Abstract
Conidium is the main infection unit and reproductive unit of pathogenic fungi. Exploring the mechanism of conidiation and its regulation contributes to understanding the pathogenicity of pathogenic fungi. Vib-1, a transcription factor, was reported to participate in the conidiation process. However, the regulation mechanism of Vib-1 in conidiation is still unclear. In this study, we analyzed the function of Vib-1 and its regulation mechanism in conidiation through knocking out and overexpression of Vib-1 in entomopathogenic fungus Metarhizium acridum. Results showed that the colonial growth of Mavib-1 disruption mutant (ΔMavib-1) was significantly decreased, and conidiation was earlier compared to wild type (WT), while overexpression of Mavib-1 led to a delayed conidiation especially when carbon or nitrogen sources were insufficient. Overexpression of Mavib-1 resulted in a conidiation pattern shift from microcycle conidiation to normal conidiation on nutrient-limited medium. These results indicated that Mavib-1 acted as a positive regulator in vegetative growth and a negative regulator in conidiation by affecting utilization of carbon and nitrogen sources in M. acridum. Transcription profile analysis demonstrated that many genes related to carbon and nitrogen source metabolisms were differentially expressed in ΔMavib-1 and OE strains compared to WT. Moreover, Mavib-1 affects the conidial germination, tolerance to UV-B and heat stresses, cell wall integrity, conidial surface morphology and conidial hydrophobicity in M. acridum. These findings unravel the regulatory mechanism of Mavib-1 in fungal growth and conidiation, and enrich the knowledge to conidiation pattern shift of filamentous fungi.
1. Introduction
The insect pathogenic fungi Metarhizium spp. and Beauveria spp. have been widely used all over the world and play an important role in the control of agricultural pests [1,2,3,4,5,6,7]. Representative species, such as Metarhizium anisopliae, Metarhizium acridum and Beauveria bassiana, have been developed as environmentally friendly biopesticides [8,9]. Entomopathogenic fungi mainly infect host insects through conidia, which are produced under complex regulation. Clarification of the regulation mechanisms of conidiation is also helpful for understanding their invasion and growth inside host insects.
Most fungi have two types of conidiation: normal conidiation (NC) and microcycle conidiation (MC) [10]. Normally, conidiophores arise from the vegetative mycelia and produce large amounts of conidia [11,12], which is the most common conidiation type for filamentous fungi [13]. The fungi can bypass the mycelia period in MC and develop secondary conidia from germ tubes or directly from conidial cells [1,4,14]. MC is a special survival mechanism of fungi under adverse conditions, such as high temperature [10,11,15], extreme pH [16], high salt content [17], and nutritional deficiencies [4,18,19,20]. Among them, nutritional deficiency is the important factor that affects the fungal growth and development.
A previous investigation showed that M. acridum produces conidia by NC on nutrient-rich medium 1/4 SDAY (Sabouraud dextrose medium with yeast extract) and MC on the nutrient-deficient medium SYA (sucrose yeast extract agar) [4]. However, adding a single carbon source (sucrose) or nitrogen source (sodium nitrate) in SYA medium can make the M. acridum switch to the NC type, indicating that nutrient element plays important roles in the conidiation pattern shift in M. acridum [20]. The underlying regulation mechanisms of changes from conidiation to vegetative growth when nutrients are changed are still largely unknown. A Vib-1 gene (vegetative incompatibility blocked) was highly upregulated when sucrose was added to SYA medium, indicating a possible role of Vib-1 in conidiation and conidiation pattern shift by affecting the nutrient utilization in M. acrdium.
Vib-1 belongs to the Ndt80/PhoG DNA binding protein family and contains a conserved DNA-binding domain NDT80_PhoG. In Neurospora crassa, Vib-1 participates in the regulation of heterokaryotic incompatibility and conidiation [21], and also regulates the expression of cell wall-degrading enzymes (PCWDEs) [22]. Vib-1 acts as a main regulator of the response to carbon and nitrogen starvation. Loss of Vib-1 leads to the imbalance of Carbon Catabolite Repression (CCR), which inhibits glucose signaling and CCR under carbon-limited conditions in N. crassa [23]. In Aspergillus nidulans, Vib-1 is required for production of extracellular proteases in response to carbon and nitrogen starvation [23,24]. However, the roles of Vib-1 in entomopathogenic fungi and its regulation mechanism in conidiation and fungal growth are not clear.
On SYA medium, the change of nutrients can directly lead to a switch between vegetative growth and conidiation, that is, switching between MC and NC. Therefore, in this study, we used SYA medium to analyze the effect of Mavib-1 on fungal development under different nutritional conditions in M. acridum through gene disruption and overexpression techniques. Exploring the function of transcription factor Mavib-1 and its mechanism of conidiation pattern shift in M. acridum will help to further understand the molecular mechanism of nutrient-regulated fungal conidiation mode shift.
2. Materials and Methods
2.1. Fungal Strains and Growth Conditions
M. acridum CQMa102 was used as a wild-type strain (WT) and stored in the China General Microbiological Culture Collection Center (CGMCC, No. 0877, NCBI Assembly No. GCF_000187405.1) [25]. The WT strain and transformants generated in this study were grown on 1/4 SDAY (1% dextrose; 0.25% peptone; 0.5% yeast extract and 2% agar) and incubated for 15 days at 28 °C. The influence of nutrient on conidiation was determined on SYA medium (sucrose 3%, yeast extract 0.5%, NaNO3 0.3%, K2HPO4 0.1%, MgSO4·7H2O 0.05%, KCl 0.05%, FeSO4·7H2O 0.001%, MnSO4·H2O 0.001% and agar 1.8%) or SYA with replaced sucrose by other carbon source. Dextrose, agar, NaNO3, MgSO4·7H2O and FeSO4·7H2O were purchased from Chengdu Kelon Chemical Co., Ltd. (Chengdu, China); yeast extract and peptone were purchased from Beijing Auboxing Biotechnology Co., Ltd. (Beijing, China); sucrose was purchased from Sinopharm Chemical Reagent Co., Ltd. (Shanghai, China). All chemicals were purchased from Chongqing Chuandong Chemical Co., Ltd. (Chongqing, China) unless stated separately.
2.2. Deletion, Complementation and Overexpression of the Mavib-1 Gene
The Mavib-1 (Gene ID: MT954969) disruption mutant was constructed by homologous recombination. For Mavib-1 deletion, upstream and downstream flanking fragments (~1 kb) of Mavib-1 coding sequences were amplified and inserted into the pK2-PB vector which harbored a bar cassette. The resulting vector pK2-PB-Mavib-1-LR was transformed into M. acridum mediated by Agrobacterium tumefaciens [26]. The Mavib-1 disruption mutants (ΔMavib-1) were screened on Czapek-Dox medium (sucrose 30 g/L, NaNO3 2 g/L, KH2PO4 1 g/L, MgSO4·7H2O 0.5 g/L, KCl 0.5 g/L, FeSO4·7H2O 0.01 g/L and agar 18 g/L) containing 500 μg/mL glufosinate-ammonium (Sigma, St. Louis, MO, USA). To rescue the deleted Mavib-1, the full-length Mavib-1 sequence including flanking regions (5.0 kb in total length) was amplified and inserted into the pK2-sur vector containing the chlorimuron ethyl resistance gene sur [27] to construct the complementary vector pK2-sur-Mavib-1. This vector was transformed into ΔMavib-1 and the complementary transformants (CP) were screened on Czapek-Dox plate containing 60 µg/mL chlorimuron ethyl (Sigma, Bellefonte, PA, USA). To create an Mavib-1 overexpression mutant, the Mavib-1 gene was fused with EGFP and driven by the promotor of constitutive glyceraldehyde 3-phosphate dehydrogenase gene (Magpd). This vector was transformed into the WT and transformants (OE) were screened on Czapek-Dox plates supplemented with 500 µg/mL glufosinate-ammonium (Sigma, St. Louis, MO, USA). The transformants were verified by PCR, and florescence was observed under a laser scanning confocal microscope (LSCM, TCS SP8, Leica, Germany).
