rufomarginatum: HFJAU1933, holotype. Scale bars: (A,D–G) 20 mm; (B,C) 30 mm.


Figure 4.
Photos of basidiomata. (A,B) Entoloma subcycneum: (A) HFJAU0985, (B) HFJAU3124, holotype; (C,D) Entoloma submurrayi: (C) HFJAU1050, (D) HFJAU3587, holotype; (E,F) Entoloma tomentosum: (E) HFJAU5159, holotype, (F) HFJAU5166. Scale bars: (A–F) 20 mm.
Figure 4.
Photos of basidiomata. (A,B) Entoloma subcycneum: (A) HFJAU0985, (B) HFJAU3124, holotype; (C,D) Entoloma submurrayi: (C) HFJAU1050, (D) HFJAU3587, holotype; (E,F) Entoloma tomentosum: (E) HFJAU5159, holotype, (F) HFJAU5166. Scale bars: (A–F) 20 mm.

Etymology. Refer to Latin “excavatus” (excavate)—the pileus is excavated.
Holotype. China, Zhejiang Province, Lishui City, Jiulongshan Reserve in Suichang County, 28°23′23″ N, 118°51′0″ E, alt. 535 m, 14 July 2020, collected by Yu-Peng Ge, Bin-Ron Ke, and Zhi-Heng Zeng, HFJAU2013.
Diagnosis. Entoloma excavatum is mainly characterized by the rather small and mycenoid basidiomata; conical to broadly conical with a depression at center; bright yellow, estriate, and glabrous pileus; relatively sparse, adnexed, and subventricose lamellae with crenulate edge; glabrous and striae stipe; cuboid basidiospores; sterile lamellae edge; cylindrical cheilocystidia; presence of clamp connections. It differs from E. overeemii E. Horak by its mycenoid basidiomata, darker estriate pileus, adnexed lamellae, larger basidiospores, cylindrical cheilocystidia, and presence of clamp connections.
Description. Basidiomata, rather small, mycenoid. Pileus 13–22 mm wide, conical to broadly conical with a depression at the center, glabrous or sparsely fibrillous, not translucently striate, with entire or serrate margin, bright yellow (3C7–8). Lamellae relatively sparse, 1.5–2.0 mm wide, with two types of lamellules, adnexed to sinuate, ventricose, concolored with cap, with serrate and concolorous edge. Stipe 30–50 × 2.0–3.0 mm, central, terete, hollow, equal, concolorous with the pileus, slightly with longitudinal or oblique striae, smooth and glabrous, base with white tomentum. Context is thin, concolorous to the surface. Odor is indistinct, taste is not tested.
Basidiospores are (8.0) 8.5–10.5 × 8.0–10.0 (10.5) μm, (av = 9.3 × 8.8 μm), Q = 1.0–1.1 (1.2) (Qm = 1.05 ± 0.03, n = 60), isodiameterical, cuboid, sporadically with five angles in side-view, thick-walled, inamyloid. Basidia are 40–52 × 11–13 μm, clavate, 4-spored, sterigmata 8.0–11 μm long, clamped. Pleurocystidia is absent. Cheilocystidia are 50–96 × 8.0–13 μm, serrulatum-type, irregular clusters in the sterile lamellae edge, cylindrical, with rounded apex. Lamellar trama are regular, made up of cylindrical hyphae 5.0–13 µm wide, with oleiferous hyphae near to the margin. Pileipellis is a cutis with transitions to a trichoderm towards the margin, made up of cylindrical hyphae 5.0–11 μm broad, thin-walled, even at septa, with rounded end and sparsely pale yellow encrusting pigment, easily dissolved in KOH solution. Stipitipellis is a cutis composed of cylindrical hyphae 5.0–9.0 μm wide, slightly constricted at septa. Clamp connections are present and abundant in all parts of the basidiocarp.
Habitat. Scattered on soil in mixed coniferous-broad-leaved forest.
Distribution. China.
Additional specimens examined: China, Zhejiang Province, Lishui City, Jiulongshan Reserve in Suichang County, 28°23′23″ N, 118°51′2″ E, alt. 496 m, 14 July 2020, collected by Yu-Peng Ge, Bin-Ron Ke, and Zhi-Heng Zeng, HFJAU4774.
Notes. In the IL trees, this specimen E. murrayi MHHNU 30,602 should be E. excavatum, since it shares 99.3% similarity with our species in ITS and clusters into a stable branch (BI-PP = 1, ML-BP = 100%). Additionally, E. murrayi and E. quadratum (Berk. and M.A. Curtis) E. Horak are closely related to the new species. E. murrayi differs from E. excavatum by the striate pileus without a depression at the center and a lack of clamp connections [38]. E. quadratum is distinguished by its orange-yellow to salmon and striate pileus without a depression at the center [38]. Morphologically, E. excavatum shares many features with E. overeemii, including the yellow, depressed umbilicate, glabrous pileus and the cuboid basidiospores. However, E. overeemii is distinguished by the omphaloid basidiomata, translucently striate pileus, adnate to decurent lamellae, smaller basidiospores (5.0–7.0 μm), clavate to vesiculose cheilocystidia, and lack of clamp connections [38].
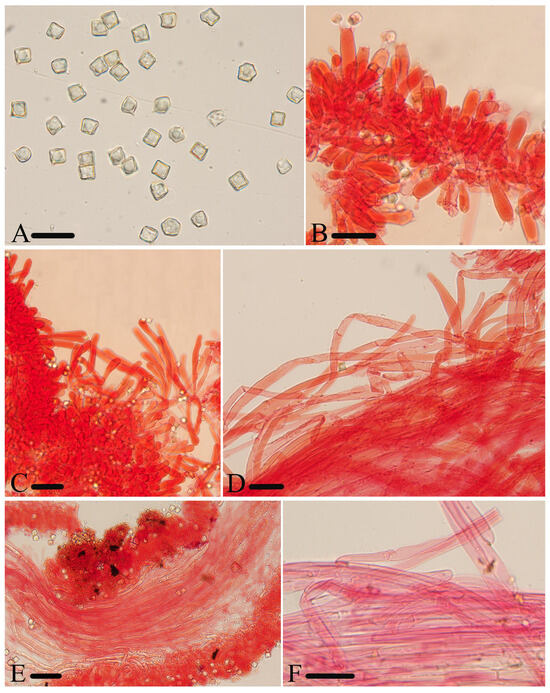
Figure 5.
Micromorphological structures of Entoloma excavatum. (A) Basidiospores. (B) Basidia. (C) Cheilocystidia. (D) Pileipellis. (E) Lamellar trama. (F) Stipitipellis. Scale bars: (A–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Figure 5.
Micromorphological structures of Entoloma excavatum. (A) Basidiospores. (B) Basidia. (C) Cheilocystidia. (D) Pileipellis. (E) Lamellar trama. (F) Stipitipellis. Scale bars: (A–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
MycoBank: MB850724
Etymology. Refer to Latin “lacteus” (milky white)—the basidiomata is entirely milk-white.
Holotype. China, Fujian Province, Wuyishan City, Zhongpeng Village, 27°55′18″ N, 117°51′13″ E, alt. 674 m, 25 June 2022, collected by Yu-Peng Ge and Meng-Hui Han, HFJAU3736.
Diagnosis. Entoloma lacticolor is mainly characterized by the milky white and glabrous pileus with obvious acute papilla; serrate lamellae edge; glabrous or furfuraceous-scaly stipe with longitudinal or oblique groove; cuboid basidiospores; presence of clamp connections. It differs from E. album Hiroë by the cylindrical to subclavate cheilocystidia and presence of clamp connections.
Description. Basidiomata are small to medium-sized. Pileus is 15–50 mm wide, conical when young, from hemispherical to flattened with age, with obvious acute papilla at the center, not hygrophanous, smooth, glabrous, marked by translucently radial striae almost up to 2/3 of the radius, with slightly cracking, straight, glabrous or squamulose margin, milky white (1A1–3A1), yellowish at center. Lamellae are medium density, 2.0–8.0 mm wide, with two types of lamellules, adnexed, ventricose, originally white, becoming pink (9A3), with a serrulate and concolourous edge. Stipe are 22–110 × 2.5–15 mm, central, cylindrical, hollow, equal, concolored with cap, with longitudinal or oblique groove, glabrous or furfuraceous-scaly, base with white tomentum. Smell and taste are indistinct.
Basidiospores are (7.5) 8.0–10.0 (10.5) × (7.0) 7.5–9.0 (10.0) μm, (av = 8.7 × 8.2 μm), Q = 1.0–1.1 (1.2) (Qm = 1.05 ± 0.04, n = 80), isodiameterical, cuboid, sporadically with five angles in side-view, thick-walled, inamyloid. Basidia are 35–53 × 10–14 μm, clavate, 4-spored, sterigmata 6.0–14 μm long, clamped. Pleurocystidia are absent. Cheilocystidia are 33–75 × 7.0–14 μm, carneogriseum-type, dispersed along the sterile lamellae edge, cylindrical to subclavate, septate, with a rounded apex. Lamellar trama is regular, made up of cylindrical hyphae 9.0–15 µm wide, intertwined by refractive hyphae near the margin. Pileipellis is a cutis to a trichoderm of cylindrical hyphae 5.0–11 μm broad, thin-walled, slightly constricted at septa, without pigment. Stipitipellis is a cutis composed of cylindrical hyphae, up to 13 μm wide, slightly constricted at septa. Clamp-connections are present in all tissue.
Habitat. Scattered or solitary on soil in mixed coniferous-broad-leaved forest.
Distribution. China
Additional specimens examined. China, Fujian Province, Wuyishan City, 27°50′9″ N, 117°46′25″ E, alt. 1905 m, 11 August 2021, collected by Jun-Qing Yan and Ze-Wei Liu, HFJAU1392, HFJAU1393, HFJAU3064; Wuyishan City, Zhongpeng Village, 27°55′15″ N, 117°51′12″ E, alt. 695 m, 25 June 2022, collected by Yu-Peng Ge, and Meng-Hui Han, HFJAU3721, HFJAU3728, HFJAU3737, HFJAU3744.
Notes. In the IL trees, E. rufomarginatum is rather close to E. lacticolor. But, the former differs in its yellow pileus, lamellae edge red-brown underlined, and shows only 94.8% similarity with new species in the ITS sequence. Morphologically, some species have aspects of E. lacticolor with cuboid basidiospores and white basidiomata, but can be separated as follows. E. albidoquadratum Manim. and Noordel. have pleurocystidia and larger basidiospores (11–16.5 × 9–14 μm) [39]. E. cycneum differs in its pileus without obviously acute papilla at the center [18]. E. peristerinum differs in its porphyrogriseum-type cheilocystidia [18]. E. minutoalbum E. Horak is a common species of New Zealand, and has smaller basidiospores (6–8 μm) [38]. E. subcycneum is distinguished by its smaller pileus and ITS region, with a similarity of 93.8%.
