
A Possibility to Use Selected Crop Post-Extraction Wastes to Improve the Composition of Cultivated Mushroom Pleurotus citrinopileatus

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
Abstract
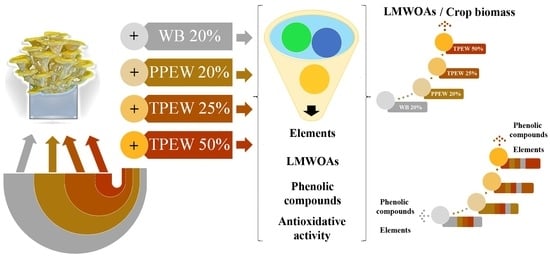
:
1. Introduction
2. Materials and Methods
2.1. Experimental Material and Experimental Design
2.2. Elemental Analysis
2.2.1. Sample Preparation
2.2.2. Instruments and Analytical Method Validation
2.3. Analysis of Low-Molecular-Weight Organic Acids and Phenolic Compounds
2.4. Determination of Total Phenolic Contents
2.5. Antioxidative Activity
2.6. The Statistical Analysis and Calculations
3. Results
3.1. Morphology and Biomass of Mushroom Fruit Bodies
3.2. Contents of Major Essential Elements
3.3. Contents of Trace Elements in Fruit Bodies
3.4. Profile and Content of LMWOAs in Mushrooms
3.5. Characteristics of the Phenolic Profile and Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ohira, I. A revision of the taxonomic status of Pleurotus citrinopileatus. Rep. Tottori Mycol. Inst. 1990, 28, 142–150. [Google Scholar]
- FAOSAT. 2016. Available online: http://faostat3.fao.org/browse/Q/QC/E (accessed on 17 July 2020).
- United States Department of Agriculture (USDA). Mushrooms National Agricultural Statistics Service (NASS) Agricultural Statistics Board United States Department of Agriculture (USDA). 2017. Available online: https://www.nass.usda.gov/ (accessed on 15 July 2020).
- Zhang, J.; Wang, G.; Li, H.; Zhuang, C.; Mizuno, T.; Ito, H.; Suzuki, C.; Okamoto, H.; Li, J. Antitumor polysaccharides from a Chinese mushroom, “yuhuangmo”, the fruit body of Pleurotus citrinopileatus. Biosci. Biotechnol. Biochem. 1994, 58, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Ziombra, M.; Czerwińska, A. Boczniaki mniej znane: Boczniak cytrynowy (Pleurotus citrinopileatus). Biul. Prod. Pieczarek Pieczarki 2006, 34, 64–67. [Google Scholar]
- Cohen, R.; Persky, L.; Hadar, Y. Biotechnological applications and potential of wood degrading mushrooms of the genus Pleurotus. Appl. Microbiol. Biotechnol. 2002, 58, 582–594. [Google Scholar] [CrossRef]
- Ponmurugan, P.; Sekhar, Y.N.; Sreesakthi, T. Effect of various substrates on the growth and quality of mushrooms. Pak. J. Biol. Sci. 2007, 10, 171–173. [Google Scholar] [CrossRef]
- Alam, N.; Yoon, K.; Lee, K.; Kim, H.; Shin, P.; Cheong, J.; Yoo, Y.; Shim, M.; Lee, M.; Lee, T. Assessment of antioxidant and phenolic compound concentrations as well as xanthine oxidase and tyrosinase inhibitory properties of different extracts of Pleurotus citrinopileatus fruit bodies. Mycobiology 2011, 39, 12–19. [Google Scholar] [CrossRef] [Green Version]
- Naik, V.; Sharma, D.; Kumar, P.; Yadav, R. Efficacy of ligno-cellulolytic fungi on recycling sericultural waste. Acta Biol. Indica 2012, 1, 47–50. [Google Scholar]
- Pandey, V.; Singh, M.; Srivastava, A.; Vishwakarma, S.; Takshak, S. Biodegradation of sugarcane bagasse by Pleurotus citrinopileatus. Cell Mol. Biol. 2012, 58, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Owaid, M.N.; Al-Saeedi, S.S.S.; Abed, I.A. Recycling of date-palm fiber to produce Pleurotus cornucopiae var. citrinopileatus mushroom. Int. J. Environ. 2016, 5, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Joshi, S.; Borkar, P.G.; Saykar, A.D.; Pawar, S.V. Assessment of biological efficiency of Pleurotus Sajor-caju, P. florida, P. citrinopileatus and Hypsizygus. Int. J. Chem. Stud. 2018, 6, 2299–2301. [Google Scholar]
