

Functional Significance of miR-4693-5p in Targeting HIF1α and Its Link to Rheumatoid Arthritis Pathogenesis


(This article belongs to the Section Small Non-Coding RNA)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
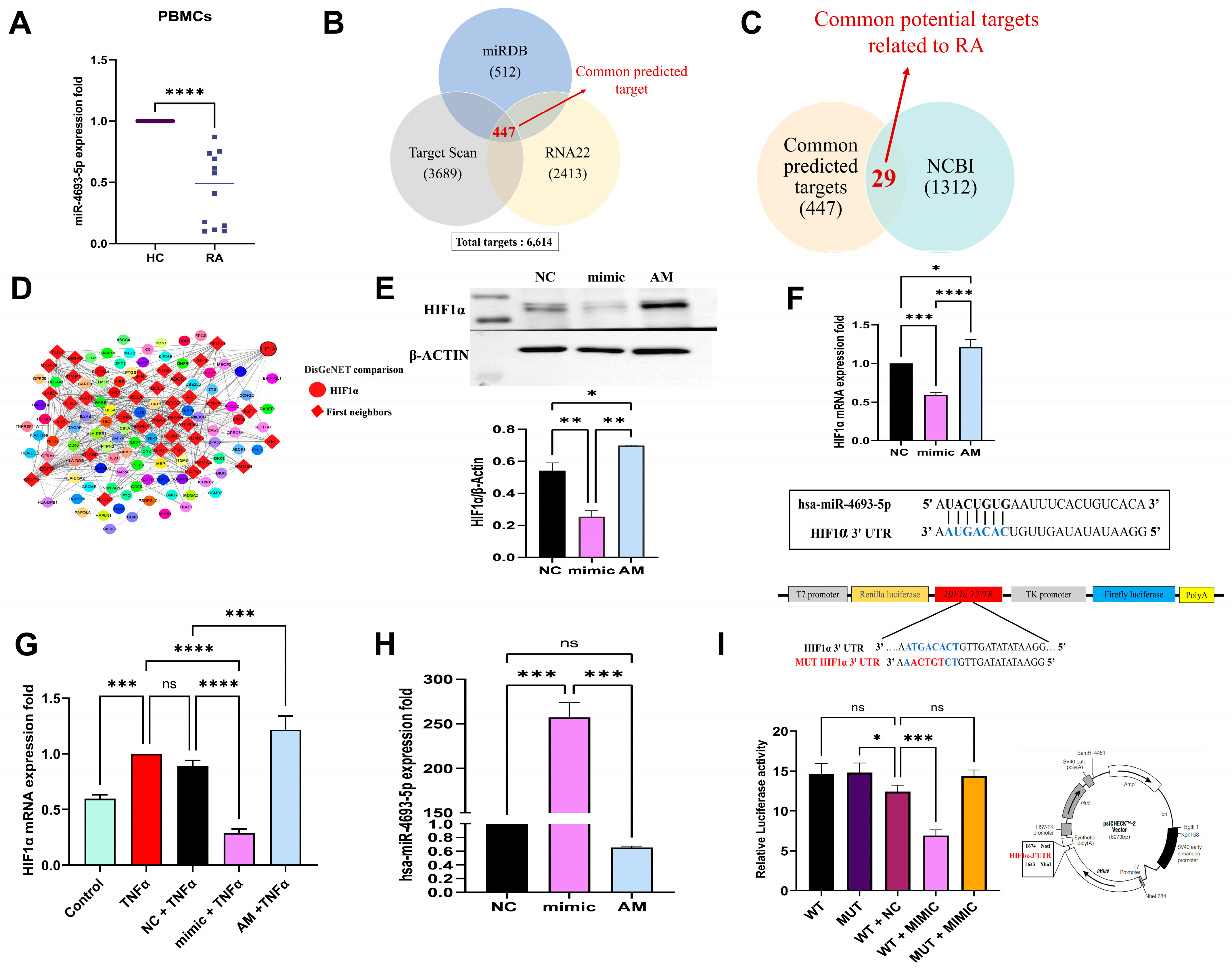
2.1. miR-4693-5p Expression Decreased in RA PBMCs
2.2. In Silico Target Prediction of miR-4693-5p
2.3. miR-4693-5p Directly Targets HIF1α
2.4. miR-4693-5p Binds to 3′ UTR of HIF1α
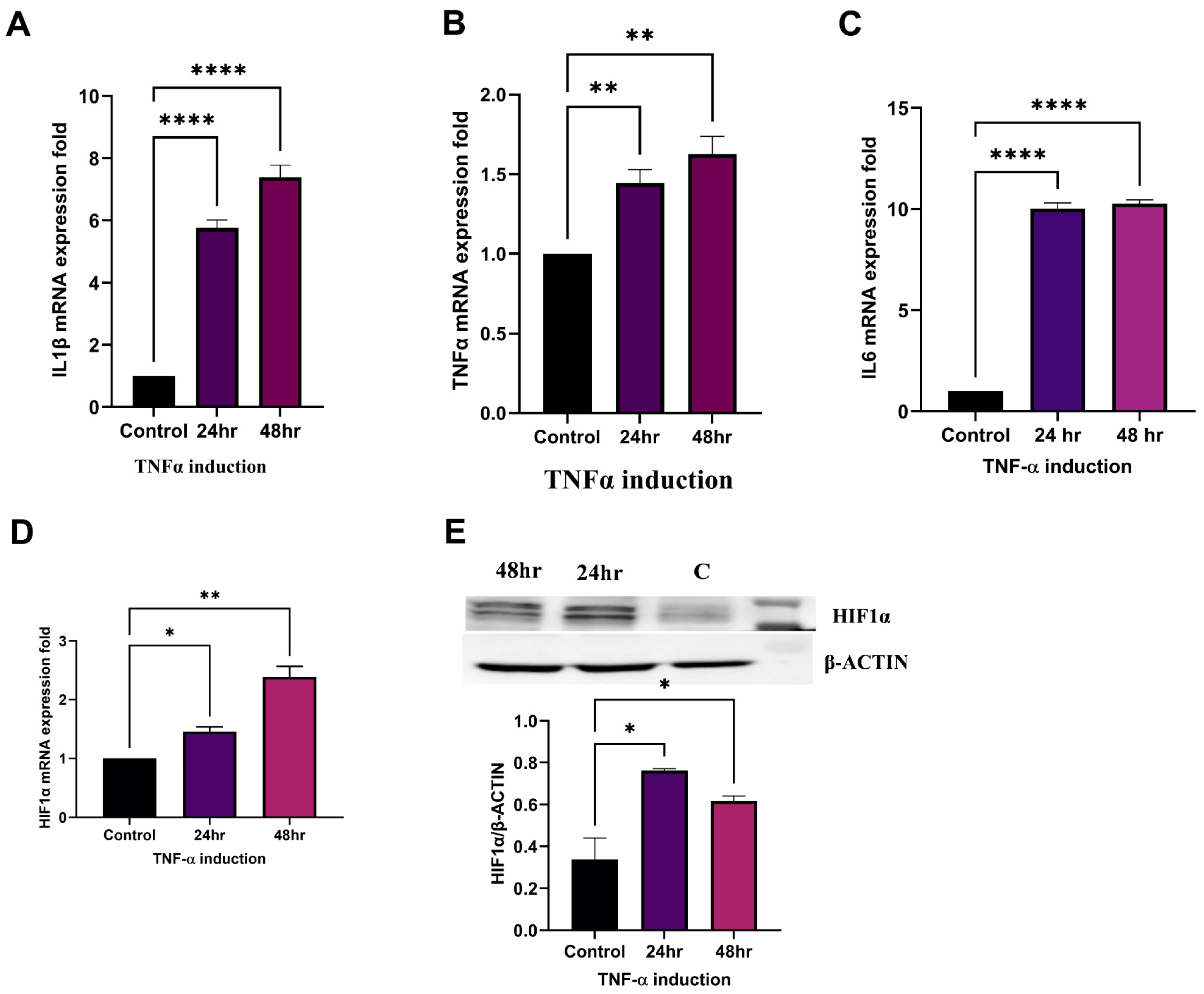
2.5. TNFα Induces HIF1α Expression and Establishment of RA
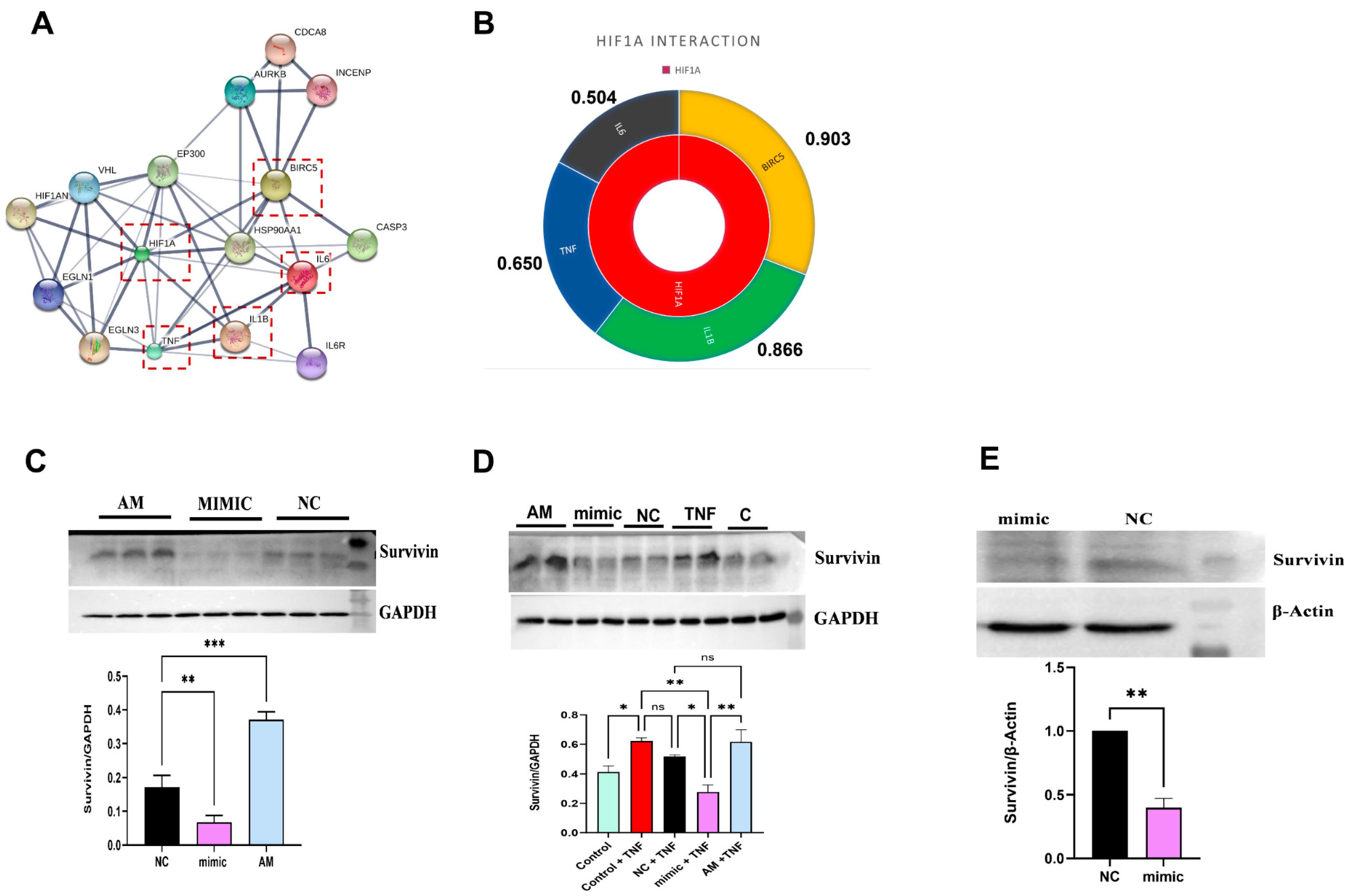
2.6. In Silico Target Interaction of HIF1α
2.7. miR-4693-5p Reduced the Expression of Survivin, a Downstream Target of HIF1α
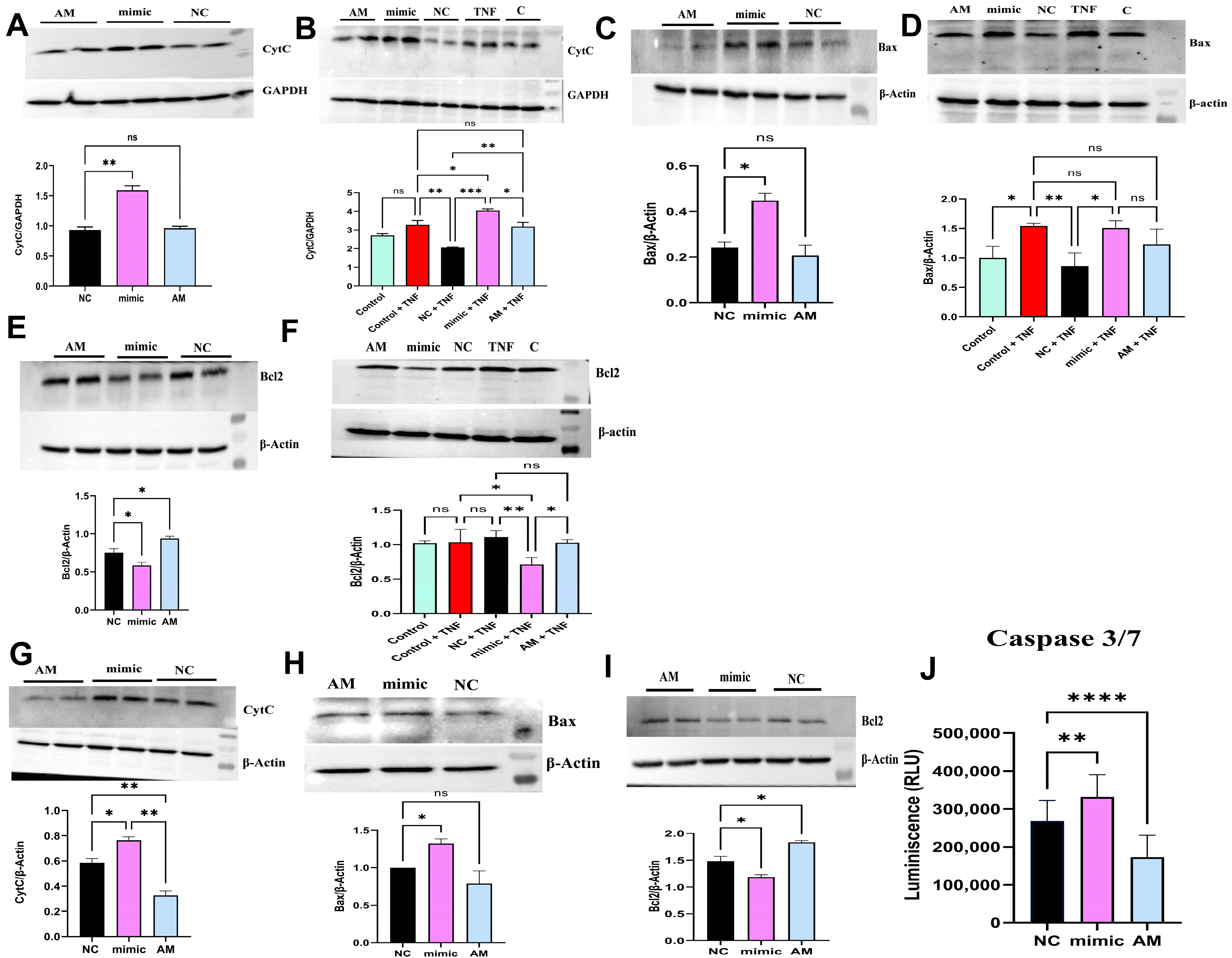
2.8. miR-4693-5p Promotes Apoptosis in RA
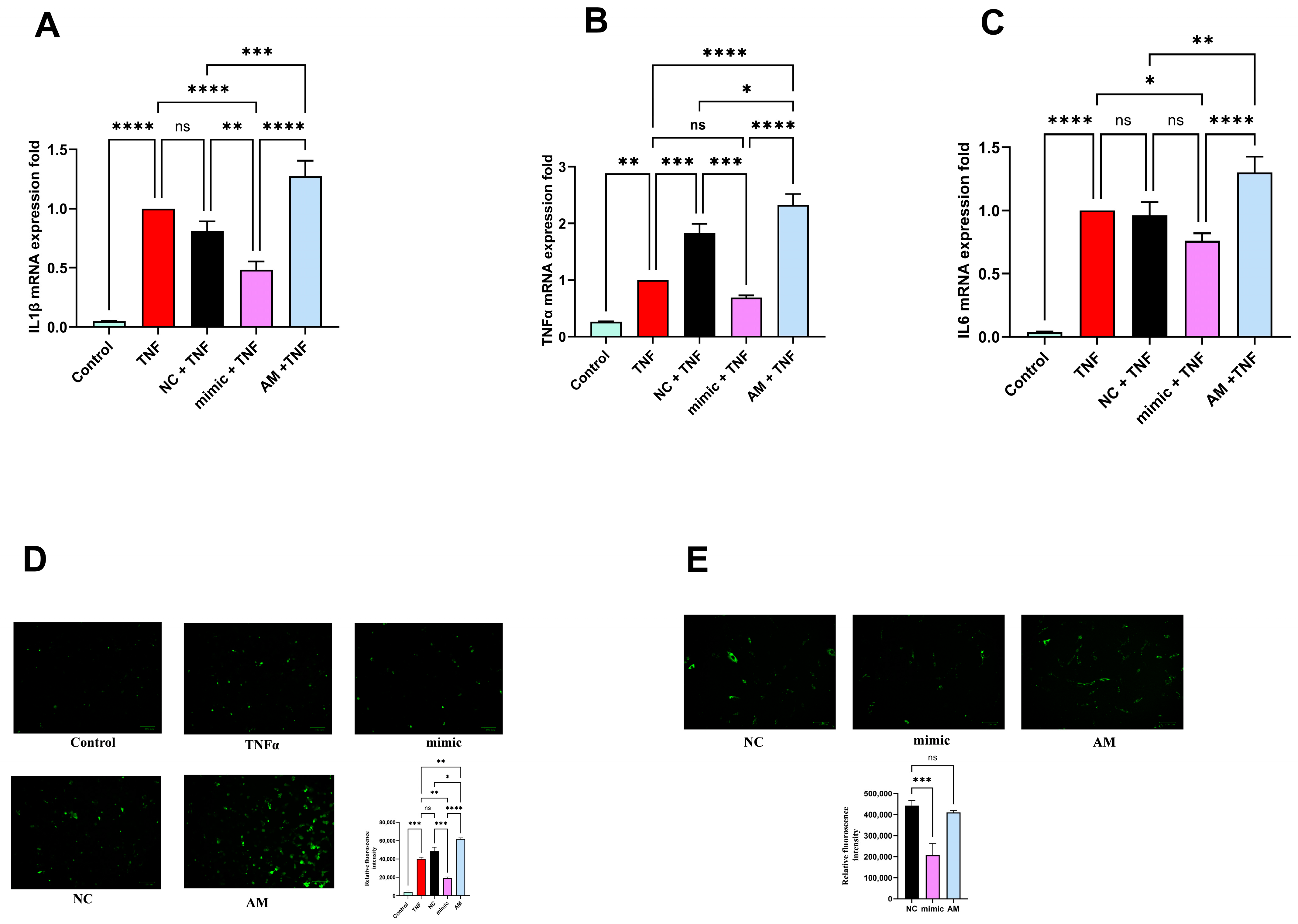
2.9. miR-4693-5p Inhibits Inflammatory Cytokines Production
2.10. miR-4693-5p Reduces Cellular ROS Level
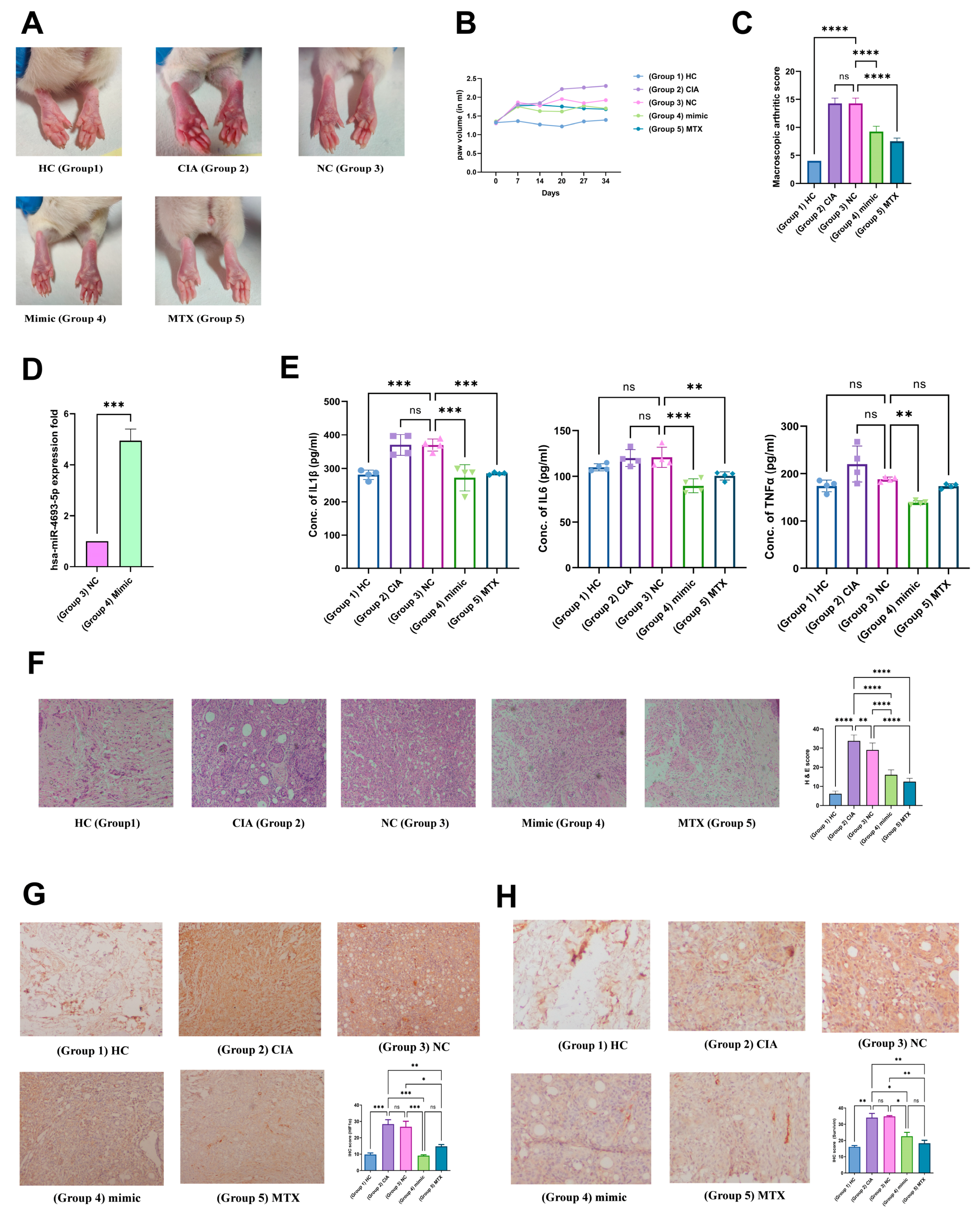
2.11. In Vivo Validation
2.11.1. miR-4693-5p Reduces Macroscopic Arthritic Score and Pro-Inflammatory Cytokines
2.11.2. Validation of the Anti-Inflammatory Effect of miR-4693-5p
2.11.3. Immunohistochemical Analysis of HIF-1α and Survivin Expression
3. Discussion
4. Materials and Methods
4.1. Sample Collection
Blood Sample Processing
4.2. miRNA Quantification in PBMCs
4.3. Target Prediction
4.4. In Vitro Studies
4.4.1. Cell Culture (SW982)
4.4.2. Primary Cell (RAFLS) Isolation
4.4.3. Cell Transfection
4.4.4. Dual-Luciferase Reporter Assay
4.4.5. Real-Time PCR
4.4.6. Western Blot (WB)