2.3. Transcription Activation Experiment
For the transcriptional activation test, full-length cDNA, N-terminal (contain NDT80_PhoG domain) and C-terminal of Mavib-1 was amplified from a cDNA library with primer pairs Mavib-1-BD-F1/Mavib-1-BD-R2, Mavib-1-BD-F1/Mavib-1-BD-R1, Mavib-1-BD-F2/Mavib-1-BD-R2 (Table S1), respectively, and the amplified fragments were respectively inserted into the vector pBKT7 (Clontech) between the NdeI and EcoRI sites under the control of the GAL4 promoter to generate plasmid pBKT7-Mavib-1 for transformation into the Yeast Y2H Gold strain. Putative prototrophic transformants were selected on a Trp-free SD medium and further transferred onto Trp/His/Ade-free SD plates containing 0.5 mM 5-Bromo-4-Chlroro-3-Indolyl-D-Galactopyranoside (X-α-gal, TaKaRa, Dalian, China) for transcriptional activation assay.
2.4. Stress Tolerance Assays
For the stress assays, aliquots of 2 µL conidial suspensions (1 × 106 conidia/mL) of the WT, ΔMavib-1, CP and OE strains were pipetted onto 1/4 SDAY plates supplemented with 1 mol/L NaCl, 0.01% SDS, 1 mol/L sorbitol (SOR) (Dingguo, Beijing, China), 500 µg/mL Congo Red (CR) (Solarbio, Beijing, China), 6 mmol/L H2O2, 50 µg/mL calcofluor white (CFW) (Solarbio, Beijing, China), respectively. The plates were incubated at 28 °C for 5 days. Stress tolerance to heat and ultraviolet radiation was assessed as described previously [4]. Half inhibition time of germination (IT50) under UV radiation or heat was then calculated for each strain. The assays were conducted with three replicates per treatment and the experiment was repeated three times.
2.5. Conidial Germination
Conidial germination assay was conducted on 1/4 SDAY plates as described previously [28]. The conidia of M. acridum WT, ΔMavib-1, CP and OE strains were suspended in sterile water (1 × 107/mL). Aliquots of 50 µL conidia suspension were spread on 1/4 SDAY plates and the plates were incubated at 28 °C. The conidial germination rate of each strain was determined every 2 h (cultured for 2, 4, 6, 8, 10 and 12 h, respectively) until conidia germinate almost completely. The half time to germination (GT50) was statistically calculated, and the data were analyzed. Three replicates in each independent experiment were conducted, and in each replicate, 300 conidia were used.
2.6. Electronic Microscopy
Ultrastructure of mature conidial cell wall surface of the WT, ΔMavib-1, CP and OE strains was examined by scanning electronic microscopy (SEM, SU8010, Hitachi, Japan) as described previously [29]. Conidia of WT, ΔMavib-1, CP and OE strains of M. acridum were collected, fixed with 2.5% glutaraldehyde solution (Servicebio, Wuhan, China) at 4 °C overnight, and then the samples were processed as follows: remove the fixation solution, rinse the samples three times with 0.1 M phosphate buffer, pH 7.0, 15 min each time; fix the sample with 1% osmic acid solution for 1–2 h; carefully remove the osmic acid waste solution, rinse the sample three times with 0.1 M, pH7.0 phosphate buffer for 15 min each time; use a gradient concentration (including 30%, 50%, 70%, 80%, 90% and 95%) ethanol solution to dehydrate the samples with each concentration for 15 min, and then treated with 100% ethanol twice, each time for 20 min. Rinse the sample with a mixture of ethanol and isoamyl acetate (v/v = 1/1) for 30 min, and then rinse with pure isoamyl acetate for 1h or leave the samples overnight. After critical point drying and coating, the samples were observed in a scanning electron microscope.
2.7. Conidial Hydrophobicity Assay
Conidial hydrophobicity was determined with hexadecane as described previously [29]. Conidia of WT, ΔMavib-1, CP and OE strains were suspended (3 × 107 conidia/mL) in reaction buffer (22.2 g/L K2HPO4, 7.26 g/L KH2PO4, 1.8 g/L urea, 0.2 g/L MgSO4, pH 7.1). Aliquots of 1 mL conidia suspension (OD470 = 0.4) were mixed well with 100 μL hexadecane and then incubated at 25 °C for 10 min. The upper hexadecane was discarded. The absorption peak of conidia suspension at 470 nm (A470) was measured after removing solidified hexadecane completely after incubation at 4 °C for 30 min. The hydrophobic index was calculated using the following formula: (A470, control-A470, hexadecane-treated)/(A470, control). The assays were conducted with three replicates per treatment and the experiment was repeated three times.
2.8. Conidiation and Colony Morphology with Various Carbon or Nitrogen Sources
SYA containing different carbon sources at 3% (w/v) (glucose, sucrose and glycerol, respectively) or SYA containing different nitrogen sources at 3% (w/v) (urea, glutamine/Gln, NaNO3, respectively) were used for colony growth assays and conidiation analysis. For conidiation observation under microscope, each plate was spread evenly with 100 µL of conidial suspension (1 × 107 conidia/mL) and incubated at 28 °C for 12–36 h. For colony growth assay, aliquots of 2 µL of conidial suspension (1 × 106 conidia/mL) of each strain were inoculated on different media and the plates were incubated at 28 °C for 5 days. For conidial yield assay, 2 µL conidial suspensions (1 × 106 conidia/mL) of WT, ΔMavib-1, CP and OE were spotted onto ¼ SDAY medium in a 24-hole plate and incubated at 28 °C. Conidia were collected after growing for 15 days and vortexed with 1 mL of 0.05% Tween 80. Conidial concentration was determined using a hemocytometer under a microscope. All experiments were performed three times.
2.9. RNA-Seq
Transcriptional profiles were measured in WT, Δmavib-1 and OE strains by RNA-Seq. Total RNA was extracted from WT, Δmavib-1, or OE strains after growing on SYA plates at 28 °C for 20 h. Total RNA was extracted from each sample using an RNA Kit (Invitrogen, Carlsbad, CA, USA) with Rnase-free DnaseI added and reverse transcribed into cDNA with an oligo-dT primer using the PrimeScript RT Master Mix (TaKaRa, Dalian, China). Sequencing libraries were prepared, and sequencing was conducted on BGISEQ-500 (BGI, Beijing, China). Genes were classed as significantly differentially expressed when false discovery rate (FDR) <0.001 and fold change ≥2.
Differentially expressed genes (DEGs) were classified and annotated using gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis.
