
Mycotic Diseases in Chelonians


Dipartimento di Scienze Veterinarie, Università degli Studi di Pisa, 56124 Pisa, Italy
*
Author to whom correspondence should be addressed.
J. Fungi 2023, 9(5), 518; https://doi.org/10.3390/jof9050518
Submission received: 28 March 2023
/
Revised: 14 April 2023
/
Accepted: 21 April 2023
/
Published: 27 April 2023
(This article belongs to the Special Issue Fungal Diseases in Animals)
Abstract
:Turtles and ‘tortoises’ populations are declining in number, the factors driving extinction risks being related to habitat loss and degradation, climate change, introduction of invasive plant and animal species, consumption by humans for food and medicinal use, and collection for international pet trade. Fungal infections represent one of the main threats for ecosystem health. The present narrative review deals with conventional and emerging mycoses of Chelonians. Although conventional mycoses in captive and pet reptiles would depend on poor husbandry, being the agents mostly opportunistic pathogens, some fungal species were reported to occur more frequently, such as the entomopathogen Purpureocillium lilacinum. Furthermore, emerging agents such as the Fusarium solani species complex have been recognized as a real threat for the surviving of some aquatic species, acting as primary pathogens. This complex has been recently included within pathogens in One Health issues. Emydomyces testavorans is recognized as an emerging threat, although, due its recent identification, information about its epidemiology is limited. Data about treatments and outcomes of mycoses in Chelonians are also referred.
Keywords:
chelonians; mycoses; turtles; tortoises; emerging mycoses; shell mycoses; skin mycoses; deep mycoses; treatment1. Introduction
Turtles, terrapins, and tortoises belong to the order Testudines and have present on our planet for hundreds of millions of years. They are characterized by extreme longevity as well as delayed maturity and long reproductive life. Furthermore, their shell is a noteworthy adaptation mechanism, allowing for their survival in both aquatic and terrestrial environments. However, today, more than 50% of 360 ascertained living species of turtles and tortoises are categorized as endangered or vulnerable by the International Union for Conservation of Nature. In more detail, 127 species out of 360 are considered as endangered or critically endangered, while seven species and ten taxa were declared extinct in the last 280 years and three of them in the past decades. This feature shows how chelonians account among the organisms with the highest risk degree of extinction when compared to other reptiles. Moreover, many other taxa, namely, marine and freshwater turtles, could be predicted to become extinct during the present century [1,2]. The factors driving this recent, dramatic, and faster decline of chelonians, as well as their extinction risks, are thought to be related to several different factors: habitat loss; degradation and fragmentation represent a major threat for biodiversity, causing the modification of habitats for agriculture; wetland loss; and human population pressure on the landscape. Climate change contributes to the exacerbation of these factors, leading to environmental desertification; these alterations could involve lower tortoise egg production as well, while sea level rise would reduce the number of possible suitable nesting sites for marine turtles. The introduction of invasive animal species acting as food competitors and predators represents a further threat, while exotic plants—mostly grasses—along with climate change, would enhance the risk of fires. The consumption of turtles, tortoises, and turtles’ eggs for human nutrition has been another impacting threat for the last 400.000 years and could have led to the extinction of large tortoise species since 17th century. The medicinal use of turtles and tortoises is also very impactful, especially in Asia. Lastly, many chelonian species are globally collected for international illegal pet trade practices [2].
In such scenarios, infectious agents could play a capital role in the decline of these populations. Among them, fungal infections may represent a main threat to ecosystem health [3].
Infectious diseases are among the top five reasons for terrestrial species’ [4] and sea turtles’ extinction; infectious hazards can also comprehend fungal affections. Furthermore, the emergence of pathogens could be driven by environmental factors, as hypothesized for the herpes virus, which is responsible for fibropapillomatosis in sea turtles [5] or infection by Ranavirus. However, available data, mostly regarding the sanitary status of sea turtles, are fragmentary, concerning the difficult-to-retrieve diseased animals and carcasses in the wild. Thus, in a lot of cases, it is difficult to correlate morbidity rate with mortality [6].
The present narrative review aimed to gather the main information available from the literature dealing with fungal diseases of Chelonians.
Mycoses in Chelonians have different features; these animals can be affected by fungi causing shell, tegumental, and/or systemic disease, which can be considered as conventional agents. However, in recent years, new emerging fungal pathogens have affected sea and freshwater turtles and their eggs, inducing not previously reported on clinical outcomes that are able to significantly reduce turtle populations worldwide [3,7]. Microsporidiosis in these species has been recently reported on, and literature records are referred to also.
2. Mycoses of Chelonians by Conventional Agents
2.1. Conventional Mycoses
Conventional mycoses of reptiles have been deeply treated by Austwick and Keymer, 1980, Kotska et al., 1997, Jacobson et al., 2000, and Parè, 2014 [8,9,10,11]; however, further records have been added since these studies were performed. The diseases were caused by environmental fungi, belonging to a wide range of different taxa. Some reports have not been accomplished by mycologists; thus, culture isolation of causative agents has not always been carried out: for this reason, it was not possible in all described cases to identify the responsible fungal species.
2.1.1. Dermatomycoses
Turtles and tortoises can be affected by mycoses involving the skin, scale armor, and carapace. Such affections are considered as related to inadequate management, mostly due to deficiencies in housing such as inappropriate temperature and moisture [12], overcrowding, poor diet, and intercurrent bacterial and parasitic affections, [13], while fungal colonization can follow trauma. In tortoises, shell fractures and skin lesions are mostly caused by vehicles, mowers, falls, or predation by dogs, foxes, or other carnivores. Motored aquatic vehicles have been also reported to damage turtle shells. Skin injuries can be induced by intra or interspecies aggression, predators, fishing lines, or fishing nets.
Inflammation, ulceration, and perforation to abscess formation or necrosis of the underlying tissues or granulomas appear as distinctive features of dermatomycoses [13,14,15]. Being reptiles lacking in subcutaneous fat, muscles and bones may be rapidly involved [13,14], especially in chelonians [15].
Shell Mycoses
Shell mycoses have been reported to involve only the carapace or to colonize skin layers.
Most necroses of carapaces would have mycotic etiologies [16]. In tortoises, the lamellae of the carapace may become weak or loose, while small impressions can develop, leading to a fracture of the shell [9,15].
The involvement of shells alone has been mostly observed in tortoises and seldom in turtles [16]. The occurrence of yeasts (Candida sp., Cryptococcus sp. Geotrichum candidum and Rhodotorula sp.), together with molds (Absidia, Alternaria, Aspergillus spp., Cladosporium, Drechslera, Fusarium, Mucor, Penicillium, Stemphylium and Ulocladium), has been recorded [17,18]. Mucorales have also been reported to be responsible for shell mycosis in several turtles [16].
A qualitative evaluation of fungal flora involved in shell lesions of Testudo spp. was investigated in an extensive study carried out in France [19]. Fungal hyphae were observed in 73% of clarified specimens, while 34 different fungal genera were cultured. Fungi belonging to Dematiaceae were observed in about 90% of samples, with Alternaria, Cladosporium, and Aureobasidium as the most represented genera (47.2%, 41.5%, and 32.1%, respectively). Keratinophilic fungi such as Chrysosporium spp., Myriodontium keratinophilum, and Scopulariopsis brevicaulis were also identified in culture, the latter with positive microscopic features also.
Fusarium semitectum (syn. Fusarium incarnatum, Fusarium pallidoroseum) was reported to be responsible for necrotizing scute disease in Gopherus berlandieri in Texas [20] and for shell mycosis in 15 Testudo hermanni living in a community [21]. The aetiological agent was cultured from the soil. All affected subjects recovered after a specific treatment, as well as after removal from the infected environment, thus indicating the role of soil-inhabiting fungi to be responsible for shell colonization in tortoises. Fusarium solani was recently cultured by severe erosive and ulcerative lesions in the carapace of Graptemys ouachitensis, involving skin also [22].
A novel species within Onygenales, Aphanoascella galapagosensis, was isolated from the diseased carapace of a Chelonoidis nigra microphyes. The mold was morphologically described, resulting in differences from other genera within the order. These findings were corroborated by the results of the D1/D2 sequences, demonstrating that this fungal isolate represented a new lineage within Onygenales. Molecular data obtained in this work were compared with the neighbor genus Aphanoascus and other related genera also [23].
Melanin-pigmented (dematiaceous) fungi responsible for phaeohyphomycoses were also reported as a cause for shell mycoses. Exophiala oligosperma was identified by molecular techniques in a Geochelone gigantea infection, involving deep bones [24]. A further recent finding described the identification and isolation of Alternaria sp. from a single case of shell mycosis in T. hermanni) [25]. Melanized fungal elements were easily noticeable in diaphanized samples (Figure 1).