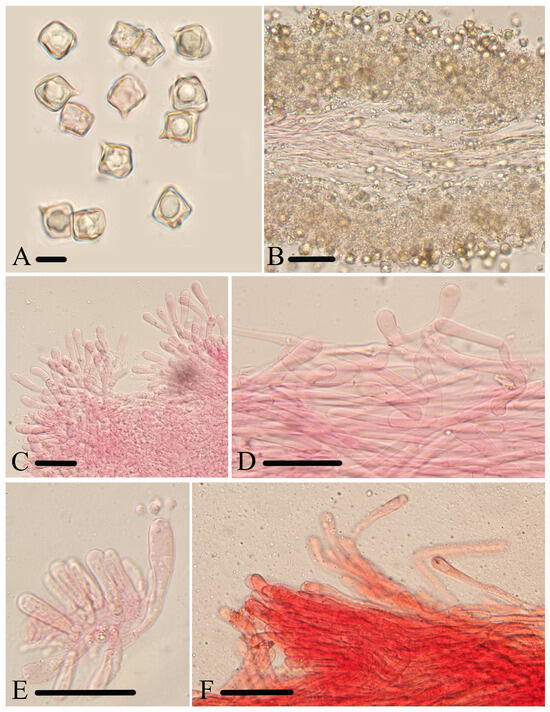
Figure 6.
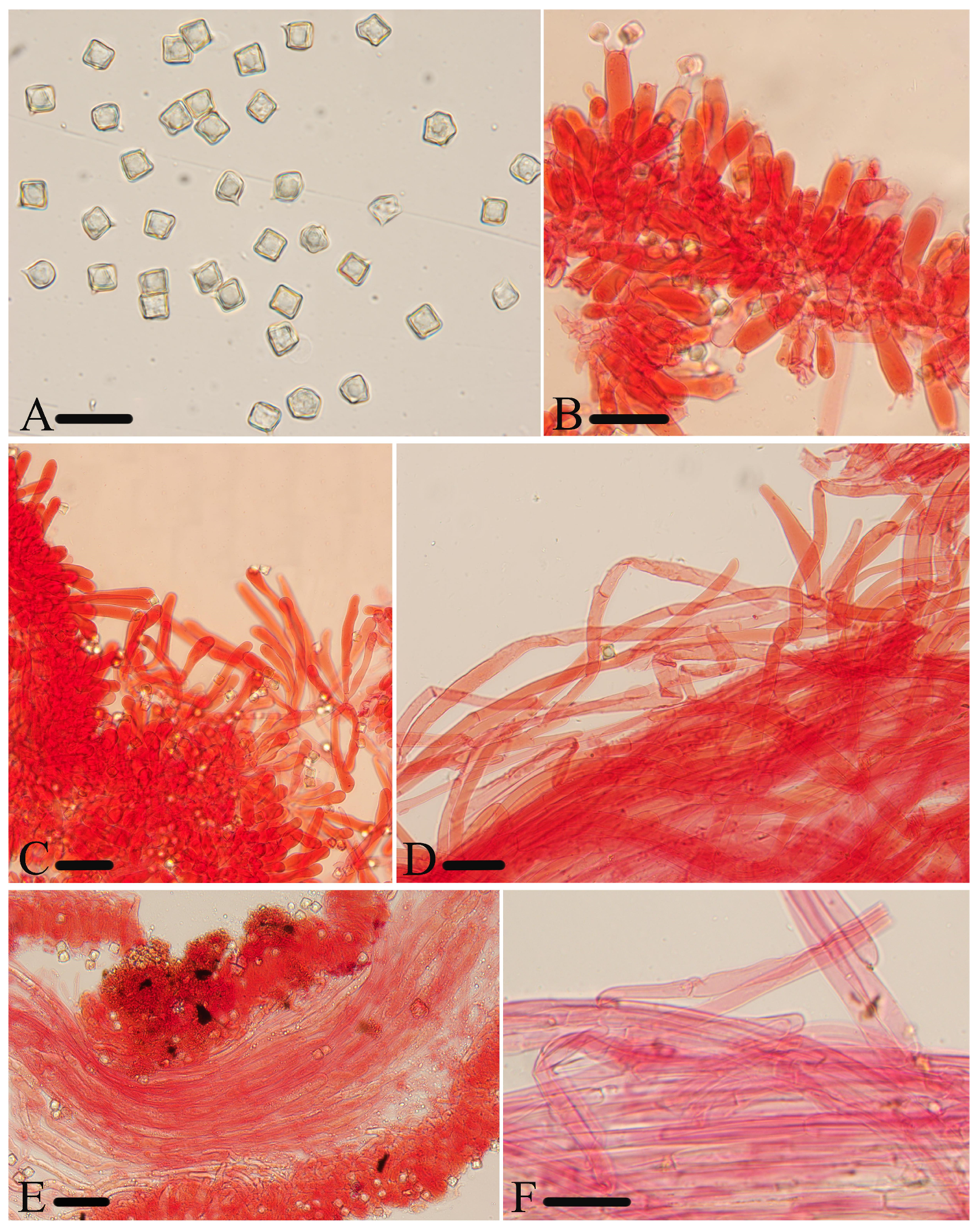
Micromorphological structures of Entoloma lacticolor. (A) Basidiospores. (B) Lamellar trama. (C) Cheilocystidia. (D) Stipitipellis. (E) Basidia. (F) Pileipellis. Scale bars: (A) 10 μm; (B–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A,B).
Figure 6.
Micromorphological structures of Entoloma lacticolor. (A) Basidiospores. (B) Lamellar trama. (C) Cheilocystidia. (D) Stipitipellis. (E) Basidia. (F) Pileipellis. Scale bars: (A) 10 μm; (B–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A,B).
MycoBank: MB851118.
Etymology. According to the features of lamellae—having tiny lateral veins.
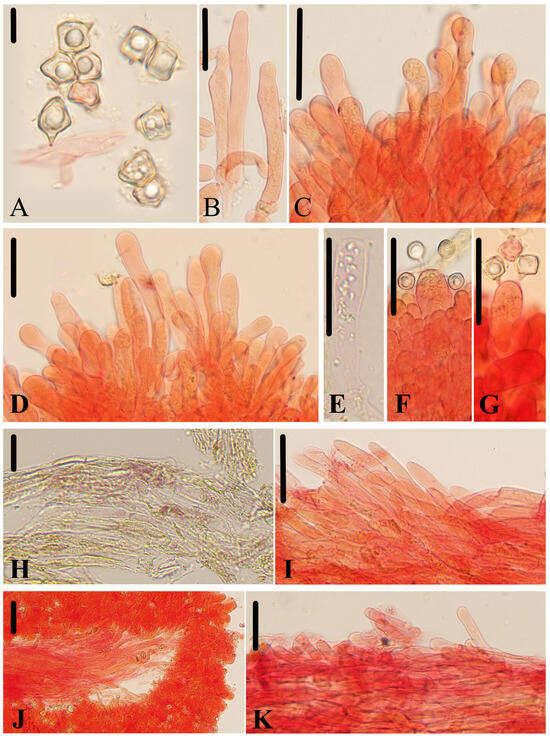
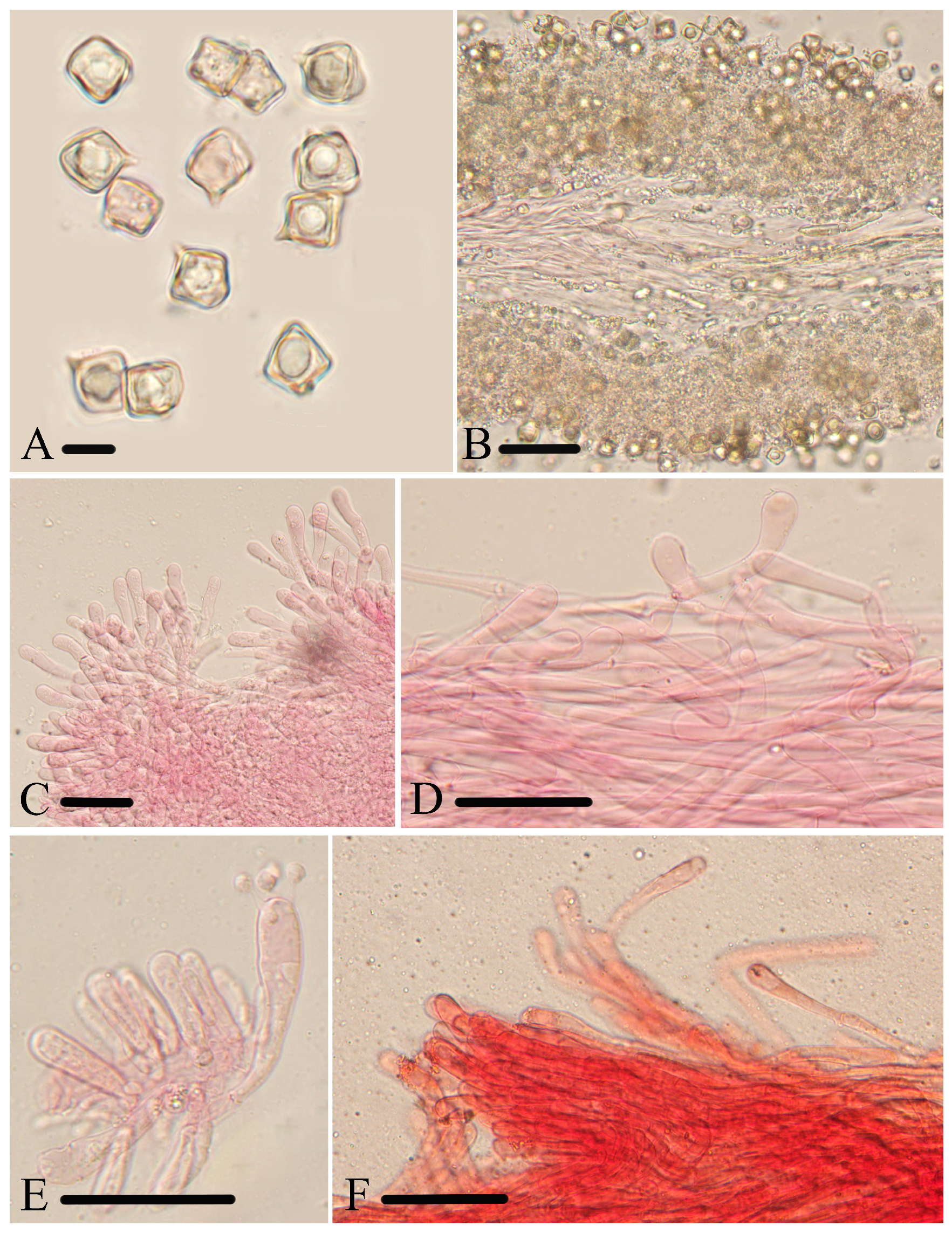
Figure 7.
Micromorphological structures of Entoloma phlebophyllum. (A) Basidiospores. (B–D) Cheilocystidia. (E–G) Basidia. (H,I) Pileipellis. (J) Lamellar trama. (K) Stipitipellis. Scale bars: (A) 10 μm; (B–K) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (H).
Figure 7.
