Let’s Get Physical: Bacterial-Fungal Interactions and Their Consequences in Agriculture and Health



Abstract
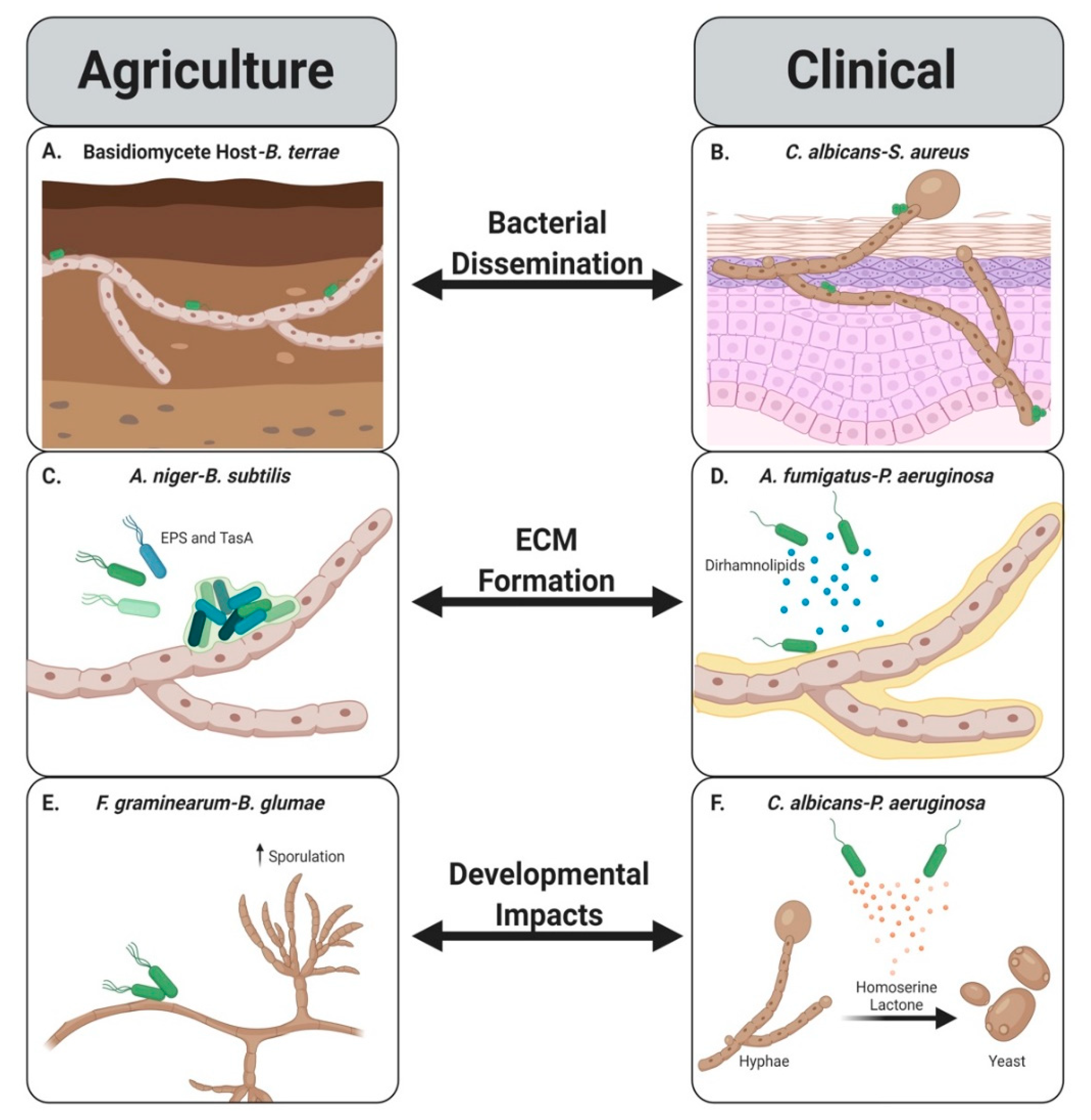
:1. Introduction
2. Bacterial-Fungal Physical Interactions
2.1. Bacterial Attachment to the External Surface of Fungi
2.1.1. Human Pathogenic Fungi and Their Bacterial Interactions
2.1.2. Hyphal Attachment in Environmental BFIs
2.2. Bacterial Invasion into Fungi
2.2.1. Specialized Endofungal Symbioses
BFIs in Mucoromycotina
BFIs in Glomeromycotina
BFIs in Mortierellomycotina
2.2.2. Facultative Endofungal Associations
3. Implications of BFIs on Microbial Fitness
3.1. Microbial Fitness Associated with Growth and Development of Microbial Partners
3.2. Factors Obtained Via BFIs
3.3. Fungal Highways for Bacterial Dissemination
4. Implications of BFIs on Host Health and Disease
4.1. Humans
4.2. Plants
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bianciotto, V.; Bandi, C.; Minerdi, D.; Sironi, M.; Tichy, H.V.; Bonfante, P. An obligately endosymbiotic mycorrhizal fungus itself harbors obligately intracellular bacteria. Appl. Environ. Microbiol. 1996, 62, 3005–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estrada-de Los Santos, P.; Palmer, M.; Chávez-Ramírez, B.; Beukes, C.; Steenkamp, E.T.; Briscoe, L.; Khan, N.; Maluk, M.; Lafos, M.; Humm, E.; et al. Whole genome analyses suggests that Burkholderia sensu lato contains two additional novel genera (Mycetohabitans gen. nov., and Trinickia gen. nov.): Implications for the evolution of diazotrophy and nodulation in the Burkholderiaceae. Genes 2018, 9, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partida-Martinez, L.P.; Hertweck, C. Pathogenic fungus harbours endosymbiotic bacteria for toxin production. Nature 2005, 437, 884–888. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Ovchinnikova, E.S.; Krom, B.P.; Harapanahalli, A.K.; Busscher, H.J.; van der Mei, H.C. Surface thermodynamic and adhesion force evaluation of the role of chitin-binding protein in the physical interaction between Pseudomonas aeruginosa and Candida albicans. Langmuir ACS J. Surf. Colloids 2013, 29, 4823–4829. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, E.S.; Krom, B.P.; van der Mei, H.C.; Busscher, H.J. Force microscopic and thermodynamic analysis of the adhesion between Pseudomonas aeruginosa and Candida albicans. Soft Matter 2012, 8, 6454–6461. [Google Scholar] [CrossRef]
- Ovchinnikova, E.S.; Krom, B.P.; Busscher, H.J.; van der Mei, H.C. Evaluation of adhesion forces of Staphylococcus aureus along the length of Candida albicans hyphae. BMC Microbiol. 2012, 12, 281. [Google Scholar] [CrossRef] [Green Version]
- Beaussart, A.; Herman, P.; El-Kirat-Chatel, S.; Lipke, P.N.; Kucharíková, S.; Van Dijck, P.; Dufrêne, Y.F. Single-cell force spectroscopy of the medically important Staphylococcus epidermidis-Candida albicans interaction. Nanoscale 2013, 5, 10894–10900. [Google Scholar] [CrossRef] [Green Version]
- Silverman, R.J.; Nobbs, A.H.; Vickerman, M.M.; Barbour, M.E.; Jenkinson, H.F. Interaction of Candida albicans cell wall Als3 protein with Streptococcus gordonii SspB adhesin promotes development of mixed-species communities. Infect. Immun. 2010, 78, 4644. [Google Scholar] [CrossRef] [Green Version]
- Peters, B.M.; Ovchinnikova, E.S.; Krom, B.P.; Schlecht, L.M.; Zhou, H.; Hoyer, L.L.; Busscher, H.J.; van der Mei, H.C.; Jabra-Rizk, M.A.; Shirtliff, M.E. Staphylococcus aureus adherence to Candida albicans hyphae is mediated by the hyphal adhesin Als3p. Microbiology (Read. Engl.) 2012, 158, 2975–2986. [Google Scholar] [CrossRef] [Green Version]
