

Differential Adaptation Has Resulted in Aggressiveness Variation of Calonectria pseudonaviculata on Hosts Buxus, Pachysandra, and Sarcococca



Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Isolates
2.2. Plant Materials for Inoculation
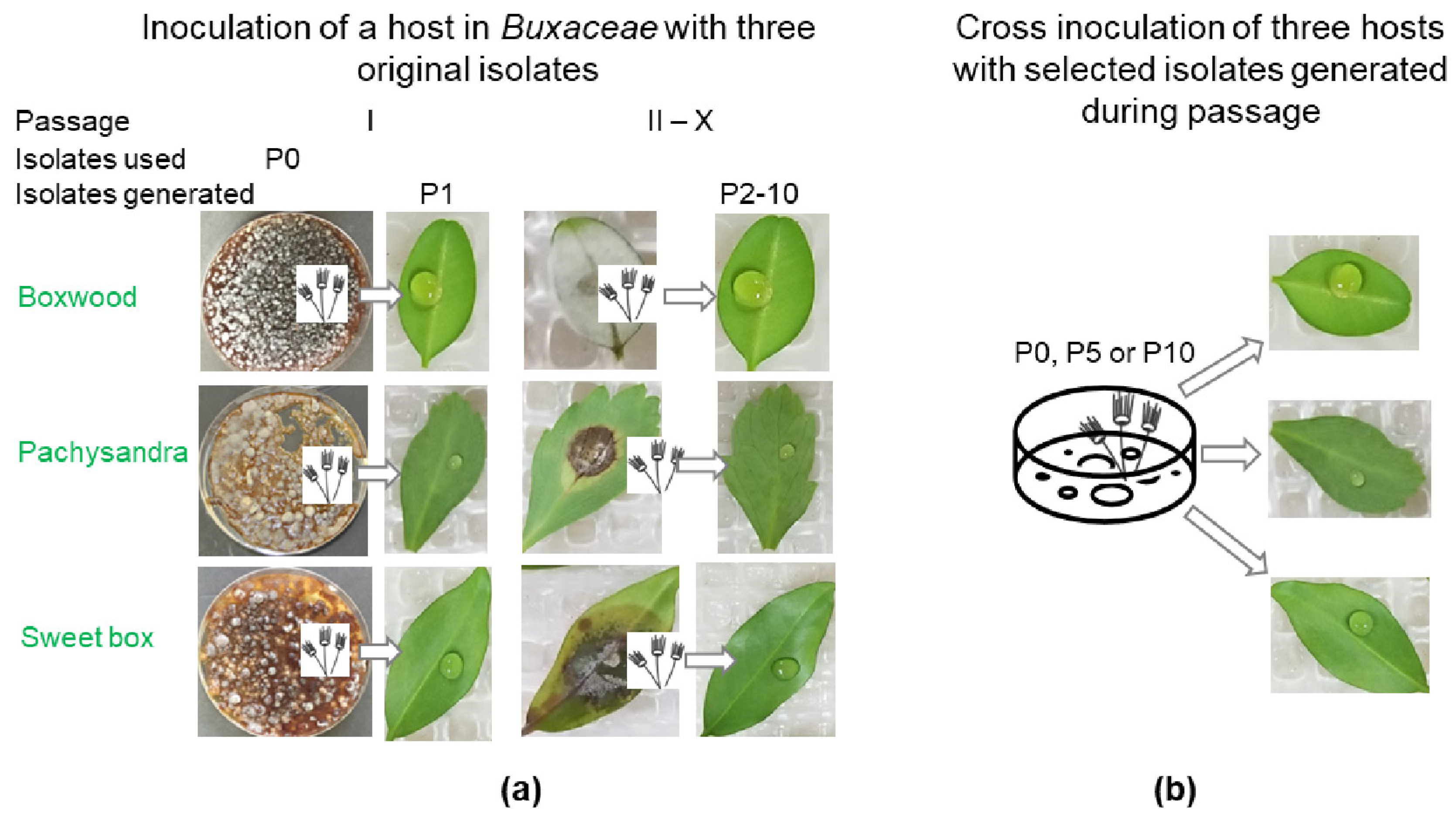
2.3. Serial Passage Experiments and Inoculum Preparation
2.4. Cps Descendant Isolation during the Passage
2.5. Comparative Study with Isolates of Different Passages
2.6. Measurement of Aggressiveness Traits
2.7. Data Analyses
3. Results
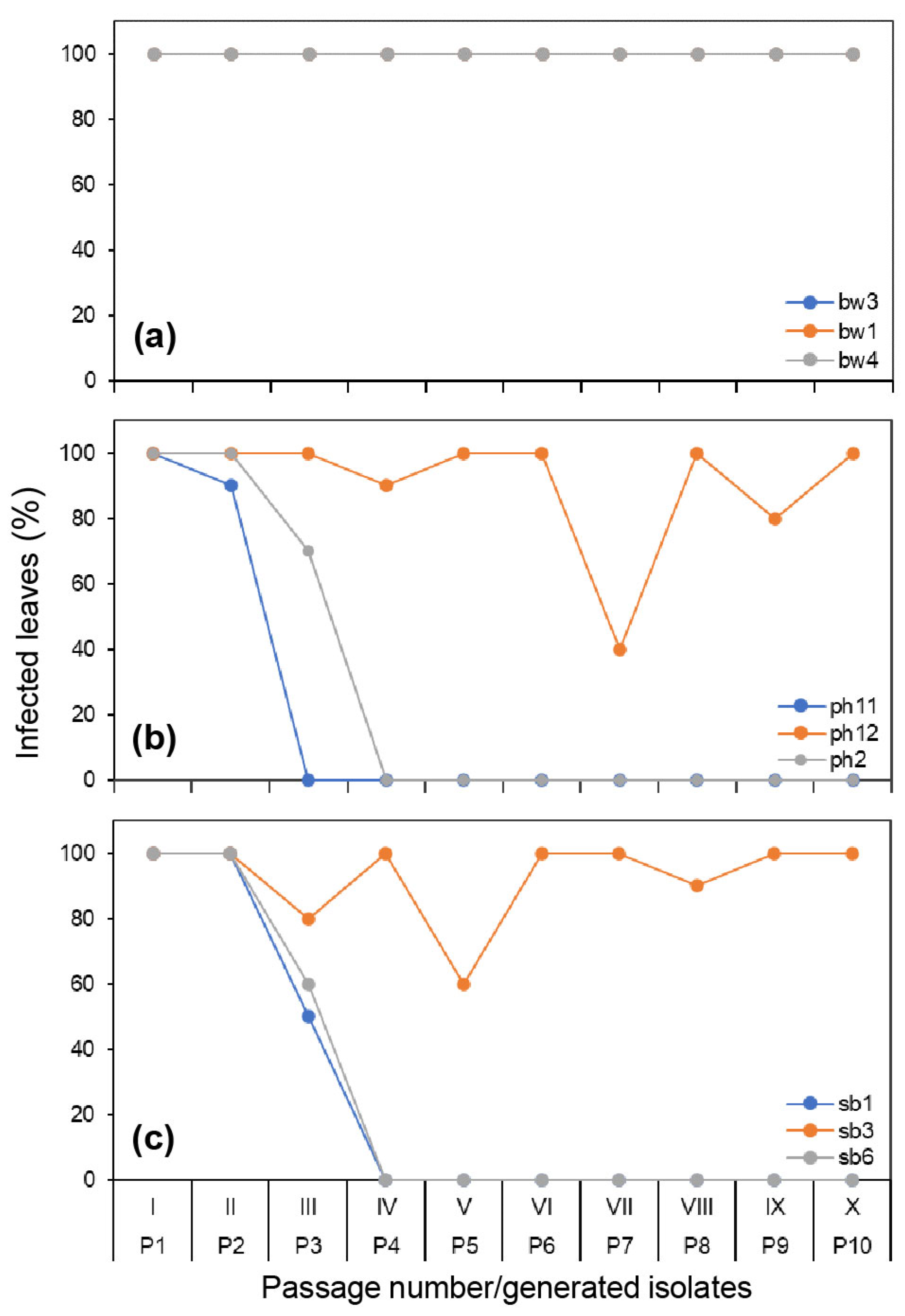
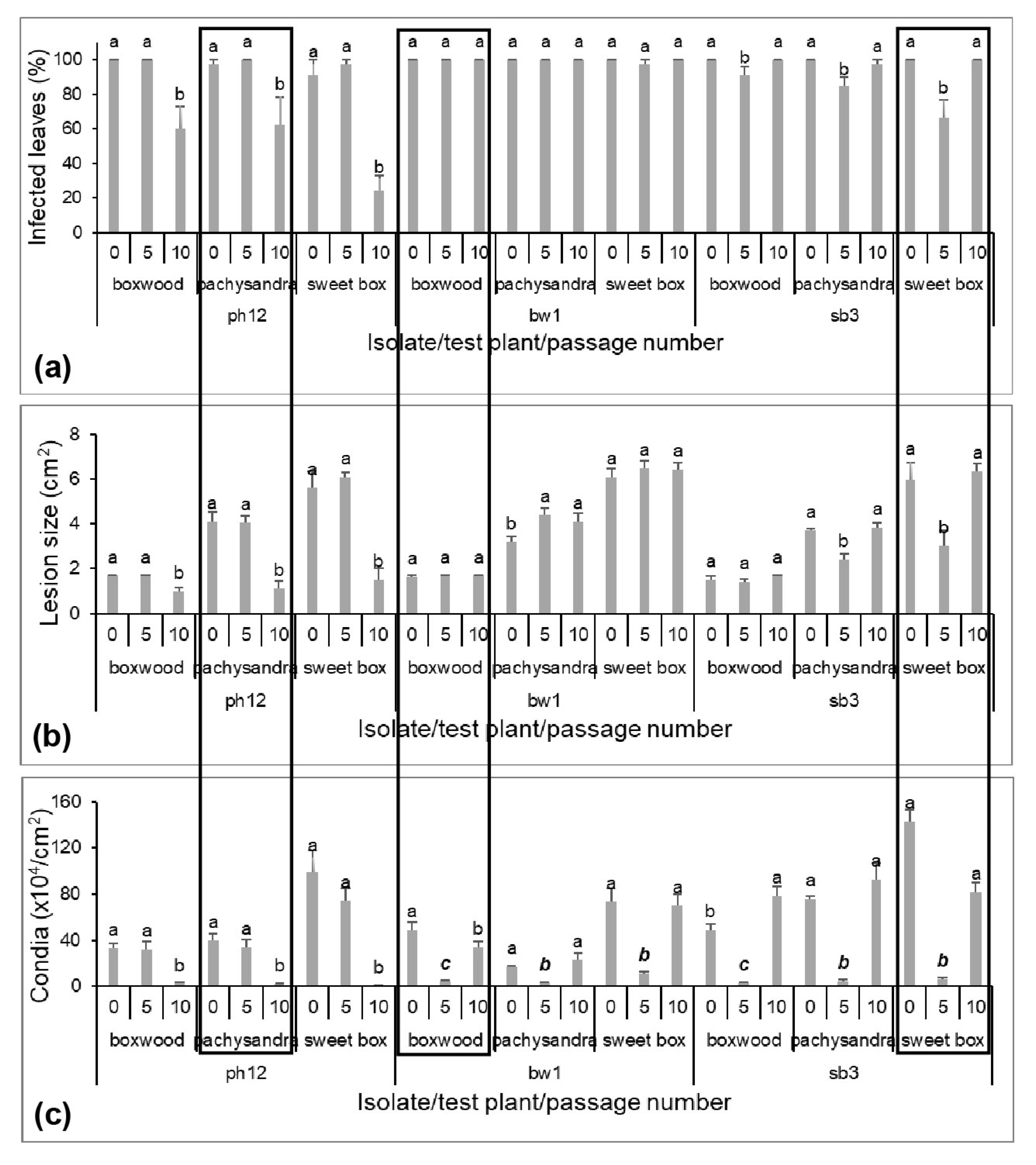
3.1. Infectivity Variation on Three Original Hosts during the Serial Passage
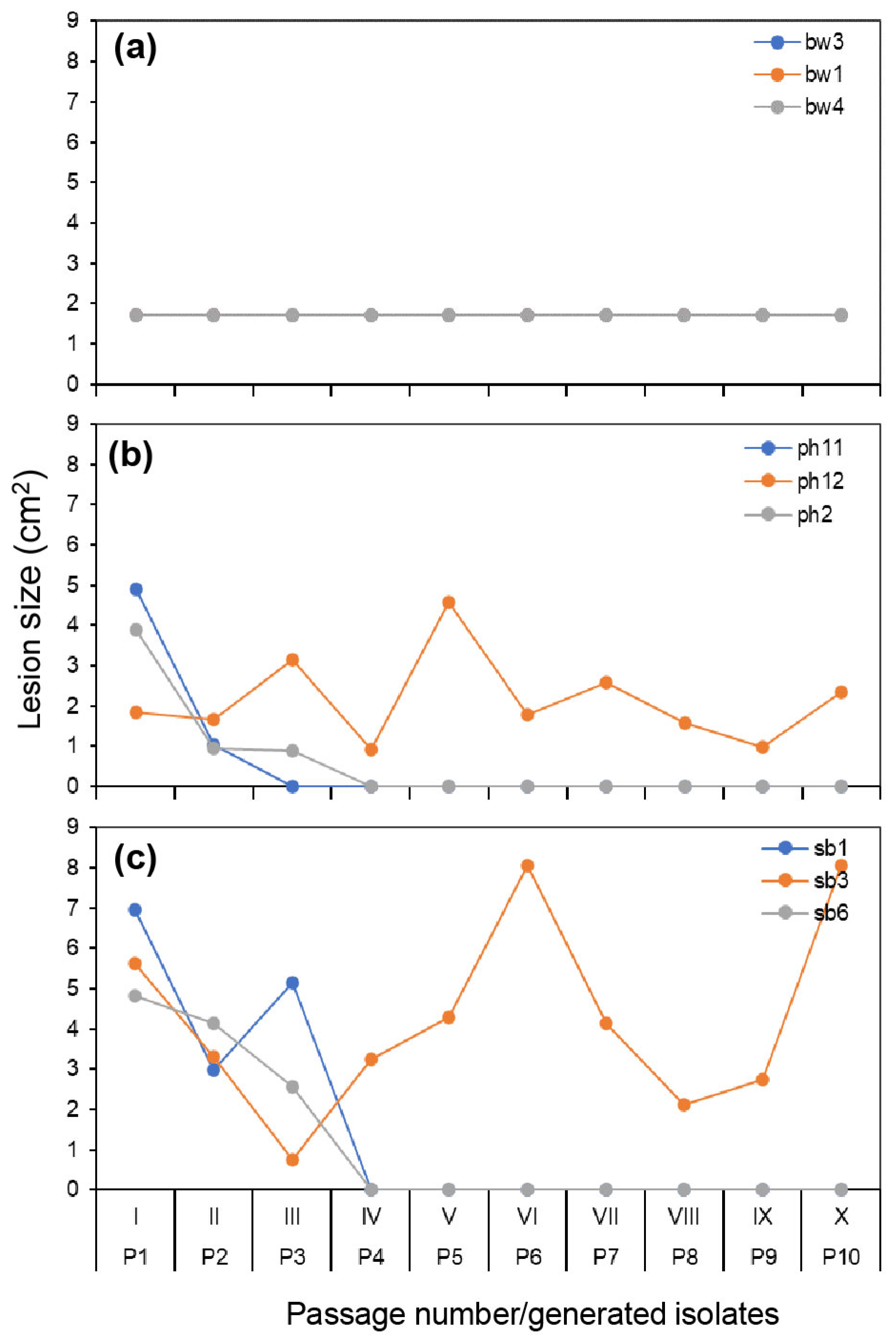
3.2. Lesion Development Variation from Original Hosts during the Serial Passage
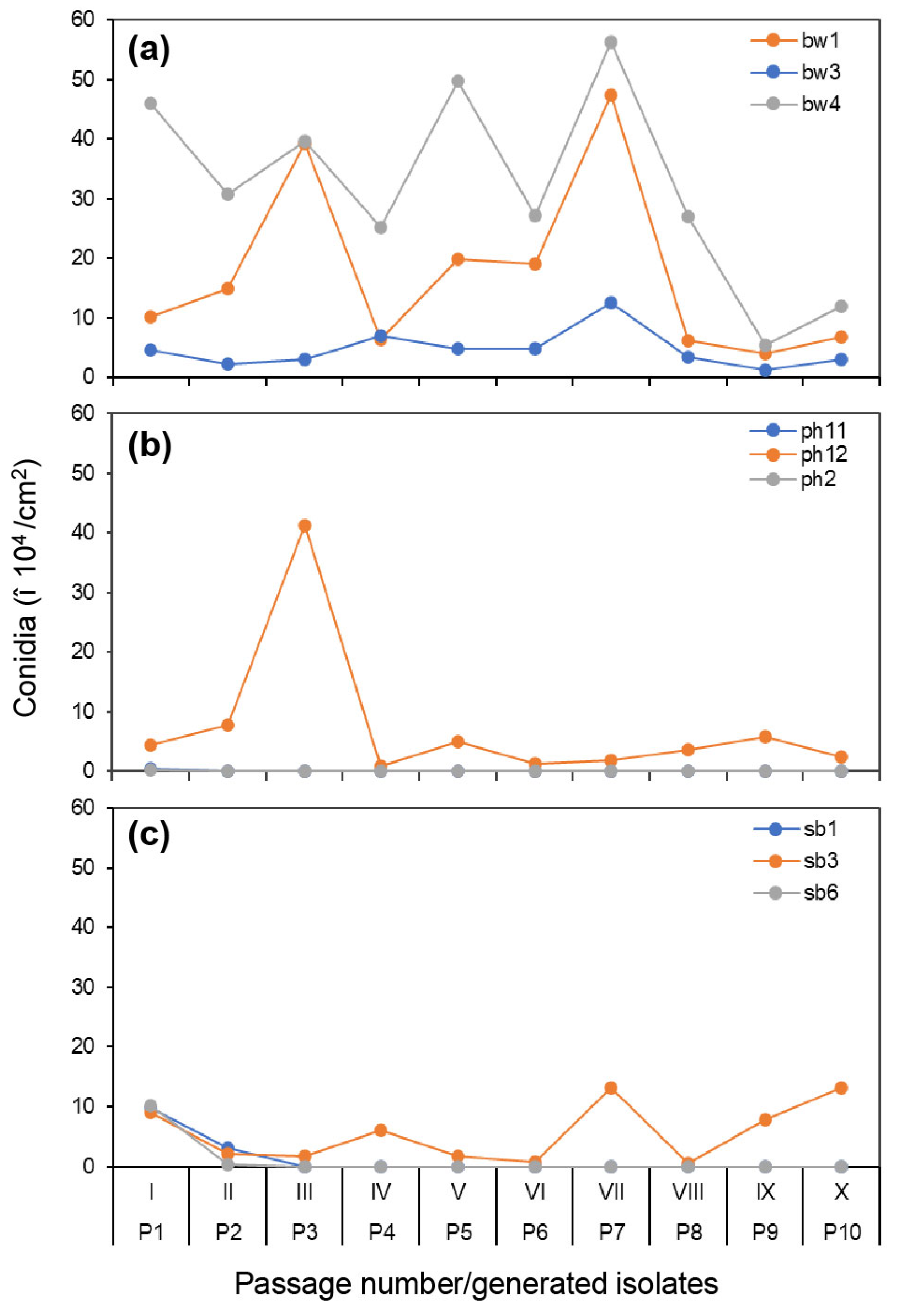
3.3. Variation in Conidia Production from Original Hosts during the Serial Passage
3.4. Responses of Original Host and Other Hosts to C. Pseudonaviculata after Passage
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- LaMondia, J.A.; Shishkoff, N. Susceptibility of boxwood accessions from the national boxwood collection to boxwood blight and potential for differences between Calonectria pseudonaviculata and C. henricotiae. Hortscience 2017, 52, 873–879. [Google Scholar] [CrossRef]
- LaMondia, J.A. Pachysandra Species and Cultivar Susceptibility to the Boxwood Blight Pathogen, Calonectria pseudonaviculata. Plant Health Prog. 2017, 18, 41–43. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Likins, M.T.; Hong, C.X. First report of blight of Sarcococca hookeriana var. humilis by Calonectria pseudonaviculata in Virginia. Plant Dis. 2016, 100, 247. [Google Scholar] [CrossRef]