Micromorphological structures of Entoloma phlebophyllum. (A) Basidiospores. (B–D) Cheilocystidia. (E–G) Basidia. (H,I) Pileipellis. (J) Lamellar trama. (K) Stipitipellis. Scale bars: (A) 10 μm; (B–K) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (H).
Holotype. China, Fujian Province, Tongmuguan Pass, 27°41′45″ N, 117°49′20″ E, alt. 391 m, 11 July 2022, collected by Jun-Qing Yan and Cheng-Feng Nie, HFJAU4261.
Diagnosis. E. phlebophyllum is recognized by the conical-to-plano convex with a depression, pink-to-maroon and glabrous pileus; adnate to decurrent lamellae with obviously tiny lateral veins and a serrulate edge; stipe with a different color from the cap; cuboid basidiospores; sterile lamellae edge; cylindrical or clavate-capitate cheilocystidia; and the presence of clamp connections. It differs from E. phleboides (Romagn.) E. Horak by the pink-to-maroon pileus, presence of cystidia, and clamp connections.
Description. Basidiomata are rather small. Pileus are 6.0–19 mm wide, broadly conical when young, convex-to-plano convex with age, with a depressed center, not hygrophanous and translucently striate, glabrous or tomentulose towards the entire and slightly inrolled margin, squamous at the center, with serrate or straight margin, pink (8A4–9A3), dark carneous (8A3–8B3) to maroon (10E8–10E7), and darker at the center. Lamellae are medium density and 2.0–3.0 mm wide, with tiny lateral veins and three types of lamellules, adnate to decurrent, originally white, becoming pink, with serrulate and a concolourous or paler edge. Stipe are 12–43 × 1.5–2.5 mm, central, cylindrical, hollow, equal or attenuated upwards, white to dirty white, glabrous or sparsely covered by white-fibrillous scales, somewhat longitudinally striate, and white tomentose at the base. Context is thin and white. Smell and taste are indistinct.
Basidiospores are (7.0) 7.5–10.0 (11.0) × (6.5) 7.0–9.0 (10.0) μm, (av = 8.5 × 8.0 μm), Q = 1.0–1.2 (1.3) (Qm = 1.07 ± 0.06, n = 80), isodiameterical or subisodiameterical, cuboid, rarely 3 or 5 angles in side-view, thick-walled, and inamyloid. Basidia are 40–55 × 11–14 μm, clavate, 4- or 2-spored, sterigmata 5.0–11 μm long, and clamped. Pleurocystidia are absent. Cheilocystidia are 37–73 × 6.0–12 μm, serrulatum-type, irregular clusters in the sterile lamellae edge, cylindrical, clavate, septate, often with rounded apex, and sometimes with tapered apex or constricted neck. Lamellar trama are regular, made up of cylindrical hyphae 4.0–10 µm wide, and intertwined by oleiferous hyphae near the margin. Pileipellis is a cutis to a trichoderm of cylindrical hyphae 5.0–13 μm broad, thin-walled, slightly constricted at septa, and with a tapered or pointed end and brownish yellow encrusting pigment and brick red intracellular pigment. Stipitipellis is a cutis composed of densely arranged, cylindrical hyphae, up to 10 μm wide, and slightly constricted at the septa. Brilliant granules are abundant and clamp connections are present in all tissue.
Habitat. Scattered on soil in mixed coniferous-broad-leaved forest.
Distribution. China.
Additional specimens examined. China, Fujian Province, Wuyi Mountain, 27°42′37″ N, 117°51′30″ E, alt. 530 m, 13 August 2021, collected by Jun-Qing Yan and Ze-Wei Liu, HFJAU3126; Fujian Province, Tongmuguan Pass, 27°41′48″ N, 117°49′19″ E, alt. 394 m, collected by Jun-Qing Yan and Cheng-Feng Nie, HFJAU4263.
Notes. E. phlebophyllum groups together with E. luteum Peck in the IL tree, and groups together with E. carneum Z.S. Bi, E. pallidoflavum (Henn. and E. Nyman) E. Horak, and E. plicatum (Largent) in the LTR tree. However, the last four species do not have tiny lateral veins between the lamellae. Apart from that, E. carneum differs in the striate pileus and shows only 98.5% similarity with E. phlebophyllum in tef-1α and rpb2 sequence, respectively [40,41]; E. luteum differs in its yellow and striate pileus, adnexed lamellae with fimbriate edge [38], and shows only 95.8% similarity with E. phlebophyllum in ITS sequence; E. pallidoflavum differs in the pale yellow and sulcate pileus [38], and shows only 95.5% similarity with E. phlebophyllum in ITS sequence; E. plicatum is distinguished by the yellow and plicate-striate pileus with mammillate umbo [34], and shows only 98% similarity with E. phlebophyllum in tef-1α and rpb2 sequence. In addition, morphologically, E. infundibuliforme Petch and E. significum Corner and E. Horak have aspects of E. phlebophyllum, but E. infundibuliforme differs in the smaller basidiospores (5.5–8 μm), presence of fusoid pleurocystidia, and lack of clamp connections [42]. E. significum is distinguished from new species by its absence of cystidia [38].
MycoBank: MB854068
Etymology. “Rufo-” means “red”, “marginata” means “margin” (Latin)—refers to the feature of the lamellae edge.
Holotype. China, Zhejiang Province, Lishui City, Baishanzu in Qingyuan County, 27°45′26″ N, 117°11′59″ E, alt. 1586 m, 8 July 2020, collected by Yu-Peng Ge and Qin Nan, HFJAU1933.
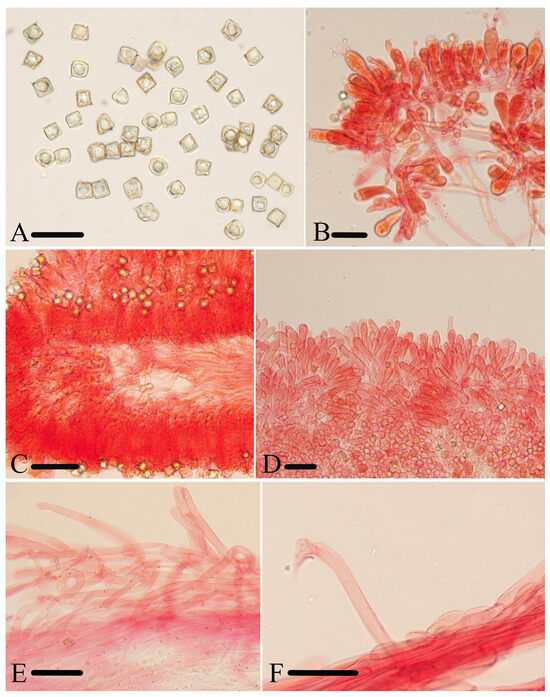
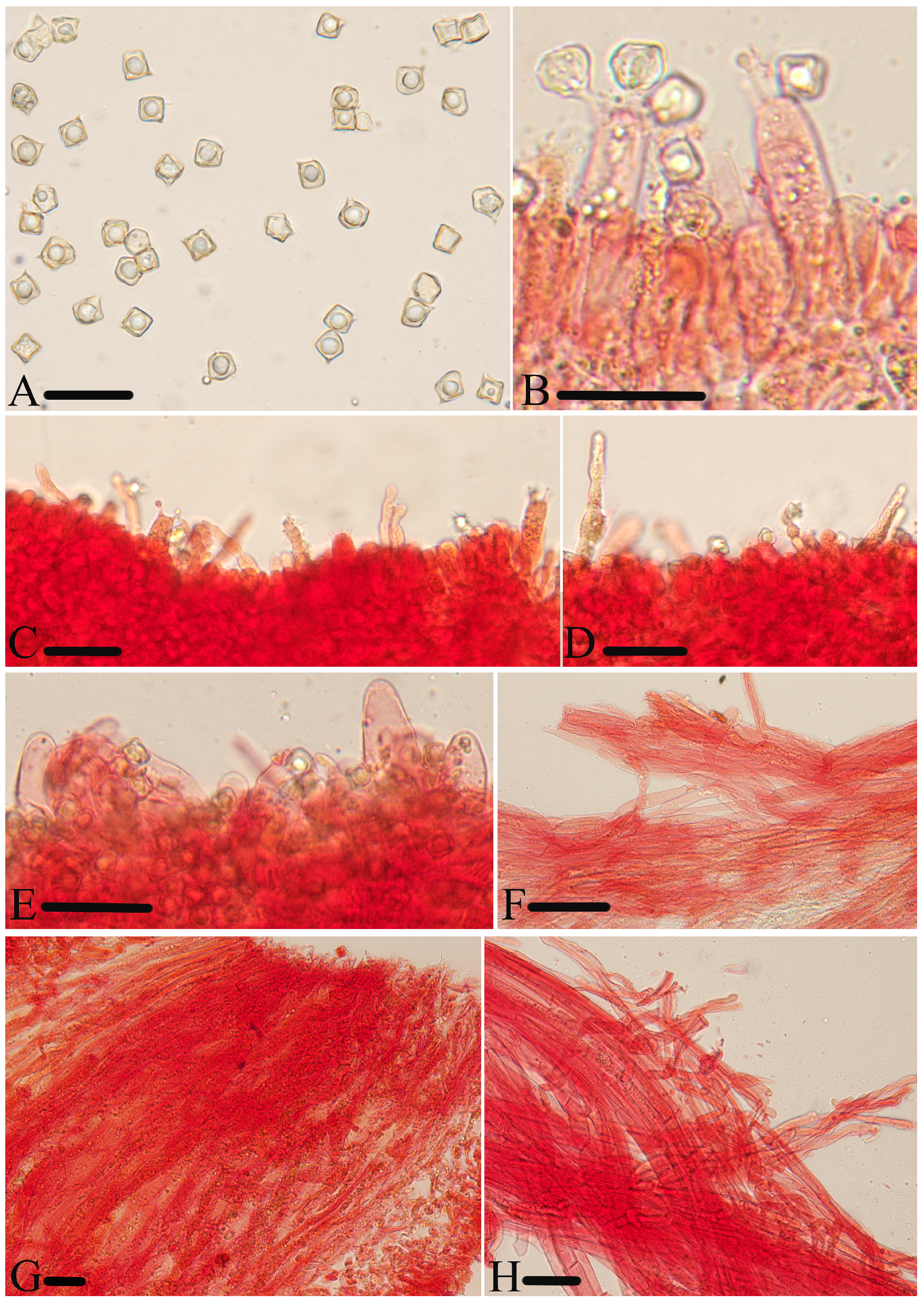
Figure 8.
Micromorphological structures of Entoloma rufomarginatum. (A) Basidiospores. (B) Basidia. (C) Lamellar trama. (D) Cheilocystidia. (E) Pileipellis. (F) Stipitipellis. Scale bars: (A–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Figure 8.