2.10. Reverse Transcription Quantitative PCR (RT-qPCR)
The gene expression levels were examined by RT-qPCR. Amplification mixtures (20 µL) contained 1 µL template cDNA, 10 µL IQ SYBR Green Supermix (2×, Bio-Rad, Foster City, CA, USA) and 1 µL of each primer (qF/qR, 10 µmol/L) [27]. The reactions were performed in iCycler system (Bio-Rad). The Magpd gene (EFY84384), amplified with primers gpd-F/gpd-R (Table S1), was used as the endogenous control, as described previously [27]. Transcript ratios of the target gene were evaluated using the 2−ΔΔCT method [30]. All PCR amplifications were conducted in triplicate, and trials were repeated three times.
2.11. Data Analysis
All measurements were analyzed using a one-way ANOVA model with the SPSS 19.0 program (SPSS Inc., Chicago, IL, USA). Tukey’s honestly significant difference test was used to evaluate means at α = 0.05.
3. Results
3.1. Structural Features and Deletion of Mavib-1
Mavib-1 gene has a full-length sequence of 1991 bp containing two introns (67 bp and 79 bp). A cDNA sequence of 1845 bp (Acc. MT954969) encodes a protein of 614 amino acids with a molecular weight of 66.90 kDa and an isoelectric point of 6.84. The Mavib-1 protein contains an NDT80_PhoG domain with 165 amino acids (Residues 171–335) (Figure S1A), which is highly conserved in Vib-1 homologs among different fungi (Figure S1B). Phylogenetic analysis revealed that Mavib-1 was clustered with other fungal NDT80_PhoG domain containing proteins (Figure S1C). To elucidate the functions of Mavib-1 in M. acridum, ΔMavib-1, CP and OE strains were constructed (Figure S2A), and the expected recombinant events were verified by PCR (Figure S2B), Semi-quantitative RT-PCR (Figure S2C) and RT-qPCR analysis (Figure S2D).
3.2. C-Terminal of Mavib-1 Has Transactivation Activity
Mavib-1-N (containing NDT80 domain), Mavib-1-C or Mavib-1 full CDS were fused to the GAL4 DNA-binding domain and expressed in yeast strain Y2H Gold. Mavib-1-C fusion strain showed blue spots on SD-Trp plates containing X-α-gal and AbA, indicating a transactivation activity at C-terminal of Mavib-1 in yeast (Figure 1A). To investigate the subcellular localization of Mavib-1, Mavib-EGFP was expressed in M. acridum. Results showed that green fluorescent signal was observed in the nucleus in both conidia and mycelia and overlapped with the staining of DAPI dye, indicating a nuclear localization of Mavib-1 (Figure 1B). Therefore, consistent with Vib-1 in A. nidulans and N. crassa [22,23], Mavib-1 can functionally act as a transcription factor in M. acridum.
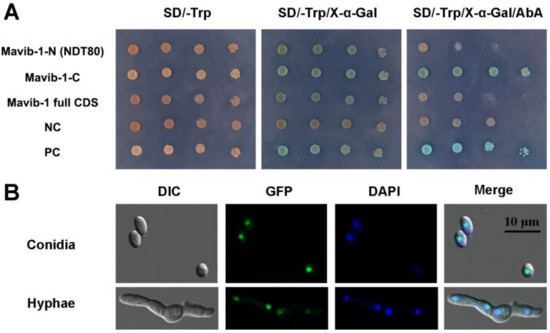
Figure 1.
Transcriptional-activation assays and subcellular localization of Mavib-1. (A) Transcriptional activation assays in yeast. Mavib-1-N (containing NDT80 domain), Mavib-1-C or Mavib-1 full CDS were fused to the GAL4 DNA-binding domain and expressed in yeast strain Y2H Gold. Strain containing only GAL4 DNA-binding domain was as negative control (NC), containing the GAL4 DNA-binding and activation domains were as positive controls (PC). All the yeast strains were cultured on SD-Trp plates, SD-Trp plates containing 0.5 mM X-α-gal and SD-Trp plates containing 0.5 mM X-α-gal and 200 ng/mL AbA (Aureobasidin A) at 30 °C for 3 days. (B) Subcellular localization of Mavib-1. The expression of Mavib-1 and EGFP fusion protein was driven by strong promoter PgpdM. The conidia and mycelia of OE strain were stained with DAPI (4′,6-diamidino-2-phenylindole, C0065, Solarbio, China), a fluorescent dye that can bind strongly to DNA, and observed under a confocal microscope (TCS SP8, Leica, Germany).
3.3. Mavib-1 Affected Stress Tolerances of M. acridum
Stress tolerances are very important characteristics for fungal potential against insect pests. Compared to the WT, the ΔMavib-1 mutant showed a significant change in conidial sensitivity to UV radiation and heat stress (Figure 2). After UV-B irradiation, conidial germination and mean IT50 values of ΔMavib-1 mutant were significantly decreased compared to WT, while OE strain did not show significant difference compared to WT (Figure 2A,B). After heat treatment, both ΔMavib-1 and OE strains presented a significant decrease in conidial germination and mean IT50 values compared to WT, but ΔMavib-1 showed a sharply decreased in heat treatment (Figure 2C,D). CP strain showed a partial recovery in UV-B and heat stress tolerance compared to WT and ΔMavib-1 mutant.
Figure 2.
Stress tolerances of conidia from WT, ΔMavib-1, CP and OE strains. (A) Germination rate of each strain on 1/4 SDAY media at 28 °C for 20 h after UV-B stress treatment at 0, 1.25, 2.5, 3.75, 5.0 h. (B) The half inactivity time (IT50) of each strain with UV-B treatment. (C) Germination rates on 1/4 SDAY at 28 °C for 20 h after 46 °C heat stress treatment at 0, 3, 6, 9, 12 h. (D) The IT50 of WT, ΔMavib-1, CP and OE strains under 46 °C heat treatment. (E) The fungal colony on 1/4 SDAY medium and 1/4 SDAY with cell wall disruptors (500 µg/mL CR, 50 µg/mL CFW), cell wall stressor (0.01% SDS), hyperosmotic stressors (1 mol/L SOR or 1 mol/L NaCl), oxidative stress (6 mmol/L H2O2), respectively. (F) Inhibition rate of colony growth. All experiments were repeated three times for statistical analysis. Different capital letters represented significant difference at p < 0.01; different lower-case letters represented significant difference at p < 0.05.
ΔMavib-1 and OE did not show significant difference in tolerance to SDS, SOR and NaCl compared to WT. The OE strain had more sensitivity to CR and CFW, while ΔMavib-1 had weaker tolerance to CR and H2O2 (Figure 2E,F), indicating a destroyed cell wall integrity in ΔMavib-1 and OE strains.
3.4. Disruption of Mavib-1 Affected Conidial Germination, Conidial Cell Wall Surface Ultrastructure, and Conidial Hydrophobicity
Germination rates of conidia of WT, ΔMavib-1, CP and OE strains were determined on 1/4 SDAY at different culture intervals. Results showed that GT50 of ΔMavib-1 and OE strains were significantly prolonged than that of WT (Figure 3A). The Mavib-1 gene affected the resistance to cell-wall-disrupting agent CFW (Figure 2E,F); we speculated that Mavib-1 might affect the conidial cell wall structure. SEM analysis showed that the conidial surface of WT and OE strains had a rough cell wall surface, while the ΔMavib-1 strain had a smoother cell wall surface compared with WT (Figure 3B).