- Chaudhary, L.C.; Singh, R.; Kamra, D.N. Biodelignification of sugarcane bagasse by Pleurotus florida and Pleurotus cornucopiae. Indian J. Microbiol. 1994, 34, 55–57. [Google Scholar]
- Ćilerdžić, J.; Galić, M.; Vukojević, J.; Brčeski, I.; Stajić, M. Potential of selected fungal species to degrade wheat straw, the most abundant plant raw material in Europe. BMC Plant Biol. 2017, 17, 249. [Google Scholar] [CrossRef] [Green Version]
- Krishna, B.; Shanmugasundaram, K.; Narendhirakannan, R.T. Mycelial growth characteristics of Pleurotus citrinopileatus and Pleurotus platypus in culture media variants. Indian J. Agric. Biochem. 2019, 32, 200–205. [Google Scholar] [CrossRef]
- Siwulski, M.; Sobieralski, K. Uprawa Grzybów Jadalnych i Leczniczych w Warunkach Naturalnych; Kurpisz Poznań: Poznań, Poland, 2004. [Google Scholar]
- Stajic, M.; Milenkovic, I.; Brceksi, I.; Vukojevic, J.; Duletic, S. Mycelial growth of edible and medicinal oyster mushroom Pleurotus ostreatus (Jacq.Fr.Kumm) on selenium enriched media. Int. J. Med. Mushrooms 2002, 4, 241–244. [Google Scholar] [CrossRef]
- Parmar, R.; Kumar, D. Study of chemical composition in wild edible mushroom Pleurotus cornucopiae (Paulet) from Himachal Pradesh, India by using fourier transforms infrared spectrometry (FTIR), gas chromatography-mass spectrometry (GCMS) and x-ray fluorescence (XRF). Biol. Forum 2015, 7, 1057. [Google Scholar]
- Tan, Y.S.; Baskaran, A.; Nallathamby, N.; Chua, K.H.; Kuppusamy, U.R.; Sabaratnam, V. Influence of customized cooking methods on the phenolic contents and antioxidant activities of selected species of oyster mushrooms (Pleurotus spp.). J. Food Sci. Technol. 2015, 52, 3058–3064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siwulski, M.; Mleczek, M.; Rzymski, P.; Budka, A.; Jasińska, A.; Niedzielski, P.; Kalač, P.; Gasecka, M.; Budzyńska, S.; Mikołajczak, P. Screening the multi-element content of Pleurotus mushroom species using inductively coupled plasma optical emission spectrometer (ICP-OES). Food Anal. Methods 2017, 10, 487–496. [Google Scholar] [CrossRef] [Green Version]
- Peraza, J.G.; Grycz, A.; Mederos, M.A.P.; Galdón, B.R.; Rodríguez, E.M.R. Total phenolic compounds and antioxidant activity in Pleurotus spp. grown on commercial and wild substrates. Glob. J. Food Sci. Nutr. 2019, 4, 1–6. [Google Scholar] [CrossRef]
- Lee, Y.L.; Huang, G.W.; Liang, Z.C.; Mau, J.L. Antioxidant properties of three extract from Pleurotus citrinopileatus. LWT Food Sci. Technol. 2007, 40, 823–833. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, B. Distribution of antioxidant activities and total phenolic contents in acetone, ethanol, water and hot water extracts from 20 edible mushrooms via sequential extraction. Austin J. Nutr. Food Sci. 2014, 2, 5. [Google Scholar]
- Khatun, S.; Islam, A.; Cakilcioglu, U.; Guler, P.; Chatterjee, N.C. Nutritional qualities and antioxidant activity of three edible oyster mushrooms (Pleurotus spp.). NJAS Wagening. J. Life Sci. 2015, 72, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, D.M.F.C.; Freitas, A.C.; Rocha-Santos, T.A.P.; Vasconcelos, M.W.; Roriz, M.; Rodríguez-Alcalá, L.M.; Gomes, A.M.P.; Duarte, A.C. Chemical composition and nutritive value of Pleurotus citrinopileatus var cornucopiae, P. eryngii, P. salmoneo stramineus, Pholiota nameko and Hericium erinaceus. J. Food Sci. Technol. 2015, 52, 6927–6939. [Google Scholar] [CrossRef]