- Schlecht, L.M.; Peters, B.M.; Krom, B.P.; Freiberg, J.A.; Hänsch, G.M.; Filler, S.G.; Jabra-Rizk, M.A.; Shirtliff, M.E. Systemic Staphylococcus aureus infection mediated by Candida albicans hyphal invasion of mucosal tissue. Microbiology (Read. Engl.) 2015, 161, 168–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoyer, L.L.; Payne, T.L.; Bell, M.; Myers, A.M.; Scherer, S. Candida albicans ALS3 and insights into the nature of the ALS gene family. Curr. Genet. 1998, 33, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Reece, E.; Segurado, R.; Jackson, A.; McClean, S.; Renwick, J.; Greally, P. Co-colonisation with Aspergillus fumigatus and Pseudomonas aeruginosa is associated with poorer health in cystic fibrosis patients: An Irish registry analysis. BMC Pulm. Med. 2017, 17, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briard, B.; Rasoldier, V.; Bomme, P.; ElAouad, N.; Guerreiro, C.; Chassagne, P.; Muszkieta, L.; Latgé, J.-P.; Mulard, L.; Beauvais, A. Dirhamnolipids secreted from Pseudomonas aeruginosa modify anjpegungal susceptibility of Aspergillus fumigatus by inhibiting β1,3 glucan synthase activity. ISME J. 2017, 11, 1578–1591. [Google Scholar] [CrossRef]
- Harriott, M.M.; Noverr, M.C. Candida albicans and Staphylococcus aureus form polymicrobial biofilms: Effects on antimicrobial resistance. Antimicrob. Agents Chemother. 2009, 53, 3914–3922. [Google Scholar] [CrossRef] [Green Version]
- Toljander, J.F.; Artursson, V.; Paul, L.R.; Jansson, J.K.; Finlay, R.D. Attachment of different soil bacteria to arbuscular mycorrhizal fungal extraradical hyphae is determined by hyphal vitality and fungal species. FEMS Microbiol. Lett. 2006, 254, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Hover, T.; Maya, T.; Ron, S.; Sandovsky, H.; Shadkchan, Y.; Kijner, N.; Mitiagin, Y.; Fichtman, B.; Harel, A.; Shanks, R.M.; et al. Mechanisms of bacterial (Serratia marcescens) attachment to, migration along, and killing of fungal hyphae. Appl. Environ. Microbiol. 2016, 82, 2585–2594. [Google Scholar] [CrossRef] [Green Version]
- Kjeldgaard, B.; Listian, S.A.; Ramaswamhi, V.; Richter, A.; Kiesewalter, H.T.; Kovács, Á.T. Fungal hyphae colonization by Bacillus subtilis relies on biofilm matrix components. Biofilm 2019, 1, 100007. [Google Scholar] [CrossRef]
- Mosse, B. Honey-coloured, sessile endogone spores-III. Wall structure. Arch. Microbiol. 1970, 74, 146–159. [Google Scholar] [CrossRef]
- Spatafora, J.W.; Chang, Y.; Benny, G.L.; Lazarus, K.; Smith, M.E.; Berbee, M.L.; Bonito, G.; Corradi, N.; Grigoriev, I.; Gryganskyi, A.; et al. A phylum-level phylogenetic classification of zygomycete fungi based on genome-scale data. Mycologia 2016, 108, 1028–1046. [Google Scholar] [CrossRef] [Green Version]
- Bonfante, P.; Venice, F. Mucoromycota: Going to the roots of plant-interacting fungi. Fungal Biol. Rev. 2020, 34, 100–113. [Google Scholar] [CrossRef]
- Partida-Martinez, L.P.; Groth, I.; Schmitt, I.; Richter, W.; Roth, M.; Hertweck, C. Burkholderia rhizoxinica sp. nov. and Burkholderia endofungorum sp. nov., bacterial endosymbionts of the plant-pathogenic fungus Rhizopus microsporus. Int. J. Syst. Evol. Microbiol. 2007, 57, 2583–2590. [Google Scholar] [CrossRef] [PubMed]
- Moebius, N.; Üzüm, Z.; Dijksterhuis, J.; Lackner, G.; Hertweck, C. Active invasion of bacteria into living fungal cells. eLife 2014, 3, e03007. [Google Scholar] [CrossRef] [Green Version]
