
Fullerol C60(OH)24 Nanoparticles and Drought Impact on Wheat (Triticum aestivum L.) during Growth and Infection with Aspergillus flavus

 ,
,  ,
,  ,
,  , and
, and
Abstract
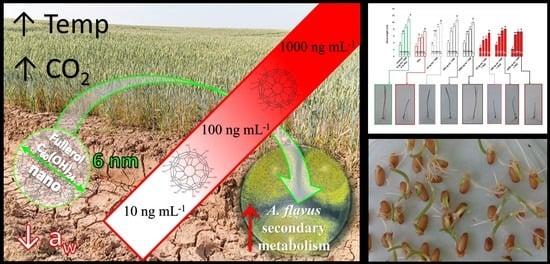
1. Introduction
2. Materials and Methods
3. Results
3.1. Fullerol C60(OH)24 Nanoparticles Characterization
3.2. Fullerol C60(OH)24 Nanoparticles and Drought Simulation Impact on Morphometric Parameters of the BC Tena Wheat Genotype
3.3. Antioxidative Enzymes Activity and TBARS Content in Shoot Extracts
3.4. Aspergillus Flavus Aggressiveness Test in the Presence of Fullerol C60(OH)24 Nanoparticles
3.5. Aspergillus Flavus NRRL 3251 Secondary Metabolite Profile after the Infection of Wheat Seeds Pre-Treated with Fullerol C60(OH)24 Nanoparticles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, J.; Alemany, L.B.; Li, W.; Benoit, D.; Fortner, J.D. Photoenhanced transformation of hydroxylated fullerene (fullerol) by free chlorine in water. Environ. Sci. Nano 2017, 4, 470–479. [Google Scholar] [CrossRef]
- Kovač, T.; Šarkanj, B.; Klapec, T.; Borišev, I.; Kovač, M.; Nevistić, A.; Strelec, I. Fullerol C60(OH)24 nanoparticles and mycotoxigenic fungi: A preliminary investigation into modulation of mycotoxin production. Environ. Sci. Pollut Res. 2017, 60, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kovač, T.; Šarkanj, B.; Klapec, T.; Borišev, I.; Kovač, M.; Nevistić, A.; Strelec, I. Antiaflatoxigenic effect of fullerene C60nanoparticles at environmentally plausible concentrations. AMB Express 2018, 8. [Google Scholar] [CrossRef]
- Kovač, T.; Borišev, I.; Crevar, B.; Čačić Kenjerić, F.; Kovač, M.; Strelec, I.; Ezekiel, C.N.; Sulyok, M.; Krska, R.; Šarkanj, B. Fullerol C60(OH)24 nanoparticles modulate aflatoxin B1 biosynthesis in Aspergillus flavus. Sci. Rep. 2018, 8, 12855. [Google Scholar] [CrossRef]
- Kovač, T.; Borišev, I.; Kovač, M.; Lončarić, A.; Čačić Kenjerić, F.; Djordjevic, A.; Strelec, I.; Ezekiel, C.N.; Sulyok, M.; Krska, R.; et al. Impact of fullerol C60(OH)24 nanoparticles on the production of emerging toxins by Aspergillus flavus. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Kovač, T.; Šarkanj, B.; Borišev, I.; Djordjevic, A.; Jović, D.; Lončarić, A.; Babić, J.; Jozinović, A.; Krska, T.; Gangl, J.; et al. Fullerol C60(OH)24 Nanoparticles Affect Secondary Metabolite Profile of Important Foodborne Mycotoxigenic Fungi In Vitro. Toxins 2020, 12, 213. [Google Scholar] [CrossRef]
- Wang, J.; Onasch, T.B.; Ge, X.; Collier, S.; Zhang, Q.; Sun, Y.; Yu, H.; Chen, M.; Prévôt, A.S.H.; Worsnop, D.R. Observation of Fullerene Soot in Eastern China. Environ. Sci. Technol. Lett. 2016, 3, 121–126. [Google Scholar] [CrossRef]
- Michalitsch, R.; Kallinger, C.; Verbandt, Y.; Veefkind, V.; Huebner, S.R. The fullerene patent landscape in Europe. Nanotechnol. Law Bus. 2008, 5, 85–94. [Google Scholar]