- Hong, C. Fighting plant pathogens together. Science 2019, 365, 229. [Google Scholar] [CrossRef] [PubMed]
- Daughtrey, M.L. Boxwood blight: Threat to ornamentals. Ann. Rev. Phytopathol. 2019, 57, 189–209. [Google Scholar] [CrossRef]
- Brand, T.; Beltz, H.; Ehsen, B.; Adhikari, U.; Daughtrey, M.L.; Luster, D.G.; Kong, P.; Hong, C. Multi-year field plantings evaluating boxwood cultivars for susceptibility to the blight pathogens (Calonectria spp.) in northern Germany. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Avenot, H.F.; King, C.; Edwards, T.; Baudoin, A.; Hong, C.X. Effects of inoculum dose, temperature, cultivar, and interrupted leaf wetness period on infection of boxwood by Calonectria pseudonaviculata. Plant Dis. 2017, 101, 866–873. [Google Scholar] [CrossRef] [Green Version]
- Kodati, S.; Allan-Perkins, E.; Cowles, R.S.; LaMondia, J. The effect of temperature, leaf wetness period, and cultivar susceptibility on boxwood blight disease development and sporulation. Plant Dis. 2022. [Google Scholar] [CrossRef]
- Pariaud, B.; Ravigné, V.; Halkett, F.; Goyeau, H.; Carlier, J.; Lannou, C. Aggressiveness and its role in the adaptation of plant pathogens. Plant Pathol. 2009, 58, 409–424. [Google Scholar] [CrossRef]
- Delmas, C.E.; Fabre, F.; Jolivet, J.; Mazet, I.D.; Richart Cervera, S.; Delière, L.; Delmotte, F. Adaptation of a plant pathogen to partial host resistance: Selection for greater aggressiveness in grapevine downy mildew. Evol. Appl. 2016, 9, 709–725. [Google Scholar] [CrossRef]
- Panepinto, J.C.; Williamson, P.R. Intersection of fungal fitness and virulence in Cryptococcus neoformans. FEMS Yeast Res. 2006, 6, 489–498. [Google Scholar] [CrossRef] [Green Version]
