
Ca2+ Signalling Differentially Regulates Germ-Tube Formation and Cell Fusion in Fusarium oxysporum

Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Conditions of F. oxysporum
2.2. Pharmacological Inhibition, Microscopy and Statistical Analysis
3. Results
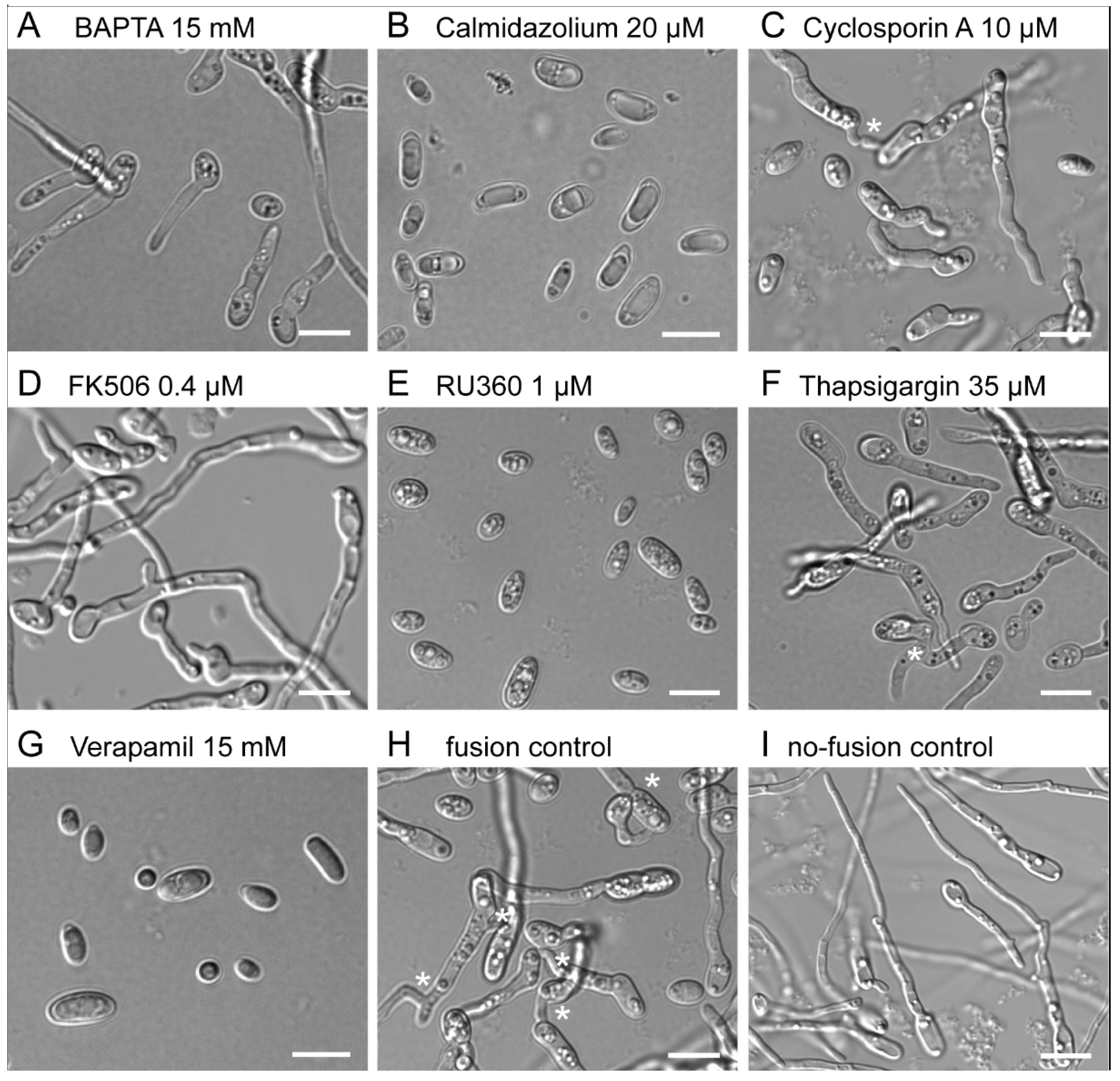
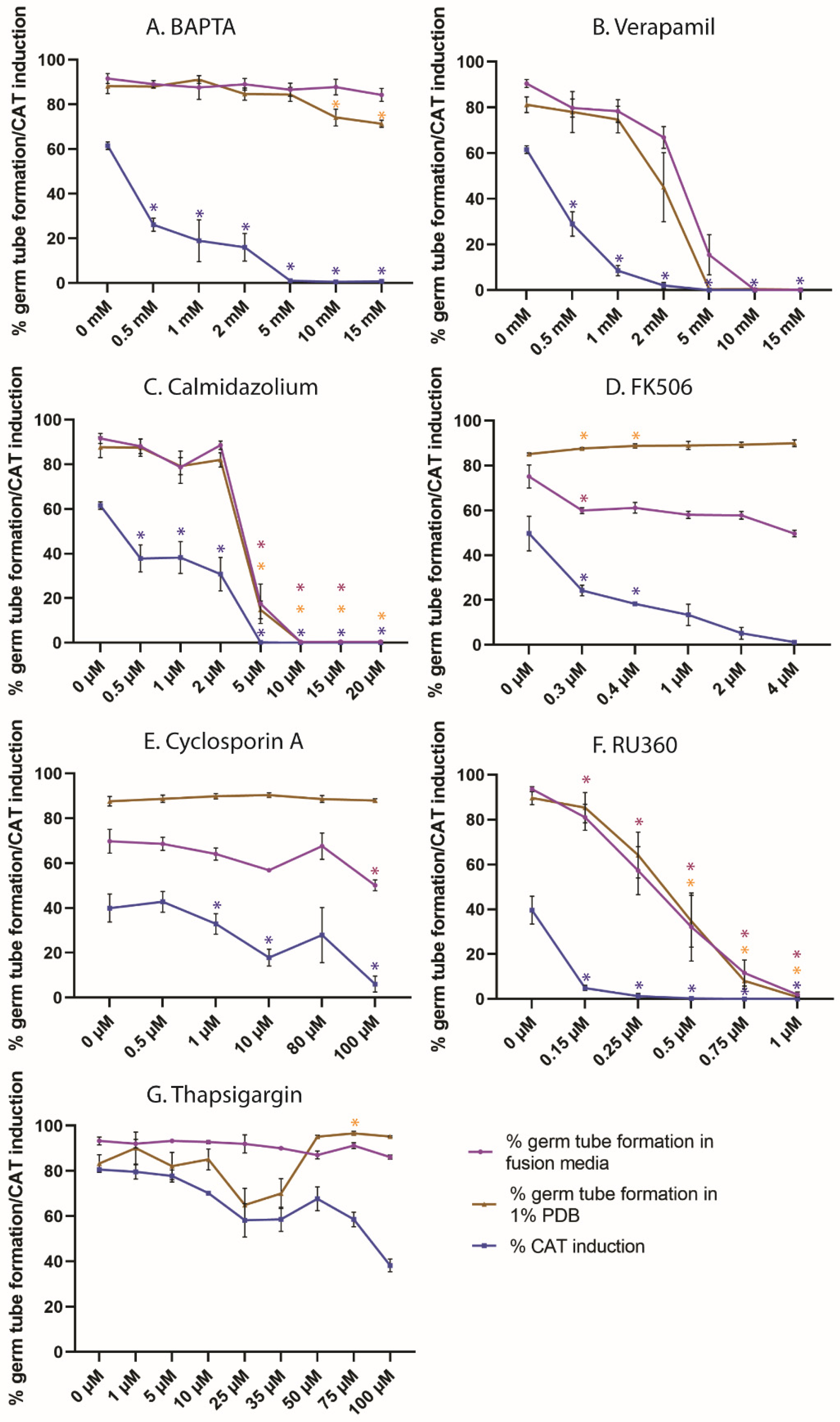
3.1. Extracellular Ca2+ Is Required for CAT Induction but Not Germ-Tube Formation
3.2. Calcium Entry/Exit through Calcium Channels Is Required for Germ-Tube Formation and CAT Induction
3.3. Calmodulin Inhibition by Calmidazolium Inhibits CAT Induction and Germ-Tube Formation
3.4. Inhibition of Calcineurin Function Inhibits CAT Induction but Not Germ-Tube Formation
3.5. Calcium Sequestration through the Mitochondrial Calcium Uniporter Is Required for CAT Induction and Germination
3.6. ER Calcium ATPase Inhibition by Thapsigargin Partially Inhibits CAT Induction but Not Germ-Tube Formation
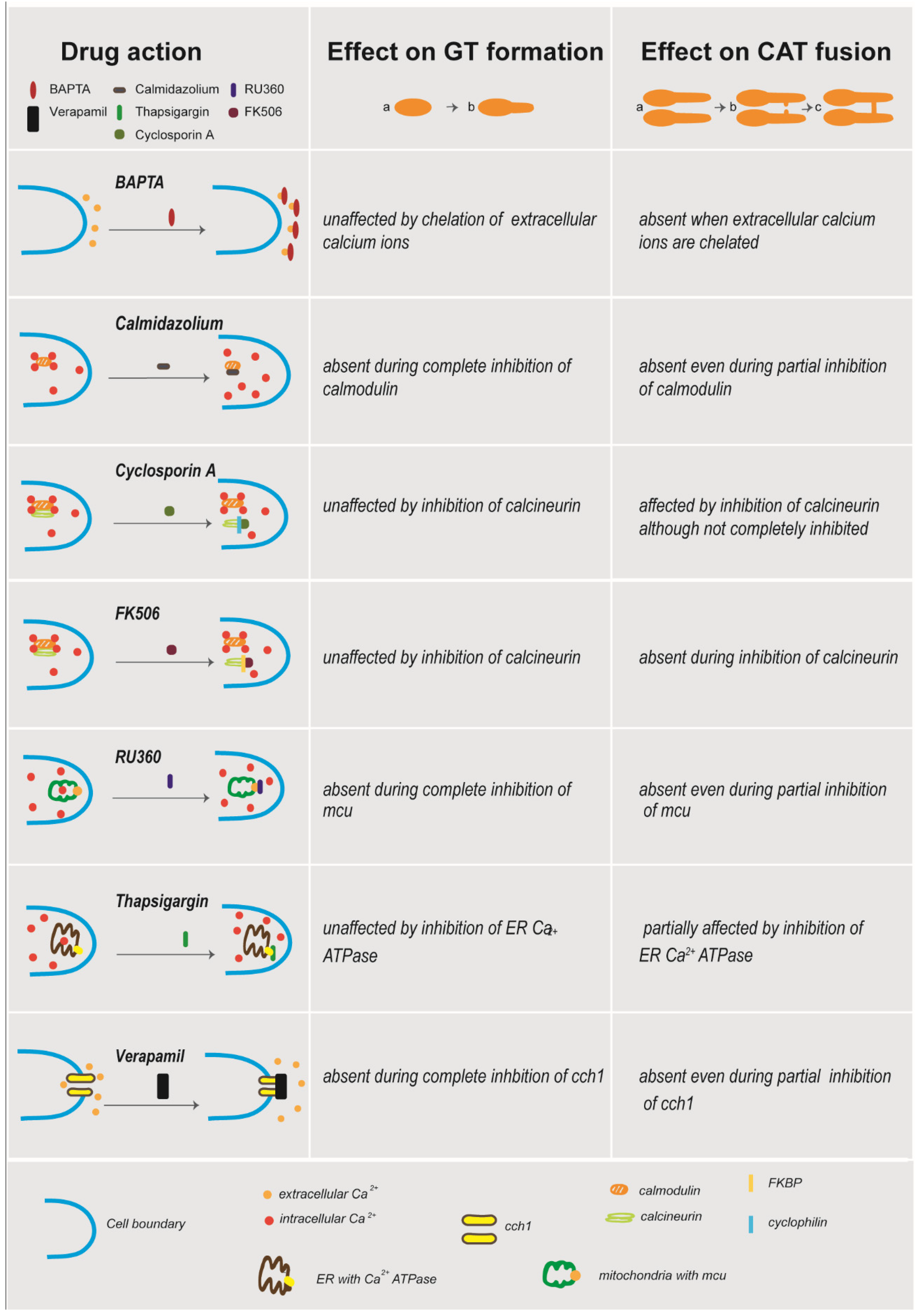
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takeshita, N.; Evangelinos, M.; Zhou, L.; Serizawa, T.; Somera-Fajardo, R.A.; Lu, L.; Takaya, N.; Nienhaus, G.U.; Fischer, R. Pulses of Ca2+ coordinate actin assembly and exocytosis for stepwise cell extension. Proc. Natl. Acad. Sci. USA 2017, 114, 5701–5706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Hou, Y.; Liu, W.; Lu, C.; Wang, W.; Sun, S. Components of the Calcium-Calcineurin Signaling Pathway in Fungal Cells and Their Potential as Antifungal Targets. Eukaryot. Cell 2015, 14, 324–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.-S.; Kim, J.-E.; Frailey, D.; Nohe, A.; Duncan, R.; Czymmek, K.J.; Kang, S. Roles of three Fusarium oxysporum calcium ion (Ca2+) channels in generating Ca2+ signatures and controlling growth. Fungal Genet. Biol. 2015, 82, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M.; Aguirre, J.; Bartnicki-García, S.; Braus, G.H.; Feldbrügge, M.; Fleig, U.; Hansberg, W.; Herrera-Estrella, A.; Kämper, J.; Kück, U.; et al. Fungal Morphogenesis, from the Polarized Growth of Hyphae to Complex Reproduction and Infection Structures. Microbiol. Mol. Biol. Rev. 2018, 82, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverman-Gavrila, L.B.; Lew, R.R. Calcium gradient dependence of Neurospora crassa hyphal growth. Microbiology 2003, 149, 2475–2485. [Google Scholar] [CrossRef] [PubMed]
- Harris, S.D. Hyphal branching in filamentous fungi. Dev. Biol. 2019, 451, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.B.; Kadotani, N.; Kasahara, S.; Tosa, Y.; Mayama, S.; Nakayashiki, H. Systematic functional analysis of calcium-signalling proteins in the genome of the rice-blast fungus, Magnaporthe oryzae, using a high-throughput RNA-silencing system. Mol. Microbiol. 2008, 68, 1348–1365. [Google Scholar] [CrossRef]