Shell mycosis due to Purpureocillium lilacinum (formerly Paecilomyces lilacinus) [26] was observed in Carettacheylis insculpta [27], and Fusarium spp. were isolated from shell lesions in C. caretta [28]. Fusarium solani was recently isolated from C. caretta with shell and skin involvement [29].
Hatchling Florida soft-shell turtles (Apalone ferox) were reported to die from mucormycosis, involving both skin and shell [30], while similar clinical outcomes caused by Trichosporon cutaneum have been described in tortoises [31]. Considering that Trichosporon jirovecii was considered a synonym of this yeast species until 1992 [32], the involvement of this latter fungal agent cannot be excluded.
Skin Mycoses
Turtles and tortoises seem to be prone to skin mycoses involving keratin layers of stratum corneum only, with no shell involvement. Mycotic granulomas, possibly caused by Aspergillus sp., were described in Sternotherus odoratus [33], while Mucor was isolated from skin lesions in Clemmys insculpta [10].
P. lilacinum was cultured and identified from the shells and skin of young Trionyx sinensis affected by white-spot disease [34].
Fusarium solani was identified as the main responsible agent for such clinical features. It was mostly identified in C. caretta [27,28,35,36,37] and in Lepidochelys kempii [38]. In some cases, the same mycotic species was isolated from the environment (sand, water) also [35,36]. The role as opportunistic pathogen of F. solani in lesions in turtles was assessed in 1999 by Cabanes [36,39]; the author demonstrated that the banding patterns yielded by an RAPD assay of clinical and environmental samples were markedly differed from those of isolates from several culture collections. Then, F. solani was recognized as a monophyletic species complex (FSSC), including more than 60 phylogenetic species [3,40,41,42]. The isolates from sea turtles have been identified as Fusarium falciforme, Fusarium keratoplasticum, and haplotypes 9 and 12 [42,43]. Furthermore, in a recent extensive study on C. caretta with superficial lesions, F. keratoplasticum, Fusarium oxysporum, Fusarium brachygibbosum, along with haplotypes 9, 12, and 27 were identified, while animals with no disease were colonized by haplotypes 9 and 12 only [27]. Similarly, F. falciforme, F. keratoplasticum, and Fusarium crassum were isolated from stranded post-hatchling C. caretta in South Africa [44].
In sea turtles, the occurrence of Fusarium spp. was suggested to be related both to the ability of these fungi to germinate within water [45] and to microclimatic factors that provoked cold stunning in turtles [37].
Regarding tortoises, T. jirovecii was identified from neck skin lesions in T. hermanni, recovering after specific antifungal treatment [46].
2.1.2. Deep and Systemic Mycoses
Deep, mostly pulmonary mycosis was reported to be more frequent in tortoises rather than in turtles [16], although further records in aquatic species are present in the literature. In these latter species, environmental fungi were described as able to colonize lesions, i.e., injury caused by the ingestion of a fishhook, as turtles are occasional scavengers. Poikilotherms are prone to adverse environmental factors. Fungal infections depend on poor cellular immunity as well as on adverse environmental conditions (overcrowding, poor water quality, drop in body temperature) or on intercurrent infections. However, the primary causes have frequently not been recognized, mostly in wildlife lacking an anamnesis, the records being made from dying subjects or carcasses.
To the best of our knowledge, the first record of generalized aspergillosis in Hydraspis hilarii dates back to 1934 [47].
Entomopathogenic fungi such as P. lilacinum, Isaria fumosorosea (formerly Paecilomyces fumosoroseus), and Beauveria bassiana were involved in deep mycoses in Aldabra tortoises, Trachemys scripta, C. caretta [48,49,50,51,52], and in systemic colonization in Eretmochelys imbricata and C. insculpta [26,53]. Beauveria bassiana, Beauveria brongniartii, Metarhizium anisopliae, Metarhizium robertsii, and a novel Metarhizium sp. were recently isolated from the lungs of C. caretta, Gopherus polyphemus, Chelonia mydas, and coelomitis in E. imbricata [54]. Paecilomyces spp. were identified as etiological agents in pneumonitis in C. mydas alone [55] or associated with Sporothrichium sp. and Cladosporium sp. [56]. The phytopathogen Colletotrichum acutatum was identified in L. kempii with signs of disseminated mycosis [57].
Penicillium griseofulvum was involved in systemic mycosis in Megalochelys (Geochelone) gigantea [58], Trichophyton sp. in Lepidochelys olivacea [59], as well as Fusarium sp. in severe pneumonia in L. kempii [60], while, presumably, Fusarium sp. and Mucor sp. were demonstrated in Emys orbicularis affected by systemic disease [61]. A disseminated phaeohyphomycosis was described in Geochelone nigra [62], the dematiacfungi Cladosporium cladosporioides and Alternaria arborescens were identified in C. caretta affected by nephritis, and Ampelomyces sp. DNA was recovered from granulomatous lesions of peritoneum of the same subject [63], as well Veronaea botryosa from the the obstructive tracheitis in 3 C. mydas [64].
Yeasts have been also identified, seeming to behave as opportunistic agents. Candida albicans was considered to be responsible for stomatitis, hyperemic or ulcerative gastritis, and enteritis in Chelonians [13,30] and was involved in severe unilateral pneumonia in Testudo graeca [65] and in C. caretta [66]. Gastrointestinal infection by Candida sp. was observed in G. gigantea [67]. Other non-albicans Candida species (mostly Candida tropicalis) were reported to be responsible for stomatitis, otitis, pneumonia, enteritis, and nephritis in tortoises and terrapins [13,30,68], while histological evidence showed that T. cutaneum was recovered from cases of stomatitis, enteritis, pneumonia, and granulomatous inflammation in multiple tissues [9], and Geotrichum candidum occurred in Geochelone elephantopus affected by dermatitis and nephritis [69]. Candida krusei was reported to be responsible for fatal, systemic mycosis in Aldabrachelys gigantea [70]. Candida palmioleophila, associated with bacteria, was recently identified in a C. mydas showing systemic disease and candidaemia [71].
Table 1 summarizes the main features of the infections described in the literature.
3. Mycoses of Chelonians by Unconventional Agents
3.1. Emerging Fungal Pathogens
In recent years, we have attempted to stop several fungal emergencies threatening several animal populations, such as amphibian chytridiomycosis, white nose syndrome in bats, snake fungal disease in wildlife, resulting in serious effects on the population level.
Similarly, sea turtle eggs fusariosis (STEF) and infections by Emydomyces testavorans have been reported as fungal emergencies in Testudines.
3.1.1. Sea Turtles’ Egg Fusariosis
This disease is reported to be responsible for low egg hatching from nests both in the wild and in hatcheries [72], mostly consisting of fungal infection by Fusarium spp., a fungal genus with worldwide distribution.
Although F. solani has been reported as present in sea turtle eggs [73,74,75], fungal identification relied on phenotypes, making it difficult to know whether the isolated molds belonged to the same species. Only recently have phylogenetic characterizations and comparisons been performed, mostly through the study of nuclear internal transcribed spacer (ITS) regions, allowing researchers to identify F. falciforme and F. keratoplasticum, both belonging to Clade III of the FSSC [3,76,77]. This complex appeared to be strongly involved in veterinary mycology [43], comprehending the etiological agents of about two thirds of all reported fusarioses [78] and acting as a true pathogen, fulfilling Koch’s postulations [76]. It represents a threat both for sea and freshwater turtles’ eggs, being possibly responsible for freshwater turtle egg fusariosis (FTEF) [79].
These fungal species have been identified in beach sand and are possibly brought by overflows from plumbing systems for human waste colonizing turtles’ nesting sites. Fusarium falciforme and F. keratoplasticum have been found in such environments [80,81]. Fusaria are environmental molds, whose taxonomy has been consistently revised, causing controversial issues. The species are grouped in complexes, some of which have been proposed as distinct genera, such as Bifusarium (for the Fusarium dimerum species complex) and Neocosmospora (for the F. solani species complex). However, they will be treated as belonging to the Fusarium genus here. Morphologically, they are characterized by banana-shaped macroconidia and are currently considered as emerging pathogens involved in plant, animal, and human infections. Fusaria are, in fact, found in the underground and aerial parts of plants on decaying vegetal materials. In particular, FSSCS are the fifth plant pathogen among the first ten responsible for human infections in hospital environments. In veterinary medicine, they are referred to as aetiological agents for different clinical presentations, mostly in aquatic animals. These fungi have recently been reviewed in a One Health perspective [82], suggesting a possible relationship between STEF and human fusarioses.
