
Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain)

 ,
,  , , ,
, , ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites and Proceedings
2.2. DNA Extraction and Sequencing
2.3. Sequencing Data and Statistical Analysis
3. Results
3.1. Water Physicochemical Data
3.2. Sequence Analysis and ASV Determination
3.3. Fungal Diversity in the Saltern
3.4. Fungal Taxonomic Assignment and Community Composition
3.5. Fungal Functional Guilds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hueso, K.; Carrasco, J.F. Las salinas de interior, un patrimonio desconocido y amenazado. Def. Patrim. Geológico Min. 2006, 6–7, 23–28. [Google Scholar]
- Murelaga, X.; Zuluaga, M.C.; Alonso, A.; Baceta, J.I.; Ortega, L.O. Guía Geológica del Diapiro de Valle Salado de Añana, 1st ed.; Diputación Foral de Álava: Vitoria-Gasteiz, Spain, 2019. [Google Scholar]
- Plata-Montero, A. El Ciclo Productivo de la Sal y las Salinas Reales a Mediados del Siglo XIX, 1st ed.; Diputación Foral de Álava: Vitoria-Gasteiz, Spain, 2006. [Google Scholar]
- Plata-Montero, A.; Landa, M. Pasado, presente y futuro del Valle Salado de Salinas de Añana (Álava, País Vasco). Rev. Int. Estud. Vascos. 2011, 56, 719–739. [Google Scholar]
- Gunde-Cimerman, N.; Polona, Z.; de Hoog, S.; Plemenitaš, A. Hypersaline waters in salterns-natural ecological niches for halophilic black yeasts. FEMS Microbiol. Ecol. 2000, 32, 235–240. [Google Scholar]
- Gostinčar, C.; Grube, M.; De Hoog, S.; Zalar, P.; Gunde-Cimerman, N. Extremotolerance in fungi: Evolution on the edge. FEMS Microbiol. Ecol. 2010, 71, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.; Kim, H.; Choi, H.S. Fungi in salterns. J. Microbiol. 2019, 57, 717–724. [Google Scholar] [CrossRef]
- Ventosa, A.; Fernández, A.B.; León, M.J.; Sánchez-Porro, C.; Rodriguez-Valera, F. The Santa Pola saltern as a model for studying the microbiota of hypersaline environments. Extremophiles 2014, 18, 811–824. [Google Scholar] [CrossRef]
- Çınar, S.; Mutlu, M.B. Comparative analysis of prokaryotic diversity in solar salterns in eastern Anatolia (Turkey). Extremophiles 2016, 20, 589–601. [Google Scholar] [CrossRef]
- Di Meglio, L.; Santos, F.; Gomariz, M.; Almansa, C.; López, C.; Antón, J.; Nercessian, D. Seasonal dynamics of extremely halophilic microbial communities in three Argentinian salterns. FEMS Microbiol. Ecol. 2016, 92, fiw184. [Google Scholar] [CrossRef] [Green Version]