- Lange, M.; Peiter, E. Calcium Transport Proteins in Fungi: The Phylogenetic Diversity of Their Relevance for Growth, Virulence, and Stress Resistance. Front. Microbiol. 2020, 10, 3100. [Google Scholar] [CrossRef]
- Boyce, K.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Gutjahr, C.; Bleckmann, A.; Dresselhaus, T. Calcium Signaling during Reproduction and Biotrophic Fungal Interactions in Plants. Mol. Plant 2015, 8, 595–611. [Google Scholar] [CrossRef] [Green Version]
- Okungbowa, F.I.; Shittu, H.O. Fusarium wilts: An overview. Environ. Res. J. 2012, 6, 83–102. [Google Scholar]
- Read, N.D.; Goryachev, A.B.; Lichius, A. The mechanistic basis of self-fusion between conidial anastomosis tubes during fungal colony initiation. Fungal Biol. Rev. 2012, 26, 1–11. [Google Scholar] [CrossRef]
- Kurian, S.M.; Di Pietro, A.; Read, N.D. Live-cell imaging of conidial anastomosis tube fusion during colony initiation in Fusarium oxysporum. PLoS ONE 2018, 13, e0195634. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, R.; Bahkali, A.H.; Abd-Elsalam, K.A.; Moslem, M.; Ben M’Barek, S.; Gohari, A.M.; Jashni, M.K.; Stergiopoulos, I.; Kema, G.H.; de Wit, P.J. Horizontal gene and chromosome transfer in plant pathogenic fungi affecting host range. FEMS Microbiol. Rev. 2011, 35, 542–554. [Google Scholar] [CrossRef] [Green Version]
- Daskalov, A.; Heller, J.; Herzog, S.; Fleißner, A.; Glass, N.L. Molecular Mechanisms Regulating Cell Fusion and Heterokaryon Formation in Filamentous Fungi. Microbiol. Spectr. 2017, 5, 215–229. [Google Scholar] [CrossRef] [Green Version]
- Goryachev, A.B.; Lichius, A.; Wright, G.D.; Read, N.D. Excitable behavior can explain the “ping-pong” mode of communication between cells using the same chemoattractant. BioEssays 2012, 34, 259–266. [Google Scholar] [CrossRef]
- Herzog, S.; Schumann, M.R.; Fleißner, A. Cell fusion in Neurospora crassa. Curr. Opin. Microbiol. 2015, 28, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.C.; Kim, H.; Mackin, N.A.; Maldonado-Báez, L.; Evangelista, C.C.; Beaudry, V.G.; Dudgeon, D.D.; Naiman, D.Q.; Erdman, S.E.; Cunningham, K.W. New Regulators of a High Affinity Ca2+ Influx System Revealed through a Genome-wide Screen in Yeast. J. Biol. Chem. 2011, 286, 10744–10754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, E.M.; Locke, E.G.; Cunningham, K.W. Differential Regulation of Two Ca2+ Influx Systems by Pheromone Signaling in Saccharomyces cerevisiae. Genetics 2001, 159, 1527–1538. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; Schnell, N.; Chattaway, J.; Davies, P.; Dixon, G.; Sanders, D. The Saccharomyces cerevisiae CCH1 gene is involved in calcium influx and mating. FEBS Lett. 1997, 419, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Paidhungat, M.; Garrett, S. A homolog of mammalian, voltage-gated calcium channels mediates yeast pheromone-stimulated Ca2+ uptake and exacerbates the cdc1(Ts) growth defect. Mol. Cell. Biol. 1997, 17, 6339–6347. [Google Scholar] [CrossRef] [Green Version]
- Muller, E.M.; Mackin, N.A.; Erdman, S.E.; Cunningham, K.W. Fig1p Facilitates Ca2+ Influx and Cell Fusion during Mating of Saccharomyces cerevisiae. J. Biol. Chem. 2003, 278, 38461–38469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavinder, B.; Trail, F. Role of Fig1, a Component of the Low-Affinity Calcium Uptake System, in Growth and Sexual Development of Filamentous Fungi. Eukaryot. Cell 2012, 11, 978–988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddish, F.N.; Miller, C.L.; Gorkhali, R.; Yang, J.J. Monitoring ER/SR Calcium Release with the Targeted Ca2+ Sensor Catch ER+. J. Vis. Exp. 2017, 123, e55822. [Google Scholar] [CrossRef]
- Nelson, G.; Kozlova-Zwinderman, O.; Collis, A.J.; Knight, M.R.; Fincham, J.R.S.; Stanger, C.P.; Renwick, A.; Hessing, J.G.M.; Punt, P.J.; Van Den Hondel, C.A.M.J.J.; et al. Calcium measurement in living filamentous fungi expressing codon-optimized aequorin. Mol. Microbiol. 2004, 52, 1437–1450. [Google Scholar] [CrossRef]
- Gonçalves, A.P.; Cordeiro, J.M.; Monteiro, J.; Muñoz, A.; Correia-De-Sá, P.; Read, N.; Videira, A. Activation of a TRP-like channel and intracellular calcium dynamics during phospholipase C-mediated cell death. J. Cell Sci. 2014, 127, 3817–3829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, U.; Bencina, M.; Eigentler, A.; Meyer, V.; Marx, F. The Aspergillus giganteus antifungal protein AFPNN5353activates the cell wall integrity pathway and perturbs calcium homeostasis. BMC Microbiol. 2011, 11, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popa, C.-V.; Dumitru, I.; Ruta, L.L.; Danet, A.F.; Farcasanu, I.C. Exogenous oxidative stress induces Ca2+ release in the yeast Saccharomyces cerevisiae. FEBS J. 2010, 277, 4027–4038. [Google Scholar] [CrossRef]