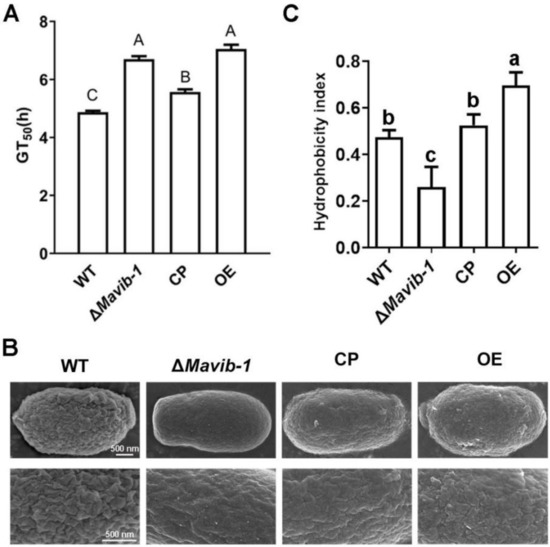
Figure 3.
Conidial germination, conidial surface morphology and hydrophobicity determination. (A) The 50% germination times (GT50) of WT, ΔMavib-1, CP and OE strains. (B) The conidial surface morphology by SEM. (C) Conidial hydrophobicity index. The trial was repeated three times. Different capital letters represented significant difference at p < 0.01 and different lower-case letters indicate significant difference, p < 0.05.
The structure of the fungal cell wall surface is closely related to the conidial characteristics. The rodlets on the conidial surface are arranged according to certain rules by fungal hydrophobins [31]. Hydrophobicity assay showed that the conidial hydrophobic index was significantly decreased in ΔMavib-1 and significantly increased in OE strain compared to WT (Figure 3C). These results indicated that deletion of Mavib-1 affected conidial germination, conidial cell wall surface ultrastructure and conidial hydrophobicity in M. acridum.
3.5. Disruption of Mavib-1 Affected Growth and Conidiation of M. acridum on Different Carbon or Nitrogen Source Media
Mavib-1 has a nutrient sensing-related NDT80_PhoG domain, and the growth, conidiation and conidial yield of WT, ΔMavib-1, CP and OE strains on media with different carbon and nitrogen sources were determined. On the nutrient-rich medium 1/4 SDAY, ΔMavib-1 and OE strains had similar colony color and size compared to WT. On nutrient-limited media, in which glucose, sucrose or glycerol was used as the main carbon source, respectively, or no extra carbon source was added, ΔMavib-1 colony had a much lighter color and significantly smaller size than WT, while the OE strain had a similar colony phenotype as WT. Conidial production analysis showed that deletion of Mavib-1 resulted in decreased conidial yield of M. acridum regardless of rich nutrient or starvation (Figure 4C). These results indicated that Mavib-1 gene is necessary for colony growth and the conidial melanin formation under nutrient deficiency in M. acridum. Microscopic observation showed that the ΔMavib-1 strain performed normal conidiation similar to WT and OE at the early growth time on 1/4 SDAY (12 h and 24 h). However, ΔMavib-1 strain produced more conidia than WT and OE at 36 h. Conidiation was also analyzed on SYA, a microcycle conidiation medium for M. acridum [4]. It clearly showed that WT and ΔMavib-1 conducted MC, while the OE strain changed to typical hyphal growth and conducted normal conidiation. On other media without a carbon source, or with glucose and glycerol as carbon source, ΔMavib-1 exhibited normal conidiation slightly earlier than WT, while the OE strain did not show conidiogenous structure until 36 h (Figure 4D).
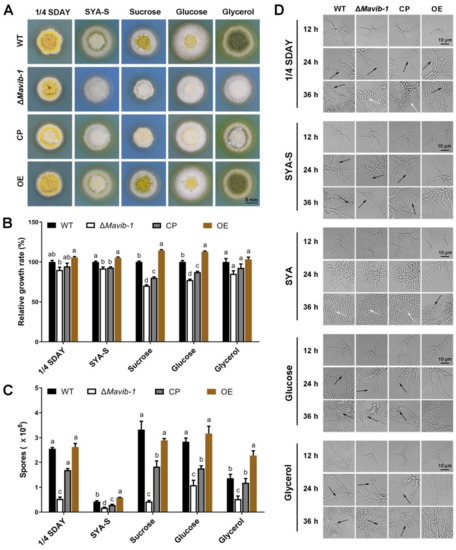
Figure 4.
Colony growth and conidiation of fungal strains on media containing different carbon sources. (A) Colony morphology of WT, ΔMavib-1, CP and OE strains. Fungal strains were cultured on the nutrient-rich medium 1/4 SDAY, and nutrient-limited medium SYA with easy-to-use or difficult-to-use carbon sources (3% w/v; easy to use: glucose; middle: sucrose, difficult to use: glycerol) for 5 d. Colony relative growth rate (B) and conidial yield (C) of WT, ΔMavib-1, CP and OE strains cultured on 1/4 SDAY or SYA with different carbon sources for 5 d. The trial was repeated three times. Error bars are standard deviations of three replicates. Different lower-case letters indicate significant difference at p < 0.05. (D) Conidiation of WT, ΔMavib-1, CP and OE strains on different media. The black arrows indicate normal conidiation, and the white arrows indicate microcycle conidiation.
Similar to the role in carbon source utilization, Mavib-1 also contributed to the utilization of different nitrogen sources, including dominant (glutamine) and non-dominant (urea and NaNO3) nitrogen sources (Figure 5A). ΔMavib-1 colonies were whitish in color on SYA medium with different nitrogen sources (Figure 5A) and had significantly smaller size than WT and CP, when urea or NaNO3 was used as the main nitrogen source or no extra nitrogen source (SYA-N) was included, while the OE strain had an obviously larger colony on these media (Figure 5A,B). These results indicated that Mavib-1 positively regulated fungal growth by affecting utilization of nitrogen sources. Conidial yield analysis showed that ΔMavib-1 had significantly decreased conidia production compared to WT when urea or NaNO3 was used as main nitrogen source, while overexpression of Mavib-1 did not significantly affect conidial yield in M. acridum (Figure 5C). Microscopic observation showed that, similar as the results on the media with different carbon sources, ΔMavib-1 strain performed MC on SYA-N and SYA medium slightly earlier compared to WT. WT strain performed hyphal growth at 20 h, while ΔMavib-1 strain had already produced a mass of conidia at that time. Conidiation of OE strain was drastically delayed compared with WT on different nitrogen source media, and no conidia was formed until 36 h on the SYA-N and SYA-Urea media (Figure 5D). These results indicated that Mavib-1 positively regulated fungal growth and negatively regulated conidiation of M. acridum by affecting utilization of different carbon or nitrogen sources especially when nutrient was insufficient. However, Mavib-1 contributed to total conidial yield in M. acridum under both nutrient-rich and nutrient-limited conditions.