- Poulin, R.; Combes, C. The Concept of Virulence: Interpretations and Implications. Parasitol. Today 1999, 15, 474–475. [Google Scholar] [CrossRef]
- Pringle, A.; Taylor, J. The fitness of filamentous fungi. Trends Microbiol. 2002, 10, 474–481. [Google Scholar] [CrossRef]
- Galvani, A.P. Epidemiology meets evolutionary ecology. Trends Ecol. Evol. 2003, 18, 132–139. [Google Scholar] [CrossRef]
- Kong, P.; Hong, C. Host responses and impact on the boxwood blight pathogen, Calonectria pseudonaviculata. Planta 2018, 249, 831–838. [Google Scholar] [CrossRef]
- Henricot, B.; Gorton, C.; Denton, G.; Denton, J. Studies on the control of Cylindrocladium buxicola using fungicides and host resistance. Plant Dis. 2008, 92, 1273–1279. [Google Scholar] [CrossRef] [Green Version]
- Ploch, S.; Choi, Y.J.; Rost, C.; Shin, H.D.; Schilling, E.; Thines, M. Evolution of diversity in Albugo is driven by high host specificity and multiple speciation events on closely related Brassicaceae. Mol. Phylogenet. Evol. 2010, 57, 812–820. [Google Scholar] [CrossRef]
- Telle, S.; Shivas, R.G.; Ryley, M.J.; Thines, M. Molecular phylogenetic analysis of Peronosclerospora (Oomycetes) reveals cryptic species and genetically distinct species parasitic to maize. Eur. J. Plant Pathol. 2011, 130, 521–528. [Google Scholar] [CrossRef]
- Rouxel, M.; Mestre, P.; Comont, G.; Lehman, B.L.; Schilder, A.; Delmotte, F. Phylogenetic and experimental evidence for host-specialized cryptic species in a biotrophic oomycete. New Phytol. 2013, 197, 251–263. [Google Scholar] [CrossRef]