- Couto-Rodríguez, R.L.; Montalvo-Rodríguez, R. Temporal analysis of the microbial community from the crystallizer ponds in Cabo Rojo, Puerto Rico, using metagenomics. Genes 2019, 10, 422. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Xu, Y.; Helfant, L. Geographical isolation, buried depth, and physicochemical traits drive the variation of species diversity and prokaryotic community in three typical hypersaline environments. Microorganisms 2020, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Filker, S.; Gimmler, A.; Dunthorn, M. Deep sequencing uncovers protistan plankton diversity in the Portuguese Ria Formosa solar saltern ponds. Extremophiles 2015, 19, 283–295. [Google Scholar] [CrossRef]
- Heo, Y.M.; Lee, H.; Kwon, S.L.; Park, M.Y.; Kang, J.E.; Kim, G.-H.; Kim, B.S.; Kim, J.-J. Fungal diversity in intertidal mudflats and abandoned solar salterns as a source for biological resources. Mar Drugs 2019, 17, 601. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.-L.; Long, Z.-J.; Ren, M.-X. Microbial community and functional prediction during the processing of salt production in a 1000-year-old marine solar saltern of South China. Sci. Total Environ. 2021, 28, 152014. [Google Scholar] [CrossRef]
- Duo Saito, R.A.; Connell, L.; Rodriguez, R.; Redman, R.; Libkind, D.; de Garcia, V. Metabarcoding analysis of the fungal biodiversity associated with Castaño Overa Glacier—Mount Tronador, Patagonia, Argentina. Fungal Ecol. 2018, 36, 8–16. [Google Scholar] [CrossRef]
- Hugoni, M.; Escalas, A.; Bernard, C.; Nicolas, S.; Jézéquel, D.; Vazzoler, F.; Sarazin, G.; Leboulander, C.; Bouvy, M.; Got, P.; et al. Spatiotemporal variations in microbial diversity across the three domains of life in a tropical thalassohaline lake (Dziani Dzaha, Mayotte Island). Mol. Ecol. 2018, 27, 4775–4786. [Google Scholar] [CrossRef]
- Li, P.D.; Jeewon, R.; Aruna, B.; Li, H.Y.; Lin, F.C.; Wang, H.K. Metabarcoding reveals differences in fungal communities between unflooded versus tidal flat soil in coastal saline ecosystem. Sci. Total Environ. 2019, 690, 911–922. [Google Scholar] [CrossRef]
- Yang, C.; Sun, J. Soil salinity drives the distribution patterns and ecological functions of Fungi in Saline-Alkali land in the Yellow River Delta, China. Front. Microbiol. 2020, 11, 594284. [Google Scholar] [CrossRef]
- Dutra-Silva, L.; Pereira, G.E.; Batista, L.R.; Matteoli, F.P. Fungal diversity and occurrence of mycotoxin producing fungi in tropical vineyards. World J. Microbiol. Biotechnol. 2021, 37, 112. [Google Scholar] [CrossRef]
- Sun, D.; Qu, J.; Huang, Y.; Lu, J.; Yin, L. Analysis of microbial community diversity of muscadine grape skins. Food Res. Int. 2021, 145, 110417. [Google Scholar] [CrossRef]