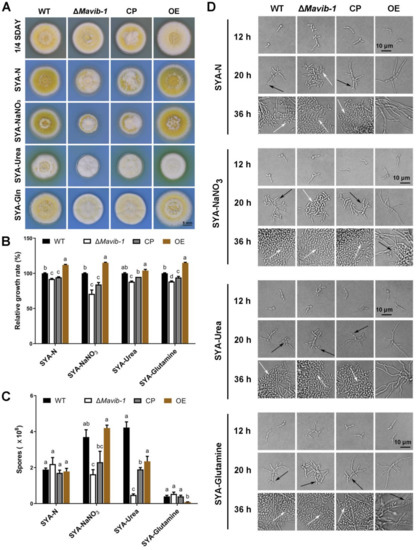
Figure 5.
Colony growth and conidiation of ΔMavib-1, CP and OE strains on media with different nitrogen sources. Colony morphology (A) and colony relative growth rate (B) of ΔMavib-1, CP and OE strains on the media 1/4 SDAY, and SYA with different nitrogen sources. (C) Conidial yield of fungal strains cultured on different media for 15 d. Media contained 0.3% nitrogen source: SYA-N (SYA without NaNO3), urea (difficult to use), NaNO3 (middle), Gln (easy to use). Each trial was repeated three times for statistical analysis. Different lower-case letters mean significant difference at p < 0.05. (D) Conidiation of ΔMavib-1, CP and OE strains on different media. The black arrows indicate NC, and the white arrows indicate MC.
3.6. Mavib-1 Affected Expression of CAZyme Genes of M. acridum
To elucidate the mechanism by which Mavib-1 affects the conidiation of M. acridum in response to different carbon and nitrogen sources, RNA-seq was performed for WT, ΔMavib-1 and OE strains cultured on SYA medium for 20 h (Accession No. PRJNA73660). The RNA-seq data were verification by RT-qPCR, and the difference fold changes obtained by the two methods were extremely significantly correlated (R = 0.890 and p < 0.01 in ΔMavib-1, R = 0.960 and p < 0.01 in OE), indicating that the RNA-seq data were reliable (Table S2). Primer sequences used for verification are listed in Table S1.
Statistical analysis was performed on the RNA-seq data of three groups. The correlation heat map showed that the correlation between the three replicates in each group was high, and the correlation between groups was small, indicating a good repeatability of sequence data (Figure S3A). Principal component analysis (PCA) showed that the three replicates in each group could be clustered together, again indicating that the data were reproducible (Figure S3B). The RNA-seq data were screened after quality control and qPCR validation and genes with |log2FC |≥ 1, Qvalue ≤ 0.01 and FPKM ≥5 were counted as DEGs (Tables S3 and S4). RNA-seq showed that there were a total of 291 DEGs (79 up-regulated, 212 down-regulated) in the ΔMavib-1 strain, and a total of 486 DEGs (318 up-regulated and 168 down-regulated) in the OE strain (Figure 6A) compared to WT. The Venn diagram showed that there were 101 shared DEGs between ΔMavib-1 and OE (66 DEGs both down-regulated, 13 DEGs both up-regulated, and 18 DEGs down-regulated in ΔMavib-1 and up-regulated in OE, 4 DEGs up-regulated in ΔMavib-1 and down-regulated in OE) (Figure 6B). The detailed information of 22 DEGs (18 downregulated and 4 upregulated in ΔMavib-1), with opposite changing trend of transcription in ΔMavib-1 and OE, is listed in Table S5. To intuitively reflect these DEGs with the opposite changing trend, a cluster heat map of 22 DEGs was constructed (Figure 6C,D).
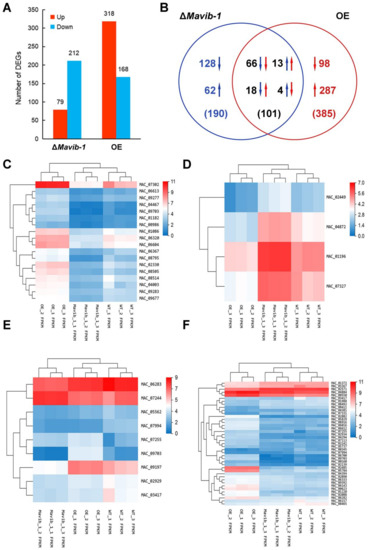
Figure 6.
DEGs analysis. (A) Statistical histogram of the number of DEGs. (B) Venn diagram of DEGs. (C) Heat map of 18 DEGs downregulated in ΔMavib-1 and upregulated in OE. The 18 DEGs consistent with the change trend of Mavib-1 were analyzed by cluster analysis of expression patterns. The distance calculation method was used: the spearman correlation coefficient between samples, the Pearson correlation coefficient between genes. (D) Cluster heat map of the expression levels of the 4 genes upregulated in ΔMavib-1 and downregulated in OE. (E) Heat map of the expression of DEGs in ΔMavib-1 annotated as CAZyme genes. (F) Heat map of the expression of DEGs in OE annotated as CAZyme genes. The up and down arrows represent upregulated or downregulated genes, respectively. Numbers in blue represent ΔMavib-1 and red represent OE. The red color denotes the upregulated DEGs, and the blue color denotes downregulated DEGs. The log2 (expression value + 1) of the sample were shown in the horizontal axis, and DEGs are shown at the right-hand vertical side.
GO classification (Figure S4), KEGG enrichment (Figure S5) and carbohydrate activity enzymes (CAZymes) annotation analysis were performed for DEGs. The up-regulated genes in ΔMavib-1 strain were mainly enriched in RNA polymerase, nitrogen metabolism and atrazine degradation, etc., and the down-regulated genes were mainly enriched in neuroactive ligand–receptor interaction, protein digestion and absorption, pancreatic secretion, influenza A and glycerolipid metabolism, etc. (Figure S5A,B). The upregulated genes in the OE strain were mainly enriched in microbial metabolism in diverse environment, metabolic pathways, and beta-alanine metabolism, etc. The downregulated genes were mainly enriched in amino sugar and nucleotide sugar metabolism, phenylpropanoid biosynthesis, neuroactive ligand–receptor interaction, protein digestion and absorption and metabolic pathways, etc. (Figure S5C,D). These results indicated that Mavib-1 affected the carbon and nitrogen source metabolic pathways of M. acridum. Aligning the DEGs with the CAZy database [32] revealed that many DEGs belong to CAZymes (9 of the 291 DEGs in the ΔMavib-1 strain listed in Table S6, and 41 of the 486 DEGs in the OE strain listed in Table S7). Cluster heatmap of DEGs belonging to CAZymes showed that the expression of CAZyme genes in ΔMavib-1 and OE strains have changed (Figure 6E,F), indicating that Mavib-1 played an important role in regulation of the expression of CAZyme genes in M. acridum. At the same time, 21 nitrogen source metabolism-related genes (listed in Table S8) were also found in DEGs. The transcription changes of these nitrogen metabolism and conidiation related genes in ΔMavib-1 and OE strains may provide explanations for the effects of Mavib-1 on nitrogen metabolism and conidiation in M. acridum.
4. Discussion
The growth and development of filamentous fungi require suitable conditions such as temperature, humidity, pH, and nutrients. Filamentous fungi can sense the nutrient conditions in the environment and regulate their development. In this study, we demonstrated the role of Vib-1 in NC and MC by affecting the nutrition utilization in entomopathogenic fungus M. acridum.