4.4.7. Protein–Protein Interaction (PPI)
4.4.8. Caspase-Glo 3/7 Assay
4.4.9. Total ROS Estimation
4.5. In Vivo Studies
4.5.1. Development of Collagen-Induced Arthritis (CIA) Rat Model
4.5.2. Macroscopic Arthritis Score Evaluation
4.5.3. miRNA Isolation from Rat Synovium
4.5.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5.5. Hematoxylin and Eosin Staining (H & E)
4.5.6. Immunohistochemistry (IHC)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid Arthritis: Pathological Mechanisms and Modern Pharmacologic Therapies. Bone Res. 2018, 6, 15. [Google Scholar] [CrossRef] [PubMed]
- Sparks, J.A. In the Clinic® Rheumatoid Arthritis. Ann. Intern. Med. 2019, 170, ITC1–ITC15. [Google Scholar] [CrossRef]
- Quiñonez-Flores, C.M.; González-Chávez, S.A.; Pacheco-Tena, C. Hypoxia and Its Implications in Rheumatoid Arthritis. J. Biomed. Sci. 2016, 23, 62. [Google Scholar] [CrossRef]
- Yap, H.Y.; Tee, S.Z.Y.; Wong, M.M.T.; Chow, S.K.; Peh, S.C.; Teow, S.Y. Pathogenic Role of Immune Cells in Rheumatoid Arthritis: Implications in Clinical Treatment and Biomarker Development. Cells 2018, 7, 161. [Google Scholar] [CrossRef] [PubMed]
- Kaikkonen, M.U.; Lam, M.T.Y.; Glass, C.K. Editor’s Choice: Non-Coding RNAs as Regulators of Gene Expression and Epigenetics. Cardiovasc. Res. 2011, 90, 430. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Flach, H.; Onizawa, M.; Wei, L.; Mcmanus, M.T.; Weiss, A. Negative Regulation of Hif1a Expression and TH17 Differentiation by the Hypoxia-Regulated MicroRNA MiR-210. Nat. Immunol. 2014, 15, 393–401. [Google Scholar] [CrossRef]