Micromorphological structures of Entoloma rufomarginatum. (A) Basidiospores. (B) Basidia. (C) Lamellar trama. (D) Cheilocystidia. (E) Pileipellis. (F) Stipitipellis. Scale bars: (A–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Diagnosis. Entoloma rufomarginatum is mainly characterized by the rather small and mycenoid basidiomata, campanulate, or conical with acute papilla, dark brownish yellow, striae, and glabrous pileus; adnate and orange-brown lamellae with lamellae edge red-brown underlined; tomentose stipe; cuboid basidiospores; sterile lamellae edge; cylindric-clavate cheilocystidia; and presence of clamp connections. It differs from E. gracilius E. Horak by its lamellae edge red-brown underlined and larger basidiospores.
Description. Basidiomata are rather small, mycenoid. Pileus are 10–20 mm wide, campanulate or conical with obvious acute papilla, translucently striate at the margin up to the centre, slightly hygrophanous, smooth, glabrous, felted-scaly towards the entire and slightly inrolled margin, light blond (4C4–5), with green tints on margin. Lamellae are moderately distant, 1.5–2.0 mm wide, with two types of lamellules, adnate, emarginate, subventricose, orange-brown (6B4–5), with undulate, concolorous edge and red-brown underlined. Stipe are 30–55 × 4.0–8.0 mm, central, terete, hollow, equal or attenuated upwards, greenish-yellow (3CD4), tomentose, somewhat with longitudinal striae, base with white tomentum. Context is thin, concolorous to the surface. Odor is indistinct, taste is not tested.
Basidiospores are (8.0) 9.0–10.0 (12.5) × (8.0)8.5–10.0 (11.0) μm, (av = 9.8 × 9.3 μm), Q = 1.0–1.1 (1.2) (Qm = 1.05 ± 0.03, n = 100), isodiameterical, cuboid, thick-walled, inamyloid. Basidia are 39–62 × 11–15 μm, clavate or narrowly vesiculose, 4- or 2-spored, sterigmata 7.0–14 μm long, clamped. Pleurocystidia are absent. Cheilocystidia are 35–113 × 6.0–11 μm, serrulatum-type, irregular clusters in the sterile lamellae edge, cylindric-clavate, with rounded, rarely acute apex. Lamellar trama are regular, made up of cylindrical hyphae 5.0–18 µm wide. Pileipellis is a transition between cutis and trichoderm, made up of hyphae 9.0–16 μm wide, thin-walled, even at septa, with a rounded end. Stipitipellis is a cutis composed of cylindrical hyphae 5.0–12 μm wide, slightly constricted at the septa, with a rounded end. Clamp connections are present in all tissue.
Habitat. Scattered on soil in mixed coniferous-broad-leaved forest.
Distribution. China.
Additional specimens examined. China, Zhejiang Province, Lishui City, Baishanzu in Qingyuan County, 27°45′26″ N, 119°11′59″ E, alt. 1574 m, 8 July 2020, collected by Yu-Peng Ge and Qin Nan, HFJAU4070, HFJAU4094.
Notes. In the IL tree, E. lacticolor is closest to E. rufomarginatum. But, the former differs in the white basidiomata, lamellae edge is not underlined, and shows only 94.8% similarity with E. rufomarginatum in ITS sequence. In addition, E. avilanum (Dennis) E. Horak, E. kamerunense (Bres.) E. Horak, and E. submurrayi are rather close to E. rufomarginatum, but the former three exhibit concolored lamellae with the cap and the lamellae edge not underlined. Apart from those, E. avilanum from Venezuela differs from E. rufomarginatum by the free lamellae with a subfimbriate edge [38]. E. kamerunense is distinguished by the absence of cystidia, and the brown plasmatic pigment in pileipellis [38].
MycoBank: MB854069.
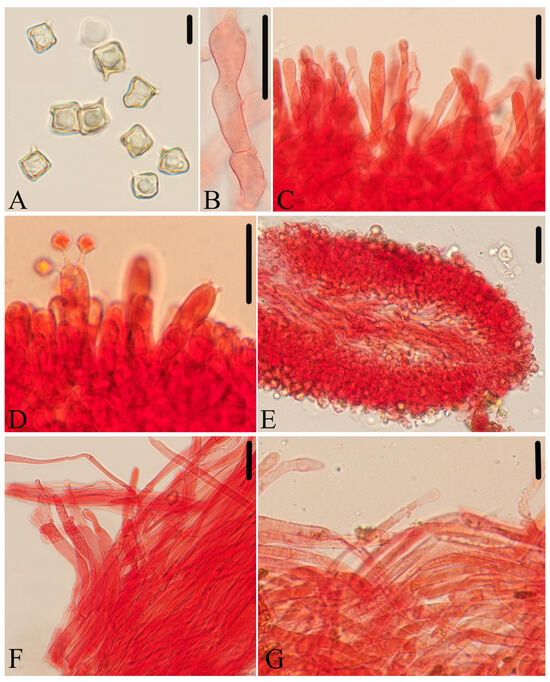
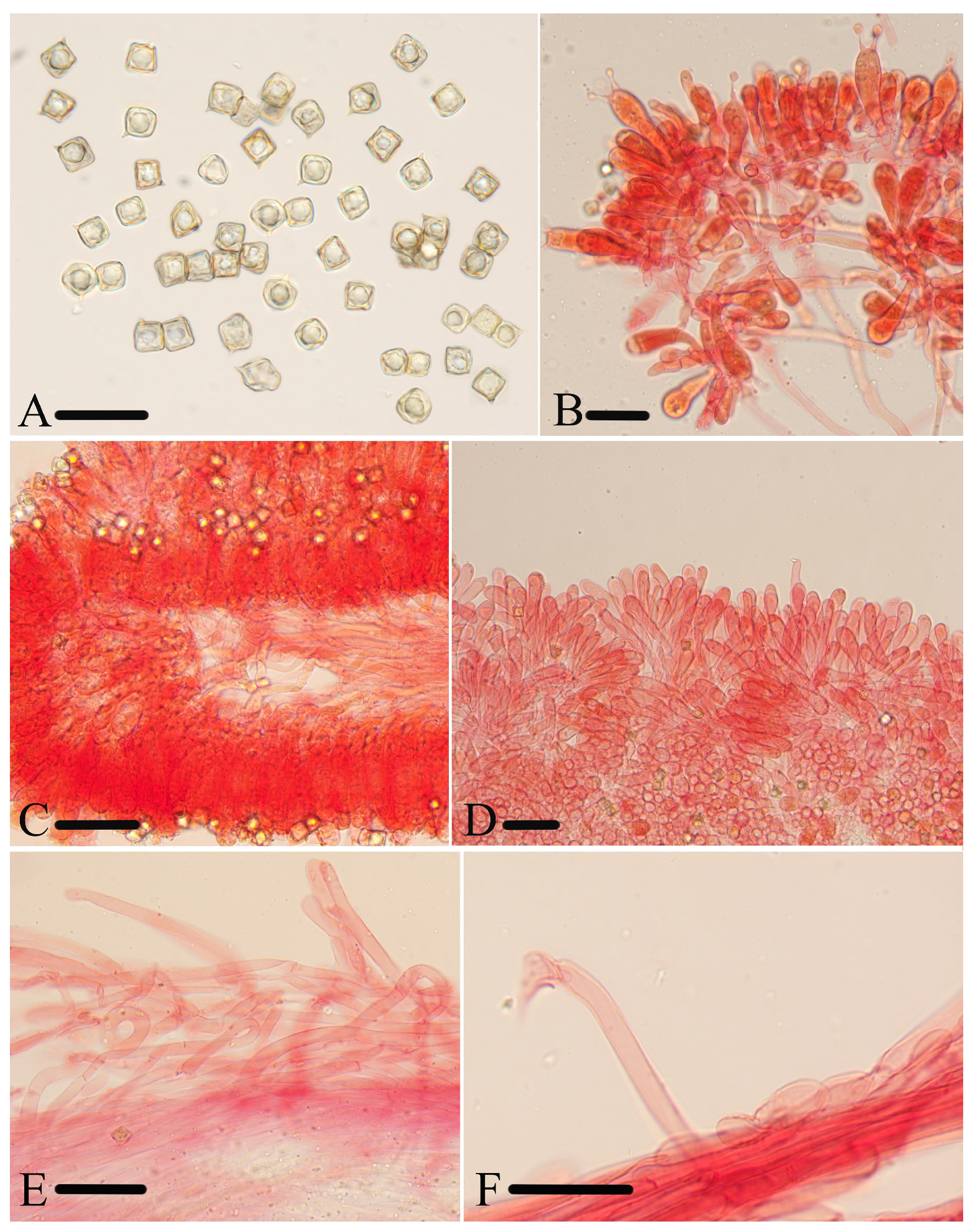
Figure 9.
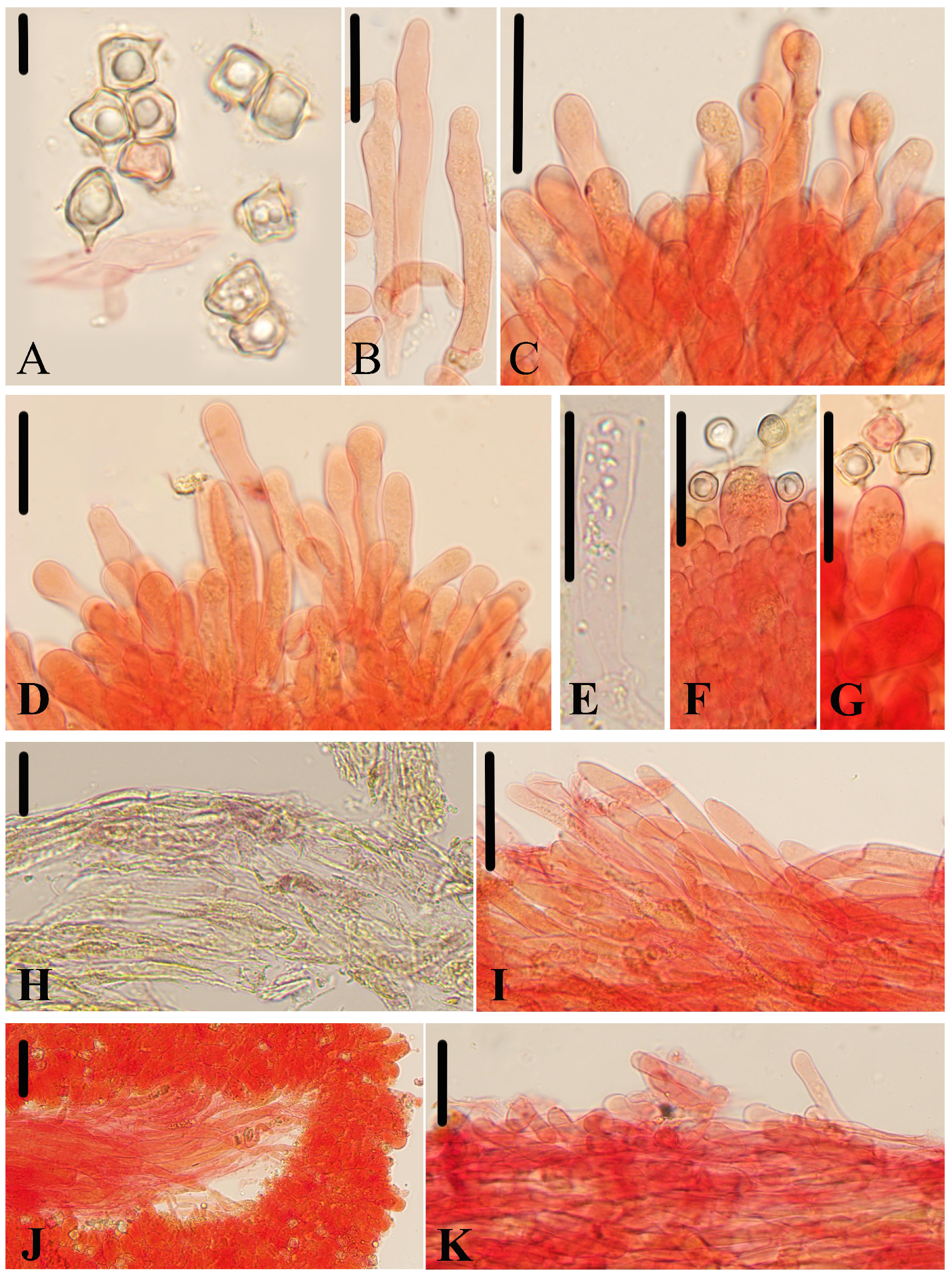
Micromorphological structures of Entoloma subcycneum. (A) Basidiospores. (B,C) Cheilocystidia. (D) Basidia. (E) Lamellar trama. (F) Pileipellis. (G) Stipitipellis. Scale bars: (A) 10 μm; (B–G) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Figure 9.