- Lackner, G.; Moebius, N.; Partida-Martinez, L.P.; Boland, S.; Hertweck, C. Evolution of an endofungal lifestyle: Deductions from the Burkholderia rhizoxinica genome. BMC Genom. 2011, 12, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehs, S.P.; Scherlach, K.; Hertweck, C. Genomics-driven discovery of a linear lipopeptide promoting host colonization by endofungal bacteria. Org. Biomol. Chem. 2018, 16, 8345–8352. [Google Scholar] [CrossRef]
- Lackner, G.; Moebius, N.; Hertweck, C. Endofungal bacterium controls its host by an hrp type III secretion system. ISME J. 2011, 5, 252–261. [Google Scholar] [CrossRef] [Green Version]
- Richter, I.; Uzum, Z.; Stanley, C.E.; Moebius, N.; Stinear, T.P.; Pidot, S.J.; Ferling, I.; Hillmann, F.; Hertweck, C. Secreted TAL effectors protect symbiotic bacteria from entrapment within fungal hyphae. bioRxiv 2020. [Google Scholar] [CrossRef]
- Carter, M.E.; Carpenter, S.C.D.; Dubrow, Z.E.; Sabol, M.R.; Rinaldi, F.C.; Lastovestsky, O.A.; Mondo, S.J.; Pawlowska, T.E.; Bogdanove, A.J. A TAL effector-like protein of symbiotic Mycetohabitans increases stress tolerance and alters the transcriptome of the fungal host Rhizopus microsporus. bioRxiv 2020. [Google Scholar] [CrossRef]
- Desirò, A.; Salvioli, A.; Ngonkeu, E.L.; Mondo, S.J.; Epis, S.; Faccio, A.; Kaech, A.; Pawlowska, T.E.; Bonfante, P. Detection of a novel intracellular microbiome hosted in arbuscular mycorrhizal fungi. ISME J. 2014, 8, 257–270. [Google Scholar] [CrossRef] [Green Version]
- Naumann, M.; Schüßler, A.; Bonfante, P. The obligate endobacteria of arbuscular mycorrhizal fungi are ancient heritable components related to the Mollicutes. ISME J. 2010, 4, 862–871. [Google Scholar] [CrossRef] [Green Version]
- Bianciotto, V.; Genre, A.; Jargeat, P.; Lumini, E.; Bécard, G.; Bonfante, P. Vertical transmission of endobacteria in the arbuscular mycorrhizal fungus Gigaspora margarita through generation of vegetative spores. Appl. Environ. Microbiol. 2004, 70, 3600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghignone, S.; Salvioli, A.; Anca, I.; Lumini, E.; Ortu, G.; Petiti, L.; Cruveiller, S.; Bianciotto, V.; Piffanelli, P.; Lanfranco, L.; et al. The genome of the obligate endobacterium of an AM fungus reveals an interphylum network of nutritional interactions. ISME J. 2012, 6, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lozano, J.M.; Bonfante, P. A Burkholderia strain living inside the arbuscular mycorrhizal fungus Gigaspora margarita possesses the vacB Gene, which is involved in host cell colonization by bacteria. Microb. Ecol. 2000, 39, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, S.; Sato, Y.; Fujimura, R.; Takashima, Y.; Hamada, M.; Nishizawa, T.; Narisawa, K.; Ohta, H. Mycoavidus cysteinexigens gen. nov., sp. nov., an endohyphal bacterium isolated from a soil isolate of the fungus Mortierella elongata. Int. J. Syst. Evol. Microbiol. 2016, 66, 2052–2057. [Google Scholar] [CrossRef]
- Li, Z.; Yao, Q.; Dearth, S.P.; Entler, M.R.; Castro Gonzalez, H.F.; Uehling, J.K.; Vilgalys, R.J.; Hurst, G.B.; Campagna, S.R.; Labbé, J.L.; et al. Integrated proteomics and metabolomics suggests symbiotic metabolism and multimodal regulation in a fungal-endobacterial system. Environ. Microbiol. 2017, 19, 1041–1053. [Google Scholar] [CrossRef]