- Yang, S.; Xu, W.; Gao, Y.; Chen, X.; Luo, Z.H. Fungal diversity in deep-sea sediments from Magellan seamounts environment of the western Pacific revealed by high-throughput Illumina sequencing. J. Microbiol. 2020, 58, 841–852. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Yu, L.Y. Geographic distance and habitat type influence fungal communities in the Arctic and Antarctic sites. Microb. Ecol. 2021, 82, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Barbero, M.D.; Viver, T.; Zabaleta, A.; Senel, E.; Gomariz, M.; Antigüedad, I.; Santos, F.; Martínez-García, M.; Rosselló-Mora, R.; Antón, J. Ancient saltern metagenomics: Tracking changes in microbes and their viruses from the underground to the surface. Environ. Microbiol. 2021, 23, 3477–3498. [Google Scholar] [CrossRef] [PubMed]
- Illumina Inc. Fungal Sequencing and Classification with the ITS Metagenomics Protocol. Available online: https://www.illumina.com/content/dam/illumina-marketing/documents/products/appnotes/its-metagenomics-app-note-1270-2018-001-web.pdf (accessed on 21 October 2021).
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugan, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Abarenkov, K.; Zirk, A.; Piirmann, T.; Pöhönen, R.; Ivanov, F.; Nilsson, H.; Kõljalg, U. UNITE QIIME Release for Fungi; Version 10.05.2021; UNITE Community: Wales, UK, 2021. [Google Scholar]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Bunbury-Blanchette, A.L.; Walker, A.K. Occurrence and distribution of fungi in saline environments. In Microorganisms in Saline Environments: Strategies and Functions. Soil Biology, 1st ed.; Giri, B., Varma, A., Eds.; Springer: Cham, Switzerland, 2019; Volume 56, pp. 19–38. [Google Scholar]
- Grossart, H.-P.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 17, 254–339. [Google Scholar] [CrossRef] [Green Version]
- Bass, D.; Howe, A.; Brown, N.; Barton, H.; Demidova, M.; Michelle, H.; Li, L.; Sanders, H.; Watkinson, S.C.; Willcock, S. Yeast forms dominate fungal diversity in the deep oceans. Proc. Biol. Sci. 2017, 274, 3069–3077. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Liu, J.J.; Banerjee, S.; White, J.F.; Zhou, N.; Zhao, Z.Y.; Zhang, K.; Hu, M.F.; Kingsley, K.; Tian, C.Y. Not by salinity alone: How environmental factors shape fungal communities in saline soils. Soil Sci. Soc. Am. J. 2019, 83, 1387–1398. [Google Scholar] [CrossRef]
- Luo, Y.; Wei, X.; Yang, S.; Gao, Y.H.; Luo, Z.H. Fungal diversity in deep-sea sediments from the Magellan seamounts as revealed by a metabarcoding approach targeting the ITS2 regions. Mycology 2020, 11, 214–229. [Google Scholar] [CrossRef]
- Tang, X.; Yu, L.; Xu, W.; Zhang, X.; Xu, X.; Wang, Q.; Wei, S.; Qiu, Y. Fungal diversity of deep-sea sediments in Mid-Oceanic Ridge area of the East Pacific and the South Indian Oceans. Botanica Marina 2020, 63, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Alsammar, H.; Delneri, D. An update on the diversity, ecology and biogeography of the Saccharomyces genus. FEMS Yeast Res. 2020, 20, foaa013. [Google Scholar] [CrossRef] [Green Version]