Micromorphological structures of Entoloma subcycneum. (A) Basidiospores. (B,C) Cheilocystidia. (D) Basidia. (E) Lamellar trama. (F) Pileipellis. (G) Stipitipellis. Scale bars: (A) 10 μm; (B–G) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Etymology. Macroscopic morphology similar to “Entoloma cycneum”.
Holotype. China, Fujian Province, Wuyishan City, 27°44′43″ N, 117°41′6″ E, alt. 792 m, 13 August 2021, collected by Jun-Qing Yan and Ze-Wei Liu, HFJAU3124.
Diagnosis. Entoloma subcycneum is mainly characterized by the rather small basidiomata; conical to applanate and pure white pileus; adnexed or sinuate lamellae; cuboid basidiospores; sterile lamellae edge; cylindric-clavate and septate cheilocystidia with various apex; and the presence of clamp connections. It differs from E. cycneum by its carneogriseum-type cheilocystidia and absence of brilliant granules.
Description. Basidiomata are rather small. Pileus are 8.0–15 mm wide, conical to hemispherical when young, becoming applanate with a small depression at the center, glabrous or erected velutinous, margin entire, sometimes undulating or cracking, when moist translucently striate up to 2/3 of the radius, pure white (3A1–4A1), with a slightly pink pigment (9A3–4). Lamellae are moderately distant, 1.5–3.5 mm wide, with two types of lamellules, adnexed, sinuate, ventricose, white to pink, with dentate, sometimes fimbriate, and concolorous edge. Stipe are 15–30 × 3–6 mm, central, terete, hollow, equal or attenuated towards the apex, concolored with cap, slightly with longitudinal striae, glabrous or densely tomentous, easily peeling, and have a base with a white tomentum. Context is thin, white. Odor is indistinct, taste is not tested.
Basidiospores are (8.0) 8.5–11.0 (11.5) × (7.0) 8.0–10.0 (11.0) μm, (av = 9.5 × 9.0 μm), Q = 1.0–1.1 (1.2) (Qm = 1.06 ± 0.04, n = 80), isodiameterical, cuboid, sporadically with five angles in side-view, thick-walled, inamyloid. Basidia are 41–54 × 11–14 μm, clavate, 4-spored, sterigmata 6.0–14 μm long, clamped. Pleurocystidia are absent. Cheilocystidia are 29–110 × 6.0–13 μm, carneogriseum-type, regularly dispersed along the sterile lamellae edge, cylindric-clavate, septate, with rounded, mucronate, acuminate apex, rarely with papilla. Lamellar trama are regular, made up of cylindrical hyphae 4.0–9.0 µm wide, intertwined by refractive hyphae near the margin. Pileipellis is a cutis with transitions to a trichoderm towards margin, made up of cylindrical hyphae 4.0–14 μm broad, thin-walled, slightly constricted at the septa, with tapered end and sparsely pale yellow encrusting pigment, easily dissolved in KOH solution. Stipitipellis is a cutis composed of cylindrical hyphae 4.0–8.0 μm wide. Clamp connections are present in all tissue.
Habitat. Scattered or solitary on soil in mixed coniferous-broad-leaved forest.
Distribution. China
Additional specimens examined. China, Jiangxi Province, Matsu Mountain, 29°37′51″ N, 116°5′25″ E, alt. 214 m, 2 July 2019, collected by Jun-Qing Yan, HFJAU0985; Fujian Province, Wuyishan City, Kuzhukeng, 27°43′52″ N, 117°51′21″ E, alt. 591 m, 25 July 2022, collected by Jun-Qing Yan and Bin-Ron Ke, HFJAU3939; Wuyi Mountain 27°44′43″ N, 117°41′6″ E, alt. 816 m, 13 August 2021, collected by Jun-Qing Yan and Ze-Wei Liu, HFJAU4738
Notes. In the two phylogenetic trees, E. subcycneum groups together with E. cycneum and E. peristerinum, but E. cycneum shows 95.4%, 98.3%, and 98.5% similarity with E. subcycneum in ITS, LSU, and tef-1α sequence, E. peristerinum shows 86.1%, 94.6%, and 94.8% similarity, respectively. In addition, E. cycneum and E. peristerinum are characterized by the presence of abundant brilliant granules, and having differentiated type of cheilocystidia and porphyrogriseum-type in E. peristerinum as well as serrulatum-type in E. cycneum [18].
Morphologically, E. caribaeum (Pegler) Courtec. and Fiard differs in the free lamellae, larger basidiospores (12–17 × 11–15 μm), 2-spored basidia, and lack of clamp connections [43]. E. cuboidoalbum Noordel. and Hauskn. is recognized by the typical omphaloid basidiomata, estriate pileus, forked lamellae, heterogeneous lamellae edge, and abundant cheilocystidia with a variable shape from cylindrico-clavate to lageniform or fusiform [44]. E. galericolor Courtec. exhibits omphaloid basidiomata, arcuate lamellae, beige brown stipe, smaller basidiospores (7–9 μm), and trichodermal pileipellis [45].
MycoBank: MB854070
Etymology. Morphology similar to “Entoloma murrayi”.
Holotype. China, Fujian Province, Nanping City, Lingxia Creek, 27°32′31″ N, 117°28′27″ E, alt. 425 m, 7 June 2022, collected by Jun-Qing Yan and Lin-Gen Chen, HFJAU3587.
Diagnosis: Entoloma submurrayi is mainly characterized by the small and mycenoid basidiomata; conical to campanulate with acute papilla, yellow, glabrous, and striae pileus; pileus margin exceeding the lamellae; adnexed lamellae; glabrous and striae stipe; cuboid basidiospores; sterile lamellae edge; cylindric-clavate cheilocystidia; 2-layered pileipellis made up of suprapellis and thinner subpellis of oleiferous hyphae; and the presence of clamp connections. It differs from E. flavoquadratum C.K. Pradeep and K.B. Vrinda by its mycenoid basidiomata and translucently striate pileus with obvious acute papilla at the center.
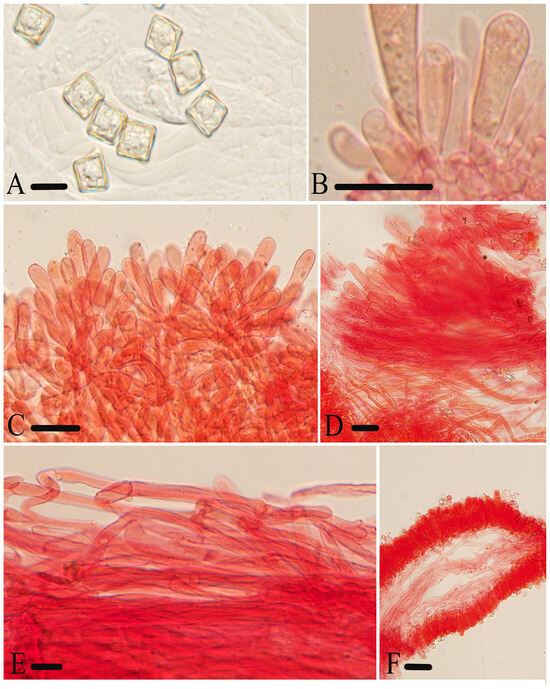
Figure 10.
Micromorphological structures of Entoloma submurrayi. (A) Basidiospores. (B) Basidia. (C) Cheilocystidia. (D) Pileipellis. (E) Stipitipellis. (E) Lamellar trama. Scale bars: (A) 10 μm; (B–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Figure 10.
Micromorphological structures of Entoloma submurrayi. (A) Basidiospores. (B) Basidia. (C) Cheilocystidia. (D) Pileipellis. (E) Stipitipellis. (E) Lamellar trama. Scale bars: (A) 10 μm; (B–F) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Description. Basidiomata are small, mycenoid. Pileus are 10–24 mm wide, campanulate, conical, or broadly conical with obvious acute papilla, translucently striate almost up to the center, smooth and glabrous, light yellow to golden yellow (3A4–5, 5AB7), lighter or fading towards rugouse margin exceeding the lamellae. Lamellae are moderately distant, 2.0–3.0 mm wide, with two types of lamellules, adnexed, sinuate, subventricose, concolored with cap, with a wavy or serrate concolorous edge. Stipe are 21–65 × 2.0–5.0 mm, central, terete, hollow, equal, white-yellow or concolorous with the pileus, with longitudinal or oblique striae, finely pruinose in the upper part elsewhere smooth and glabrous, and have a base with white tomentum. Context is thin, concolorous to the surface. Odor is indistinct, taste is not tested.
Basidiospores are (8.0)8.5–11.0 (12.0) × 8.0–10.5 (11.5) μm, (av = 9.6 × 9.2 μm), Q = 1.0–1.1 (1.2) (Qm = 1.04 ± 0.03, n = 100), isodiameterical, cuboid, thick-walled, inamyloid. Basidia are 42–60 × 11–16 μm, clavate, 4-spored, sterigmata 7.0–11 μm long, clamped. Pleurocystidia are absent. Cheilocystidia are 47–94 × 8.0–14 μm, serrulatum-type, irregular clusters in the sterile lamellae edge, cylindric-clavate, with rounded, rarely acute apex. Lamellar trama are regular, made up of cylindrical hyphae 5.0–18 µm wide. Pileipellis is a 2-layered, suprapellis cutis with transitions to a trichoderm towards the center, made up of cylindrical hyphae 10–15 μm broad, thin-walled, slightly constricted at septa, with rounded or acute end; subpellis made up of thinner cylindrically oleiferous hyphae, up to 9.0 μm wide, all with pale yellow membranal pigment. Stipitipellis is a cutis composed of cylindrical hyphae 5.0–12 μm wide, with rounded or acute end. Clamp connections are abundant in all parts of the basidiocarp.