- Frantzeskakis, L.; Kracher, B.; Kusch, S.; Yoshikawa-Maekawa, M.; Bauer, S.; Pedersen, C.; Spanu, P.D.; Maekawa, T.; Schulze-Lefert, P.; Panstruga, R. Signatures of host specialization and a recent transposable element burst in the dynamic one-speed genome of the fungal barley powdery mildew pathogen. BMC Genom. 2018, 19, 381. [Google Scholar] [CrossRef]
- Giraud, T.; Gladieux, P.; Gavrilets, S. Linking the emergence of fungal plant diseases with ecological speciation. Trends Ecol. Evol. 2010, 25, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, S.M.; Benson, D.M.; Crouch, J.A.; Dart, N.L.; Daughtrey, M.L.; Hong, C.; Ivors, K.L.; LaMondia, J.A.; Marra, R.E.; Palmer, C.L.; et al. Boxwood blight and the dawn of a research collaboration (Abstr.). Phytopathology 2014, 104, S147. [Google Scholar]
- Dart, N.L.; Hong, C.X. Significance of soil inoculum in the epidemiology of boxwood blight caused by Calonectria pseudonaviculata. Phytopathology 2013, 103, 33–34. [Google Scholar]
- Gehesquière, B.; Crouch, J.A.; Marra, R.E.; Van Poucke, K.; Rys, F.; Maes, M.; Gobin, B.; Höfte, M.; Heungens, K. Characterization and taxonomic reassessment of the box blight pathogen Calonectria pseudonaviculata, introducing Calonectria henricotiae sp. nov. Plant Pathol. 2016, 65, 37–52. [Google Scholar] [CrossRef]
- Castroagudín, V.L.; Yang, X.; Daughtrey, M.L.; Luster, D.G.; Pscheidt, J.W.; Weiland, J.E.; Crouch, J.A. Boxwood Blight Disease: A Diagnostic Guide. Plant Health Prog. 2020, 21, 291–300. [Google Scholar] [CrossRef]
- LeBlanc, N.; Gehesquière, B.; Salgado-Salazar, C.; Heungens, K.; Crouch, J.A. Limited genetic diversity across pathogen populations responsible for the global emergence of boxwood blight identified using SSRs. Plant Pathol. 2019, 68, 861–868. [Google Scholar] [CrossRef]