FSSCs have been mostly described in C. caretta, C. mydas, Dermochelys coriaceae, Eretmochelys imbricata, L. olivacea, L. kempi, and Natator depressus [3]. However, eggs of freshwater turtles can be affected also. Fusarium keratoplasticum, as well as other members of the FSSC, have been found in Podocnemis unifilis [79,83]. Furthermore, the invasive alien species Trachemys scripta has recently been suggested as a carrier of FSSCs in Mediterranean freshwater marshes [84].
Sea turtles’ eggs are buried in the sand and in nesting sites, mostly in slimy and silty environments [3], which have suitable conditions for the growth of these fungi, which feed on organic matter deriving from hatched and failed eggs [85]. Affected eggs can show yellow, blue, or red areas on eggshells, but more severe infections are able to develop in gray hyphal mats [76]. Hyphae can produce a network on damaged eggs, colonizing neighboring eggs also [86]. The molds produce enzymes and organic acids that are able to dissolve the shell, invading the embryos [76,87]. However, these fungi can also be present in nests with asymptomatic eggs [76], suggesting that factors other than climatic conditions would play a role in STEF [85]. FSSC members widely occur in terrestrial environments and could reach the oceans by run offs, colonizing floating particles [85]. The role of the ingestion of insoluble organic and inorganic particles and microplastics as substrates for the growth of potentially pathogenic fungi, released with feces by sea turtles, have been discussed [88,89] to elucidate the occurrence of fungal cells in nests. Furthermore, the effect of mycotoxins cannot be ruled out, as well as the protective activity of protease inhibitors produced by some turtles. Another important feature is represented by the identification of microbiomes in Fusarium-infected sea turtle eggs in protection against STEF. A preliminary study involving microbial community occurances in the nesting sites of E. imbricata in Ecuador, in order to evaluate bacteria with antifungal activity, allowed researchers to determine that Actinobacteria isolated from healthy eggshells, mostly Streptomyces, as well Amycolaptosis, Micromonospora, and Plantactinospora, showed in vitro activity against F. falciforme [90]. Despite the small sample size of the study, these findings shone a light on the microflora associated with turtle eggs and are corroborated by recent studies on symbiotic bacteria of bobtail squids (Euprymna scolopes). This bacterial community produced secondary metabolites that were able to inhibit the overgrowth of fungi and bacteria responsible for egg fouling, with marked antifungal activity against F. keratoplasticum [91,92].
3.1.2. Emydomyces testavorans Infection
Emydomyces testavorans is a member of the order Onygenales, recently isolated and characterized by ulcerative skin and shell lesions in several species of freshwater turtles from the USA [93,94]. This disease is considered a threat for the conservation of Actinemys marmorata [95] and presents similarities with Septicemic Cutaneous Ulcerative Disease, a bacterial syndrome found in captive turtles. However, E. testavorans infection differs from other shell mycoses, the general conditions of animals not being involved [95]. The lesions consist of keratin inclusion cysts, limited by keratinized squamous epithelium; hyperkeratosis, inflammation, and osteonecrosis are observed as shell lesions [96]. Affected animals show shell pitting also. Cysts can expand into celomatic cavity, compressing internal organs.
The disease is often underdiagnosed, especially when external lesions cannot be appreciated [94,97], and the etiological agent, until today, is not recognized as a true pathogen [97], unlike the other emerging Onygenales, Ophydiomyces and Nannizziopsis, in snakes and chameleons. The ecological situations and the environmental conditions of the areas in which E. testavorans infection has been reported on should be further investigated to elucidate the epidemiology of this emergent mycosis. However, seasonal variations, as observed for snake infections, cannot be ruled out [94]. Young Macrochelys temminckii can show different clinical signs such as rhinitis, paronychia, nail loss, skin ulcers, plastron ulceration, excessive shedding, and death. Shedding may persist when animal husbandry is ameliorated, and the disease becomes chronic [97].
3.1.3. Microsporidioses
Microsporidia are a group of obligate intracellular single-celled eukariotes, lacking mitochondria, recently included in the Kingdom Fungi [98]. They spread through spores and would infect a huge range of hosts, including reptiles [99]. Microsporidiosis was first reported in four tortoises (Testudo hermanni boettgeri) from Germany, with intestinal and respiratory clinical signs. About 20 subjects living together with the 3 examined tortoises showed respiratory and digestive clinical signs, and most of them died within three weeks from the onset of illness. The animals became symptomatic after hibernation and died within early summer. Histopathological findings allowed researchers to detect granulomatous and necrotizing hepatitis and/or pneumonia, containing large amounts of spores, also observed in the small intestine in one animal (Eydner et al., 2017) [100]. Bacteriological cultures reported as positive for Enterobacteriaceae and Streptococcus spp. Microsporidial organisms were identified neither at a species nor a genus level. Regardless, the number and location of filament coils allowed the researchers to exclude the occurrence of Encephalitozoon spp., described in Pogona vitticeps [101,102].
4. Treatment of Mycoses in Chelonians
Many reports deal with description of clinical and mycological pictures in wild Chelonians; sea turtles; and, except for epidemics, in cases in captive or pet subjects and stranded animals brought up at rehabilitation centers. Nevertheless, data about specific antimycotic treatments, follow ups, and outcomes are scarce. Animal species, etiological agents, clinical presentations, drug posologies, and outcomes are reported in Table 3.
5. Conclusions
Chelonians can be affected by mycoses caused both by conventional and emerging aetiologic agents.
Conventional mycoses in captive and pet reptiles would depend on poor husbandry, the agents being mostly opportunistic pathogens. Other possible causes for wild animals are stressors such as concurrent bacterial infections, parasites, wounds, ingestion of foreign bodies, exposure to environmental contaminants, or being caught in fishing nets. However, some fungal species seem to occur more frequently. Fusarium spp. are frequently involved with different clinical features, as previously reported. However, the entomopathogenic mold P. lilacinum has been consistently observed in Chelonians’ mycoses, suggesting a tropism of this organism toward reptile tissues. This fungal species has been recently recovered from unhatched eggs in T. hermanni associated with bacteria (Pseudomonas aeruginosa, Bacillus sp. and Escherichia coli) [103]. Furthermore, P. lilacinum is considered a primary pulmonary pathogen in green tree pythons [104], and the related species Purpureocillium lavendulum has been isolated from lesions in several reptile species [105].
The impact of fungal affection on turtles and tortoises’ health has not been fully understood yet, although some emerging agents such FSSCs have been recognized as real threats to the survival of some aquatic species and has been recently included within pathogens with One Health issues [82]. Furthermore, in the last few years, the role of the FSSC as a primary pathogen has been assessed. This feature has not been recognized for E. testavorans, due to its recent identification.
Future work should include a multidisciplinary approach to mycotic diseases in these animal species. In fact, data about the risk management for fungal disease hazards are limited by the scarce number of experts in this area, resulting in difficulty and, consequently, a lack of diagnostic testing in a lot of regional management units [6].
To enhance the study of etiologic agents, with particular paid attention to strain virulence, researchers should endeavor to research the pathogenesis of such infections and the sensitivities of fungal isolates versus antimycotic drugs.