- Duncan, T.V. Applications of nanotechnology in food packaging and food safety: Barrier materials, antimicrobials and sensors. J. Colloid Interface Sci. 2011, 363, 1–24. [Google Scholar] [CrossRef]
- Pycke, B.F.G.; Chao, T.-C.; Herckes, P.; Westerhoff, P.; Halden, R.U. Beyond nC60: Strategies for identification of transformation products of fullerene oxidation in aquatic and biological samples. Anal. Bioanal. Chem. 2012, 404, 2583–2595. [Google Scholar] [CrossRef][Green Version]
- Dai, L. Synthesis of fullerene- and fullerol-containing polymers. J. Mater. Chem. 1998, 8, 325–330. [Google Scholar] [CrossRef]
- Zhou, A.H.; Zhang, J.D.; Xie, Q.J.; Yao, S.Z. Application of double-impedance system and cyclic voltammetry to study the adsorption of fullerols (C60(OH)n) on biological peptide-adsorbed gold electrode. Biomaterials 2001, 22, 2515–2524. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, H.; Ruan, L.; Chen, L.; Li, H.; Chang, X.L.; Zhang, X.; Yang, S.T. Bioaccumulation of 13C-fullerenol nanomaterials in wheat. Environ. Sci. Nano 2016, 3, 799–805. [Google Scholar] [CrossRef]
- Medina, A.; Akbar, A.; Baazeem, A.; Rodriguez, A.; Magan, N. Climate change, food security and mycotoxins: Do we know enough? Fungal Biol. Rev. 2017, 31, 143–154. [Google Scholar] [CrossRef]
- Bebber, D.P.; Ramotowski, M.A.T.; Gurr, S.J. Crop pests and pathogens move polewards in a warming world. Nat. Clim. Chang. 2013, 3, 985–988. [Google Scholar] [CrossRef]
- Helfer, S. Rust fungi and global change. New Phytol. 2014, 201, 770–780. [Google Scholar] [CrossRef]
- Trnka, M.; Rötter, R.P.; Ruiz-Ramos, M.; Kersebaum, K.C.; Olesen, J.E.; Žalud, Z.; Semenov, M.A. Adverse weather conditions for European wheat production will become more frequent with climate change. Nat. Clim. Chang. 2014, 4, 637–643. [Google Scholar] [CrossRef]
- Medina, Á.; Rodríguez, A.; Magan, N. Climate change and mycotoxigenic fungi: Impacts on mycotoxin production. Curr. Opin. Food Sci. 2015, 5, 99–104. [Google Scholar] [CrossRef]
- Fayez, K.A.; Bazaid, S.A. Improving drought and salinity tolerance in barley by application of salicylic acid and potassium nitrate. J. Saudi Soc. Agric. Sci. 2014, 13, 45–55. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New Phytol. 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Ladjal, M.; Epron, D.; Ducrey, M. Effects of drought preconditioning on thermotolerance of photosystem II and susceptibility of photosynthesis to heat stress in cedar seedlings. Tree Physiol. 2000, 20, 1235–1241. [Google Scholar] [CrossRef]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen species signaling in plants. Antioxid. Redox Signal. 2006, 8, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Zong, W.; Zhong, X.; You, J.; Xiong, L. Genome-wide profiling of histone H3K4-tri-methylation and gene expression in rice under drought stress. Plant Mol. Biol. 2013, 81, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Biosci, I.J.; Ahadzadeh, B.; Zaeifizadeh, M.; Shahbazi, H.; Ghasemi, M. Effect of terminal drought stress on the germination and growth potential wheat cultivars seeds. Int. J. Biosci. 2014, 6655, 134–146. [Google Scholar] [CrossRef]