- Harrower, K.M. Specialization of Leptosphaeria nodorum to alternative graminaceous hosts. Trans. Br. Mycol. Soc. 1977, 68, 101–103. [Google Scholar] [CrossRef]
- Rufty, R.C.; Hebert, T.T.; Murphy, C.F. Variation in virulence in isolates of Septoria nodorum. Phytopathology 1981, 71, 593–596. [Google Scholar] [CrossRef]
- Graf, K.; Hube, B.; Brunke, S. Experimental Evolution of Candida by Serial Passaging in Host Cells. In Host-Fungal Interactions: Methods and Protocols; Bignell, E., Ed.; Springer: New York, NY, USA, 2021; pp. 145–154. [Google Scholar]
- Groth, J.V.; Ozmon, E.A. Contrasting effects of asexual reproduction and random mating on changes in virulence frequency in a field collection of Uromyces appendiculatus. Phytopathology 1994, 84, 566–569. [Google Scholar] [CrossRef]
- Agha, R.; Gross, A.; Rohrlack, T.; Wolinska, J. Adaptation of a chytrid parasite to its Cyanobacterial host Is hampered by host Intraspecific diversity. Front. Microbiol. 2018, 9, 921. [Google Scholar] [CrossRef] [Green Version]
- Kong, P.; Likins, T.M.; Hong, C.X. First report of Pachysandra terminalis leaf spots by Calonectria pseudonaviculata in Virginia. Plant Dis. 2017, 101, 509. [Google Scholar] [CrossRef]
- Enjalbert, J.; Duan, X.; Leconte, M.; Hovmøller, M.S.; De Vallavieille-Pope, C. Genetic evidence of local adaptation of wheat yellow rust (Puccinia striiformis f. sp. tritici) within France. Mol. Ecol. 2005, 14, 2065–2073. [Google Scholar] [CrossRef]
- Gavrilets, S. Fitness Landscapes and the Origin of Species (MPB-41); Princeton University Press: Princeton, NJ, USA, 2004. [Google Scholar]
- Johnson, P.A.; Hoppensteadt, F.C.; Smith, J.J.; Bush, G.L. Conditions for sympatric speciation: A diploid model incorporating habitat fidelity and non-habitat assortative mating. Evol. Ecol. 1996, 10, 187–205. [Google Scholar] [CrossRef]