Author Contributions
Conceptualization, S.N. and F.M.; methodology, S.N. and F.M.; data curation, S.N. and F.M.; writing—original draft preparation, S.N. and F.M.; writing—review and editing, S.N. and F.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rhodin, A.G.J.; Stanford, C.B.; Dijk, P.P.V.; Eisemberg, C.; Luiselli, L.; Mittermeier, R.A.; Hudson, R.; Horne, B.D.; Goode, E.V.; Kuchling, G.; et al. Global Conservation status of turtles and tortoises (Order testudines). Chelonian Conserv. Biol. 2018, 17, 135. [Google Scholar] [CrossRef]
- Stanford, C.B.; Iverson, J.B.; Rhodin, A.G.J.; Paul van Dijk, P.; Mittermeier, R.A.; Kuchling, G.; Berry, K.H.; Bertolero, A.; Bjorndal, K.A.; Blanck, T.E.G.; et al. Turtles and Tortoises Are in Trouble. Curr. Biol. 2020, 30, R721–R735. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Ramírez, J.M.; Abella-Pérez, E.; Phillott, A.D.; Sim, J.; van West, P.; Martín, M.P.; Marco, A.; Diéguez-Uribeondo, J. Global Distribution of Two Fungal Pathogens Threatening Endangered Sea Turtles. PLoS ONE 2014, 9, e85853. [Google Scholar] [CrossRef]
- Smith, K.F.; Sax, D.F.; Lafferty, K.D. Evidence for the Role of Infectious Disease in Species Extinction and Endangerment. Conserv. Biol. 2006, 20, 1349–1357. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.G. Sea Turtles: Old Viruses and New Tricks. Curr. Biol. 2004, 14, R842–R843. [Google Scholar] [CrossRef]
- Mashkour, N.; Jones, K.; Kophamel, S.; Hipolito, T.; Ahasan, S.; Walker, G.; Jakob-Hoff, R.; Whittaker, M.; Hamann, M.; Bell, I.; et al. Disease Risk Analysis in Sea Turtles: A Baseline Study to Inform Conservation Efforts. PLoS ONE 2020, 15, e0230760. [Google Scholar] [CrossRef]
- Reynolds, H.T.; Raudabaugh, D.B.; Lilje, O.; Matthew, C.; Allender, M.C.; Miller, A.M.; Gleason, F.H. Emerging mycoses and fungus-like diseases of vertebrate wildlife. In The Fungal Community: Its Organization and Role in the Ecosystem; Dighton, J., White, J.F., Eds.; CRC Taylor and Francis: Boca Raton, FL, USA, 2017; pp. 286–403. [Google Scholar]
- Austwick, P.; Keymer, I. Fungi and actinomycetes. In Diseases of the Reptilia; Cooper, J.E., Jackson, O.F., Eds.; Academic Press: New York, NY, USA, 1981; pp. 193–231. [Google Scholar]
- Kostka, V.M.; Hoffmann, L.; Balks, E.; Eskens, U.; Wimmershof, N. Review of the literature and investigations on the prevalence and consequences of yeasts in reptiles. Vet. Rec. 1997, 140, 282–287. [Google Scholar] [CrossRef]
- Jacobson, E.R.; Cheatwood, J.L.; Maxwell, L.K. Mycotic diseases of reptiles. Semin. Avian Exot. Pet Med. 2000, 9, 94–101. [Google Scholar] [CrossRef]
- Pare, J.A. Update on fungal infections in reptiles. In Current Therapy in Reptile Medicine and Surgery; Mader, D.R., Divers, S., Eds.; Elsevier Health Sciences: Saint Louis, MO, USA, 2014; pp. 53–56. [Google Scholar]
- Hatt, J.M. Dermatological diseases in reptiles. Schweizer Archiv für Tierheilkunde 2010, 152, 123–130. [Google Scholar] [CrossRef]
- Zwart, P.; Schroder, H.D. Reptilien. In Handbuch der Zootierkrankheiten; Ippen, R., Schroder, H.D., Elze, K., Eds.; Akademia Verlag: Berlin, Germany, 1985; p. 349. [Google Scholar]
- Gabrisch, K.; Zwart, P. (Eds.) Krankheiten der Heimtiere; Schlutersche Verlagsanstalt: Berlin, Germany, 1995; p. 694. [Google Scholar]
- Hafeli, W.; Schildger, B.J. Krankheiten der Zoo- und Wildtiere; Goltenboth, E., Klos, H.G., Eds.; Blackwell Wissenschaftsverlag: Berlin, Germany, 1995; p. 530. [Google Scholar]
- Hunt, T.J. Notes on Diseases and Mortality in Testudines. Herpetologica 1957, 13, 19–23. [Google Scholar]
- Bourdeau, P.; Tronco, N. Pathologie des tortues de compagnie: Bilan de consultations à Maisons-Alfort. In Proceedings of the first International Congress of Chelonian Pathology, Gonfaron, France, 25–27 April 1992; pp. 174–188. [Google Scholar]
- Bouvard, J. Contribution a L’étude des Affections Tegumentaires des Tortues Terrestres Mediterraneennes. Observations Personnelles dans le Village des Tortues de Gonfaron (France). Ph.D. Thesis, Ecole Nationale Veterinaire de Maisons-Alfort, Maisons-Alfort, France, 22 September 1992. [Google Scholar]
- Flamant, F.; de Gentile, L.; Chermette, R.; Chabasse, D.; Bouchara., J.P. Flore fongique des lésions de la carapace des tortues terrestres de compagnie dans l’Ouest de la France. J. Mycol. Méd. 2003, 13, 67–72. [Google Scholar]
- Rose, F.L.; Koke, J.; Koehn, R.; Smith, D. Identification of the etiological agent for necrotizing scute disease in the Texas tortoise. J. Wildl. Dis. 2001, 37, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Nardoni, S.; Lungonelli, P.; Papini, R.; Mugnaini, L.; Mancianti, F. Shell Mycosis in a Group of Hermann’s Tortoises (Testudo hermanni). Vet. Rec. 2012, 170, 76. [Google Scholar] [CrossRef] [PubMed]
- Brilhante-Simoes, P.; Duque, D.; Sampaio, F.; Silva, A.; Lopes, R.; Salinas, D.; Fernandes, H.; Garces, A. First Report of a Cutaneous and Shell Mycosis in an Ouachita Map Turtle (Graptemys ouachitensis, Cagle, 1953) by Fusarium solani. Veterinarska Stanica 2023, 54, 3. [Google Scholar] [CrossRef]
- Sutton, D.A.; Marín, Y.; Thompson, E.H.; Wickes, B.L.; Fu, J.; García, D.; Swinford, A.; De Maar, T.; Guarro, J. Isolation and characterization of a new fungal genus and species, Aphanoascella galapagosensis, from carapace keratitis of a Galapagos tortoise (Chelonoidis nigra microphyes). Med. Mycol. 2013, 51, 113–120. [Google Scholar] [CrossRef]
- Stringer, E.M.; Garner, M.M.; Proudfoot, J.S.; Ramer, J.C.; Bowman, M.R.; Heng, H.G.; Bradway, D.S. Phaeohyphomycosis of the carapace in an Aldabra tortoise (Geochelone gigantea). J. Zoo Wildl. Med. 2009, 40, 160–167. [Google Scholar] [CrossRef]
- Nardoni, S. (Dipartimento di Scienze Veterinarie, Università degli Studi di Pisa, Pisa, Italy). Unpublished data. 2011. [Google Scholar]
- Luangsa-ard, J.; Houbraken, J.; Van Doorn, T.; Hong, S.-B.; Borman, A.M.; Hywel-Jones, N.L.; Samson, R.A. Purpureocillium, a new genus for the medically important Paecilomyces lilacinus: Purpureocillium, a new fungal genus for P. lilacinus. FEMS Microbiol. Lett. 2011, 321, 141–149. [Google Scholar] [CrossRef]
- Lafortune, M.; Wellehan, J.F.X.; Terrell, S.P.; Jacobson, E.R.; Heard, D.; Kimbrough, J.W. Shell and systemic hyalohyphomycosis in fly river turtles, Carettochelys insculpta, caused by Paecilomyces lilacinus. J. Herpetol. Med. Surg. 2005, 15, 15–19. [Google Scholar] [CrossRef]