Habitat. Scattered or solitary on soil in mixed coniferous-broad-leaved forest.
Distribution. China.
Additional specimens examined. China, Jiangxi Province, Tongmuguan Pass of Wuyi Mountain, 27°48′54″ N, 117°43′7″ E, alt. 1130 m, 7 July 2019, collected by Jun-Qing Yan, HFJAU1050; Jiujiang City, Lushan Botanical Garden, 29°32′51″ N, 115°59′0″ E, alt. 1125 m, 9 July 2019, collected by Hong-Zhao Pan, HFJAU1062; Fujian Province, Wuyishan City, 27°42′38″ N, 117°51′30″ E, alt. 521 m, 13 August 2021, collected by Qin Na, Yu-Peng Ge, and Yu-Lan Sun, HFJAU3152.
Notes. In the IL trees, E. submurrayi formed a sister lineage with E. murrayi, but the latter differs in its pileus margin not exceeding the lamellae, lack of clamp connections [38], and differences in the ITS region, with a similarity of 95%. Additionally, E. cremeoluteum (Largent) Noordel. and Co-David, E. flavoquadratum and E. pseudomurrayi Eyssart., Ducousso and Buyck are rather similar to the new species. Nevertheless, they exhibit distinct differences. E. cremeoluteum is characterized by the rostrate-ventricose cheilocystidia and pleurocystidia, and rare clamp connections [4]. E. flavoquadratum is characterized by its tricholomatoid basidiomata and shorter cheilocystidia (18.5–36 μm) [46]. E. pseudomurrayi is distinguished by the cheilocystidia of various shapes, from clavate, cylindrical, and long lanceolate to moniliform [47].
MycoBank: MB854071
Etymology. According to the features of pileus—tomentose.
Holotype. China, Fujian Province, Wuyishan City, Wuyi Mountain, 27°58′49″ N, 118°3′49″ E, alt. 1392 m, 17 August 2023, collected by Nian-Kai Zeng, Cheng-Feng Nie, Hua-Zhi Qin, Hui Deng, Tian Jiang, and Run-Xiang Zhao, HFJAU5159.
Diagnosis. Entoloma tomentosum is mainly characterized by the rather small basidiomata; conical to plano-convex with or without papilla, tomentose or velvety-scaly, white to pale yellow, and striate pileus; pileus margin exceeding the lamellae; adnate, lamellae; tomentose or glabrous stipe; cuboid basidiospores; heterogeneous lamella edge; versiform cheilocystidia with brown-yellow contents; and the presence of clamp connections. It differs from E. albidoquadratum by its smaller basidiospores, heterogeneous lamella edge, and absence of pleurocystidia.
Description. Basidiomata are rather small. Pileus are 12–25 mm wide, conical, campanulate to plano-convex with or without papilla, densely or sparsely tomentose or velvety-scaly, not translucently striate when young, becoming strikingly translucently striate up to the center on maturity, with straight or slightly inrolled margin, exceeding the lamellae, white to pale yellow (1–4A1 to 1–4A3), sometimes yellowish green at the center (30C8). Lamellae are sparse to moderately distant, 1.5–2.0 mm wide, with two types of lamellules, adnexed, ventricose, white at first, becoming pink, with wavy and concolorous edge. Stipe are 30–60 × 2.0–4.0 mm, central, terete, hollow, equal, white, glabrous or tomentose, and have a base with white tomentum. Context is thin, white. Odor is indistinct, taste is not tested.
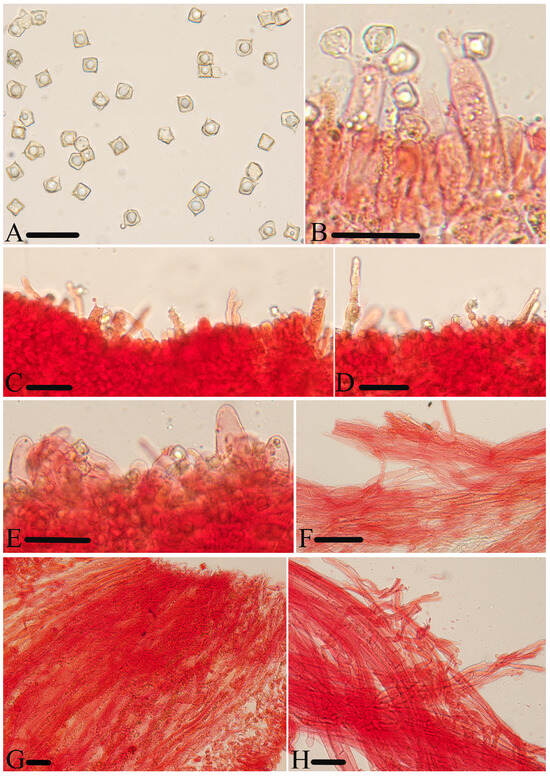
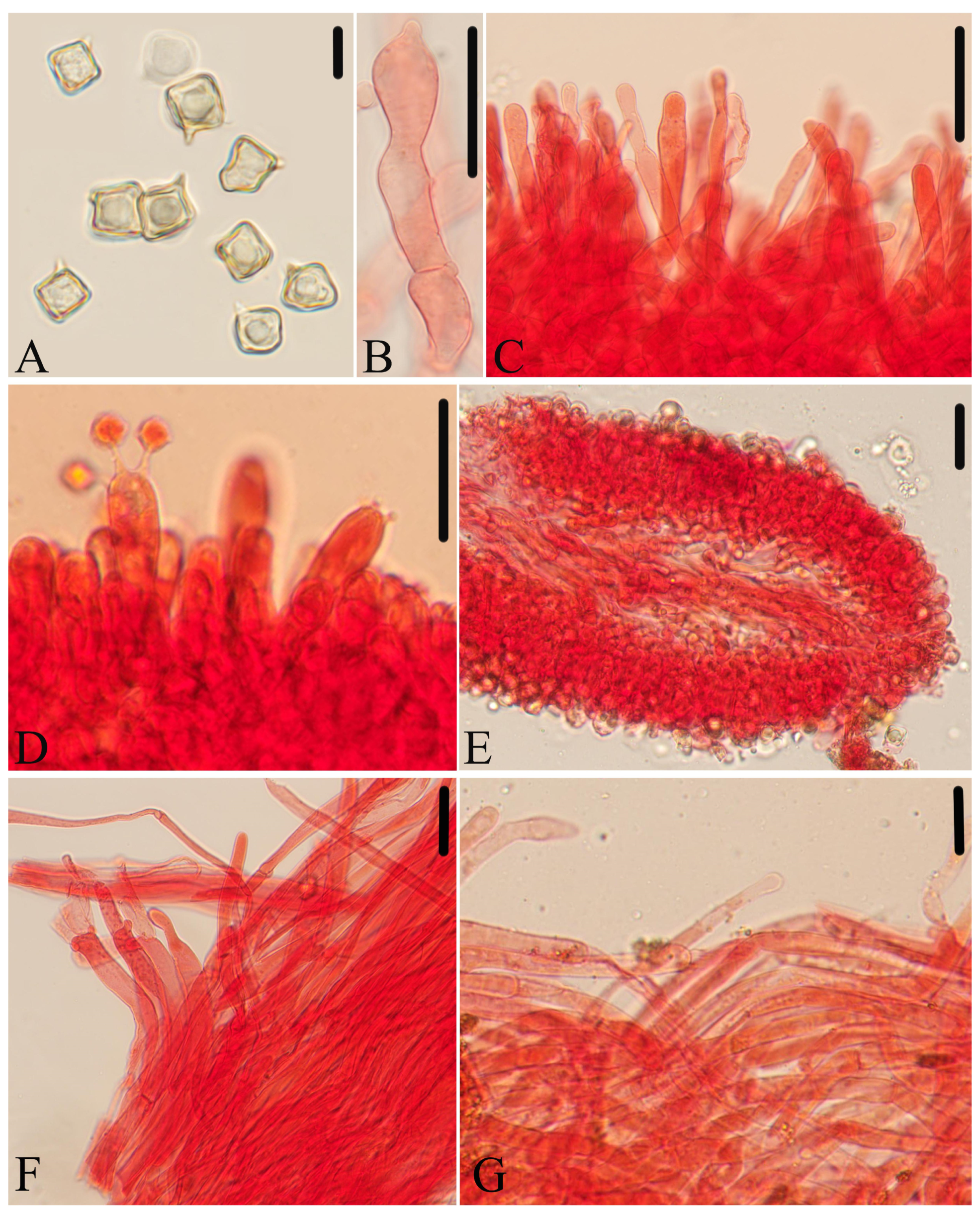
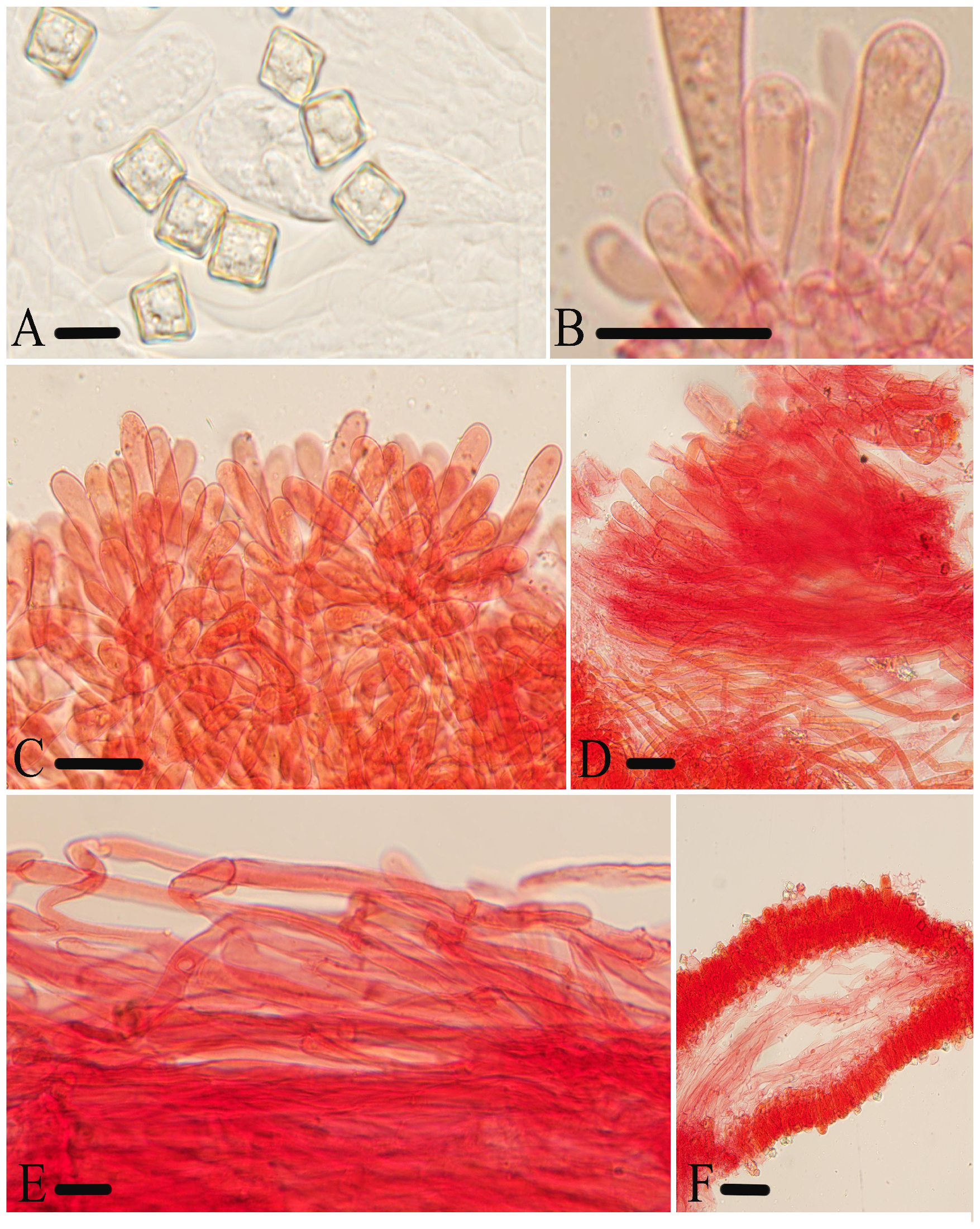
Figure 11.