- Mustafa, G.; Komatsu, S. Toxicity of heavy metals and metal-containing nanoparticles on plants. Biochim. Biophys. Acta Proteins Proteom. 2016, 1864, 932–944. [Google Scholar] [CrossRef]
- Cox, A.; Venkatachalam, P.; Sahi, S.; Sharma, N. Silver and titanium dioxide nanoparticle toxicity in plants: A review of current research. Plant Physiol. Biochem. 2016, 107, 147–163. [Google Scholar] [CrossRef]
- Borišev, M.; Borišev, I.; Župunski, M.; Arsenov, D.; Pajević, S.; Ćurćić, Ž.; Vasin, J.; Djordjevic, A. Drought impact is alleviated in sugar beets (Beta vulgaris L.) by foliar application of fullerenol nanoparticles. PLoS ONE 2016, 11, 1–20. [Google Scholar] [CrossRef]
- Xiong, J.L.; Li, J.; Wang, H.C.; Zhang, C.L.; Naeem, M.S. Fullerol improves seed germination, biomass accumulation, photosynthesis and antioxidant system in Brassica napus L. under water stress. Plant Physiol. Biochem. 2018, 129, 130–140. [Google Scholar] [CrossRef]
- Panova, G.G.; Ktitorova, I.N.; Skobeleva, O.V.; Sinjavina, N.G.; Charykov, N.A.; Semenov, K.N. Impact of polyhydroxy fullerene (fullerol or fullerenol) on growth and biophysical characteristics of barley seedlings in favourable and stressful conditions. Plant Growth Regul. 2016, 79, 309–317. [Google Scholar] [CrossRef]
- Avanasi, R.; Jackson, W.A.; Sherwin, B.; Mudge, J.F.; Anderson, T.A. C60 fullerene soil sorption, biodegradation, and plant uptake. Environ. Sci. Technol. 2014, 48, 2792–2797. [Google Scholar] [CrossRef]
- Kovač, M.; Šubarić, D.; Bulaić, M.; Kovač, T.; Šarkanj, B. Yesterday masked, today modified; what do mycotoxins bring next? Arch. Ind. Hyg. Toxicol. 2018, 69, 196–214. [Google Scholar] [CrossRef]
- Kovač, M.; Bulaić, M.; Jakovljević, J.; Nevistić, A.; Rot, T.; Kovač, T. Mycotoxins, Pesticide Residues, and Heavy Metals Analysis of Croatian Cereals. Microorganisms 2021, 9, 216. [Google Scholar] [CrossRef] [PubMed]
- Sulyok, M.; Stadler, D.; Steiner, D. Validation of an LC-MS/MS-based dilute-and-shoot approach for the quantification of >500 mycotoxins and other secondary metabolites in food crops: Challenges and solutions. Anal. Bioanal. Chem. 2020, 412, 2607–2620. [Google Scholar] [CrossRef]
- Mirkov, S.M.; Djordjevic, A.N.; Andric, N.L.; Andric, S.A.; Kostic, T.S.; Bogdanovic, G.M.; Vojinovic-Miloradov, M.B.; Kovacevic, R.Z. Nitric oxide-scavenging activity of polyhydroxylated fullerenol, C60(OH)24. Nitric Oxide 2004, 11, 201–207. [Google Scholar] [CrossRef]
- Gharoobi, B.; Ghorbani, M.; Nezhad, M.G. Effects of different levels of osmotic potential on germination percentage and germination rate of barley, corn and canola. Iran J. Plant Physiol. 1990, 2, 413–418. [Google Scholar]