- Blomberg, A. Metabolic surprises in Saccharomyces cerevisiae during adaptation to saline conditions: Questions, some answers and a model. FEMS Microbiol. Lett. 2000, 182, 1–8. [Google Scholar] [CrossRef]
- Naumov, G.I.; Naumova, E.S.; Martynenko, N.N.; Masneuf-Pomaréde, I. Taxonomy, ecology, and genetics of the yeast Saccharomyces bayanus: A new object for science and practice. Microbiology 2011, 80, 735–742. [Google Scholar] [CrossRef]
- Zalar, P.D.; De Hoog, G.S.; Schroers, H.J.; Crous, P.W.; Groenewald, J.Z.; Gunde-Cimerman, N. Phylogeny and ecology of the ubiquitous saprobe Cladosporium sphaerospermum, with descriptions of seven new species from hypersaline environments. Stud. Micol. 2007, 58, 157–183. [Google Scholar] [CrossRef]
- Zalar, P.; de Hood, G.S.; Gunde-Cimerman, N. Trimmatostroma salinum, a new species from hypersaline water. Stud. Mycol. 1999, 43, 57–62. [Google Scholar]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The genus Metschnikowia in enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Grangeteau, C.; Gerhards, D.; Rousseaux, S.; von Wallbrunn, C.; Alexandre, H.; Guilloux-Benatier, M. Diversity of yeast strains of the genus Hanseniaspora in the winery environment: What is their involvement in grape must fermentation? Food Microbiol. 2015, 50, 70–77. [Google Scholar] [CrossRef]
- Windholtz, S.; Dutilh, L.; Lucas, M.; Maupeu, J.; Vallet-Courbin, A.; Farris, L.; Coulon, J.; Masneuf-Pomarède, I. Population dynamics and yeast diversity in early winemaking stages without sulfites revealed by three complementary approaches. Appl. Sci. 2021, 11, 2494. [Google Scholar] [CrossRef]
- Jančič, S.; Zalar, P.; Kocev, D.; Schroers, H.J.; Džeroski, S.; Gunde-Cimerman, N. Halophily reloaded: New insights into the extremophilic life-style of Wallemia with the description of Wallemia hederae sp. nov. Fungal Divers. 2016, 76, 97–118. [Google Scholar] [CrossRef] [Green Version]
- Lambevska, A.; Rusevska, K.; Karadelev, M. New data on the taxonomy, distribution and ecology of the genus Peniophora Cooke (Basidiomycota, Fungi) in the Republic of Macedonia. Maced. J. Ecol. Environ. 2013, 15, 69–79. [Google Scholar]
- Glushakova, A.M.; Kachalkin, A.V. Endophytic yeasts in Malus domestica and Pyrus communis fruits under anthropogenic impact. Microbiology 2017, 86, 128–135. [Google Scholar] [CrossRef]
- DeLong-Duhon, S.G.; Bagley, R.K. Phylogeny, morphology, and ecology resurrect previously synonymized species of North American Stereum. bioRxiv 2020. [Google Scholar] [CrossRef]