- Henricot, B.; Culham, A. Cylindrocladium buxicola, a new species affecting Buxus spp., and its phylogenetic status. Mycologia 2002, 94, 980–997. [Google Scholar] [CrossRef]
- Parisa, K.; Saeed, R.; Mansoureh, M.; Hamidreza, Z.; Hadi, K. Genetic and phenotypic variation of Calonectria pseudonaviculata isolates causing boxwood blight disease in the Hyrcanian forest of Iran. Agric. Res. Tech. Open Access J. 2018, 19, 556081. [Google Scholar] [CrossRef] [Green Version]
- Sacristán, S.; García-Arenal, F. The evolution of virulence and pathogenicity in plant pathogen populations. Mol. Plant. Pathol. 2008, 9, 369–384. [Google Scholar] [CrossRef]
- Ebert, D. Experimental evolution of parasites. Science 1998, 282, 1432–1436. [Google Scholar] [CrossRef] [Green Version]
- Shipunov, A.; Lee, H.J.; Choi, J.; Pay, K.; DeSpiegelaire, S.; Floden, A. Not out of the box: Phylogeny of the broadly sampled Buxaceae. bioRxiv 2020. [Google Scholar] [CrossRef]
- Batdorf, L.R. Boxwood Handbook; A Practical Guide to Knowing and Growing Boxwood, 3rd ed.; Greater Valley Publications, Inc.: Winchester, VI, USA, 2005. [Google Scholar]
- Sealy, J.R. A revision of the genus Sarcococca (Buxaceae). Bot. J. 1986, 92, 117–159. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Parker, I.M. Rapid evolution in a plant-pathogen interaction and the consequences for introduced host species. Evol. Appl. 2010, 3, 144–156. [Google Scholar] [CrossRef]
- Anderson, R.M.; May, R.M. Coevolution of hosts and parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Van Der Frank, S.A. Models of parasite virulence. Q. Rev. Biol. 1996, 71, 37–78. [Google Scholar] [CrossRef] [PubMed]