- Cafarchia, C.; Paradies, R.; Figueredo, L.A.; Iatta, R.; Desantis, S.; Di Bello, A.V.F.; Zizzo, N.; Van Diepeningen, A.D. Fusarium spp. in loggerhead sea turtles (Caretta caretta): From colonization to infection. Vet. Pathol. 2020, 57, 139–146. [Google Scholar] [CrossRef]
- Lai, O.; Tinelli, A.; Soloperto, S.; Marzano, G.; Tosches, M.; Leone, R.; Gelli, D.; Belloli, C.; Crescenzo, G. Fusarium solani hyalohyphomycosis in loggerhead sea turtles (Caretta caretta): A diagnostic and therapeutical challenge. Vet. Ital. 2020, 56, 123–132. [Google Scholar] [CrossRef]
- Jacobson, E.R.; Calderwood, M.B.; Clubb, S.L. Mucormycosis in hatchling Florida softshell turtles. J. Am. Vet. Med. Assoc. 1980, 177, 835–837. [Google Scholar] [PubMed]
- Schildger, B.J.; Frank, H.; Gobel, T.; Weiss, R. Mycotic infections of the integument and inner organs in reptiles. Herpetopathologia 1991, 2, 81–97. [Google Scholar]
- Guého, E.; Smith, M.T.; de Hoog, G.S.; Billon-Grand, G.; Christen, R.; Batenburg-van der Vegte, W.H. Contributions to a revision of the genus Trichosporon. Antonie Van Leeuwenhoek 1992, 6, 289–316. [Google Scholar] [CrossRef] [PubMed]
- Frye, F.L.; Dutra, F.R. Mycotic granulomata involving the forefeet of a turtle. Vet. Med. Small Anim. Clin. 1974, 69, 1554–1556. [Google Scholar] [PubMed]
- Li, X.; Zhang, C.; Fang, W.; Lin, F. White-Spot Disease of Chinese soft-shelled turtles (Trionyx sinens) caused by Paecilomyces lilacinus. J. Zhejiang Univ. Sci. B 2008, 9, 578–581. [Google Scholar] [CrossRef]
- Rebell, G. Fusarium infections in human and veterinary medicine, In Fusarium: Diseases, Biology and Taxonomy; Nelson, P.E., Tousson, T.A., Cook, R.J., Eds.; The Pennsylvania State University Press: University Park, PA, USA, 1981; pp. 210–220. [Google Scholar]
- Cabañes, F.J.; Alonso, J.M.; Castellá, G.; Alegre, F.; Domingo, M.; Pont, S. Cutaneous hyalohyphomycosis caused by Fusarium solani in a loggerhead sea turtle (Caretta caretta L.). J. Clin. Microbiol. 1997, 35, 3343–3345. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Hartmann, M.; Hennequin, C.; Catteau, S.; Béatini, C.; Blanc, V. Clusters of Fusarium solani infection in juvenile captive born Caretta caretta sea turtles. J. Mycol. Med. 2017, 27, 113–118. [Google Scholar] [CrossRef]
- Williams, S.R.; Sims, M.A.; Roth-Johnson, L.; Wickes, B. Surgical Removal of an Abscess Associated with Fusarium Solani from a Kemp’s Ridley Sea Turtle (Lepidochelys kempii). J. Zoo Wildl. Med. 2012, 43, 402–406. [Google Scholar] [CrossRef]
- Castellá, G.; Cano, J.; Guarro, J.; Cabañes, F.J. DNA Fingerprinting of Fusarium solani Isolates Related to a Cutaneous Infection in a Sea Turtle. Med. Mycol. 1999, 37, 223–226. [Google Scholar] [CrossRef]
- Zhang, N.; O’Donnell, K.; Sutton, D.A.; Nalim, F.A.; Summerbell, R.C.; Padhye, A.A.; Geiser, D.M. Members of the Fusarium solani species complex that cause infections in both humans and plants are common in the environment. J. Clin. Microbiol. 2006, 44, 2186–2190. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Rinaldi, M.G.; Sarver, B.A.J.; Balajee, S.A.; Schroers, H.-J.; Summerbell, R.C.; Robert, V.A.R.G.; Crous, P.W.; Zhang, N.; et al. Internet-accessible DNA sequence database for identifying Fusaria from human and animal infections. J. Clin. Microbiol. 2010, 48, 3708–3718. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Sutton, D.A.; Fothergill, A.; McCarthy, D.; Rinaldi, M.G.; Brandt, M.E.; Zhang, N.; Geiser, D.M. Molecular phylogenetic diversity, multilocus haplotype nomenclature, and in vitro antifungal resistance within the Fusarium solani species complex. J. Clin. Microbiol. 2008, 46, 2477–2490. [Google Scholar] [CrossRef]
- O’Donnell, K.; Sutton, D.A.; Wiederhold, N.; Robert, V.A.R.G.; Crous, P.W.; Geiser, D.M. Veterinary fusarioses within the United States. J. Clin. Microbiol. 2016, 54, 2813–2819. [Google Scholar] [CrossRef] [PubMed]
- Greeff-Laubscher, M.R.; Jacobs, K. Fusarium species isolated from post-hatchling loggerhead sea turtles (Caretta caretta) in South Africa. Sci. Rep. 2022, 12, 5874. [Google Scholar] [CrossRef]
- Wadhwani, K.; Srivastava, A.K. Fungi from otitis media of agricultural field workers. Mycopathologia 1984, 88, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Nardoni, S.; Salvadori, M.; Poli, A.; Rocchigiani, G.; Mancianti, F. Cutaneous lesions due to Trichosporon jirovecii in a tortoise (Testudo hermanni). Med. Mycol. Case Rep. 2017, 18, 18–20. [Google Scholar] [CrossRef] [PubMed]
- Hamerton, A.E. Report on the deaths occurring in the Society’s garden during the year 1933. Proc. Zoo Soc. Lond. 1934, 104, 39. [Google Scholar] [CrossRef]
- Heard, D.J.; Cantor, G.H.; Jacobson, E.R.; Purich, B.; Ajello, L.; Padhye, A.A. Hyalohyphomycosis caused by Paecilomyces lilacinus in an Aldabra tortoise. J. Am. Vet. Med. Assoc. 1986, 189, 1143–1145. [Google Scholar]
- Georg, L.K.; Williamson, W.M.; Tilden, E.B.; Getty, R.E. Mycotic pulmonary disease of captive giant tortoises due to Beauveria bassiana and Paecilomyces fumoso-roseus. Sabouraudia 1962, 2, 80. [Google Scholar] [CrossRef]
- Cabo, J.F.G.; Serrano, J.E.; Asensio, M.C.B. Mycotic pulmonary disease by Beauveria bassiana in a captive tortoise. Mycoses 1995, 38, 167–169. [Google Scholar] [CrossRef]
- Schumacher, V.L.; Mangold, B.; Lenzycki, J.; Hinckley, L.; Sutton, D.A.; Frasca, S. Occurrence of fruiting structures allows determination of Purpureocillium lilacinum as an inciting agent of pleuritis and pneumonia in a loggerhead sea turtle (Caretta caretta) by histopathologic correlation to culture. Med. Mycol. Case Rep. 2014, 6, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Arpini, C.M.; Nóbrega, Y.C.; Castheloge, V.D.; Neves, D.S.; Tadokoro, C.E.; da Costa, G.L.; Oliveira, M.M.E.; de Deus Santos, M.R. Purpureocillium lilacinum infection in captive loggerhead sea turtle hatchlings. Med. Mycol. Case Rep. 2019, 23, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Posthaus, H.; Krampe, M.; Pagan, O.; Gueho, E.; Suter, C.; Bacciarini, L. Systemic paecilomycosis in a hawksbill turtle (Eretmochelys imbricata). J. Mycol. Med. 1997, 7, 223–226. [Google Scholar]
- Horgan, M.D.; Alexander, A.B.; Innis, C.; Stacy, B.A.; Gai, J.J.; Pesavento, P.A.; Highland, M.A.; Liguori, B.L.; Norton, T.M.; Wellehan, J.F.X.; et al. Pulmonary and coelomic mycoses due to Metarhizium and Beauveria species in reptiles. J. Zoo Wildl. Med. 2022, 53, 605–612. [Google Scholar] [CrossRef]