Mavib-1 shares the NDT80_PhoG domain with Ndt80 of S. cerevisiae [33], Vib-1, Fsd-1 and NCU04729 of N. crassa [34,35], and XprG and NdtA of A. nidulans [36,37,38]. Mavib-1 was localized at nuclear in both conidia and hyphae, while Vib-1-GFP was found in nuclear in hyphae and cytoplasmically localized in conidiophores and immature conidia in N. crassa [21]. This might cause some divergent roles of Vib-1 in different fungi. Proteins containing NDT80_PhoG domains had been reported in model fungi yeast and N. crassa. In S. cerevisiae, Ndt80 is a meiosis-specific transcription factor that regulates the expression of metaphase genes and also regulated by the pachytene checkpoint [39,40,41]. In filamentous fungi, sporogenesis is a procedure of producing a new cell through cell division by mitosis, which Vib-1 might be involved in the regulation of conidiation. Consistent with our result, Vib-1 was involved in the regulation of conidiation in N. crassa [21].
Our findings indicate that Mavib-1 positively regulates fungal growth, and negatively regulates conidiation especially under nutrient deficient conditions. Consistent with the decreased hyphal extension in Vib-1 disruption mutant in N. crassa [21], ΔMavib-1 had shortened hyphae when growing on nutrient-rich medium. However, conidiation was decreased in ΔMavib-1 and showed no changes when Mavib-1 was upregulated, in contrast with no changes in Vib-1 mutant and decreased conidiation in Vib-1 overexpression strain in N. crassa [21], indicating a divergent role of Vib-1 in different fungi. When under carbon-limited conditions, deletion of N. crassa Vib-1 causes a growth defect on cellulose biomass [23]. Some other Ndt80-containing proteins were reported to contribute to growth and nutrient utilization. Ndt80 transcription factor RON1 is essential for hyphal growth and N-acetylglucosamine (GlcNAc) metabolism in Crytococcus neoformans [42]. In A. nidulans, the Ndt80-like protein XprG positively regulates the expression of extracellular proteases, mycotoxins and penicillins in response to carbon starvation [37]. In M. acridum, disruption of the Vib-1 led to a more serious defect in growth and conidiation initiation under carbon and nitrogen limited condition, but OE strain displayed a contrast phenotype with ΔMavib-1, demonstrating more important roles of Mavib-1 in both carbon and nitrogen utilization under nutrient limitation. Therefore, the explanation for the conidiation pattern change of Mavib-1 mutants was as follows. When Mavib-1 was impaired, M. acridum could not obtain enough nutrients to support hyphal growth and then the conidiation was induced, while hyphal growth was retained and strengthened owing to an improvement of nutrient utilization during overexpression of Mavib-1.
Consistent with the inhibited hyphal growth in ΔMavib-1 and promoted growth in OE strain on plate, RNA-Seq analysis revealed that DEGs of ΔMavib-1 and OE strains included many CAZyme members, which can degrade, modify or create glycosidic bonds. In ΔMavib-1, the DEGs related to CAZyme were mostly downregulted (seven downregulated in nine DEGs) compared to upregulation of most CAZyme DEGs (27 upregulated in 41 DEGs) in OE strain. This indicated a more active carbon metabolism when Mavib-1 was overexpressed. Similar in N. crassa, Vib-1 can repress glucose signaling and CCR [23], and cellulose digestion-related genes were increased and glucose metabolism related CAZyme were repressed under carbon starvation [22,23] in Vib-1 disruption mutant.
In Aspergillus niger, nutrient limitation, such as severe carbon limitation, led to a zero growth rate and induction of conidiation [43,44]. M. acridum show similar growth phenotype on minimal medium (Figure S6A), on which all WT and mutant strains conducted MC and no hyphal growth was observed under microscope (Figure S6B). This made it impossible to analyze the shift between hyphal growth and conidiation. On minimal medium, OE strain had a larger colony size and a better growth than ΔMavib-1, which was consistent with the result on nutrient-limited medium SYA (Figure S6A). Therefore, we preferred analysis of hyphal growth and conidiation on nutrient-limited media SYA, on which M. acridum might change to normal conidiation when conidiation was in dysregulation.
The fungal cell wall acts as an environmental barrier to the host immune system and plays a crucial role in the pathogenic process and stress tolerance of entomopathogenic fungi [45]. Elements involved in pathways of cell wall integrity would affect the fungal virulence and stress tolerance to heat, UV irradiation or chemical stressors [46,47,48]. In M. acridum, Vib-1 disruption or overexpression affected many genes related to CAZyme, which would also affect the cell wall synthesis. The conidial pigment formation-related gene laccase, also a virulence factor [49], was downregulated in ΔMavib-1 and upregulated in OE strain (Table S5), which was consistent with the light color of colony and decreased pathogenicity in ΔMavib-1. Agreed with decreased UV tolerance, riboflavine-aldehyde-forming enzyme gene (MAC_01768), contributed to UV tolerance and virulence in Metarhizium [50], was significantly downregulated in ΔMavib-1. SEM showed that ΔMavib-1 had abnormal conidial surface morphology compared to WT and OE. These changes in gene transcription or cell wall structure might lead to changes tolerance to environmental stresses and virulence. ΔMavib-1 did not show opposite transcription changes with Mavib-1-OE strain in all the tested phenotype, such as decreased virulence, decreased UV and heat tolerance in ΔMavib-1, while there were no changes for these phenotypes in OE. This may be due to the complexity of biological regulation. After a gene is knocked out or overexpressed, it may lead to expression changes of other related genes to make up for the defects, and finally result in an atypical phenotype. Previous reports showed that the damage of conidial cell wall surface results in reduced pathogenicity, but enhanced thermotolerance in B. bassiana [31]. Moreover, conidial characteristics are closely related to the adherence and invasion ability of entomopathogenic fungi [29,51]. The changes in conidial characteristics in ΔMavib-1 would possibly affect the fungal pathogenicity. Some pathogenicity-related genes were also affected by Mavib-1. For example, citrinin biosynthesis transcriptional activator CtnR [52] and polyketide synthase protein (MAC_01101) [53], were differentially expressed in ΔMavib-1 or OE strains (Tables S3 and S4). These results suggested that Vib-1 might contribute to virulence in M. acridum.
5. Conclusions
In summary, our study indicated that the transcription factor Mavib-1 positively regulated fungal growth and negatively regulated conidiation by affecting utilization of carbon and nitrogen sources in M. acridum. Owing to the contribution in the regulation of nutrient utilization, Mavib-1 contributed to the conidiation pattern shift from microcycle conidiation to normal conidiation under nutrient deficiency. Moreover, Mavib-1 is important for stress tolerance in M. acridum. Further analysis of the divergent upstream and downstream targets of Mavib-1 in regulating fungal growth, conidiation, stress tolerance and virulence will help explore the underlying regulative mechanisms of Vib-1 in response to nutrient and other environmental stimuli.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8060594/s1, Figure S1: Sequence analysis of Mavib-1. Figure S2: Construction of Mavib-1 disruption (ΔMavib-1), complementation (CP) and overexpression (OE) strains of the Mavib-1 gene in M. acridum. Figure S3: RNA-seq data quality control analysis. Figure S4: GO classification of DEGs. Figure S5: DEGs KEGG pathway enrichment analysis. Figure S6: Colony growth and conidiation of WT, ΔMavib-1, CP and OE strains on minimal medium (CZA). Table S1: Primers used in construction of Mavib-1 deletion mutant, complemented strain and overexpression. Table S2: Verification of DGEs results by RT-qPCR analysis. Table S3: DEGs of ΔMavib-1 compared to WT cultured on SYA medium for 20 h. Table S4: DEGs of Mavib-1 OE compared to WT cultured on SYA medium for 20 h. Table S5: DEGs with opposite changing trends of transcription in ΔMavib-1 and OE. Table S6 CAZyme in DEGs of Δmavib-1. Table S7: CAZyme in DEGs of OE. Table S8: DEGs related to nitrogen metabolism.