- Vujanovic, V. Tremellomycetes yeasts in kernel ecological niche: Early indicators of enhanced competitiveness of endophytic and mycoparasitic symbionts against wheat pathobiota. Plants 2021, 10, 905. [Google Scholar] [CrossRef]
- Baldrian, P.; Vetrovský, T.; Lepinay, C.; Kohout, P. High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Divers. 2021. [Google Scholar] [CrossRef]
- Lücking, R.; Aima, M.C.; Robbertse, B.; Miller, A.N.; Aoki, T.; Ariyawansa, H.A.; Cardinali, G.; Crous, P.W.; Druzhinina, I.S.; Geiser, D.M.; et al. Fungal taxonomy and sequence-based nomenclature. Nat. Microbiol. 2021, 6, 540–548. [Google Scholar] [CrossRef]
- Grossart, H.-P.; Wurzbacher, C.; James, T.Y.; Kagami, M. Discovery of dark matter fungi in aquatic ecosystems demands a reappraisal of the phylogeny and ecology of zoosporic fungi. Fungal Ecol. 2016, 19, 28–38. [Google Scholar] [CrossRef] [Green Version]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. mBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andermann, T.; Torres Jiménez, M.F.; Matos-Maraví, P.; Batista, R.; Blanco-Pastor, J.L.; Gustafsson, A.L.S.; Kistler, L.; Liberal, I.M.; Oxelman, B.; Bacon, C.D.; et al. A guide to carrying out a phylogenomic target sequence capture project. Front. Genet. 2020, 10, 1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Liu, C.; Wang, J.; Zhang, L. Soil pH determines fungal diversity along an elevation gradient in Southwestern China. Sci. China Life Sci. 2018, 61, 718–726. [Google Scholar] [CrossRef]
- Mohamed, D.J.; Martiny, J.B. Patterns of fungal diversity and composition along a salinity gradient. ISME J. 2011, 5, 379–388. [Google Scholar] [CrossRef]
- Zhu, S.; Lei, Y.; Wang, C.; Wei, Y.; Wang, C.; Sun, Y. Patterns of yeast diversity distribution and its drivers in rhizosphere soil of Hami melon orchards in different regions of Xinjiang. BMC Microbiol. 2021, 21, 1–18. [Google Scholar] [CrossRef]
- Taguiam, J.D.; Evallo, E.; Balendres, M.A. Epicoccum species: Ubiquitous plant pathogens and effective biological control agents. Eur. J. Plant Pathol. 2021, 159, 713–725. [Google Scholar] [CrossRef]
- Zalar, P.; Kocuvan, M.A.; Plemenitas, A.; Gunde-Cimerman, N. Halophilic black yeast colonize wood immersed in hypersaline water. Botanica Marina 2005, 48, 323–326. [Google Scholar] [CrossRef]
- Raghukumar, S. The marine environment and the role of fungi. In Fungi in Coastal and Oceanic Marine Ecosystems; Springer: Cham, Switzerland, 2017; pp. 17–38. [Google Scholar]
- Kirschner, R. Cercosporella and Ramularia. Mycologia 2009, 101, 110–119. [Google Scholar] [CrossRef]