- Teng, J.; Goto, R.; Iida, K.; Kojima, I.; Iida, H. Ion-channel blocker sensitivity of voltage-gated calcium-channel homologue Cch1 in Saccharomyces cerevisiae. Microbiology 2008, 154, 3775–3781. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Ding, X.; Zhang, B.; Xu, N.; Jia, C.; Mao, J.; Zhang, B.; Xing, L.; Li, M. Inhibitory effect of verapamil on Candida albicans hyphal development, adhesion and gastrointestinal colonization. FEMS Yeast Res. 2014, 14, 633–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunagawa, M.; Kosugi, T.; Nakamura, M.; Sperelakis, N. Pharmacological Actions of Calmidazolium, a Calmodulin Antagonist, in Cardiovascular System. Cardiovasc. Drug Rev. 2006, 18, 211–221. [Google Scholar] [CrossRef]
- Hooks, M.A. Tacrolimus, a New Immunosuppressant—A Review of the Literature. Ann. Pharmacother. 1994, 28, 501–511. [Google Scholar] [CrossRef] [PubMed]
- Lamoth, F.; Alexander, B.D.; Juvvadi, P.R.; Steinbach, W.J. Antifungal activity of compounds targeting the Hsp90-calcineurin pathway against various mould species: Table 1. J. Antimicrob. Chemother. 2015, 70, 1408–1411. [Google Scholar] [CrossRef] [Green Version]
- Juvvadi, P.R.; Lamoth, F.; Steinbach, W.J. Calcineurin as a multifunctional regulator: Unraveling novel functions in fungal stress responses, hyphal growth, drug resistance, and pathogenesis. Fungal Biol. Rev. 2014, 28, 56–69. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Handschumacher, R. Specific interaction of the cyclophilin-cyclosporin complex with the B subunit of calcineurin. J. Biol. Chem. 1993, 268, 14040–14044. [Google Scholar] [CrossRef]
- Amor, K.T.; Ryan, C.; Menter, A. The use of cyclosporine in dermatology: Part I. J. Am. Acad. Dermatol. 2010, 63, 925–946. [Google Scholar] [CrossRef]
- Hong, M.-P.; Vu, K.; Bautos, J.; Gelli, A. Cch1 Restores Intracellular Ca2+ in Fungal Cells during Endoplasmic Reticulum Stress. J. Biol. Chem. 2010, 285, 10951–10958. [Google Scholar] [CrossRef] [Green Version]
- Brini, M.; Carafoli, E. Calcium Pumps in Health and Disease. Physiol. Rev. 2009, 89, 1341–1378. [Google Scholar] [CrossRef] [Green Version]
- Garg, V.; Kirichok, Y. Keeping a lid on calcium uptake. eLife 2016, 5, 341. [Google Scholar] [CrossRef] [Green Version]
- Baughman, J.M.; Perocchi, F.; Girgis, H.S.; Plovanich, M.; Belcher-Timme, C.A.; Sancak, Y.; Bao, X.R.; Strittmatter, L.; Goldberger, O.; Bogorad, R.L.; et al. Integrative genomics identifies MCU as an essential component of the mitochondrial calcium uniporter. Nature 2011, 476, 341–345. [Google Scholar] [CrossRef] [Green Version]
- De Stefani, D.; Raffaello, A.; Teardo, E.; Szabò, I.; Rizzuto, R. A forty-kilodalton protein of the inner membrane is the mitochondrial calcium uniporter. Nature 2011, 476, 336–340. [Google Scholar] [CrossRef]