- Storti, P.; Bolzoni, M.; Donofrio, G.; Airoldi, I.; Guasco, D.; Toscani, D.; Martella, E.; Lazzaretti, M.; Mancini, C.; Agnelli, L.; et al. Hypoxia-Inducible Factor (HIF)-1α Suppression in Myeloma Cells Blocks Tumoral Growth in Vivo Inhibiting Angiogenesis and Bone Destruction. Leukemia 2013, 27, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Sahu, S.; Kumari, P.; Gopi, S.R.; Malhotra, R.; Biswas, S. Genome-Wide Identification and Functional Annotation of MiRNAs in Anti-Inflammatory Plant and Their Cross-Kingdom Regulation in Homo Sapiens. J. Biomol. Struct. Dyn. 2017, 35, 1389–1400. [Google Scholar] [CrossRef] [PubMed]
- Saquib, M.; Agnihotri, P.; Monu, X.; Biswas, S. Exogenous MiRNA: A Perspective Role as Therapeutic in Rheumatoid Arthritis. Curr. Rheumatol. Rep. 2021, 23, 43. [Google Scholar] [CrossRef]
- Hou, C.; Wang, D.; Zhang, L. MicroRNA-34a-3p Inhibits Proliferation of Rheumatoid Arthritis Fibroblast-like Synoviocytes. Mol. Med. Rep. 2019, 20, 2563. [Google Scholar] [CrossRef]
- Huang, Q.; Li, B.; Lin, C.; Chen, X.; Wang, T.; Liu, J.; Liu, Y.; Lu, R.; Liao, S.; Ding, X. MicroRNA Sequence Analysis of Plasma Exosomes in Early Legg-Calvé-Perthes Disease. Cell. Signal. 2022, 91, 110184. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, D.; Qin, Y.; Qiu, C.; Zhou, Y.; Dai, M.; Li, L.; Sun, Q.; Jiang, Y. TRIP13, Identified as a Hub Gene of Tumor Progression, Is the Target of MicroRNA-4693-5p and a Potential Therapeutic Target for Colorectal Cancer. Cell Death Discov. 