- Lin, S.Y.; Chien, S.C.; Wang, S.Y.; Mau, J.L. Non-volatile taste components and antioxidant properties of fruit body and mycelium with high ergothioneine content from the culinary-medicinal golden oyster mushroom Pleurotus citrinopileatus (Agaricomycetes). Int. J. Med. Mushrooms 2016, 18, 689–698. [Google Scholar] [CrossRef]
- Yıldız, S.; Yılmaz, A.; Can, Z.; Kılıç, C.; Yıldız, Ü.C. Total phenolic, flavonoid, tannin contents and antioxidant properties of Pleurotus ostreatus and Pleurotus citrinopileatus cultivated on various sawdust. GIDA J. Food 2017, 42, 315–323. [Google Scholar] [CrossRef]
- Chang, S.; Miles, P. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact, 2nd ed.; CRC Press LLC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Hu, S.; Wang, J.; Lien, J.; Liaw, E.; Lee, M. Antihyperglycemic effect of polysaccharide from fermented broth of Pleurotus citrinopileatus. Appl. Microbiol. Biotechnol. 2005, 70, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Liang, Z.; Chia, Y.; Lien, J.; Chen, K.; Lee, M.; Wang, J. Antihyperlipidemic and Antioxidant Effects of Extracts from Pleurotus citrinopileatus. J. Agric. Food Chem. 2006, 54, 2103–2110. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, C.; Wang, J. The evaluation of using mushroom sawdust waste for cultivation of Pleurotus citrinopileatus. Fungi Sci. 2005, 20, 27–34. [Google Scholar]
- Li, Y.; Liu, Q.; Wang, H.; Ng, T. A novel lectin with potent antitumor, mitogenic and HIV-1 reverse transcriptase inhibitory activities from the edible mushroom Pleurotus citrinopileatus. Biochim. Biophys. Acta 2008, 1780, 51–57. [Google Scholar] [CrossRef]
- Minato, K. Immunomodulation activity of a polysaccharide fraction of a culinary-medicinal mushroom, Pleurotus citrinopileatus Singer (Agaricomycetideae) in vitro. Int. J. Med. Mushrooms 2008, 10, 235–244. [Google Scholar] [CrossRef]
- Chen, J.; Wang, Y.; Wu, J. A glycoprotein extracted from golden oyster mushroom Pleurotus citrinopileatus exhibiting growth inhibitory effect against U937 leukemia cells. J. Agric. Food Chem. 2009, 57, 6706–6711. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ma, C.; Tsai, P.; Wang, Y.; Wu, J. In vitro antitumor and immunomodulatory effects of the protein PCP-3A from mushroom Pleurotus citrinopileatus. J. Agric. Food Chem. 2010, 58, 12117–12122. [Google Scholar] [CrossRef]
- Chen, J.; de Mejia, E.G.; Wu, J. Inhibitory effect of a glycoprotein isolated from golden oyster mushroom (Pleurotus citrinopileatus) on the lipopolysaccharide-induced inflammatory reaction in RAW 264.7 Macrophage. J. Agric. Food Chem. 2011, 59, 7092–7097. [Google Scholar] [CrossRef]
- Xue, B.; Jin, Z.; Shi, S. Antioxidant Activities of Pleurotus citrinopileatus in Fresh and Dired Character. Food Res. Dev. 2011, 3–10. [Google Scholar]
- Wu, S.; Gao, S.; Liu, H.; Sun, X.; Hao, L.; Jia, L.; Pang, L.; Jia, S.; Jia, M. Intracellular polysaccharide and its antioxidant activity by Pleurotus citrinopileatus SM-01. Macromol. Res. 2013, 21, 660–668. [Google Scholar] [CrossRef]
- Younis, A.M.; Abdel-Aziz, M.M.; Yosri, M. Evaluation of Some Biological Applications of Pleurotus citrinopileatus and Boletus edulis Fruit Bodies. Curr. Pharm. Biotechnol. 2019, 20, 1309. [Google Scholar] [CrossRef] [PubMed]
- Dubost, N.J.; Ou, B.; Beelman, R.B. Quantification of polyphenols and ergothioneine in cultivated mushrooms and correlation to total antioxidant capacity. Food Chem. 2007, 105, 727–735. [Google Scholar] [CrossRef]