- Zelter, A.; Bencina, M.; Bowman, B.J.; Yarden, O.; Read, N.D. A comparative genomic analysis of the calcium signaling machinery in Neurospora crassa, Magnaporthe grisea, and Saccharomyces cerevisiae. Fungal Genet. Biol. 2004, 41, 827–841. [Google Scholar] [CrossRef]
- Benčina, M.; Bagar, T.; Lah, L.; Kraševec, N. A comparative genomic analysis of calcium and proton signaling/homeostasis in Aspergillus species. Fungal Genet. Biol. 2009, 46, S93–S104. [Google Scholar] [CrossRef] [PubMed]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium–calcineurin signalling components in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef]
- Galagan, J.E.; Calvo, S.E.; Borkovich, K.A.; Selker, E.U.; Read, N.D.; Jaffe, D.; Fitzhugh, W.; Ma, L.-J.; Smirnov, S.; Purcell, S.; et al. The genome sequence of the filamentous fungus Neurospora crassa. Nature 2003, 422, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Prole, D.L.; Taylor, C.W. Identification and Analysis of Cation Channel Homologues in Human Pathogenic Fungi. PLoS ONE 2012, 7, e42404. [Google Scholar] [CrossRef]
- Gonçalves, A.P.; Cordeiro, J.M.; Monteiro, J.; Lucchi, C.; Correia-De-Sá, P.; Videira, A. Involvement of mitochondrial proteins in calcium signaling and cell death induced by staurosporine in Neurospora crassa. Biochim. et Biophys. Acta (BBA)-Bioenerg. 2015, 1847, 1064–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploetz, R.C. Fusarium Wilt of Banana. Phytopathology 2015, 105, 1512–1521. [Google Scholar] [CrossRef] [Green Version]
- Nucci, F.; Nouér, S.A.; Capone, D.; Anaissie, E.; Nucci, M. Fusariosis. Semin. Respir. Crit. Care Med. 2015, 36, 706–714. [Google Scholar] [CrossRef]
- Esnakula, A.K.; Summers, I.; Naab, T.J. Fatal Disseminated Fusarium Infection in a Human Immunodeficiency Virus Positive Patient. Case Rep. Infect. Dis. 2013, 2013, 379320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, Y.-H.; Hsu, L.-H.; Wang, H.-F.; Lai, Y.-H.; Chen, Y.-L. Calcineurin Regulates Conidiation, Chlamydospore Formation and Virulence in Fusarium oxysporum f. sp. lycopersici. Front. Microbiol. 2020, 11, 539702. [Google Scholar] [CrossRef] [PubMed]
- Harren, K.; Tudzynski, B. Cch1 and Mid1 Are Functionally Required for Vegetative Growth under Low-Calcium Conditions in the Phytopathogenic Ascomycete Botrytis cinerea. Eukaryot. Cell 2013, 12, 712–724. [Google Scholar] [CrossRef]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef]
- Jin, L.; Harrison, S.C. Crystal structure of human calcineurin complexed with cyclosporin A and human cyclophilin. Proc. Natl. Acad. Sci. USA 2002, 99, 13522–13526. [Google Scholar] [CrossRef] [Green Version]