- Glazebrook, J.S.; Campbell, R.S.F. A survey of the diseases of marine turtles in northern Australia. I Farmed turtles. Dis. Aquat. Org 1990, 9, 83–95. [Google Scholar] [CrossRef]
- Jacobson, E.R.; Gaskin, J.M.; Shields, R.P.; White, F.W. Mycotic pneumonia in mariculture-reared green sea turtles. J. Am. Vet. Med. Assoc. 1979, 175, 929–933. [Google Scholar] [PubMed]
- Manire, C.A.; Rhinehart, H.L.; Sutton, D.A.; Thompson, E.H.; Rinaldi, M.G.; Buck, J.D.; Jacobson, E. Disseminated mycotic infection caused by Colletotrichum acutatum in a Kemp’s ridley sea turtle (Lepidochelys kempi). J. Clin. Microbiol. 2002, 40, 4273–4280. [Google Scholar] [CrossRef] [PubMed]
- Orós, J.; Ramírez, A.S.; Poveda, J.B.; Rodríguez, J.L.; Fernández, A. Systemic mycosis caused by Penicillium griseofulvum in a Seychelles giant tortoise (Megalochelys gigantea). Vet. Rec. 1996, 139, 295–296. [Google Scholar] [CrossRef]
- Orós, J.; Calabuig, P.; Arencibia, A.; Camacho, M.; Jensen, H. Systemic mycosis caused by Trichophyton spp. in an olive ridley sea turtle (Lepidochelys olivacea): An immunohistochemical study. N. Z. Vet. J. 2011, 59, 92–95. [Google Scholar] [CrossRef]
- Orós, J.; Delgado, C.; Fernández, L.; Jensen, H.E. Pulmonary hyalohyphomycosis caused by Fusarium spp. in a Kemp’s ridley sea turtle (Lepidochelys kempi): An immunohistochemical study. N. Z. Vet. J. 2004, 52, 150–152. [Google Scholar] [CrossRef]
- Aleksić-Kovačević, S.; Ozvegy, J.; Krstić, N.; Rusvai, M.; Jakab, C.; Stanimirović, Z.; Becskei, Z. Skin and skeletal system lesions of european pond turtles (Emys orbicularis) from natural habitats. Acta Vet. Hung. 2014, 62, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Manharth, A.; Lemberger, K.; Mylniczenko, N.; Pinkerton, M.; Pessier, A.P.; Kammeyer, P.; De Hoog, S. Disseminated phaeohyphomycosis due to an Exophiala species in a Galapagos tortoise, Geochelone nigra. J. Herpetol. Med. Surg. 2005, 15, 20–26. [Google Scholar] [CrossRef]
- Domiciano, I.G.; Domit, C.; Trigo, C.C.; de Alcântara, B.K.; Headley, S.A.; Bracarense, A.P.F.R.L. Phaeohyphomycoses in a free-ranging loggerhead turtle (Caretta caretta) from Southern Brazil. Mycopathologia 2014, 178, 123–128. [Google Scholar] [CrossRef]
- Donnelly, K.; Waltzek, T.B.; Wellehan, J.F.X.; Sutton, D.A.; Wiederhold, N.P.; Stacy, B.A. Phaeohyphomycosis resulting in obstructive tracheitis in three green sea Turtles Chelonia mydas stranded along the Florida coast. Dis. Aquat. Organ. 2015, 113, 257–262. [Google Scholar] [CrossRef]
- Hernandez-Divers, S.J. Pulmonary candidiasis caused by Candida albicans in a Greek tortoise (Testudo graeca) and treatment with intrapulmonary amphotericin B. J. Zoo Wildl. Med. 2001, 32, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Orós, J.; Arencibia, A.; Fernández, L.; Jensen, H.E. Intestinal candidiasis in a loggerhead sea turtle (Caretta Caretta): An immunohistochemical study. Vet. J. 2004, 167, 202–207. [Google Scholar] [CrossRef]
- Juniantito, V.; Izawa, T.; Kuwamura, M.; Yonezawa, M.; Ito, S.; Yamate, J. Gastrointestinal candidiasis in an Aldabra giant tortoise (Geochelone gigantea). J. Vet. Med. Sci. 2009, 71, 1269–1272. [Google Scholar] [CrossRef]
- Zwart, P.; Buitelaar, M.N. Candida tropicalis enteric infections and their treatment in Chelonians. In Proceedings of the Annual American Association of Zoo Veterinarians, Washington DC, USA, 18–20 October 1980; pp. 58–59. [Google Scholar]
- Ruiz, J.M.; Arteaga, E.; Martinez, J.; Rubio, E.M.; Torres, J.M. Cutaneous and renal geotrichosis in a giant tortoise (Geochelone elephantopus). Sabouraudia 1980, 18, 51–59. [Google Scholar] [CrossRef]
- Iannaccone, M.; Basso, P.R.; Congiu, T.; Cavicchio, P.; Ulivi, V.; Campolo, M. Multiple Organ Dysfunction Syndrome (MODS) Induced by Candida krusei in an Aldabra giant tortoise (Aldabrachelys gigantea) and confirmed by electron microscopy analysis. Med. Mycol. Case Rep. 2018, 21, 44–48. [Google Scholar] [CrossRef]
- Wang, W.-L.; Sun, P.-L.; Kao, C.-F.; Li, W.-T.; Cheng, I.-J.; Yu, P.-H. Disseminated candidiasis and candidemia caused by Candida palmioleophila in a green sea turtle (Chelonia mydas). Animals 2021, 11, 3480. [Google Scholar] [CrossRef]
- Hoh, D.Z.; Lee, H.-H.; Wada, N.; Liu, W.-A.; Lu, M.R.; Lai, C.-K.; Ke, H.-M.; Sun, P.-F.; Tang, S.-L.; Chung, W.-H.; et al. Comparative genomic and transcriptomic analyses of trans-kingdom pathogen Fusarium solani species complex reveal degrees of compartmentalization. BMC Biol. 2022, 20, 236. [Google Scholar] [CrossRef] [PubMed]
- Phillott, A.D. Penetration of the eggshell and invasion of embryonic tissue by fungi colonizing sea turtle eggs. Herpetofauna 2004, 34, 44–47. [Google Scholar]
- Güçlü, Ö.; Bıyık, H.; Şahiner, A. Mycoflora identified from loggerhead turtle (Caretta caretta) egg shells and nest sand at Fethiye beach, Turkey. Afr. J. Microbiol. Res. 2010, 4, 408–413. [Google Scholar]
- Milena, S.C.N.; Carina, C.D.M.M.; Luciana, G.D.O. Mycobiota from the eggs, nests and stillbirths of Eretmochelys imbricata Linneus 1766 (Testudines: Cheloniidae) in Pernambuco State, Brazil. Afr. J. Microbiol. Res. 2015, 9, 1195–1199. [Google Scholar] [CrossRef]
- Sarmiento-Ramírez, J.M.; Abella, E.; Martín, M.P.; Tellería, M.T.; López-Jurado, L.F.; Marco, A.; Diéguez-Uribeondo, J. Fusarium solani is responsible for mass mortalities in nests of loggerhead sea turtle, Caretta caretta, in Boavista, Cape Verde. FEMS Microbiol. Lett. 2010, 312, 192–200. [Google Scholar] [CrossRef]
- Brofft Bailey, J.; Lamb, M.; Walker, M.; Weed, C.; Stephenson Craven, K. Detection of potential fungal pathogens Fusarium falciforme and F. keratoplasticum in unhatched loggerhead turtle eggs using a molecular approach. Endanger. Species Res. 2018, 36, 111–119. [Google Scholar] [CrossRef]
- O’Donnell, K.; Al-Hatmi, A.M.S.; Aoki, T.; Brankovics, B.; Cano-Lira, J.F.; Coleman, J.J.; de Hoog, G.S.; Di Pietro, A.; Frandsen, R.J.N.; Geiser, D.M.; et al. No to Neocosmospora: Phylogenomic and practical reasons for continued inclusion of the Fusarium solani species complex in the genus Fusarium. mSphere 2020, 5, e00810-20. [Google Scholar] [CrossRef]
- García-Martín, J.M.; Sarmiento-Ramírez, J.M.; Diéguez-Uribeondo, J. Beyond sea turtles: Fusarium keratoplasticum in eggshells of Podocnemis unifilis, a threatened Amazonian freshwater turtle. J. Fungi 2021, 7, 742. [Google Scholar] [CrossRef]