- Reis, F.S.; Martins, A.; Barros, L.; Ferreira, I.C.F.R. Antioxidant properties and phenolic profile of the mostwidely appreciated cultivated mushrooms: A comparative study between in vivo and in vitro samples. Food Chem. Toxicol. 2012, 50, 1201–1207. [Google Scholar] [CrossRef]
- Taofiq, O.; Calhelha, R.C.; Heleno, S.; Barros, L.; Martins, A.; Santos-Buelga, C.; Quieroz, M.J.R.P.; Ferreira, I.C. The contribution of phenolic acids to the anti-inflammatory activity of mushrooms: Screening in phenolic extracts, individual parent molecules and synthesized glucuronated and methylated derivatives. Food Res. Int. 2015, 76, 821–827. [Google Scholar] [CrossRef] [Green Version]
- Islam, T.; Yu, X.; Xu, B. Phenolic profiles, antioxidant capacities and metal chelating ability of edible mushrooms commonly consumed in China. LWT Food Sci. Technol. 2016, 72, 423–431. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P.; Kozak, L. The effect of selenium on phenolics and flavonoids in selected edible white rot fungi. LWT Food Sci. Technol. 2015, 163, 726–731. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Atila, F.; Tuzel, Y.; Fernández, J.A.; Cano, A.F.; Sen, F. The effect of some agro–industrial wastes on yield, nutritional characteristics and antioxidant activities of Hericium erinaceus isolates. Sci. Hortic. 2018, 238, 246–254. [Google Scholar] [CrossRef]
- Fasehah, S.N.; Shah, A. Effect of using various substrates on cultivation of Pleurotus Sajor Caju. J. Eng. Sci. Technol. 2017, 12, 1104–1110. [Google Scholar]
- Suganthi, V.; Krishnakumari, S. Efficacy of different agricultural by product substrates and moisture content on mycelial growth of Pleurotus cornucopiae. J. Pharmacol. Phytochem. 2018, 7, 3407–3409. [Google Scholar]
- Mukhopadhyay, S.B. Oyster Mushroom Cultivation on Water Hyacinth Biomass: Assessment of Yield Performances, Nutrient, and Toxic Element Contents of Mushrooms. In An Introduction to Mushroom; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Siwulski, M.; Sobieralski, K.; Korszun, S. Effect of the leaf extract from the maidenhair tree (Ginkgo biloba L.) on the growth of the oyster mushroom (Pleurotus spp.) mycelium. Herba Pol. 2007, 53, 28–32. [Google Scholar]
- Valentão, P.; Lopes, G.; Valente, M.; Barbosa, P.; Andrade, P.B.; Silva, B.M.; Baptista, P.; Seabra, R.M. Quantification of nine organic acids in wild mushrooms. J. Agric. Food Chem. 2005, 53, 3626–3630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Gu, Z.; Yang, Y.; Zhou, S.; Liu, Y.; Zhang, J. Non-volatile taste components of several cultivated mushrooms. Food Chem. 2014, 143, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Pereira, C.; Ferreira, C.F.R. Optimized analysis of organic acids in edible mushrooms from Portugal by ultra fast liquid chromatography and photodiode array detection. Food Anal. Methods 2013, 6, 309–316. [Google Scholar] [CrossRef]
- Magdziak, Z.; Mleczek, M.; Siwulski, M. Characteristics of organic acid profiles in 16 species of wild growing edible mushrooms. J. Environ. Sci. Health B 2017, 52, 784–789. [Google Scholar] [CrossRef]
- Stamets, P. Growing Gourmet & Medicinal Mushrooms, 3rd ed.; Random House USA Inc.: Barkeley, KA, USA, 2016. [Google Scholar]