Author Contributions
Conceptualization, Y.C.; Methodology, X.S.; Validation, X.S. and H.L.; Investigation, X.S.; Resources, Y.C. and Y.X.; Writing—Original Draft Preparation, X.S.; Writing—Review and Editing, Y.C. and Y.X.; Visualization, X.S.; Supervision, Y.C. and Y.X.; Project Administration, Y.C.; Funding Acquisition, Y.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Natural Science Foundation of China (No. 31772222) and Natural Science Foundation Project of CQ CSTC (cstc2021jcyj-msxmX0261).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article and its Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pendland, J.C.; Hung, S.Y.; Boucias, D.G. Evasion of host defense by in vivo-produced protoplast-like cells of the insect mycopathogen Beauveria bassiana. J. Bacteriol. 1993, 175, 5962–5969. [Google Scholar] [CrossRef]
- Khurana, N.; Saxena, R.K.; Gupta, R.; Kuhad, R.C. Light-independent conidiation in Trichoderma spp.: A novel approach to microcycle conidiation. World J. Microbiol. Biotechnol. 1993, 9, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, J.M.; Charnley, A.K. New insights into the mechanisms of fungal pathogenesis in insects. Trends Microbiol. 1996, 4, 197–203. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Peng, G.X.; Xia, Y.X. Microcycle conidiation and the conidial properties in the entomopathogenic fungus Metarhizium acridum on agar medium. Biocontrol Sci. Technol. 2010, 20, 809–819. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- St. Leger, R.J.; Wang, C. Genetic engineering of fungal biocontrol agents to achieve greater efficacy against insect pests. Appl. Microbiol. Biot. 2010, 85, 901–907. [Google Scholar] [CrossRef]
- Ortiz-Urquiza, A.; Luo, Z.; Keyhani, N.O. Improving mycoinsecticides for insect biological control. Appl. Microbiol. Biot. 2015, 99, 1057–1068. [Google Scholar] [CrossRef]
- Peng, G.; Xia, Y. The mechanism of the mycoinsecticide diluent on the efficacy of the oil ormulation of insecticidal fungus. Biocontrol 2011, 56, 893–902. [Google Scholar] [CrossRef]
- Wang, C.S.; Wang, S.B. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Hanlin, R. Microcycle conidiation-A review. Mycoscience 1994, 35, 113–123. [Google Scholar] [CrossRef]
- Anderson, J.G.; Smith, J.E. The production of conidiophores and conidia by newly germinated conidia of Aspergillus niger (microcycle conidiation). J. Gen. Microbiol. 1971, 69, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Bosch, A.; Yantorno, O. Microcycle conidiation in the entomopathogenic fungus Beauveria bassiana bals. (vuill.). Process Biochem. 1999, 34, 707–716. [Google Scholar] [CrossRef]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Z.; Xia, Y.X. Identification of genes preferentially expressed during microcycle conidiation of Metarhizium anisopliae using suppression subtractive hybridization. FEMS Microbiol. Lett. 2008, 286, 71–77. [Google Scholar] [CrossRef]
- Sekiguchi, J.; Gaucher, G.M.; Costerton, J.W. Microcycle conidiation in Penicillium urticae: An ultrastructural investigation of conidiogenesis. Can. J. Microbiol. 1975, 21, 2069–2083. [Google Scholar] [CrossRef]
- Gestel, J.F.E.V. Microcycle conidiation in Penicillium italicum. Exp. Mycol. 1983, 7, 287–291. [Google Scholar] [CrossRef]
- Vézina, C.; Singh, K.; Sehgal, S.N. Sporulation of filamentous fungi in submerged culture. Mycologia 1965, 57, 722–736. [Google Scholar] [CrossRef]
- Saxena, R.K.; Khurana, N.; Kuhad, R.C.; Gupta, R. D-glucose soluble starch, a novel medium for inducing microcyclic conidiation in Aspergillus. Mycol. Res. 1992, 96, 490–494. [Google Scholar] [CrossRef]
- Ahearn, D.G.; Price, D.; Simmons, R.B.; Mavo, A.; Zhang, S.T.; Crow, S.A. Microcycle conidiation and medusa head conidiophores of Aspergilli on indoor construction materials and air filters from hospitals. Mycologia 2007, 99, 1–6. [Google Scholar] [CrossRef]
- Wang, Z.L.; Jin, K.; Xia, Y.X. Transcriptional analysis of the conidiation pattern shift of the entomopathogenic fungus Metarhizium acridum in response to different nutrients. BMC Genom. 2016, 17, 586. [Google Scholar] [CrossRef]
- Dementhon, K.; Iyer, G.; Glass, N.L. VIB-1 is required for expression of genes necessary for programmed cell death in Neurospora crassa. Eukaryot. Cell 2006, 5, 2161–2173. [Google Scholar] [CrossRef] [PubMed]
- Wu, V.W.; Thieme, N.; Huberman, L.B.; Dietschmann, A.; Kowbel, D.J.; Lee, J.; Calhoun, S.; Singan, V.R.; Lipzen, A.; Xiong, Y.; et al. The regulatory and transcriptional landscape associated with carbon utilization in a filamentous fungus. Proc. Natl. Acad. Sci. USA 2020, 117, 6003–6013. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Sun, J.; Glass, N.L. VIB1, a link between glucose signaling and carbon catabolite repression, is essential for plant cell wall degradation by Neurospora crassa. PLoS Genet. 2014, 10, e1004500. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.E.; Bernardo, S.M.; Cheetham, B.F. The interaction of induction, repression and starvation in the regulation of extracellular proteases in Aspergillus nidulans: Evidence for a role for CreA in the response to carbon starvation. Curr. Genet. 2008, 54, 47–55. [Google Scholar] [CrossRef]
- Gao, Q.; Jin, K.; Ying, S.H.; Zhang, Y.; Xiao, G.; Shang, Y.; Duan, Z.; Hu, X.; Xie, X.Q.; Zhou, G.; et al. Genome sequencing and comparative transcriptomics of the model entomopathogenic fungi Metarhizium anisopliae and, M. acridum. PLoS Genet. 2011, 7, e1001264. [Google Scholar] [CrossRef]
- Dos Reis, M.C.; Pelegrinelli Fungaro, M.H.; Delgado Duarte, R.T.; Furlaneto, L.; Furlaneto, M.C. Agrobacterium tumefaciens-mediated genetic transformation of the entomopathogenic fungus Beauveria bassiana. J. Microbiol. Methods 2004, 58, 197–202. [Google Scholar] [CrossRef]
- Luo, S.; He, M.; Cao, Y.Q.; Xia, Y.X. The tetraspanin gene MaPls1 contributes to virulence by affecting germination, appressorial function and enzymes for cuticle degradation in the entomopathogenic fungus, Metarhizium acridum. Environ. Microbiol. 2013, 15, 2966–2979. [Google Scholar] [CrossRef]
- Liu, J.; Cao, Y.Q.; Xia, Y.X. Mmc, a gene involved in microcycle conidiation of the entomopathogenic fungus Metarhizium anisopliae. J. Invertebr. Pathol. 2010, 105, 132–138. [Google Scholar] [CrossRef]
- Chen, X.; Liu, Y.Z.; Keyhani, N.O.; Xia, Y.X.; Cao, Y.X. The regulatory role of the transcription factor Crz1 in stress tolerance, pathogenicity, and its target gene expression in Metarhizium acridum. Appl. Microbiol. Biot. 2017, 101, 5033–5043. [Google Scholar] [CrossRef]
- Livak, K.; Schmittgen, T. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, S.; Xia, Y.X.; Kim, B.; Keyhani, N.O. Two hydrophobins are involved in fungal spore coat rodlet layer assembly and each play distinct roles in surface interactions, development and pathogenesis in the entomopathogenic fungus, Beauveria bassiana. Mol. Microbiol. 2011, 80, 811–826. [Google Scholar] [CrossRef] [PubMed]
- CAZy Database. Available online: http://www.cazy.org/ (accessed on 18 March 2021).