- Sharmin, D.; Guo, Y.; Nishizawa, T.; Ohshima, S.; Sato, Y.; Takashima, Y.; Narisawa, K.; Ohta, H. Comparative genomic insights into endofungal lifestyles of two bacterial endosymbionts, Mycoavidus cysteinexigens and Burkholderia rhizoxinica. Microbes Environ. 2018, 33, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Glaeser, S.P.; Alabid, I.; Imani, J.; Haghighi, H.; Kämpfer, P.; Kogel, K.H. The abundance of endofungal bacterium Rhizobium radiobacter (syn. Agrobacterium tumefaciens) increases in its fungal host Piriformospora indica during the Tripartite Sebacinalean Symbiosis with higher plants. Front. Microbiol. 2017, 8, 629. [Google Scholar] [CrossRef]
- Baltrus, D.A.; Dougherty, K.; Arendt, K.R.; Huntemann, M.; Clum, A.; Pillay, M.; Palaniappan, K.; Varghese, N.; Mikhailova, N.; Stamatis, D.; et al. Absence of genome reduction in diverse, facultative endohyphal bacteria. Microb. Genom. 2017, 3, e000101. [Google Scholar] [CrossRef]
- Hoffman, M.T.; Arnold, A.E. Diverse bacteria inhabit living hyphae of phylogenetically diverse fungal endophytes. Appl. Environ. Microbiol. 2010, 76, 4063–4075. [Google Scholar] [CrossRef] [Green Version]
- Spraker, J.E.; Sanchez, L.M.; Lowe, T.M.; Dorrestein, P.C.; Keller, N.P. Ralstonia solanacearum lipopeptide induces chlamydospore development in fungi and facilitates bacterial entry into fungal tissues. ISME J. 2016, 10, 2317–2330. [Google Scholar] [CrossRef] [Green Version]
- Hazarika, D.J.; Gautom, T.; Parveen, A.; Goswami, G.; Barooah, M.; Modi, M.K.; Boro, R.C. Mechanism of interaction of an endofungal bacterium Serratia marcescens D1 with its host and non-host fungi. PLoS ONE 2020, 15, e0224051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duque, A.; Samad, A.; Nybroe, O.; Antonielli, L.; Sessitsch, A.; Compant, S. Interaction between endophytic Proteobacteria strains and Serendipita indica enhances biocontrol activity against fungal pathogens. Plant Soil 2020. [Google Scholar] [CrossRef]
- Pent, M.; Bahram, M.; Põldmaa, K. Fruitbody chemistry underlies the structure of endofungal bacterial communities across fungal guilds and phylogenetic groups. ISME J. 2020, 14, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Partida-Martínez, L.P. The fungal holobiont: Evidence from early diverging fungi. Environ. Microbiol. 2017, 19, 2919–2923. [Google Scholar] [CrossRef]
- Briard, B.; Bomme, P.; Lechner, B.E.; Mislin, G.L.A.; Lair, V.; Prévost, M.-C.; Latgé, J.-P.; Haas, H.; Beauvais, A. Pseudomonas aeruginosa manipulates redox and iron homeostasis of its microbiota partner Aspergillus fumigatus via phenazines. Sci. Rep. 2015, 5, 8220. [Google Scholar] [CrossRef]
- Mowat, E.; Rajendran, R.; Williams, C.; McCulloch, E.; Jones, B.; Lang, S.; Ramage, G. Pseudomonas aeruginosa and their small diffusible extracellular molecules inhibit Aspergillus fumigatus biofilm formation. FEMS Microbiol. Lett. 2010, 313, 96–102. [Google Scholar] [CrossRef] [Green Version]
- Briard, B.; Heddergott, C.; Latgé, J.-P. Volatile compounds emitted by Pseudomonas aeruginosa stimulate growth of the fungal pathogen Aspergillus fumigatus. mBio 2016, 7, e00219-16. [Google Scholar] [CrossRef] [Green Version]
- Margalit, A.; Carolan, J.C.; Shehan, D.; Kavanagh, K. The Aspergillus fumigatus secretome alters the proteome of <em>Pseudomonas aeruginosa</em> to stimulate bacterial growth: Implications for co-infection. Mol. Cell. Proteom. 2020. [Google Scholar] [CrossRef]