- Osman, M.E.; El-Shapy, A.A.E.; Meligry, D.A.; Ayid, M.M. Survey for fungal decaying archaeological wood and their enzymatic activity. Int. J. Conserv. Sci. 2014, 5, 295–308. [Google Scholar]
- Blancard, D. Diagnosis of parasitic and nonparasitic diseases. In Tomato Diseases, 2nd ed.; Blancard, D., Ed.; CRC Press: London, UK, 2013; pp. 35–411. [Google Scholar]
- Abdo, H.; Catacchio, C.R.; Ventura, M.; D’Addabbo, P.; Alexandre, H.; Guilloux-Bénatier, M.; Rousseaux, S. The establishment of a fungal consortium in a new winery. Sci. Rep. 2020, 10, 7962. [Google Scholar] [CrossRef]

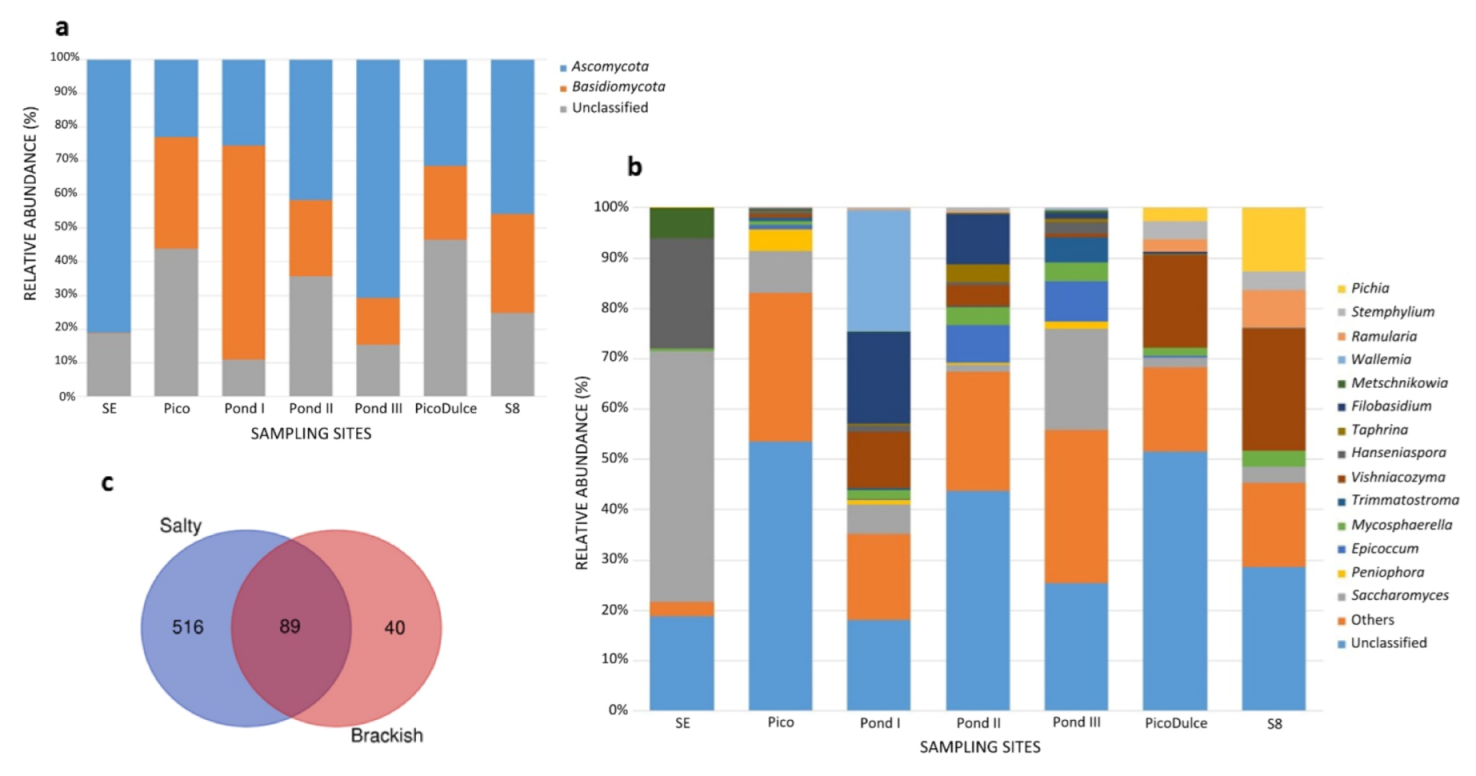

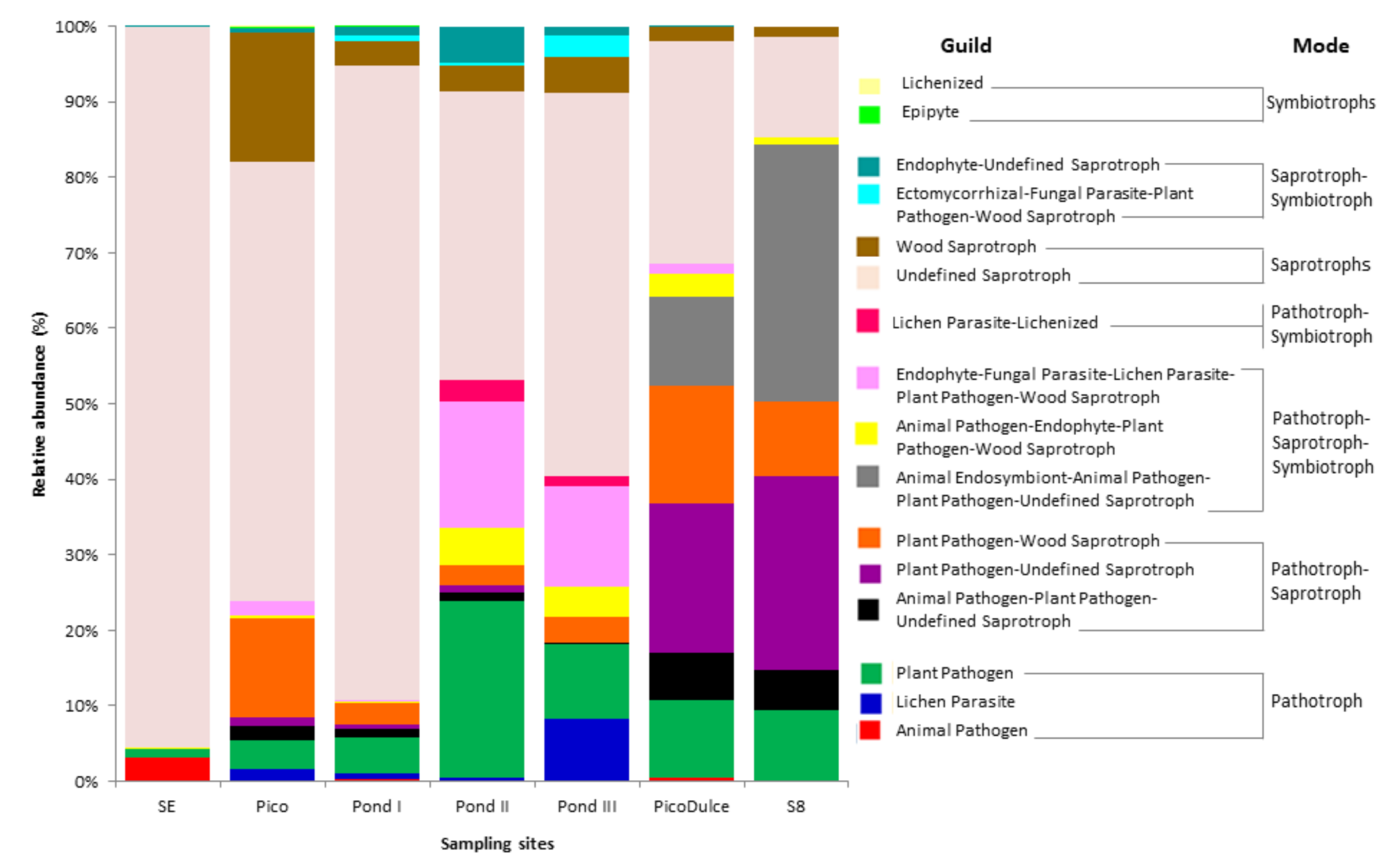

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Site | Observed ASVs | Chao1 | Shannon | Simpson | Pielou’s Evenness |
|---|---|---|---|---|---|
| Santa Engracia Spring | 71 | 116 | 3.59 | 0.77 | 0.58 |
| Pico Spring | 354 | 633 | 6.27 | 0.95 | 0.74 |
| Pond I | 264 | 452 | 5.44 | 0.93 | 0.67 |
| Pond II | 396 | 699 | 6.47 | 0.94 | 0.75 |
| Pond III | 263 | 337 | 6.07 | 0.94 | 0.75 |
| Pico Dulce Spring | 262 | 430 | 6.15 | 0.96 | 0.76 |
| S8 | 93 | 93 | 4.89 | 0.91 | 0.74 |
| Parameter | Pearson’s Coefficient Test (p Values) | |
|---|---|---|
| Shannon Index | Simpson Index | |
| NaCl | 0.896 | 0.732 |
| Temperature | 0.314 | 0.262 |
| pH | 0.029 * | 0.015 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azpiazu-Muniozguren, M.; Perez, A.; Rementeria, A.; Martinez-Malaxetxebarria, I.; Alonso, R.; Laorden, L.; Gamboa, J.; Bikandi, J.; Garaizar, J.; Martinez-Ballesteros, I. Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain). J. Fungi 2021, 7, 1074. https://doi.org/10.3390/jof7121074
Azpiazu-Muniozguren M, Perez A, Rementeria A, Martinez-Malaxetxebarria I, Alonso R, Laorden L, Gamboa J, Bikandi J, Garaizar J, Martinez-Ballesteros I. Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain). Journal of Fungi. 2021; 7(12):1074. https://doi.org/10.3390/jof7121074
Chicago/Turabian StyleAzpiazu-Muniozguren, Maia, Alba Perez, Aitor Rementeria, Irati Martinez-Malaxetxebarria, Rodrigo Alonso, Lorena Laorden, Javier Gamboa, Joseba Bikandi, Javier Garaizar, and Ilargi Martinez-Ballesteros. 2021. "Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain)" Journal of Fungi 7, no. 12: 1074. https://doi.org/10.3390/jof7121074
APA StyleAzpiazu-Muniozguren, M., Perez, A., Rementeria, A., Martinez-Malaxetxebarria, I., Alonso, R., Laorden, L., Gamboa, J., Bikandi, J., Garaizar, J., & Martinez-Ballesteros, I. (2021). Fungal Diversity and Composition of the Continental Solar Saltern in Añana Salt Valley (Spain). Journal of Fungi, 7(12), 1074. https://doi.org/10.3390/jof7121074