2022, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Pauley, K.M.; Satoh, M.; Chan, A.L.; Bubb, M.R.; Reeves, W.H.; Chan, E.K.L. Upregulated MiR-146a Expression in Peripheral Blood Mononuclear Cells from Rheumatoid Arthritis Patients. Arthritis Res. Ther. 2008, 10, R101. [Google Scholar] [CrossRef] [PubMed]
- Riolo, G.; Cantara, S.; Marzocchi, C.; Ricci, C. MiRNA Targets: From Prediction Tools to Experimental Validation. Methods Protoc. 2021, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Kaloni, D.; Chakraborty, D.; Tiwari, A.; Biswas, S. In Silico Studies on the Phytochemical Components of Murraya Koenigii Targeting TNF-α in Rheumatoid Arthritis. J. Herb. Med. 2020, 24, 100396. [Google Scholar] [CrossRef]
- Hu, F.; Mu, R.; Zhu, J.; Shi, L.; Li, Y.; Liu, X.; Shao, W.; Li, G.; Li, M.; Su, Y.; et al. Hypoxia and Hypoxia-Inducible Factor-1α Provoke Toll-like Receptor Signalling-Induced Inflammation in Rheumatoid Arthritis. Ann. Rheum. Dis. 2014, 73, 928–936. [Google Scholar] [CrossRef]
- Khansai, M.; Phitak, T.; Klangjorhor, J.; Udomrak, S.; Fanhchaksai, K.; Pothacharoen, P.; Kongtawelert, P. Effects of Sesamin on Primary Human Synovial Fibroblasts and SW982 Cell Line Induced by Tumor Necrosis Factor-Alpha as a Synovitis-like Model. BMC Complement. Altern. Med. 2017, 17, 532. [Google Scholar] [CrossRef]
- Bai, H.; Ge, S.; Lu, J.; Qian, G.; Xu, R. Hypoxia Inducible Factor-1α-Mediated Activation of Survivin in Cervical Cancer Cells. J. Obstet. Gynaecol. Res. 2013, 39, 555–563. [Google Scholar] [CrossRef]
- Mobahat, M.; Narendran, A.; Riabowol, K. Survivin as a Preferential Target for Cancer Therapy. Int. J. Mol. Sci. 2014, 15, 2494. [Google Scholar] [CrossRef]