- Hogan, D.A.; Vik, Å.; Kolter, R. A Pseudomonas aeruginosa quorum-sensing molecule influences Candida albicans morphology. Mol. Microbiol. 2004, 54, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Partida-Martinez, L.P.; Monajembashi, S.; Greulich, K.O.; Hertweck, C. Endosymbiont-dependent host reproduction maintains bacterial-fungal mutualism. Curr. Biol. CB 2007, 17, 773–777. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.K.; Jakobsen, M. Possible role of microbial interactions for growth and sporulation of Penicillium roqueforti in Danablu. Lait 1997, 77, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Adams, A.S.; Currie, C.R.; Cardoza, Y.; Klepzig, K.D.; Raffa, K.F. Effects of symbiotic bacteria and tree chemistry on the growth and reproduction of bark beetle fungal symbionts. Can. J. For. Res. 2009, 39, 1133–1147. [Google Scholar] [CrossRef]
- Deveau, A.; Palin, B.; Delaruelle, C.; Peter, M.; Kohler, A.; Pierrat, J.C.; Sarniguet, A.; Garbaye, J.; Martin, F.; Frey-Klett, P. The mycorrhiza helper Pseudomonas fluorescens BBc6R8 has a specific priming effect on the growth, morphology and gene expression of the ectomycorrhizal fungus Laccaria bicolor S238N. New Phytol. 2007, 175, 743–755. [Google Scholar] [CrossRef]
- Frey-Klett, P.; Burlinson, P.; Deveau, A.; Barret, M.; Tarkka, M.; Sarniguet, A. Bacterial-fungal interactions: Hyphens between agricultural, clinical, environmental, and food microbiologists. Microbiol. Mol. Biol. Rev. MMBR 2011, 75, 583–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, L.; Lin, Q.; Yao, Q.; Zhu, H. Population and function analysis of cultivable bacteria associated with spores of arbuscular mycorrhizal fungus. Gigaspora Margarita 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, U.; Ouziad, F.; Marner, F.J.; Bothe, H. The bacterium Paenibacillus validus stimulates growth of the arbuscular mycorrhizal fungus Glomus intraradices up to the formation of fertile spores. FEMS Microbiol. Lett. 2006, 254, 258–267. [Google Scholar] [CrossRef] [Green Version]
- Venkatesh, N.; Keller, N.P. Mycotoxins in conversation with bacteria and fungi. Front. Microbiol. 2019, 10, 403. [Google Scholar] [CrossRef]
- Reece, E.; Doyle, S.; Greally, P.; Renwick, J.; McClean, S. Aspergillus fumigatus inhibits Pseudomonas aeruginosa in co-culture: Implications of a mutually antagonistic relationship on virulence and inflammation in the CF airway. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Salvioli, A.; Ghignone, S.; Novero, M.; Navazio, L.; Venice, F.; Bagnaresi, P.; Bonfante, P. Symbiosis with an endobacterium increases the fitness of a mycorrhizal fungus, raising its bioenergetic potential. ISME J. 2016, 10, 130–144. [Google Scholar] [CrossRef]
- Dearth, S.P.; Castro, H.F.; Venice, F.; Tague, E.D.; Novero, M.; Bonfante, P.; Campagna, S.R. Metabolome changes are induced in the arbuscular mycorrhizal fungus Gigaspora margarita by germination and by its bacterial endosymbiont. Mycorrhiza 2018, 28, 421–433. [Google Scholar] [CrossRef]
- Bharadwaj, D.P.; Lundquist, P.-O.; Persson, P.; Alström, S. Evidence for specificity of cultivable bacteria associated with arbuscular mycorrhizal fungal spores. FEMS Microbiol. Ecol. 2008, 65, 310–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abeysinghe, G.; Kuchira, M.; Kudo, G.; Masuo, S.; Ninomiya, A.; Takahashi, K.; Utada, A.S.; Hagiwara, D.; Nomura, N.; Takaya, N.; et al. Fungal mycelia and bacterial thiamine establish a mutualistic growth mechanism. bioRxiv 2020. [Google Scholar] [CrossRef]
- Bravo, D.; Cailleau, G.; Bindschedler, S.; Simon, A.; Job, D.; Verrecchia, E.; Junier, P. Isolation of oxalotrophic bacteria able to disperse on fungal mycelium. FEMS Microbiol. Lett. 2013, 348, 157–166. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.; van Elsas, J.D. Mechanisms and ecological implications of the movement of bacteria in soil. Appl. Soil Ecol. 2018, 129, 112–120. [Google Scholar] [CrossRef]