- Lončarić, A.; Stuburić, M.; Kovač, M.; Nevistić, A.; Šarkanj, B.; Kovač, T.; Crevar, B. Disruption of Aspergillus Flavus Cells: A Bead Mill Homogenization Method. Food Health Dis. 2019, 8, 13–18. [Google Scholar]
- Siegel, B.Z.; Galston, A.W. The isoperoxidases of Pisum sativum. Plant Physiol. 1967, 42, 221–226. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods Enzymatic Analysis; Academic Press: Cambridge, MA, USA, 1974; pp. 673–684. [Google Scholar]
- Raymond, J.; Rakariyatham, N.; Azanza, J.L. Purification and some properties of polyphenoloxidase from sunflower seeds. Phytochemistry 1993, 34, 927–931. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Gospodaryov, D. V Catalases protect cellular proteins from oxidative modification in Saccharomyces cerevisiae. Cell Biol. Int. 2005, 29, 187–192. [Google Scholar] [CrossRef]
- Malachová, A.; Sulyok, M.; Beltrán, E.; Berthiller, F.; Krska, R. Optimization and validation of a quantitative liquid chromatography-tandem mass spectrometric method covering 295 bacterial and fungal metabolites including all regulated mycotoxins in four model food matrices. J. Chromatogr. A 2014, 1362, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Purahong, W.; Alkadri, D.; Nipoti, P.; Pisi, A.; Lemmens, M.; Prodi, A. Validation of a modified Petri-dish test to quantify aggressiveness of Fusarium graminearum in durum wheat. Eur. J. Plant Pathol. 2012, 132, 381–391. [Google Scholar] [CrossRef]
- Pituch, K.A.; Stevens, J.P. Applied Multivariate Statistics for the Social Sciences, 6th ed.; Taylor & Francis: New York, NY, USA, 2016; ISBN 9780415836661. [Google Scholar]
- Jović, D.S.; Seke, M.N.; Djordjevic, A.N.; Mrdanović, J.; Aleksić, L.D.; Bogdanović, G.M.; Pavić, A.B.; Plavec, J. Fullerenol nanoparticles as a new delivery system for doxorubicin. RSC Adv. 2016, 6, 38563–38578. [Google Scholar] [CrossRef]
- Vraneš, M.; Borišev, I.; Tot, A.; Armaković, S.; Armaković, S.; Jović, D.; Gadžurić, S.; Djordjevic, A. Self-assembling, reactivity and molecular dynamics of fullerenol nanoparticles. Phys. Chem. Chem. Phys. 2017, 19, 135–144. [Google Scholar] [CrossRef]
- Rauscher, H.; Roebben, G.; Roebben, G.; Sanfeliu, A.B.; Emons, H.; Gibson, N.; Koeber, R.; Linsinger, T.; Rasmussen, K.; Sintes, J.R.; et al. Towards a Review of the EC Recommendation for a Definition of the Term “Nanomaterial”, Part. 3: Scientific-Technical Evaluation of Options to Clarify the Definition and to Facilitate Its Implementation; Joint Research Centre of the European Commission, European Union: Brussels, Belgium, 2015. [Google Scholar]
- Khodakovskaya, M.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon nanotubes are able to penetrate plant seed coat and dramatically affect seed germination and plant growth. ACS Nano 2009, 3, 3221–3227. [Google Scholar] [CrossRef] [PubMed]