Micromorphological structures of Entoloma tomentosum. (A) Basidiospores. (B) Basidia. (C,D) Cheilocystidia. (E,F) Pileipellis. (G) Lamellar trama. (H) Stipitipellis. Scale bars: (A–H) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Figure 11.
Micromorphological structures of Entoloma tomentosum. (A) Basidiospores. (B) Basidia. (C,D) Cheilocystidia. (E,F) Pileipellis. (G) Lamellar trama. (H) Stipitipellis. Scale bars: (A–H) 30 μm. All structures were observed in 5% KOH, and 1% Congo red was used as the stain, except (A).
Basidiospores are (7.5) 8.0–10.0 (11.0) × (7.0) 8.0–9.5 (10.5) μm, (av = 9.1 × 8.6 μm), Q = 1.0–1.1 (1.2) (Qm = 1.06 ± 0.04, n = 100), isodiameterical, cuboid, sporadically with five angles in side-view, thick-walled, inamyloid. Basidia are 41–53 × 10–14 μm, clavate, 4- or 2-spored, sterigmata 7.0–13 μm long, clamped. Pleurocystidia are absent. Cheilocystidia are 35–113 × 6.0–11 μm, regularly dispersed along the heterogeneous edge, versiform, cylindrical, long lanceolate, or moniliform, with rounded or acute apex and brown-yellow contents. Lamellar trama are regular, made up of cylindrical hyphae 5.0–13 µm wide, intertwined by oleiferous hyphae near the margin. Pileipellis is a cutis with transitions to a trichoderm, made up of cylindrical hyphae 8.0–17 μm broad, thin-walled, somewhat constricted at septa, with acute end and pale yellow epiparietal pigment. Stipitipellis is a cutis composed of cylindrical hyphae 5.0–9.0 μm wide, even at the septa, with a rounded end. Brilliant granules are abundant and clamp connections are present in all tissue.
Habitat. Scattered on soil or rotten wood in mixed coniferous-broad-leaved forest.
Distribution. China.
Additional specimens examined. China, Fujian Province, Wuyishan City, Wuyi Mountain, 27°58′49″ N, 118°3′49″ E, alt. 1392 m, 16 August 2023, collected by Nian-Kai Zeng, Cheng-Feng Nie, Hua-Zhi Qin, Hui Deng, Tian Jiang, and Run-Xiang Zhao, HFJAU5116; 17 August 2023, HFJAU5153, HFJAU5160, HFJAU5166.
Notes. In the two phylogenetic trees, E. pallidoflavum is the closest species to the new species, but differs in the cylindrical and hyaline cheilocystidia [38], and shows only 98.4%, 96.8%, and 98.5% similarity with new species in ITS, tef-1α, and rpb2 sequences. Morphologically, the new species has much in common with E. albidoquadratum from India, regarding the white-to-pale-yellow pileus and the versiform cheilocystidia. However, E. albidoquadratum is distinguished by its fimbriate and sterile lamella edge, larger basidiospores (11–16.5 × 9–14 μm), hyaline cheilocystidia, and the presence of pleurocystidia that are similar in appearance to cheilocystidia [39].
4. Discussion
According to the previous studies of Karstedt et al., the subgenus Cubospora formed a monophyly placed in the/Inocephalus-Cyanula, with the synapomorphies of mycenoid, collybioid, or tricholomatoid habit and cuboid with dihedral base basidiospores [9].
In China, about 20 species of Entoloma subgenus Cubospora were distributed in southern regions, with the white species typically identified as E. album, the orange species as E. quadratum, and the yellow species as E. murrayi. The new species of this study exhibit significant differences from the three aforementioned species. Additionally, some sequences that have been identified as E. quadratum and E. murrayi are noteworthy for not clustering on a single branch but forming many independent branches. This suggests that the current understanding of the distribution of these two species may not be as widespread as previously reported, and there may be some new species that are morphologically very similar to these two species which have not yet been reported.
5. Conclusions
Seven new species described from subtropical regions of China were well supported based on phylogenetic analysis and morphological characteristics. This study further confirmed a taxonomic relationship among the species of subgenus Cubospora and enhanced the species diversity of subtropical regions of China.
6. Key to Related Species
1. Pileus white or pale beige 2
1’ Pileus other colored 11
2. Lamellae edge heterogeneous 3
2’ Lamellae edge sterile or fertile 4
3. Pileus depressed, estriate; lamellae decurrent; basidiospores 9.0–11.5 × 7.5–10.5 μm
E. cuboidoalbum
3’ Pileus epapillose or papillose, striate; lamellae adnexed; basidiospores 8.0–10 × 8.0–9.5 μm
E. tomentosum
4. Clampless 5
4’ Clamped 6
5. Pileus campanulate to hemispherical with acute papilla; lamellae adnexed to adnate with serrate edge; basidiospores 7.0–9.5 μm
E. album
5’ Pileus applanate with depression; lamellae adnate to decurent with fimbriate edge; basidiospores 5.0–7.0 μm
E. overeemii
6. Basidiospores 11–16.5 × 9.0–14 μm; cheilocystidia versiform; pleurocystidia present
E. albidoquadratum
6’ Basidiospores smaller; cheilocystidia not versiform; pleurocystidia absent
7
7. Pileus obviously papillate 8
7’ Pileus epapillate or unobviously papillate 9
8. Pileus not hygrophanous; lamellae adnexed; cheilocystidia carneogriseum-type; brilliant granules absent E. lacticolor
8’ Pileus hygrophanous; lamellae adnate-emarginate; cheilocystidia porphyrogriseum-type; brilliant granules abundant E. peristerinum
9. Cheilocystidia carneogriseum-type; pileus glabrous or erected-velutinous; lamellae adnexed E. subcycneum
9’ Cheilocystidia other type 10
10. Pileus 10–25 mm, smooth and glabrous; caulocystidia present
E. cycneum
10’ Pileus 30–100 mm, fibrillose; caulocystidia absent
E. pallidoflavum
11. Pileus pink to brown 12
11’ Pileus orange to yellow 13
12. Lamellae with tiny lateral veins; pileus striate
E. phlebophyllum
12’ Lamellae without tiny lateral veins; pileus striate
E. carneum
13. Pileus orange, conical or campanulate with or without distinct acute papilla, glabrous to fibrillose adnexed to almost free with fimbriate edge; brilliant granules abundant E. quadratum
13’ Pileus yellow to brown yellow 14
14. Neither cystidia nor clamp connections E. phleboides
14’ Either cystidia or clamp connections or both 15
15. Cystidia absent; basidiospores 6.0–9.0 μm E. gracilius
15’ Cystidia present; basidiospores larger 16
16. Clampless; pileus innately fibrillose; lamellae adnexed to adnate
E. murrayi
16’ Clamped 17
17. Lamellae edge red-brown underlined
E. rufomarginatum
17’ Lamellae edge not underlined 18
18. Pileus with depressed centre, mycenoid, glabrous, estriate; lamellae sparse, adnexed; stipe smooth and glabrous E. excavatum
18’ Pileus without depressed centre 19
19. Pleurocystidia present, rostrate-ventricose; pileus broadly campanulate, hygrophanous; clamp connections presence, but rare
E. cremeoluteum
19’ Pleurocystidia absent 20
20. Cheilocystidia versiform, empty or filled with an emulsified or crystallized yellow pigment E. pseudomurrayi
20’ Cheilocystidia not versiform 21
21. Pileus without papilla, convex to hemispherical, glabrous to fibrillose, striate; lamellae adnexed to almostfree with fimbriate edge E. luteum
21’ Pileus with papilla 22
22. Pileus margin exceeding the lamellae 23
22’ Pileus margin not exceeding the lamellae 24
23. Pileus tricholomatoid, estriate; caulocystidia present
E. flavoquadratum
23’ Pileus mycenoid, striate; caulocystidia absent
E. submurrayi
24. Lamellae free; stipe fibrillose; on rotten wood
E. avilanum
24’ Lamellae adnexed; stipe glabrous; on soil E. plicatum
Author Contributions
Conceptualization, J.-Q.Y.; Data curation, J.-Q.Y.; Formal analysis, L.-G.C.; Funding acquisition, J.-Q.Y.; Investigation, L.D., H.C., and Z.-H.Z.; Methodology, S.-N.W.; Project administration, J.-Q.Y.; Resources, H.Z. and Z.-H.Z.; Software, L.D. and H.C.; Supervision, H.Z.; Visualization, L.-G.C.; Writing—original draft, L.-G.C.; Writing—review and editing, S.-N.W. and J.-Q.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financed by the National Natural Science Foundation of China (31960008), Jiangxi Provincial Natural Science Foundation (20224BAB205003), Fujian Provincial Natural Science Foundation (2023J01379), and the Project of FAAS (XTCXGC2021007).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All alignments for phylogenetic analyses were deposited in TreeBASE (http://www.treebase.org); the following links were available: http://purl.org/phylo/treebase/phylows/study/TB2:S31496?x-access-code=a3f46a59482fe0426a9347d545e5ba6e&format=html (accessed on 18 August 2024)).