- Pion, M.; Bshary, R.; Bindschedler, S.; Filippidou, S.; Wick, L.Y.; Job, D.; Junier, P. Gains of bacterial flagellar motility in a fungal world. Appl. Environ. Microbiol. 2013, 79, 6862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuno, S.; Päzolt, K.; Rabe, C.; Neu, T.R.; Harms, H.; Wick, L.Y. Fungal mycelia allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-unsaturated systems. Environ. Microbiol. 2010, 12, 1391–1398. [Google Scholar] [CrossRef]
- Banitz, T.; Frank, K.; Wick, L.Y.; Harms, H.; Johst, K. Spatial metrics as indicators of biodegradation benefits from bacterial dispersal networks. Ecol. Indic. 2016, 60, 54–63. [Google Scholar] [CrossRef]
- Worrich, A.; König, S.; Miltner, A.; Banitz, T.; Centler, F.; Frank, K.; Thullner, M.; Harms, H.; Kästner, M.; Wick, L.Y. Mycelium-like networks increase bacterial dispersal, growth, and biodegradation in a model ecosystem at various water potentials. Appl. Environ. Microbiol. 2016, 82, 2902. [Google Scholar] [CrossRef] [Green Version]
- Kohlmeier, S.; Smits, T.H.; Ford, R.M.; Keel, C.; Harms, H.; Wick, L.Y. Taking the fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environ. Sci. Technol. 2005, 39, 4640–4646. [Google Scholar] [CrossRef]
- Warmink, J.A.; van Elsas, J.D. Migratory response of soil bacteria to Lyophyllum sp. strain Karsten in soil microcosms. Appl. Environ. Microbiol. 2009, 75, 2820–2830. [Google Scholar] [CrossRef] [Green Version]
- Nazir, R.; Tazetdinova, D.I.; van Elsas, J.D. Burkholderia terrae BS001 migrates proficiently with diverse fungal hosts through soil and provides protection from antifungal agents. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef]
- Warmink, J.A.; Nazir, R.; Corten, B.; van Elsas, J.D. Hitchhikers on the fungal highway: The helper effect for bacterial migration via fungal hyphae. Soil Biol. Biochem. 2011, 43, 760–765. [Google Scholar] [CrossRef]
- Yang, P.; da Rocha Calixto, R.O.; van Elsas, J.D. Migration of Paraburkholderia terrae BS001 along old fungal hyphae in soil at various pH levels. Microb. Ecol. 2018, 76, 443–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Kastman, E.K.; Guasto, J.S.; Wolfe, B.E. Fungal networks shape dynamics of bacterial dispersal and community assembly in cheese rind microbiomes. Nat. Commun. 2018, 9, 336. [Google Scholar] [CrossRef] [Green Version]
- Simon, A.; Bindschedler, S.; Job, D.; Wick, L.Y.; Filippidou, S.; Kooli, W.M.; Verrecchia, E.P.; Junier, P. Exploiting the fungal highway: Development of a novel tool for the in situ isolation of bacteria migrating along fungal mycelium. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [Green Version]
- Kalan, L.; Loesche, M.; Hodkinson, B.P.; Heilmann, K.; Ruthel, G.; Gardner, S.E.; Grice, E.A. Redefining the chronic-wound microbiome: Fungal communities are prevalent, dynamic, and associated with delayed healing. mBio 2016, 7, e01058-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, O.A.; Noverr, M.C.; Peters, B.M. Candida albicans impacts Staphylococcus aureus alpha-toxin production via extracellular alkalinization. mSphere 2019, 4, e00780-19. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Cheng, W.; He, X.; Liu, Y. The co-colonization prevalence of Pseudomonas aeruginosa and Aspergillus fumigatus in cystic fibrosis: A systematic review and meta-analysis. Microb. Pathog. 2018, 125, 122–128. [Google Scholar] [CrossRef]