- Khodakovskaya, M.V.; Kim, B.S.; Kim, J.N.; Alimohammadi, M.; Dervishi, E.; Mustafa, T.; Cernigla, C.E. Carbon nanotubes as plant growth regulators: Effects on tomato growth, reproductive system, and soil microbial community. Small 2013, 9, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Burman, U.; Kumar, P. Plant Response to Engineered Nanoparticles. Nanomater. Plants Algae Microorg. 2018, 1, 103–118. [Google Scholar] [CrossRef]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Aslani, F.; Bagheri, S.; Muhd Julkapli, N.; Juraimi, A.S.; Hashemi, F.S.G.; Baghdadi, A. Effects of engineered nanomaterials on plants growth: An overview. Sci. World J. 2014, 2014, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Salah, S.M.; Yajing, G.; Dongdong, C.; Jie, L.; Aamir, N.; Qijuan, H.; Weimin, H.; Mingyu, N.; Jin, H. Seed priming with polyethylene glycol regulating the physiological and molecular mechanism in rice (Oryza sativa L.) under nano-ZnO stress. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Dursun, A.; Ekinci, M. Effects of different priming treatments and priming durations on germination percentage of parsley (Petroselinum crispum L.) seeds. Agric. Sci. 2010, 1, 17–23. [Google Scholar] [CrossRef][Green Version]
- Trpkovic, A.; Todorovic-Markovic, B.; Trajkovic, V. Toxicity of pristine versus functionalized fullerenes: Mechanisms of cell damage and the role of oxidative stress. Arch. Toxicol. 2012, 86, 1809–1827. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, X.; Zhao, Y.; Lin, J.; Shu, C.; Wang, C.; Fang, X. Fullerene-induced increase of glycosyl residue on living plant cell wall. Environ. Sci. Technol. 2013, 47, 7490–7498. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.Y.; Xiong, F.X.; Fan, Y.K.; Li, J.; Wang, H.Z.; Xing, G.M.; Yan, F.M.; Tai, F.J.; He, R. Facile and scalable fabrication engineering of fullerenol nanoparticles by improved alkaline-oxidation approach and its antioxidant potential in maize. J. Nanoparticle Res. 2016, 18, 338. [Google Scholar] [CrossRef]
 - diameter of the fullerol C60(OH)24 aqueous nanoparticles.
- diameter of the fullerol C60(OH)24 aqueous nanoparticles.
- diameter of the fullerol C60(OH)24 aqueous nanoparticles.
- diameter of the fullerol C60(OH)24 aqueous nanoparticles.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | APX | CAT | POD | PPO | TBARS [pmol/g] |
|---|---|---|---|---|---|
| (nkat/g DW) | |||||
| Control; 0 ng mL−1 FNP | 0.1 ± 0.1 bc | 152.9 ± 12.2 ab | 3.3 ± 0.4 d | 1.0 ± 0.0 a | 6.2 ± 2.1 c |
| PEG | 0.8 ± 0.1 bcd | 168.3 ± 27.5 a | 4.5 ± 0.9 c | 1.1 ± 0.0 a | 57.7 ± 6.6 a |
| 10 ng mL−1 FNP | 0.6 ± 0.08 de | 165.1 ± 24.4 a | 2.9 ± 0.0 d | 1.1 ± 0.0 a | 18.6 ± 3.5 c |
| 100 ng mL−1 FNP | 0.6 ± 0.1 cde | 110.1 ± 0.0 bc | 4.4 ± 0.7 c | 0.9 ± 0.1 a | 51.3 ± 1.0 ab |
| 1000 ng mL−1 FNP | 1.1 ± 0.0 b | 99.7 ± 13.7 cd | 6.1 ± 0.1 b | 1.2 ± 0.3 a | 44.6 ± 6.2 b |
| 10 ng mL−1 FNP+ PEG | 0.3 ± 0.0 e | 89.7 ± 9.3 cd | 6.2 ± 0.6 b | 1.1 ± 0.0 a | 56.8 ± 0.2 a |
| 100 ng mL−1 FNP+ PEG | 0.7 ± 0.0 bcd | 63.0 ± 9.4 de | 3.1 ± 0.3 d | 1.0 ± 0.1 a | 67.1 ± 1.0 a |