- Osherov, N.; May, G.S. The molecular mechanisms of conidial germination. FEMS Microbiol. Lett. 2001, 199, 153–160. [Google Scholar] [CrossRef]
- Lee, S.C.; Lee, Y.H. Calcium/calmodulin-dependent signaling for appressorium formation in the plant pathogenic fungus Magnaporthe grisea. Mol. Cells 1998, 8, 698–704. [Google Scholar]
- Som, T.; Kolaparthi, V.S. Developmental decisions in Aspergillus nidulans are modulated by Ras activity. Mol. Cell. Biol. 1994, 14, 5333–5348. [Google Scholar] [CrossRef]
- Palma-Guerrero, J.; Hall, C.R.; Kowbel, D.; Welch, J.; Taylor, J.W.; Brem, R.B.; Glass, N.L. Genome Wide Association Identifies Novel Loci Involved in Fungal Communication. PLoS Genet. 2013, 9, e1003669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichert, M.; Fleißner, A. Anastomosis and Heterokaryon Formation. In Genetic Transformation Systems in Fungi; van den Berg, M.A., Maruthachalam, K., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 2, pp. 3–21. [Google Scholar]
- Schumann, M.R.; Brandt, U.; Adis, C.; Hartung, L.; Fleißner, A. Plasma Membrane Integrity During Cell–Cell Fusion and in Response to Pore-Forming Drugs Is Promoted by the Penta-EF-Hand Protein PEF1 in Neurospora crassa. Genetics 2019, 213, 195–211. [Google Scholar] [CrossRef]
- Lichius, A.; Goryachev, A.; Fricker, M.; Obara, B.; Castro-Longoria, E.; Read, N.D. CDC-42 and RAC-1 regulate opposite chemotropisms in Neurospora crassa. J. Cell Sci. 2014, 127, 1953–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichius, A.; Roca, G.M.; Read, N.D. How to Distinguish Conidial Anastomosis Tubes (CATs) from Germ Tubes, and to Discriminate between Cell Fusion Mutants Blocked in CAT Formation and Homing. The Neurospora Protocol Guide. 2010. Available online: http://www.fgsc.net/neurosporaprotocols/LICHIUS_ROCA%20_READ_2010_CAT_formation_protocol_final.pdf (accessed on 27 November 2021).
- Courchesne, W.E. Characterization of a Novel, Broad-Based Fungicidal Activity for the Antiarrhythmic Drug Amiodarone. J. Pharmacol. Exp. Ther. 2002, 300, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Lew, R.R.; Abbas, Z.; Anderca, M.I.; Free, S.J. Phenotype of a Mechanosensitive Channel Mutant, mid-1, in a Filamentous Fungus, Neurospora crassa. Eukaryot. Cell 2008, 7, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavinder, B.; Hamam, A.; Lew, R.R.; Trail, F. Mid1, a Mechanosensitive Calcium Ion Channel, Affects Growth, Development, and Ascospore Discharge in the Filamentous Fungus Gibberella zeae. Eukaryot. Cell 2011, 10, 832–841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troppens, D.M.; Chu, M.; Holcombe, L.J.; Gleeson, O.; O’Gara, F.; Read, N.; Morrissey, J.P. The bacterial secondary metabolite 2,4-diacetylphloroglucinol impairs mitochondrial function and affects calcium homeostasis in Neurospora crassa. Fungal Genet. Biol. 2013, 56, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, M.; Cunningham, K.W. Calcium Release and Influx in Yeast: TRPC and VGCC Rule Another Kingdom. Sci. Signal. 