- D’Ignazio, L.; Bandarra, D.; Rocha, S. NF-κB and HIF Crosstalk in Immune Responses. FEBS J. 2016, 283, 413. [Google Scholar] [CrossRef]
- Tafani, M.; Sansone, L.; Limana, F.; Arcangeli, T.; De Santis, E.; Polese, M.; Fini, M.; Russo, M.A. The Interplay of Reactive Oxygen Species, Hypoxia, Inflammation, and Sirtuins in Cancer Initiation and Progression. Oxid. Med. Cell. Longev. 2016, 2016, 3907147. [Google Scholar] [CrossRef]
- Bai, L.; Bai, Y.; Yang, Y.; Zhang, W.; Huang, L.; Ma, R.; Wang, L.; Duan, H.; Wan, Q. Baicalin Alleviates Collagen-Induced Arthritis and Suppresses TLR2/MYD88/NF-ΚB P65 Signaling in Rats and HFLS-RAs. Mol. Med. Rep. 2020, 22, 2833. [Google Scholar] [CrossRef]
- Falconer, J.; Murphy, A.N.; Young, S.P.; Clark, A.R.; Tiziani, S.; Guma, M.; Buckley, C.D. Synovial Cell Metabolism and Chronic Inflammation in Rheumatoid Arthritis. Arthritis Rheumatol. 2018, 70, 984. [Google Scholar] [CrossRef]
- Fitriana, M.; Hwang, W.L.; Chan, P.Y.; Hsueh, T.Y.; Liao, T.T. Roles of MicroRNAs in Regulating Cancer Stemness in Head and Neck Cancers. Cancers 2021, 13, 1742. [Google Scholar] [CrossRef]
- Zhu, X.; Wu, L.; Mo, X.; Xia, W.; Guo, Y.; Wang, M.; Zeng, K.; Wu, J.; Qiu, Y.; Lin, X.; et al. Identification of PBMC-Expressed MiRNAs for Rheumatoid Arthritis. Epigenetics 2020, 15, 386. [Google Scholar] [CrossRef]
- Saquib, M.; Agnihotri, P.; Monu, X.; Biswas, S. Interrelated Grid of Non-Coding RNA: An Important Aspect in Rheumatoid Arthritis Pathogenesis. Mol. Biol. Rep. 2023, 50, 6217–6232. [Google Scholar] [CrossRef]