- Gąscka, M.; Magdziak, Z.; Siwulski, M.; Jasińska, A.; Budzyńska, S.; Rzymski, P.; Kalač, P.; Niedzielski, P.; Mleczek, M.; Pankiewicz, J. Effect of Thymus vulgaris post-extraction waste and spent coffee grounds on quality of cultivated Pleurotus eryngii. J. Food Process. Preserv. 2020, 44, e14648. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin–Ciocalteu reagent. Method Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, M.; Lu, L.; Sun, L.; Xu, M. Nitric oxide fumigation stimulates flavonoid and phenolic accumulation and enhances antioxidant activity of mushroom. Food Chem. 2012, 135, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Kalač, P. Mineral Composition and Radioactivity of Edible Mushrooms, 1st ed.; Elsevier: New York, NY, USA, 2019. [Google Scholar]
- Tabata, T.; Ogura, T. Absorption of Calcium and Magnesium to the Fruit Body of Aragekikurage (Auricularia polytricha (Mont.) Sacc.) from Sawdust Culture Media Supplemented with Calcium and Magnesium Salts. Food Sci. Technol. Res. 2003, 9, 250–253. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Kalia, A.; Sodhi, H.S. Selenium biofortification of Pleurotus species and its effects on yield, phytochemical profiles, and protein chemistry of fruiting bodies. J. Food Biochem. 2018, 42, e12467. [Google Scholar] [CrossRef]
- Sturion, G.L.; Oetterer, M. Composição química de cogumelos comestíveis (Pleurotus spp.) originados de cultivos em diferentes substratos. Ciênc. Tecnol. Aliment. 1995, 15, 189–193. [Google Scholar]
- Singh, M.P.; Singh, V.K. Yield performance and nutritional analysis of Pleurotus citrinopileatus on different agrowastes and vegetable wastes. In Proceedings of the 7th International Conference on Mushroom Biology and Mushroom Products, Arcachon, France, 4–7 October 2011. [Google Scholar]
- Owaid, M.N.; Abed, A.M.; Nassar, B.M. Recycling cardboard wastes to produce blue oyster mushroom Pleurotus ostreatus in Iraq. Emir. J. Food Agric. 2015, 27, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Donini, L.P.; Bernardi, E.; Minotto, E.; Nascimento, J.S. Growing Shimeji on elephant grass substrate supplemented with different types of sharps. Sci. Agrar. 2009, 10, 67–74. [Google Scholar]
- Fanadzo, M.; Zireva, D.T.; Dube, E.; Mashingaidze, A.B. Evaluation of various substrates and supplements for biological efficiency of Pleurotus sajor-caju and Pleurotus ostreatus. Afr. J. Biotechnol. 2010, 9, 2756–2761. [Google Scholar] [CrossRef]
- Kalmis, E.; Azbar, N.; Yildiz, H.; Kalyoncu, F. Feasibility of using olive mill effluent (OME) as a wetting agent during the cultivation of oyster mushroom, Pleurotus ostreatus, on wheat straw. Bioresour. Technol. 2010, 99, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Kabata-Pendias, A.; Pendias, H. Biogeochemia Pierwiastków Śladowych. [Biogeochemistry of Trace Elements]; Wydawnictwo Naukowe PWN: Warsaw, Poland, 1999. (In Polish) [Google Scholar]
- Li, W.; Chen, W.; Yang, Y.; Zhang, J.; Feng, J.; Yu, H.; Zhou, S.; Li, X.; Liu, Y. Effects of culture substrates on taste component content and taste quality of Lentinula edodes. Int. J. Food Sci. Technol. 2017, 52, 981–991. [Google Scholar] [CrossRef]
- Ousmanova, D.; Parker, W. Fungal generation of organic acids for removal of lead from contaminated soil. Water Air Soil Pollut. 2007, 179, 365–380. [Google Scholar] [CrossRef]
- Brennan, M.; Port, G.L.; Gormley, R. Post-harvest treatment with citric acid or hydrogen peroxide to extend the shelf life of fresh sliced mushrooms. LWT Food Sci. Technol. 2000, 33, 285–289. [Google Scholar] [CrossRef]
- Yadav, M.; Jain, S.; Tomar, R.; Prasad, G.B.; Yadav, H. Medicinal and biological potential of pumpkin: An updated review. Nutr. Res. Rev. 2010, 23, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Montesano, D.; Rocchetti, G.; Putnik, P.; Lucini, L. Bioactive profile of pumpkin: An overview on terpenoids and their health-promoting properties. Curr. Opin. Food Sci. 2018, 22, 81–87. [Google Scholar] [CrossRef]