2002, 2002, pe17. [Google Scholar] [CrossRef]
- Kim, H.-S.; Kim, J.-E.; Son, H.; Frailey, D.; Cirino, R.; Lee, Y.-W.; Duncan, R.; Czymmek, K.; Kang, S. Roles of three Fusarium graminearum membrane Ca2+ channels in the formation of Ca2+ signatures, growth, development, pathogenicity and mycotoxin production. Fungal Genet. Biol. 2018, 111, 30–46. [Google Scholar] [CrossRef]
- Brand, A.C.; Morrison, E.; Milne, S.; Gonia, S.; Gale, C.A.; Gow, N. Cdc42 GTPase dynamics control directional growth responses. Proc. Natl. Acad. Sci. USA 2014, 111, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Brand, A.; Shanks, S.; Duncan, V.M.; Yang, M.; Mackenzie, K.; Gow, N.A. Hyphal Orientation of Candida albicans Is Regulated by a Calcium-Dependent Mechanism. Curr. Biol. 2007, 17, 347–352. [Google Scholar] [CrossRef] [Green Version]
- Simonin, A.R.; Rasmussen, C.G.; Yang, M.; Glass, N.L. Genes encoding a striatin-like protein (ham-3) and a forkhead associated protein (ham-4) are required for hyphal fusion in Neurospora crassa. Fungal Genet. Biol. 2010, 47, 855–868. [Google Scholar] [CrossRef]
- Fu, C.; Iyer, P.; Herkal, A.; Abdullah, J.; Stout, A.; Free, S.J. Identification and Characterization of Genes Required for Cell-to-Cell Fusion in Neurospora crassa. Eukaryot. Cell 2011, 10, 1100–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, C.; Ao, J.; Dettmann, A.; Seiler, S.; Free, S.J. Characterization of the Neurospora crassa Cell Fusion Proteins, HAM-6, HAM-7, HAM-8, HAM-9, HAM-10, AMPH-1 and WHI-2. PLoS ONE 2014, 9, e107773. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, M.; Cunningham, K.W. Mitogen-activated Protein Kinase Stimulation of Ca2+ Signaling Is Required for Survival of Endoplasmic Reticulum Stress in Yeast. Mol. Biol. Cell 2003, 14, 4296–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Silao, F.G.S.; Bigol, U.G.; Bungay, A.A.C.; Nicolas, M.G.; Heitman, J.; Chen, Y.-L. Calcineurin Is Required for Pseudohyphal Growth, Virulence, and Drug Resistance in Candida lusitaniae. PLoS ONE 2012, 7, e44192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blankenship, J.; Heitman, J. Calcineurin Is Required for Candida albicans To Survive Calcium Stress in Serum. Infect. Immun. 2005, 73, 5767–5774. [Google Scholar] [CrossRef] [Green Version]