- Koella, J.C.; Agnew, P. A correlated response of a parasite’s virulence and life cycle to selection on its host’s life history. J. Evol. Biol. 1999, 12, 70–79. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.E.; Lafond, T.; Müller-Graf, C.D.; Nithiuthai, S.; Brey, P.T.; Koella, J.C. Experimental evaluation of the relationship between lethal or non-lethal virulence and transmission success in malaria parasite infections. BMC Evol. Biol. 2004, 4, 30. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate Name [15] | Passage Isolate Name X | Host Plant | Date of Original Isolation |
|---|---|---|---|
| bw1 | bw1-P0 | Buxus sempervirens ‘Suffruticosa’ | September 2016 |
| bw3 | bw3-P0 | B. sempervirens ‘Suffruticosa’ | |
| bw4 | bw4-P0 | B. sempervirens ‘Suffruticosa’ | |
| p2 | ph2-P0 | Pachysandra terminalis | May 2016 |
| p11 | ph11-P0 | P. terminalis | |
| p12 | ph12-P0 | P. terminalis | |
| sb1 | sb1-P0 | Sarcococca hookeriana var. humilis | October 2015 |
| sb3 | sb3-P0 | S. hookeriana var. humilis | October 2015 |
| sb6 | sb6-P0 | S. hookeriana var. humilis | November 2016 |
| - | bw1-P5 | B. sempervirens ‘Suffruticosa’ | 18 December 2017 |
| - | bw1-P10 | B. sempervirens ‘Suffruticosa’ | 15 March 2018 |
| - | ph12-P5 | P. terminalis | 18 December 2017 |
| - | ph12-P10 | P. terminalis | 16 March 2018 |
| - | sb3-P5 | S. hookeriana var. humilis | 18 December 2017 |
| - | sb3-P10 | S. hookeriana var. humilis | 26 March 2018 |
| Variation Source | Buxus (Boxwood) | Pachysandra (Pachysandra) | Sarcococca (Sweet Box) | p-Value Among Hosts |
|---|---|---|---|---|
| (p-Value Among Isolates) | ||||
| Infection rate (%) | 100 (1) | 45.7 (0.0002) | 48.0 (0.0002) | 0.0048 |