- Rekiel, A.; Grzesiuk, K.; Sońta, M. Dynia Pastewna—Uprawa, Pozyskanie, Badania, Wykorzystanie Paszowe. Przegląd Hodowlany nr 2; Szkola Gospodarstwa Wiejskiego w Warszawie: Warsaw, Poland, 2019. (In Polish) [Google Scholar]
- Imelouane, B.; Amhamdi, H.; Wathelet, J.P.; Ankit, M.; Khedid, K.; Bachiri, E.L. Chemical composition and antimicrobial activity of essential oil of Thyme (Thymus vulgaris) from Eastern Morocco. Int. J. Agric. Biol. 2009, 11, 205–208. [Google Scholar]
- Zhao, L.; Zhao, G.; Zhao, Z.; Chen, P.; Tong, J.; Hu, X. Selenium distribution in a Se-enriched mushroom species of the genus Ganoderma. J. Agric. Food Chem. 2004, 52, 3954–3959. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, C.; Yang, J.; Chen, J.; Yuan, T.; Zhao, J. Collection of group characteristics of Pleurotus eryngii using machine vision. In Proceedings of the International Conference on Computer and Computing Technologies in Agriculture, Jilin, China, 12–15 August 2009; Springer: Berlin/Heidelberg, Germany,, 2009; pp. 98–103. [Google Scholar]
- Stojković, D.S.; Kovačević-Grujičić, N.; Reis, F.S.; Davidović, S.; Barros, L.; Popocić, J.; Petrović, I.; Pavić, A.; Glamočlija, J.; Ćirić, A.; et al. Chemical composition of the mushroom Meripilus giganteus Karst. and bioactive properties of its methanolic extract. LWT Food Sci. Technol. 2017, 79, 454–462. [Google Scholar] [CrossRef] [Green Version]
- Karamn, M.; Tesanovic, K.; Gorjanovic, S.; Pastor, F.T.; Simanovic, M.; Glimac, M.; Pejin, B. Polarography as a technique of choice for the evaluation of total antioxidant activity: The case study of selected Coprinus Comatus extracts and quinic acid, their antidiabetic ingredient. Nat. Prod. Res. 2019, 13, 1711–1716. [Google Scholar] [CrossRef]
- Gürgen, S.; Yildiz, S. Artificial neural network approach for protection of the color of dried golden and pink oyster mushrooms with pretreatments. Color Res. Appl. 2019, 44, 1006–1016. [Google Scholar] [CrossRef]
- Gogoi, P.; Chutia, P.; Singh, P.; Mahant, C.L. Effect of optimized ultrasound-assisted aqueous and ethanolic extraction of Pleurotus citrinopileatus mushroom on total phenol, flavonoids and antioxidant properties. J. Food Process. Eng. 2019, 42, e13172. [Google Scholar] [CrossRef]
- Nattoh, G.; Gatebe, E.; Musieba, F.; Mathara, J. Bioprospecting optimal phenology for bioactive molecules in native golden yellow Pleurotus citrinopileatus Singer. Asian Pac. J. Trop. Biomed. 2016, 6, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Rodriguez, A.; Soler-Rivas, C.; Polonia, I.; Wichers, J.H. Effect of olive mill waste (OMW) supplementation to oyster mushrooms substrates on the cultivation parameters and fruit bodies quality. Int. Biodeterior. Biodegrad. 2010, 64, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Pandey, R.; Vyas, D. Antioxidant potentiality of Pleurotus ostreatus (MTCC142) cultivated on different agro wastes. Asian J. Plant Sci. Res. 2015, 5, 22–27. [Google Scholar]
- Sardar, H.; Ali, M.A.; Anjum, M.A.; Nawaz, F.; Hussain, S.; Naz, S.; Karimi, S.M. Agro-industrial residues influence mineral elements accumulation and nutritional composition of king oyster mushroom (Pleurotus eryngii). Sci. Hortic. 2017, 225, 327–334. [Google Scholar] [CrossRef]