- Rakhecha, B.; Agnihotri, P.; Dakal, T.C.; Saquib, M.; Monu, X.; Biswas, S. Anti-Inflammatory Activity of Nicotine Isolated from Brassica Oleracea in Rheumatoid Arthritis. Biosci. Rep. 2022, 42, 20211392. [Google Scholar] [CrossRef]
- Lee, S.H.; Golinska, M.; Griffiths, J.R. HIF-1-Independent Mechanisms Regulating Metabolic Adaptation in Hypoxic Cancer Cells. Cells 2021, 10, 2371. [Google Scholar] [CrossRef]
- Mokuda, S.; Miyazaki, T.; Ito, Y.; Yamasaki, S.; Inoue, H.; Guo, Y.; Kong, W.S.; Kanno, M.; Takasugi, K.; Sugiyama, E.; et al. The Proto-Oncogene Survivin Splice Variant 2B Is Induced by PDGF and Leads to Cell Proliferation in Rheumatoid Arthritis Fibroblast-like Synoviocytes. Sci. Rep. 2015, 5, 9795. [Google Scholar] [CrossRef]
- Mita, A.C.; Mita, M.M.; Nawrocki, S.T.; Giles, F.J. Survivin: Key Regulator of Mitosis and Apoptosis and Novel Target for Cancer Therapeutics. Clin. Cancer Res. 2008, 14, 5000–5005. [Google Scholar] [CrossRef]
- Bae, T.; Hallis, S.P.; Kwak, M.K. Hypoxia, Oxidative Stress, and the Interplay of HIFs and NRF2 Signaling in Cancer. Exp. Mol. Med. 2024, 1–14. [Google Scholar] [CrossRef]
- Hua, C.; Daien, C.I.; Combe, B.; Landewe, R. Diagnosis, Prognosis and Classification of Early Arthritis: Results of a Systematic Review Informing the 2016 Update of the EULAR Recommendations for the Management of Early Arthritis. RMD Open 2017, 3, e000406. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Sharma, S.; Saroha, A.; Bhakuni, D.S.; Malhotra, R.; Zahur, M.; Oellerich, M.; Das, H.R.; Asif, A.R. Identification of Novel Autoantigen in the Synovial Fluid of Rheumatoid Arthritis Patients Using an Immunoproteomics Approach. PLoS ONE 2013, 8, e56246. [Google Scholar] [CrossRef] [PubMed]