- Short, D.P.G.; O’Donnell, K.; Geiser, D.M. Clonality, recombination, and hybridization in the plumbing-inhabiting human pathogen Fusarium keratoplasticum inferred from multilocus sequence typing. BMC Evol. Biol. 2014, 14, 91. [Google Scholar] [CrossRef]
- Short, D.P.G.; O’Donnell, K.; Thrane, U.; Nielsen, K.F.; Zhang, N.; Juba, J.H.; Geiser, D.M. Phylogenetic relationships among members of the Fusarium solani species complex in human infections and the descriptions of F. keratoplasticum sp. nov. and F. petroliphilum stat. nov. Fungal Genet. Biol. 2013, 53, 59–70. [Google Scholar] [CrossRef]
- Sáenz, V.; Alvarez-Moreno, C.; Pape, P.L.; Restrepo, S.; Guarro, J.; Ramírez, A.M.C. A One health perspective to recognize Fusarium as important in clinical practice. J. Fungi 2020, 6, 235. [Google Scholar] [CrossRef] [PubMed]
- Carranco, A.S.; Gillingham, M.A.F.; Wilhelm, K.; Torres, M.d.L.; Sommer, S.; Romo, D. Transcending sea turtles: First report of hatching failure in eggs of an Amazonian freshwater turtle with symptoms of the fungal emerging disease fusariosis. Transbound Emerg. Dis. 2022, 69, e3282–e3288. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ríos, M.; Martín-Torrijos, L.; Diéguez-Uribeondo, J. The invasive alien Red-eared slider turtle, Trachemys scripta, as a carrier of STEF-disease pathogens. Fungal Biol. 2022, 126, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Gleason, F.H.; Allerstorfer, M.; Lilje, O. Newly emerging diseases of marine turtles, especially sea turtle egg fusariosis (SEFT), caused by species in the Fusarium solani complex (FSSC). Mycology 2020, 11, 184–194. [Google Scholar] [CrossRef]
- Phillott, A.D.; Parmenter, C.J. The distribution of failed eggs and the appearance of fungi in artificial nests of green (Chelonia mydas) and loggerhead (Caretta caretta) sea turtles. Aust. J. Zool 2001, 49, 713. [Google Scholar] [CrossRef]
- Phillott, A.D.; Parmenter, C.J.; McKillup, S.C. Calcium dof eggshell after fungal invasion of sea turtle eggs. Chelonian Conserv. Biol. 2006, 5, 146. [Google Scholar] [CrossRef]
- Das, M.P.; Kumar, S. Microbial deterioration of low density polyethylene by Aspergillus and Fusarium sp. Int. J. ChemTech Res. 2014, 6, 299–305. [Google Scholar]
- Oberbeckmann, S.; Löder, M.G.J.; Labrenz, M. Marine microplastic-associated biofilms—A review. Environ. Chem. 2015, 12, 551. [Google Scholar] [CrossRef]
- Sarmiento-Ramírez, J.M.; van der Voort, M.; Raaijmakers, J.M.; Diéguez-Uribeondo, J. Unravelling the microbiome of eggs of the endangered sea turtle Eretmochelys imbricata identifies bacteria with activity against the emerging pathogen Fusarium falciforme. PLoS ONE 2014, 9, e95206. [Google Scholar] [CrossRef]
- Kerwin, A.H.; Nyholm, S.V. Reproductive system symbiotic bacteria are conserved between two distinct populations of Euprymna scolopes from Oahu, Hawaii. mSphere 2018, 3, e00531-17. [Google Scholar] [CrossRef]
- Suria, A.M.; Tan, K.C.; Kerwin, A.H.; Gitzel, L.; Abini-Agbomson, L.; Bertenshaw, J.M.; Sewell, J.; Nyholm, S.V.; Balunas, M.J. Hawaiian Bobtail squid symbionts inhibit marine bacteria via production of specialized metabolites, including new bromoalterochromides BAC-D/D’. mSphere 2020, 5, e00166-20. [Google Scholar] [CrossRef] [PubMed]
- Woodburn, D.B.; Miller, A.N.; Allender, M.C.; Maddox, C.W.; Terio, K.A. Emydomyces testavorans, a new genus and species of Onygenalean fungus isolated from shell lesions of freshwater aquatic turtles. J. Clin. Microbiol. 2019, 57, e00628-18. [Google Scholar] [CrossRef] [PubMed]
- Lambert, M.R.; Hernández-Gómez, O.; Krohn, A.R.; Mutlow, A.; Patterson, L.; Rosenblum, E.B.; Timmer, M.; Willis, J.; Bushell, J. Turtle shell disease fungus (Emydomyces testavorans): First documented occurrence in California and prevalence in free-living turtles. Ichthyol. Herpetol. 2021, 109, 958–962. [Google Scholar] [CrossRef]
- Haman, K.; Hallock, L.; Schmidt, T.; Holman, E.; Murphie, B. Shell disease in Northwestern Pond Turtles (Actinemys marmorata) in Washington State, USA. Herpetol. Rev. 2019, 50, 495–502. [Google Scholar]
- Woodburn, D.B.; Kinsel, M.J.; Poll, C.P.; Langan, J.N.; Haman, K.; Gamble, K.C.; Maddox, C.; Jeon, A.B.; Wellehan, J.F.X.; Ossiboff, R.J.; et al. Shell lesions associated with Emydomyces testavorans infection in freshwater aquatic turtles. Vet. Pathol. 2021, 58, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Adamovicz, L.; Allender, M.C.; Gibbons, P.M. Emerging infectious diseases of chelonians: An update. Vet. Clin. N. Am. Exot. Anim. Pract. 2020, 23, 263–283. [Google Scholar] [CrossRef]
- Lee, S.C.; Corradi, N.; Byrnes, E.J.; Torres-Martinez, S.; Dietrich, F.S.; Keeling, P.J.; Heitman, J. Microsporidia Evolved from Ancestral Sexual Fungi. Curr. Biol. 2008, 18, 1675–1679. [Google Scholar] [CrossRef]
- Vergneau-Grosset, C.; Larrat, S. Microsporidiosis in Vertebrate Companion Exotic Animals. J. Fungi 2015, 2, 3. [Google Scholar] [CrossRef]
- Eydner, M.; Donhauser, J.; Beineke, A.; Guenther, P.; Blahak, S. Microsporidiosis in Four Tortoises (Testudo hermanni Boettgeri). Vet. Pathol. 2017, 54, 704–709. [Google Scholar] [CrossRef]
- Shibasaki, K.; Tokiwa, T.; Sukegawa, A.; Kondo, H.; Tamukai, K.; Haga, Y.; Ike, K. First Report of Fatal Disseminated Microsporidiosis in Two Inland Bearded Dragons Pogona Vitticeps in Japan. JMM Case Rep. 2017, 4, e005089. [Google Scholar] [CrossRef]
- Wünschmann, A.; Armién, A.G.; Childress, A.L.; Wellehan, J.F.X.; Giannitti, F. Intrapericardial Encephalitozoon Pogonae-Associated Arteritis with Fatal Hemopericardium in Two Juvenile Central Bearded Dragons. J. Vet. Diagn. Investig. 2019, 31, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Dovč, A.; Stvarnik, M.; Lindtner Knific, R.; Gregurić Gračner, G.; Klobučar, I.; Zorman Rojs, O. Monitoring of unhatched eggs in Hermann’s Tortoise (Testudo hermanni) after artificial incubation and possible improvements in hatching. Animals 2021, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.; Loncaric, I.; Richter, B.; Spergser, J. Fatal Purpureocillium lilacinum pneumonia in a green tree python. J. Vet. Diagn. Investig 2018, 30, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, V.; Klasen, L.; Schneider, J.; Hübel, J.; Cramer, K. Pulmonary fungal granulomas and fibrinous pneumonia caused by different hypocrealean fungi in reptiles. Vet. Microbiol. 2018, 225, 58–63. [Google Scholar] [CrossRef]
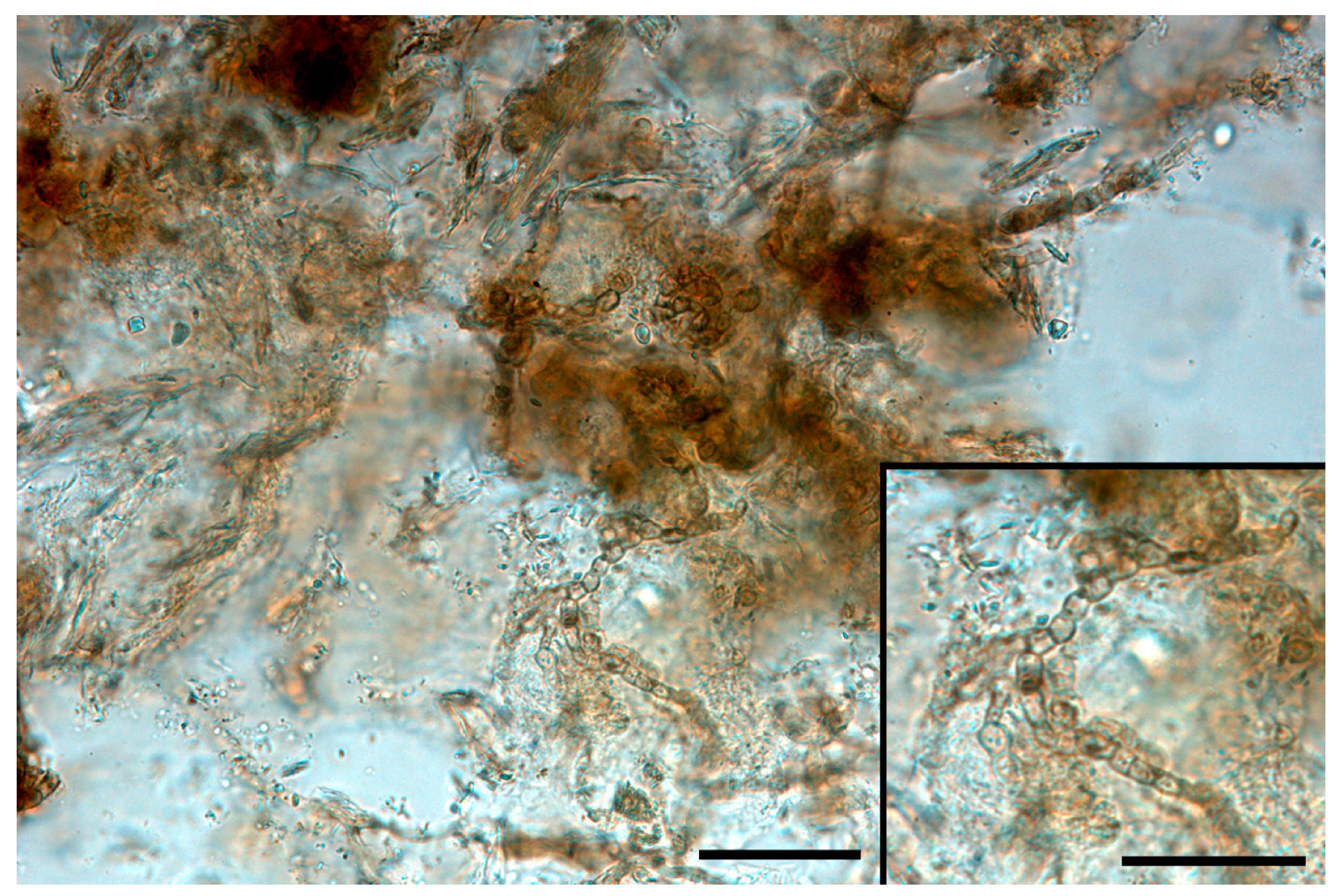
Figure 1.