- Freitas, A.C.; Antunes, M.B.; Rodrigues, D.; Sousa, S.; Amorim, M.; Barroso, M.F.; Carvalho, A.; Ferrador, S.M.; Gomes, A.M. Use of coffee by-products for the cultivation of Pleurotus citrinopileatus and Pleurotus salmoneo-stramineus and its impact on biological properties of extracts thereof. Int. J. Food Sci. Technol. 2018, 53, 1914–1924. [Google Scholar] [CrossRef]
- da Paz, M.F.; Breyer, C.A.; Longhi, R.F.; Vicente, M.S.; Oviedo, P. Determining the basic composition and total phenolic compounds of Pleurotus sajor-caju cultivated in three different substrates by solid state bioprocess. J. Biotechnol. Biodivers. 2012, 3, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.H.; Weng, Y.M.; Lin, S.M.; Yu, Z.R.; Wang, B.J. Molecular weight affected antioxidant, hypoglycemic and hypotensive activities of cold water extract from Pleurotus citrinopileatus. J. Food Sci. 2017, 82, 2456–2461. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Kalogeropoulos, N.; Kaliora, A.C.; Zervakis, G.I. Toward an increased functionality in oyster (Pleurotus) mushrooms produced on grape marc or olive mill wastes serving as sources of bioactive compounds. J. Agric. Food Chem. 2018, 66, 5971–5983. [Google Scholar] [CrossRef]
- Koutrotsios, G.; Larou, E.; Mountzouris, K.C.; Zervakis, G. Detoxification of olive millwastewater and bioconversion of olive crop residues into high-value-added biomass by the choice edible mushroom Hericium erinaceus. Appl. Biochem. Biotechnol. 2016, 180, 195–209. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Control | WB 20% | PPEW 20% | TPEW 25% | TPEW 50% |
|---|---|---|---|---|---|
| Ca | 0.39 | 0.56 | 0.64 | 0.71 | 0.76 |
| K | 1.20 | 1.83 | 2.42 | 2.03 | 1.90 |
| Mg | 1.13 | 1.66 | 1.49 | 0.66 | 1.63 |
| Na | 1.18 | 1.07 | 0.72 | 1.78 | 1.77 |
| S | 0.37 | 0.94 | 0.60 | 1.00 | 1.16 |
| Ag | 0.95 | 1.11 | 0.33 | 1.04 | 1.13 |
| Al | 0.40 | 1.14 | 0.83 | 1.44 | 1.27 |
| B | 0.15 | 0.33 | 0.15 | 1.60 | 3.11 |
| Ba | 1.06 | 0.67 | 1.88 | 1.92 | 0.91 |
| Bi | 2.69 | 0.67 | 0.14 | 0.80 | 0.94 |
| Cd | 3.05 | 0.39 | 2.75 | 0.15 | 1.33 |
| Ce | 4.43 | 2.65 | 1.30 | 0.77 | 0.72 |
| Cr | 0.65 | 0.73 | 0.44 | 1.07 | 0.81 |
| Cu | 4.66 | 2.54 | 1.39 | 2.04 | 1.86 |
| Er | 5.31 | 1.14 | 2.18 | 0.76 | 2.06 |
| Fe | 1.92 | 0.75 | 1.21 | 1.88 | 1.59 |
| Ge | 3.29 | 1.06 | 0.18 | 0.65 | 1.09 |
| Hf | 2.73 | 0.16 | 0.30 | 0.75 | 0.38 |
| Hg | 0.53 | 0.11 | 0.32 | 0.30 | 0.26 |
| In | 1.19 | 0.36 | 0.35 | 0.48 | 0.53 |
| Ir | 1.25 | 0.37 | 0.08 | 0.58 | 0.38 |
| La | 1.17 | 0.61 | 0.20 | 0.76 | 0.50 |
| Mn | 6.96 | 0.65 | 0.98 | 0.90 | 0.67 |
| Mo | 2.74 | 0.48 | 0.56 | 0.66 | 0.32 |
| Nd | 2.24 | 1.06 | 0.29 | 0.67 | 0.35 |
| Ni | 0.74 | 0.37 | 0.17 | 0.37 | 0.40 |
| Os | 2.64 | 0.56 | 0.64 | 1.17 | 0.86 |
| Pb | 0.86 | 0.16 | 0.05 | 0.25 | 0.23 |
| Pr | 1.86 | 0.53 | 0.73 | 0.39 | 0.80 |
| Pt | 0.90 | 0.48 | 0.14 | 0.31 | 0.34 |
| Rb | 2.02 | 1.00 | 0.60 | 0.71 | 1.02 |
| Ru | 3.26 | 0.38 | 1.43 | 1.52 | 0.72 |
| Sb | 1.33 | 0.82 | 0.46 | 0.91 | 0.89 |
| Se | 2.46 | 1.24 | 0.40 | 0.22 | 0.77 |
| Si | 1.38 | 0.79 | 0.41 | 0.93 | 0.75 |
| Sn | 9.97 | 1.22 | 1.86 | 0.06 | 1.59 |
| Sr | 2.67 | 0.72 | 0.85 | 1.37 | 1.19 |
| Te | 4.32 | 1.91 | 3.88 | 1.59 | 3.59 |
| Th | 11.3 | 2.09 | 0.73 | 0.60 | 2.31 |
| Ti | 4.79 | 1.17 | 0.26 | 0.91 | 0.71 |
| W | 3.18 | 1.82 | 0.23 | 0.82 | 0.72 |
| Zn | 1.33 | 1.28 | 1.67 | 1.93 | 1.29 |
| Zr | 1.65 | 1.07 | 0.52 | 1.27 | 0.87 |
| Acid | Control | WB 20% | PPEW 20% | TPEW 25% | TPEW 50% |
|---|---|---|---|---|---|
| Acetic | nd | 0.129 a ± 0.0632 | nd | nd | nd |