- Monu, X.; Agnihotri, P.; Saquib, M.; Sarkar, A.; Chakraborty, D.; Kumar, U.; Biswas, S. Transthyretin and Receptor for Advanced Glycation End Product’s Differential Levels Associated with the Pathogenesis of Rheumatoid Arthritis. J. Inflamm. Res. 2021, 14, 5581–5596. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, D.; Sarkar, A.; Mann, S.; Monu, X.; Agnihotri, P.; Saquib, M.; Malik, S.; Kumavat, R.; Mathur, A.; Biswas, S. Estrogen-Mediated Differential Protein Regulation and Signal Transduction in Rheumatoid Arthritis. J. Mol. Endocrinol. 2022, 69, R25–R43. [Google Scholar] [CrossRef]
- Wangyang, Y.; Yi, L.; Wang, T.; Feng, Y.; Liu, G.; Li, D.; Zheng, X. MiR-199a-3p Inhibits Proliferation and Induces Apoptosis in Rheumatoid Arthritis Fibroblast-like Synoviocytes via Suppressing Retinoblastoma 1. Biosci. Rep. 2018, 38, BSR20180982. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.M.; Zorman, J.; Horton, M.; Dubey, S.; Ter Meulen, J.; Vora, K.A. Caspase Activation as a Versatile Assay Platform for Detection of Cytotoxic Bacterial Toxins. J. Clin. Microbiol. 2013, 51, 2970. [Google Scholar] [CrossRef]
- Kim, H.; Xue, X. Detection of Total Reactive Oxygen Species in Adherent Cells by 2′,7′-Dichlorodihydrofluorescein Diacetate Staining. J. Vis. Exp. 2020, 2020, e60682. [Google Scholar] [CrossRef]