- Carolus, H.; Van Dyck, K.; Van Dijck, P. Candida albicans and Staphylococcus species: A threatening twosome. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef]
- Krüger, W.; Vielreicher, S.; Kapitan, M.; Jacobsen, I.D.; Niemiec, M.J. Fungal-bacterial interactions in health and disease. Pathogens 2019, 8, 70. [Google Scholar] [CrossRef] [Green Version]
- Diaz, P.I.; Strausbaugh, L.D.; Dongari-Bagtzoglou, A. Fungal-bacterial interactions and their relevance to oral health: Linking the clinic and the bench. Front. Cell. Infect. Microbiol. 2014, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, A.Y.; Hogan, D.A.; Mylonakis, E. Medically important bacterial–fungal interactions. Nat. Rev. Microbiol. 2010, 8, 340–349. [Google Scholar] [CrossRef] [PubMed]
- De Sordi, L.; Mühlschlegel, F.A. Quorum sensing and fungal–bacterial interactions in Candida albicans: A communicative network regulating microbial coexistence and virulence. FEMS Yeast Res. 2009, 9, 990–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Briard, B.; Mislin, G.L.A.; Latgé, J.-P.; Beauvais, A. Interactions between Aspergillus fumigatus and pulmonary bacteria: Current state of the field, new data, and future perspective. J. Fungi 2019, 5, 48. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, M.; Sugiyama, H.; Kizawa, K.; Hori, R.; Mitsuyama, J.; Araki, H.; Shimakura, M.; Minami, S.; Watanabe, Y.; Yamaguchi, K. A new model of pulmonary superinfection with Aspergillus fumigatus and Pseudomonas aeruginosa in mice. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2000, 6, 155–161. [Google Scholar] [CrossRef]
- Mitsuyama, J.; Kizawa, K.; Minami, S.; Watanabe, Y.; Yamaguchi, K. Evaluation of antimicrobial agents using an experimental pulmonary superinfection model with Aspergillus fumigatus and Pseudomonas aeruginosa in leukopenic mice. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2003, 9, 144–150. [Google Scholar] [CrossRef]
- Sharma, A.; Hofmeyr, A.; Bansal, A.; Thakkar, D.; Lam, L.; Harrington, Z.; Bhonagiri, D. COVID-19 associated pulmonary aspergillosis (CAPA): An Australian case report. Med. Mycol. Case Rep. 2020. [Google Scholar] [CrossRef]
- Fernandez, N.B.; Caceres, D.H.; Beer, K.D.; Irrazabal, C.; Delgado, G.; Farias, L.; Chiller, T.M.; Verweij, P.E.; Stecher, D. Ventilator-associated pneumonia involving Aspergillus flavus in a patient with coronavirus disease 2019 (COVID-19) from Argentina. Med. Mycol. Case Rep. 2020. [Google Scholar] [CrossRef]
- Koehler, P.; Cornely, O.A.; Böttiger, B.W.; Dusse, F.; Eichenauer, D.A.; Fuchs, F.; Hallek, M.; Jung, N.; Klein, F.; Persigehl, T.; et al. COVID-19 associated pulmonary aspergillosis. Mycoses 2020, 63, 528–534. [Google Scholar] [CrossRef]
- Whipps, J.M. Microbial interactions and biocontrol in the rhizosphere. J. Exp. Bot. 2001, 52, 487–511. [Google Scholar] [CrossRef]
- Eljounaidi, K.; Lee, S.K.; Bae, H. Bacterial endophytes as potential biocontrol agents of vascular wilt diseases–Review and future prospects. Biol. Control. 2016, 103, 62–68. [Google Scholar] [CrossRef]
- Dewey, F.M.; Wong, Y.L.; Seery, R.; Hollins, T.W.; Gurr, S.J. Bacteria associated with Stagonospora (Septoria) nodorum increase pathogenicity of the fungus. New Phytol. 1999, 144, 489–497. [Google Scholar] [CrossRef]
- Obasa, K.; White, F.F.; Fellers, J.; Kennelly, M.; Liu, S.; Katz, B.; Tomich, J.; Moore, D.; Shinogle, H.; Kelley, K. A dimorphic and virulence-enhancing endosymbiont bacterium discovered in Rhizoctonia solani. Phytobiomes J. 2017, 1, 14–23. [Google Scholar] [CrossRef]