Acknowledgments
The authors are very grateful for assistance of Yu-Peng Ge and Meng-Hui Han in the field specimen collection and the anonymous reviewers of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kummer, P. Der Führer in die Pilzkunde: Anleitung zum Methodischen, Leichten und Sichern Bestimmen der in Deutschland Vorkommenden Pilze: Mit Ausnahme der Schimmel- und Allzu Winzigen Schleim- und Kern-Pilzchen; Verlag von E. Luppe’s Buchhandlung: Zerbst, Germany, 1871; p. 146. [Google Scholar]
- Co-David, D.; Langeveld, D.; Noordeloos, M.E. Molecular phylogeny and spore evolution of Entolomataceae. Persoonia 2009, 23, 147–176. [Google Scholar] [CrossRef] [PubMed]
- Noordeloos, M.E. Introduction to the taxonomy of the genus Entoloma sensu lato (Agaricales). Persoonia 1981, 11, 121–151. [Google Scholar]
- Largent, D.L. Entolomatoid Fungi of the Western United States and Alaska; Mad River Press: Eureka, CA, USA, 1994; p. 495. [Google Scholar]
- He, X.L.; Horak, E.; Wang, D.; Li, T.H.; Peng, W.H.; Gan, B.C. Descriptions of five new species in Entoloma subgenus Claudopus from China, with molecular phylogeny of Entoloma s.l. MycoKeys 2019, 61, 1–26. [Google Scholar] [CrossRef]
- Asif, M.; Saba, M.; Akram, W.; Maula, F.; Raza, M. Integrated morphological and molecular approaches unveil a new species and a new record of the genus Entoloma (Entolomataceae) from southern Punjab, Pakistan. Phytotaxa 2024, 653, 219–240. [Google Scholar] [CrossRef]
- He, X.L.; Li, T.H.; Jiang, Z.D.; Shen, Y.H. Two new cuboid-spored species of Entoloma s.l. (Agaricales, Entolomataceae) from southern China. Cryptogam. Mycol. 2015, 36, 237–249. [Google Scholar] [CrossRef]
- Noordeloos, M.E. Entoloma s.l. Fungi Europaei 5; Libreria Editrice Giovanna Biella: Saronno, Italy, 1992; p. 760. [Google Scholar]
- Karstedt, F.; Capelari, M.; Baroni, T.J.; Largent, D.L.; Bergemann, S.E. Phylogenetic and morphological analyses of species of the Entolomataceae (Agaricales, Basidiomycota) with cuboid basidiospores. Phytotaxa 2019, 391, 1–27. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. The Methuen Handbook of Colour; Eyre Methuen Ltd.: London, UK, 1978; p. 252. [Google Scholar]
- Horak, E. Röhrlinge und Blätterpilze in Europa: Bestimmungsschlüssel für Polyporales (pp), Boletales, Agaricales, Russulales; Moser, M., Ed.; Elsevier, Spektrum, Akad. Verl.: Munich, Germany, 2005; p. 557. [Google Scholar]
- Bas, C. Morphology and subdivision of Amanita and a monograph of its section Lepidella. Persoonia 1969, 5, 285–573. [Google Scholar]
- Noordeloos, M.E.; Dima, B.; Morozova, O.V.; Reschke, K.; Jansen, G.M.; Brandrud, T.E.; Jordal, J.B.; Bendiksen, E.; Vila, J. Entoloma Sensu Lato. Subgenera Cyanula, Leptonia, Nolanea, Trichopilus, and the/Rhombisporum Clade; Candusso Edtrice: Origgio, Italy, 2022; p. 968. [Google Scholar]
- Wang, S.N.; Fan, Y.G.; Yan, J.Q. Iugisporipsathyra reticulopilea gen. et sp. nov. (Agaricales, Psathyrellaceae) from tropical China produces unique ridge-ornamented spores with an obvious suprahilar plage. MycoKeys 2022, 90, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Hopple, J.S.; Vilgalys, R. Phylogenetic Relationships in the Mushroom Genus Coprinus and Dark-Spored Allies Based on Sequence Data from the Nuclear Gene Coding for the Large Ribosomal Subunit RNA: Divergent Domains, Outgroups, and Monophyly. Mol. Phylogenet. Evol. 1999, 13, 1–19. [Google Scholar] [CrossRef]
- Kluting, K.; Baroni, T.J.; Bergemann, S.E. Toward a stable classification of genera within the Entolomataceae: A phylogenetic re-evaluation of the Rhodocybe-Clitopilus clade. Mycologia 2014, 106, 1127–1142. [Google Scholar] [CrossRef]
- Bau, T.; Yan, J.Q. A new genus and four new species in the /Psathyrella s.l. clade from China. MycoKeys 2021, 80, 115–131. [Google Scholar] [CrossRef]
- Morozova, O.V.; Pham, T.H.G. New species of Entoloma subgenera Cubospora and Leptonia (Agaricales, Basidiomycota) from central Vietnam. J. Fungi 2023, 9, 621. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Briefings Bioinf. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Haeseler, A.V.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Haeseler, A.V.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Dettman, J.R.; Jacobson, D.J.; Taylor, J.W. A multilocus genealogical approach to phylogenetic species recognition in the model eukaryote Neurospora. Evolution 2003, 57, 2667–2919. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Popov, E.; Alexandrova, A. Three new species within the genus Entoloma (Basidiomycota, Agaricales) with clamped basidia and a serrulatum-type lamellae edge, and their phylogenetic position. Mycol. Prog. 2017, 17, 381–392. [Google Scholar] [CrossRef]
- Karstedt, F.; Bergemann, S.E.; Capelari, M. Five Nolanea spp. nov. from Brazil. Mycotaxon 2020, 135, 589–612. [Google Scholar] [CrossRef]
- Reschke, K.; Noordeloos, M.E.; Manz, C.; Hofmann, T.A.; Rodríguez-Cedeño, J.; Dima, B.; Piepenbring, M. Fungal diversity in the tropics: Entoloma spp. in Panama. Mycol. Prog. 2022, 21, 93–145. [Google Scholar] [CrossRef]
- Reschke, K.; Morozova, O.V.; Dima, B.; Cooper, J.A.; Corriol, G.; Biketova, A.Y.; Piepenbring, M.; Noordeloos, M.E. Phylogeny, taxonomy, and character evolution in Entoloma subgenus Nolanea. Persoonia 2022, 49, 136–170. [Google Scholar] [CrossRef]
- He, X.L.; Li, T.H.; Jiang, Z.D.; Shen, Y.H. Four new species of Entoloma s.l. (Agaricales) from southern China. Mycol. Prog. 2012, 11, 915–925. [Google Scholar] [CrossRef]
- Crous, P.W.; Osieck, E.R.; Jurjević, Ž.; Boers, J.; Iperen, A.V.; Starink, M.; Dima, B.; Balashov, S.; Bulgakov, T.; Johnston, P.R.; et al. Fungal Planet description sheets: 1284–1382. Persoonia 2021, 47, 1284–1382. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Zhang, P. Atlas of Macrofung in Hunan; Hunan Normal University Press: Changsha, China, 2019; p. 426. [Google Scholar]
- He, X.L.; Li, T.H.; Xi, P.G.; Jiang, Z.D.; Shen, Y.H. Phylogeny of Entoloma s.l. subgenus Pouzarella with descriptions of five new species from China. Fungal Divers. 2013, 58, 227–243. [Google Scholar] [CrossRef]
- He, X.L.; Peng, W.H.; Gan, B.C. Morphological and molecular evidence for a new species in Entoloma subgenus Claudopus from Sichuan Province, southwest China. Mycoscience 2015, 56, 326–331. [Google Scholar] [CrossRef]
- Baroni, T.J.; Hofstetter, V.; Largent, D.L.; Vilgalys, R. Entocybe is proposed as a new genus in the Entolomataceae (Agaricomycetes, Basidiomycota) based on morphological and molecular evidence. N. Am. Fungi 2011, 6, 1–19. [Google Scholar] [CrossRef]
- Largent, D.L.; Henkel, T.W.; Siegel, N.; Koch, R.; Séné, O.; Hageman, K.; Aime, M.C. New species of Entolomataceae from Cameroon. Fungal Syst Evol. 2019, 5, 151–168. [Google Scholar] [CrossRef]
- Largent, D.L.; Bergemann, S.E.; Abell-Davis, S.E.; Kluting, K.; Cummings, G.A. Three new Inocephalus species with cuboid basidiospores from New South Wales and Queensland, Australia. Mycotaxon 2013, 123, 301–319. [Google Scholar] [CrossRef]
- Morozova, O.V.; Noordeloos, M.E.; Vila, J. Entoloma subgenus Leptonia in boreal-temperate Eurasia: Towards a phylogenetic species concept. Persoonia 2014, 32, 141–169. [Google Scholar] [CrossRef]
- Largent, D.L.; Kluting, K.; Anderson, N.M.; Bergemann, S. New leptonioid species from New South Wales and northeastern Queensland, Australia. Mycotaxon 2016, 131, 153–176. [Google Scholar] [CrossRef]
- Moncalvo, J.-M.; Vilgalys, R.; Redhead, S.; Johnson, J.E.; James, T.Y.; Aime, M.C.; Valerie, H.; Verduin, S.J.W.; Larsson, E.; Baroni, T.J.; et al. One Hundred and Seventeen Clades of Eu-agarics. Mol. Phylogenet. Evol. 2002, 23, 357–400. [Google Scholar] [CrossRef] [PubMed]
- Horak, E. On cuhoid-spored species of Entoloma (Agaricales). Sydowia 1976, 28, 171–236. [Google Scholar]
- Manimohan, P.; Noordeloos, M.E.; Dhanya, A.M. Studies on the genus Entoloma (Basidiomycetes, Agaricales) in Kerala State, India. Persoonia 2006, 19, 45–93. [Google Scholar]
- Morozova, O.V.; Popov, E.; Kovalenko, A. Studies on Mycobiota of Vietnam. I. Genus Entoloma: New Records and New Species. Mikol. I Fitopatol. 2012, 46, 184–200. [Google Scholar]
- Bi, Z.S.; Zheng, G.Y.; Li, T.H. Taxonomic studies on the genus Entoloma from Guangdong. Acta Mycol. Sin. 1986, 5, 161–169. [Google Scholar] [CrossRef]
- Petch, T. Additions to Ceylon Fungi. Ann. R. Bot. Gard. Perad. 1917, 6, 195–256. [Google Scholar]
- Pegler, D.N. Agaric Flora of the Lesser Antilles; Her Majesty’s Stationery Office: London, UK, 1983; p. 668. [Google Scholar]
- Noordeloos, M.E.; Hausknecht, A. New and interesting Entoloma species from Central Europe. Österr. Z. Für Pilzkd. 2009, 18, 169–182. [Google Scholar]
- Courtecuisse, R. Macromycètes intéressantes, rares ou nouveaux - 6 - Entolomataceae. Doc. Mycol. 1993, 23, 1–38. [Google Scholar]
- Pradeep, C.K.; Vrinda, K.B.; Chandrasekharan, B.; Baroni, T.J. Additions to the quadrate-spored Entoloma (Agaricales) in Kerala State, India. Mycosphere 2016, 7, 642–648. [Google Scholar] [CrossRef]
- Eyssartier, G.; Ducousso, M.; Buyck, B. Entoloma pseudomurrayi sp. nov., un nouvel entolome jaune des forêts néo-calédoniennes de montagne à Nothofagus. Cryptogam. Mycol. 2010, 31, 3–8. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).