| 1000 ng mL−1 FNP+ PEG | 1.9 ± 0.3 a | 30.6 ± 6.1 e | 7.4 ± 0.2 a | 1.1 ± 0.0 a | 74.9 ± 12.2 a |
| Treatment | AUHPC | AUDPCStandard |
|---|---|---|
| Control, 0 ng mL−1 FNP + A. flavus | 243 ± 13.7 a | 0.45 ± 0.04 a |
| 10 ng mL−1 FNP + A. flavus | 212 ± 24.4 a | 0.54 ± 0.07 a |
| 100 ng mL−1 FNP + A. flavus | 260 ± 13.2 a | 0.40 ± 0.04 a |
| 1000 ng mL−1 FNP + A. flavus | 275 ± 11.7 a | 0.36 ± 0.03 a |
| Treatment |  3-Nitropropionic acid 3-Nitropropionic acid
|  Aflatoxin B1 Aflatoxin B1
|  Aflatoxin B2 Aflatoxin B2
|  Aflatoxicol Aflatoxicol
|  Kojic acid Kojic acid
|  O-Methylsterigmatocystin O-Methylsterigmatocystin
|
|---|---|---|---|---|---|---|
| (ng mL−1) | ||||||
| Control; 0 ng mL−1 FNP | 9.01 ± 0.13 cd | <LOD d | <LOD b | <LOD a | <LOD b | <LOD b |
| Control, 0 ng mL−1 FNP + A. flavus | 25.5 ± 0.09 c | 6.63 ± 0.06 A | 0.07 ± 0 a | <LOD a | 16.8 ± 0.32 b | 0.17 ± 0 a |
| 10 ng mL−1 FNP | 7.95 ± 0.30 cd | <LOD D | <LOD b | <LOD a | <LOD b | <LOD b |
| 100 ng mL−1 FNP | 13.9 ± 0.29 cd | <LOD D | <LOD b | <LOD a | <LOD b | <LOD b |
| 1000 ng mL−1 FNP | 1.33 ± 0.06 d | <LOD D | <LOD b | <LOD a | <LOD b | <LOD b |
| FNP 10 ng mL−1 FNP + A. flavus | 5.03 ± 0.22 cd | 4.94 ± 0.13 C | 0.01 ± 0 b | 0.37 ± 0.16 a | 30.6 ± 0.20 b | 0.11 ± 0.04 ab |
| FNP 100 ng mL−1 FNP + A. flavus | 79.6 ± 0.07 b | 5.83 ± 0.12 B | 0 ± 0 b | 0.35 ± 0.04 a | 1338 ± 189 a | 0.15 ± 0 a |
| FNP 1000 ng mL−1 FNP + A. flavus | 1776 ± 10.8 a | 6.77 ± 0.43 A | 0.02 ± 0.02 b | 0.19 ± 0.15 a | 51.7 ± 2.23 b | 0.11 ± 0.04 ab |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovač, T.; Marček, T.; Šarkanj, B.; Borišev, I.; Ižaković, M.; Jukić, K.; Lončarić, A.; Krska, T.; Sulyok, M.; Krska, R. Fullerol C60(OH)24 Nanoparticles and Drought Impact on Wheat (Triticum aestivum L.) during Growth and Infection with Aspergillus flavus. J. Fungi 2021, 7, 236. https://doi.org/10.3390/jof7030236
Kovač T, Marček T, Šarkanj B, Borišev I, Ižaković M, Jukić K, Lončarić A, Krska T, Sulyok M, Krska R. Fullerol C60(OH)24 Nanoparticles and Drought Impact on Wheat (Triticum aestivum L.) during Growth and Infection with Aspergillus flavus. Journal of Fungi. 2021; 7(3):236. https://doi.org/10.3390/jof7030236
Chicago/Turabian StyleKovač, Tihomir, Tihana Marček, Bojan Šarkanj, Ivana Borišev, Maja Ižaković, Katarina Jukić, Ante Lončarić, Tamara Krska, Michael Sulyok, and Rudolf Krska. 2021. "Fullerol C60(OH)24 Nanoparticles and Drought Impact on Wheat (Triticum aestivum L.) during Growth and Infection with Aspergillus flavus" Journal of Fungi 7, no. 3: 236. https://doi.org/10.3390/jof7030236
APA StyleKovač, T., Marček, T., Šarkanj, B., Borišev, I., Ižaković, M., Jukić, K., Lončarić, A., Krska, T., Sulyok, M., & Krska, R. (2021). Fullerol C60(OH)24 Nanoparticles and Drought Impact on Wheat (Triticum aestivum L.) during Growth and Infection with Aspergillus flavus. Journal of Fungi, 7(3), 236. https://doi.org/10.3390/jof7030236