- Sobrova, P.; Adam, V.; Vasatkova, A.; Beklova, M.; Zeman, L.; Kizek, R. Deoxynivalenol and its toxicity. Interdiscip. Toxicol. 2010, 3, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Saha, C.; Nag, M.; Mandal, D.; Naiya, H.; Sen, D.; Mitra, S.; Kumar, M.; Bose, D.; Mukherjee, G.; et al. A Tripartite Interaction among the Basidiomycete Rhodotorula mucilaginosa-fixing endobacteria, and rice improves plant nitrogen nutrition. Plant Cell 2020, 32, 486. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.; Romo, J.A.; Lamendella, R.; Kumamoto, C.A. The role of fungi in C. difficile infection: An underappreciated transkingdom interaction. Fungal Genet. Biol. 2019, 129, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calcagno, A.; Ghisetti, V.; Burdino, E.; Trunfio, M.; Allice, T.; Boglione, L.; Bonora, S.; Di Perri, G. Co-infection with other respiratory pathogens in COVID-19 patients. Clin. Microbiol. Infect. 2020. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mediators | BFI | Citation |
|---|---|---|
| Surface molecules | ||
| Als Proteins and O-mannosylations | S. epidermidis-C. albicans | Beaussart, 2013 |
| Als3p | S. gordonii-C. albicans S. aureus-C. albicans | Silverman, 2010 Peters, 2012 |
| Als adhesins, SasF, Atl | S. aureus-C. albicans | Schlecht, 2015 |
| CbpD | P. aeruginosa-C. albicans | Ovchinnikova, 2013 |
| T2SS | M. rhizoxinica-R. microsporus | Moebius, 2014 |
| T3SS | M. rhizoxinica-R. microsporus | Lackner, 2011 |
| T2SS, T3SS, T4SS, gspD, sec system | M. rhizoxinica-R. microsporus | Ghignone, 2012 |
| TAL effector | M. rhizoxinica-R. microsporus | Richter, 2020; Carter, 2020 |
| Genes | ||
| gspD, secB | Ca. G. gigasporum-G. margarita | Ghignone, 2011 |
| vacB | Burkholderia sp.-G. margarita | Ruiz-Lozano & Bonfante, 2000 |
| spo0A | B. subtilis-A. niger B. subtilis-Ag. bisporus | Kjeldgaard, 2019 |
| Secreted Factors | ||
| EPS and TasA | B. subtilis-A. niger B. subtilis-A. bisporus | Kjeldgaard, 2019 |
| GAG, pyo-melanin, 1,8-dihydroxynaphthalene- melanin | P. aeruginosa-A. fumigatus | Briard, 2017 |
| ECM | S. aureus- C. albicans | Harriot and Noverr, 2009 |
| Holrhizin A | M. rhizoxinica-R. microsporus | Moebius, 2014 |
| Ralsolamycin | R. solanacearum-A. flavus | Spraker, 2016 |
| Progidiosin, T6SS, TssJ, murein lipoprotein | S. marcescens-M. irregularis | Hazarika, 2020 |
| Other | ||
| Acid-Base Attractive Forces | P. aeruginosa-C. albicans | Ovchinnikova, 2012 |
| Fungal Viability | B. cereus-Glomus sp. P. peoriae-Glomus sp. P. brasilensis-Glomus sp. S. marcescens-A. fumigatus S. marcescents-R. oryzae | Toljander, 2006 Hover, 2016 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steffan, B.N.; Venkatesh, N.; Keller, N.P. Let’s Get Physical: Bacterial-Fungal Interactions and Their Consequences in Agriculture and Health. J. Fungi 2020, 6, 243. https://doi.org/10.3390/jof6040243
Steffan BN, Venkatesh N, Keller NP. Let’s Get Physical: Bacterial-Fungal Interactions and Their Consequences in Agriculture and Health. Journal of Fungi. 2020; 6(4):243. https://doi.org/10.3390/jof6040243
Chicago/Turabian StyleSteffan, Breanne N., Nandhitha Venkatesh, and Nancy P. Keller. 2020. "Let’s Get Physical: Bacterial-Fungal Interactions and Their Consequences in Agriculture and Health" Journal of Fungi 6, no. 4: 243. https://doi.org/10.3390/jof6040243
APA StyleSteffan, B. N., Venkatesh, N., & Keller, N. P. (2020). Let’s Get Physical: Bacterial-Fungal Interactions and Their Consequences in Agriculture and Health. Journal of Fungi, 6(4), 243. https://doi.org/10.3390/jof6040243