Melanized short hyphae in a diaphanized sample from Testudo hermanni carapace (bar = 200 μm). Insert: detail of fungal elements (bar = 100 μm).
Figure 1.
Melanized short hyphae in a diaphanized sample from Testudo hermanni carapace (bar = 200 μm). Insert: detail of fungal elements (bar = 100 μm).

{kind=link}
Table 1.
Main fungal diseases, with epidemiological patterns, transmission routes, pathogenetic features, and prevention strategies.
Table 1.
Main fungal diseases, with epidemiological patterns, transmission routes, pathogenetic features, and prevention strategies.
| Species | Etiologic Agent | Epidemiological Pattern * | Transmission Route § | Pathogenetic Features | Lesions | Prevention Strategies | Ref. |
|---|---|---|---|---|---|---|---|
| Gopherus berlendieri | Fusarium semitectum | Group infection © | Environment | fungal keratinases | superficial | [20] | |
| Testudo hermanni | Fusarium semitectum | Group infection © | Environment | fungal keratinases | superficial | moving | [21] |
| Graptemys ouachitensis | Fusarium solani | Single case © | fungal keratinases and poor management | superficial | improved management practices | [22] | |
| Chelonoidis nigra | Aphanoascella galapagosensis | Single case and group © | superficial | none | [23] | ||
| Geochelone gigantea | Exophiala oligosperma | Single case © | superficial | none | [24] | ||
| Testudo hermanni | Alternaria sp. | Single case © | superficial | none | [25] | ||
| Carettocheylis insculpta | Purpureocillium lilacinum | Group infection (W) | immunosuppression, stress of shipment, poor management | superficial | water changes and good management practices | [26] | |
| Caretta caretta | Fusarium spp | Group infection © (R) | Environment | stressors, poor management | superficial pneumonia | improved management practices | [27] |
| Caretta caretta | Fusarium solani | Group infection © (R) | cold stunning | superficial | [28] | ||
| Apalone ferox | Mucorales | Group infection © | mixed bacterial infection | superficial | [29] | ||
| Trionyx sinensis | Purpureocillium lilacinum | Group infection © | poor management | superficial | improved management practices | [34] | |
| Caretta caretta | Fusarium solani | Single case © (R) | Environment | immunosuppression for trauma, surgery, rehabilitation | superficial | improved management practices | [36] |
| Caretta caretta | Fusarium solani | Group infection © | Environment through wound | fungal keratinases, stress for stranding, and poor management | superficial | improved management practices | [37] |
| Lepidochelys kempii | Fusarium solani | Single case © (R) | Through wound | cold stunning, mixed bacterial infection | superficial | [38] | |
| Caretta caretta | FSSC | Group infection (W) (R) | stranded | superficial | preventing pathogen introduction and diffusion | [44] | |
| T. hermanni | Trichosporon jirovecii | Single case © | Through wound | Infection of a skin lesion | superficial | preventing infections of open wounds | [46] |
| Caretta caretta | Purpureocillium lilacinum | Single case © | Water | poor management, introduction of infected hosts | pneumonia | improved management practices | [51] |
| Caretta caretta | Purpureocillium lilacinum | Group infection © | captive stress, nutritional deficits, poor management | superficial pneumonia | improved management practices | [52] | |
| Lepidochelys kempii | Colletotrichum acutatum | Single case © (R) | Cold stunning | [57] | |||
| Megalocheylis gigantea | Penicillium griseofulvum | Single case © | stress consequent to burns | systemic | [58] | ||
| Chelonia mydas | Veronaea botryosa | Group infection (W) | Respiratory | stranded animals | obstructive tracheitis | [64] | |
| Chelonia mydas | Candida palmioleophila | Single case © R | shell fracture | systemic | [71] |
Legend: * © captive animals; (W) wild animals; (R) animals introduced in a rehabilitation center. § All the listed fungi are environmental inhabitants, but the captions refer to confirmed occurrences of agents in the environment.
Table 2.
Fusarioses of Chelonians.
| Animal Species | Agent | Country | Disease | Reference |
|---|---|---|---|---|
| Gopherus berlandieri | Fusarium semitectum | Texas | Necrotizing scute disease | [20] |
| Testudo hermani | F. semitectum | Italy | Shell mycosis | [21] |
| Graptemys ouachitensis | Fusarium solani | Portugal | Shell mycosis | [57] |
| Caretta caretta | FSSC * | Italy | Skin and shell | [27] |
| Caretta caretta | F. solani | Italy | Skin, shell and bone | [28] |
| Caretta caretta | F. solani | Spain | Skin | [36] |
| Caretta caretta | F. solani | France | Skin and systemic | [37] |
| Caretta caretta | FSSC ** | South Africa | Skin | [44] |
| Lepidochels kempii | F. solani | USA | Abscess | [38] |
| Emys orbicularis | Fusarium sp. and Mucor | Serbia | Systemic | [66] |
| Caretta caretta | F. solani | Cape Verde | STEF | [76] |
| Chelonia mydas | FSSC | Australia, Ecuador | STEF | [3] |
| Eretmochelys imbricata | FSSC | Ecuador | STEF | [3] |
| Lepidochelys olivacea | FSSC | Ecuador | STEF | [3] |
| Dermochelys coriacea | FSSC | Colombia, Costa Rica | STEF | [3] |
| Natator depressus | FSSC | Australia | STEF | [3] |
| Podocnemis unifilis | FSSC | Ecuador | FTEF | [79] |
| Podocnemis unifilis | FSSC | Ecuador | FTEF | [83] |
Legend: FSSC: Fusarium solani species complex; STEF: sea turtle egg fusariosis; FTEF: freshwater turtles. * F. keratoplasticum, Fusarium oxysporum, Fusarium brachygibbosum, and haplotypes 9, 12, and 27. ** F. falciforme, F. keratoplasticum, and Fusarium crassum.
Table 3.
Mycoses of chelonians submitted to specific antimycotic treatments.
| Animal Species | Agent | Disease | Antimycotic Drugs | Outcome | Reference |
|---|---|---|---|---|---|
| Aldabrachelys gigantea | P. lilacinum | systemic | Ketoconazole 10 mg/kg PO | death | [48] |
| Aldabrachelys gigantea | C. krusei | systemic | Itraconazole 5 mg/kg/die PO | death | [70] |
| Lepidochelys kempi | Co. acutatum | systemic | Fluconazole 0.75 mg/kg SC/48 h, then Itraconazole 5 mg/kg/die PO | death | [57] |
| Caretta caretta | F. solani | shell | Topical 10% iodine in alcohol and topical ketoconazole | recovery | [36] |
| Caretta caretta | F. solani | Skin and systemic | Posaconazole 0.2 mg/kg/48 h PO | recovery by 33% | [37] |
| Testudo graeca | C. albicans | Pneumonia | Amphotericin B 0.1 mg/kg/die intrapulmonary | recovery | [65] |
| A. marmarota | E. testavorans | Skin shell | Topical iodine, terbinafine | recovery | [97] |
| Carettochelys insculpta | P. lilacinum | shell | Malachite green and formaldehyde dips, itraconazole 10 mg/kg/48 h PO | recovery | [25] |
| T. hermanni | F. semitectum | shell | Povidone iodine and Iruxol ointment/daily | recovery | [21] |
| T. hermanni | T. jirovecii | skin | Povidone iodine and Iruxol ointment/daily | recovery | [45] |
| T. hermanni | Alternaria sp | shell | Topical 10% iodine in alcohol | recovery | [24] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Nardoni, S.; Mancianti, F. Mycotic Diseases in Chelonians. J. Fungi 2023, 9, 518. https://doi.org/10.3390/jof9050518
AMA Style
Nardoni S, Mancianti F. Mycotic Diseases in Chelonians. Journal of Fungi. 2023; 9(5):518. https://doi.org/10.3390/jof9050518
Chicago/Turabian StyleNardoni, Simona, and Francesca Mancianti. 2023. "Mycotic Diseases in Chelonians" Journal of Fungi 9, no. 5: 518. https://doi.org/10.3390/jof9050518
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.