- Nocedal, I.; Mancera, E.; Johnson, A.D. Gene regulatory network plasticity predates a switch in function of a conserved transcription regulator. eLife 2017, 6, e23250. [Google Scholar] [CrossRef] [PubMed]
- Borkovich, K.A.; Alex, L.A.; Yarden, O.; Freitag, M.; Turner, G.E.; Read, N.D.; Seiler, S.; Bell-Pedersen, D.; Paietta, J.; Plesofsky, N.; et al. Lessons from the genome sequence of Neurospora crassa: Tracing the path from genomic blueprint to multicellular organism. Microbiol. Mol. Biol. Rev. 2004, 68, 1–108. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, E.A.; Glass, N.L. Meiotic regulators Ndt80 and Ime2 have different roles in Saccharomyces and Neurospora. Genetics 2010, 185, 1271–1282. [Google Scholar] [CrossRef] [PubMed]
- Katz, M.E.; Gray, K.A.; Cheetham, B.F. The Aspergillus nidulans xprG (phoG) gene encodes a putative transcriptional activator involved in the response to nutrient limitation. Fungal Genet. Biol. 2006, 43, 190–199. [Google Scholar] [CrossRef]
- Katz, M.E.; Braunberger, K.; Yi, G.; Cooper, S.; Nonhebel, H.M.; Gondro, C. A p53-like transcription factor similar to Ndt80 controls the response to nutrient stress in the filamentous fungus, Aspergillus nidulans. F1000 Res. 2013, 2, 72. [Google Scholar] [CrossRef] [PubMed]
- Krohn, N.G.; Brown, N.A.; Colabardini, A.C.; Reis, T.; Savoldi, M.; Dinamarco, T.M.; Goldman, M.H.; Goldman, G.H. The Aspergillus nidulans ATM kinase regulates mitochondrial function, glucose uptake and the carbon starvation response. G3 Bethesda 2014, 4, 49–62. [Google Scholar] [CrossRef]
- Lamoureux, J.S.; Glover, J.N.M. Principles of protein-DNA recognition revealed in the structural analysis of Ndt80-MSE DNA complexes. Structure 2006, 14, 555–565. [Google Scholar] [CrossRef]
- Chu, S.; Herskowitz, I. Gametogenesis in yeast is regulated by a transcriptional cascade dependent on Ndt80. Mol. Cell. 1998, 1, 685–696. [Google Scholar] [CrossRef]
- Hepworth, S.R.; Friesen, H.; Segall, J. NDT80 and the meiotic recombination checkpoint regulate expression of middle sporulation-specific genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 5750–5761. [Google Scholar] [CrossRef]
- Naseem, S.; Min, K.; Spitzer, D.; Gardin, J.; Konopka, J.B. Regulation of hyphal growth and N-acetylglucosamine catabolism by two transcription factors in Candida albicans. Genetics 2017, 206, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Nitsche, B.M.; Jørgensen, T.R.; Akeroyd, M.; Meyer, V.; Ram, A.F.J. The carbon starvation response of Aspergillus niger during submerged cultivation: Insights from the transcriptome and secretome. BMC Genom. 2012, 13, 380. [Google Scholar] [CrossRef] [PubMed]
- Jørgensen, T.R.; Nitsche, B.M.; Lamers, G.E.; Arentshorst, M.; van den Hondel, C.A.; Ram, A.F. Transcriptomic insights into the physiology of Aspergillus niger approaching a specific growth rate of zero. Appl. Environ. Microbiol. 2010, 76, 5344–5355. [Google Scholar] [CrossRef] [PubMed]
- Hopke, A.; Brown, A.J.P.; Hall, R.A.; Wheeler, R.T. Dynamic fungal cell wall architecture in stress adaptation and immune evasion. Trends Microbiol. 2018, 26, 284–295. [Google Scholar] [CrossRef] [PubMed]
- Latgé, J.P.; Mouyna, I.; Tekaia, F.; Baeuvais, A.; Debeaupuis, J.P.; Nierman, W. Specific molecular features in the organization and biosynthesis of the cell wall of Aspergillus fumigatus. Med. Mycol. 2005, 43 (Suppl. 1), S15–S22. [Google Scholar] [CrossRef]
- Rispail, N.; Soanes, D.M.; Ant, C.; Czajkowski, R.; Grünler, A.; Huguet, R.; Perez-Nadales, E.; Poli, A.; Sartorel, E.; Valiante, V.; et al. Comparative genomics of MAP kinase and calcium-calcineurin signaling components in plant and human pathogenic fungi. Fungal Genet. Biol. 2009, 46, 287–298. [Google Scholar] [CrossRef]
- Levin, D.E. Regulation of cell wall biogenesis in Saccharomyces cerevisiae: The cell wall integrity signaling pathway. Genetics 2011, 189, 1145–1175. [Google Scholar] [CrossRef]
- Mayer, A.M.; Staples, R.C. Laccase: New functions for an old enzyme. Phytochemistry 2002, 60, 551–565. [Google Scholar] [CrossRef]
- Pereira-Junior, R.A.; Huarte-Bonnet, C.; Paixão, F.R.S.; Roberts, D.W.; Luz, C.; Pedrini, N.; Fernandes, É.K.K. Riboflavin induces Metarhizium spp. to produce conidia with elevated tolerance to UV-B, and upregulates photolyases, laccases and polyketide synthases genes. J. Appl. Microbiol. 2018, 125, 159–171. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic fungi: New insights into host-pathogen interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar] [CrossRef]
- Ballester, A.R.; Marcet-Houben, M.; Levin, E.; Sela, N.; Selma-Lazaro, C.; Carmona, L.; Wisniewski, M.; Droby, S.; Gonzalez-Candelas, L.; Gabaldon, T. Genome, transcriptome, and functional analyses of Penicillium expansum provide new insights into secondary metabolism and pathogenicity. Mol. Plant Microbe Interact. 2015, 28, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Noar, R.D.; Daub, M.E. A polyketide synthase gene cluster required for pathogenicity of Pseudocercospora fijiensis on banana. PLoS ONE 2021, 16, e0258981. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).