| Citric | 0.941 d ± 0.0171 | nd | 689 c ± 33.7 | 1055 b ± 19.4 | 5536 a ± 239 |
| Fumaric | nd | nd | 7.49 a ± 0.366 | nd | nd |
| Maleic | nd | 0.587 a ± 0.0287 | nd | nd | nd |
| Malic | nd | 10.5 c ± 0.211 | 134 b ± 6.50 | 49.9 c ± 6.93 | 3262 a ± 138 |
| Malonic | 1.92 c ± 0.350 | nd | 471 b ± 23.0 | 663 a ± 12.2 | nd |
| Oxalic | nd | 0.148 c ± 0.0724 | 53.9 a ± 2.61 | 41.8 b ± 3.77 | nd |
| Quinic | 0.609 c ± 0.0112 | 2.36 c ± 0.115 | nd | 581 b ± 10.7 | 2431 a ± 105 |
| Succinic | 1.71 b ± 0.000311 | 4.11 b ± 0.201 | nd | 49.7 a ± 5.9 | nd |
| Sum | 5.19 d ± 0.948 | 17.8 d ± 1.871 | 1355 c ± 66.2 | 2440 b ± 145 | 11229 a ± 485 |
| Compound | Control | WB 20% | PPEW 20% | TPEW 25% | TPEW 50% |
|---|---|---|---|---|---|
| Caffeic | nd | 151 ± 11.9 | nd | nd | nd |
| Catechin | nd | 407 a ± 14.1 | 25.9 b ± 1.83 | 20.2 b ± 0.919 | 2.21 c ± 0.171 |
| Chlorogenic acid | 2.73 b ± 0.143 | 853.4 a ± 0.00775 | 3.23 b ± 0.0841 | nd | nd |
| p-Coumaric acid | nd | 251 ± 21.1 | nd | nd | nd |
| 2,5-DHBA | nd | 2162 a ± 112.3 | 33.7 b ± 0.757 | 23.3 b ± 02.67 | 55.8 b ± 4.76 |
| Ferulic acid | nd | 152 a ± 04.50 | nd | 2.0 b ± 0.151 | 2.10 b ± 0.553 |
| Gallic acid | 13.2 b ± 0.989 | 2190 a ± 149 | 60.7 b ± 01.32 | 36.0 b ± 1.38 | 148 b ± 21.6 |
| 4-HBA | nd | nd | 143a ± 14.5 | 66.1 b ± 1.87 | nd |
| Protocatechuic acid | nd | 622 a ± 16.1 | 22.3 b ± 0.481 | 23.6 b ± 0.745 | 24.3 b ± 3.61 |
| Rutin | nd | 497 a ± 25.4 | nd | 3.07 b ± 0.121 | 2.98 b ± 0.165 |
| Salicylic acid | nd | 858 a ± 38.5 | 1.63 b ± 0.135 | 7.69 b ± 0.455 | 6.90 b ± 0.177 |
| Sinapic acid | nd | 331 a ± 18.1 | 1.92 b ± 1.28 | nd | nd |
| Syringic acid | 16.5 b ± 1.26 | 298 a ± 12.9 | 15.5 bc ± 1.32 | 9.12 bc ± 0.309 | nd |
| trans-Cinnamic acid | nd | 23.3 ± 2.91 | nd | nd | nd |
| Vanillic acid | 4.01 b ± 0.209 | 1690 a ± 131 | 57.1 b ± 2.42 | 77.9 b ± 4.05 | 145 b ± 1.5 |
| Total phenolic content [mg GAE g−1 DM] | 2.03 c ± 0.137 | 11.3 a ± 1.03 | 3.22 bc ± 0.300 | 2.22b c ± 0.203 | 3.42 b ± 0.321 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magdziak, Z.; Gąsecka, M.; Stuper-Szablewska, K.; Siwulski, M.; Budzyńska, S.; Jasińska, A.; Niedzielski, P.; Kalač, P.; Mleczek, M. A Possibility to Use Selected Crop Post-Extraction Wastes to Improve the Composition of Cultivated Mushroom Pleurotus citrinopileatus. J. Fungi 2021, 7, 894. https://doi.org/10.3390/jof7110894
Magdziak Z, Gąsecka M, Stuper-Szablewska K, Siwulski M, Budzyńska S, Jasińska A, Niedzielski P, Kalač P, Mleczek M. A Possibility to Use Selected Crop Post-Extraction Wastes to Improve the Composition of Cultivated Mushroom Pleurotus citrinopileatus. Journal of Fungi. 2021; 7(11):894. https://doi.org/10.3390/jof7110894
Chicago/Turabian StyleMagdziak, Zuzanna, Monika Gąsecka, Kinga Stuper-Szablewska, Marek Siwulski, Sylwia Budzyńska, Agnieszka Jasińska, Przemysław Niedzielski, Pavel Kalač, and Mirosław Mleczek. 2021. "A Possibility to Use Selected Crop Post-Extraction Wastes to Improve the Composition of Cultivated Mushroom Pleurotus citrinopileatus" Journal of Fungi 7, no. 11: 894. https://doi.org/10.3390/jof7110894
APA StyleMagdziak, Z., Gąsecka, M., Stuper-Szablewska, K., Siwulski, M., Budzyńska, S., Jasińska, A., Niedzielski, P., Kalač, P., & Mleczek, M. (2021). A Possibility to Use Selected Crop Post-Extraction Wastes to Improve the Composition of Cultivated Mushroom Pleurotus citrinopileatus. Journal of Fungi, 7(11), 894. https://doi.org/10.3390/jof7110894