| Lesion size (cm2) | 1.7 (1) | 1.1 (0.0171) | 2.3 (0.0120) | 0.4612 |
| Conidia (×10 k/cm2) | 18.0 (0.0003) | 2.5 (0.0191) | 2.6 (0.0104) | <0.0001 |
| Infection cycle (days) | 12.9 (1) | 12.9 (1) | 15.8 (1) | 0.1034 |
| Variation Source | Infection (%) | Lesion Size (cm2) | Conidia (104/cm2) |
|---|---|---|---|
| Passage | <0.0001 | <0.0001 | <0.0001 |
| Isolate | <0.0001 | 0.0010 | <0.0001 |
| Host | 0.0005 | <0.0001 | <0.0001 |
| Experiment | 0.0174 | 0.7007 | 0.3627 |
| Passage × Isolate | <0.0001 | <0.0001 | <0.0001 |
| Passage × Host | 0.4008 | 0.1618 | <0.0001 |
| Passage × Experiment | <0.0001 | 0.1352 | 0.1644 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, P.; Daughtrey, M.L.; Hong, C. Differential Adaptation Has Resulted in Aggressiveness Variation of Calonectria pseudonaviculata on Hosts Buxus, Pachysandra, and Sarcococca. J. Fungi 2023, 9, 181. https://doi.org/10.3390/jof9020181
Kong P, Daughtrey ML, Hong C. Differential Adaptation Has Resulted in Aggressiveness Variation of Calonectria pseudonaviculata on Hosts Buxus, Pachysandra, and Sarcococca. Journal of Fungi. 2023; 9(2):181. https://doi.org/10.3390/jof9020181
Chicago/Turabian StyleKong, Ping, Margery L. Daughtrey, and Chuanxue Hong. 2023. "Differential Adaptation Has Resulted in Aggressiveness Variation of Calonectria pseudonaviculata on Hosts Buxus, Pachysandra, and Sarcococca" Journal of Fungi 9, no. 2: 181. https://doi.org/10.3390/jof9020181
APA StyleKong, P., Daughtrey, M. L., & Hong, C. (2023). Differential Adaptation Has Resulted in Aggressiveness Variation of Calonectria pseudonaviculata on Hosts Buxus, Pachysandra, and Sarcococca. Journal of Fungi, 9(2), 181. https://doi.org/10.3390/jof9020181