- Eckstein, L.A.; Van Quill, K.R.; Bui, S.K.; Uusitalo, M.S.; O’Brien, J.M. Cyclosporin A Inhibits Calcineurin/Nuclear Factor of Activated T-Cells Signaling and Induces Apoptosis in Retinoblastoma Cells. Investig. Opthalmology Vis. Sci. 2005, 46, 782–790. [Google Scholar] [CrossRef] [Green Version]
- Fleissner, A.; Leeder, A.C.; Roca, M.G.; Read, N.; Glass, N.L. Oscillatory recruitment of signaling proteins to cell tips promotes coordinated behavior during cell fusion. Proc. Natl. Acad. Sci. USA 2009, 106, 19387–19392. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Solvent | Source | Concentration |
|---|---|---|---|
| BAPTA | Water | Invitrogen Life Technologies | 0–15 mM |
| Calmidazolium | DMSO | Acros Organics | 0–20 µM |
| Cyclosporin A | DMSO | InvivoGen | 0–100 µM |
| FK506 | DMSO | InvivoGen | 0–20 µM |
| RU360 | Water | Calbiochem | 0–1 µM |
| Thapsigargin | DMSO | Sigma Aldrich | 0–100 µM |
| Verapamil | Water | Sigma Aldrich | 0–15 mM |
| Pharmacological Agent | Mode of Action in Mammalian Cells | MIC for Germ-Tube Formation | MIC for CAT Induction | |
|---|---|---|---|---|
| 1% PDB | Fusion Media | |||
| BAPTA | extracellular Ca2+ ion chelation | no inhibition | no inhibition | 5 mM |
| Calmidazolium | inhibits calmodulin | 10 µM | 10 µM | 5 µM |
| Cyclosporin A | inhibits calcineurin | no inhibition | no inhibition | >100 µM |
| FK506 | inhibits calcineurin | no inhibition | no inhibition | 4 µM |
| RU360 | inihibits mitochondrial Ca2+ uniporter | 0.75 µM | 1 µM | 0.15 µM |
| Thapsigargin | inhibits ER Ca2+ ion sequestration | no inhibition | no inhibition | no full inhibition |
| Verapamil | inhibits Cch1 | 5 mM | 10 mM | 2 mM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurian, S.M.; Lichius, A.; Read, N.D. Ca2+ Signalling Differentially Regulates Germ-Tube Formation and Cell Fusion in Fusarium oxysporum. J. Fungi 2022, 8, 90. https://doi.org/10.3390/jof8010090
Kurian SM, Lichius A, Read ND. Ca2+ Signalling Differentially Regulates Germ-Tube Formation and Cell Fusion in Fusarium oxysporum. Journal of Fungi. 2022; 8(1):90. https://doi.org/10.3390/jof8010090
Chicago/Turabian StyleKurian, Smija M., Alexander Lichius, and Nick D. Read. 2022. "Ca2+ Signalling Differentially Regulates Germ-Tube Formation and Cell Fusion in Fusarium oxysporum" Journal of Fungi 8, no. 1: 90. https://doi.org/10.3390/jof8010090
APA StyleKurian, S. M., Lichius, A., & Read, N. D. (2022). Ca2+ Signalling Differentially Regulates Germ-Tube Formation and Cell Fusion in Fusarium oxysporum. Journal of Fungi, 8(1), 90. https://doi.org/10.3390/jof8010090