- Kang, X.; Jiao, Y.; Zhou, Y.; Meng, C.; Zhou, X.; Song, L.; Jiao, X.; Pan, Z. MicroRNA-5112 Targets IKKγ to Dampen the Inflammatory Response and Improve Clinical Symptoms in Both Bacterial Infection and DSS-Induced Colitis. Front. Immunol. 2022, 13, 779770. [Google Scholar] [CrossRef]
- Bendele, A.; Mccomb, J.; Gould, T.; Mcabee, T.; Sennello, G.; Chlipala, E.; Guy, M. Animal Models of Arthritis: Relevance to Human Disease. Toxicol. Pathol. 1999, 27, 134–142. [Google Scholar] [CrossRef]
- af Klint, E.; Catrina, A.I.; Matt, P.; Neregråd, P.; Lampa, J.; Ulfgren, A.K.; Klareskog, L.; Lindblad, S. Evaluation of Arthroscopy and Macroscopic Scoring. Arthritis Res. Ther. 2009, 11, R81. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, N.; Furukawa, K.; Endo, Y.; Shimizu, T.; Sumiyoshi, R.; Umeda, M.; Koga, T.; Kawashiri, S.Y.; Igawa, T.; Ichinose, K.; et al. Methotrexate Alters the Expression of Microrna in Fibroblast-like Synovial Cells in Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 11561. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saquib, M.; Agnihotri, P.; Sarkar, A.; Malik, S.; Mann, S.; Chakraborty, D.; Joshi, L.; Malhotra, R.; Biswas, S. Functional Significance of miR-4693-5p in Targeting HIF1α and Its Link to Rheumatoid Arthritis Pathogenesis. Non-Coding RNA 2024, 10, 22. https://doi.org/10.3390/ncrna10020022
Saquib M, Agnihotri P, Sarkar A, Malik S, Mann S, Chakraborty D, Joshi L, Malhotra R, Biswas S. Functional Significance of miR-4693-5p in Targeting HIF1α and Its Link to Rheumatoid Arthritis Pathogenesis. Non-Coding RNA. 2024; 10(2):22. https://doi.org/10.3390/ncrna10020022
Chicago/Turabian StyleSaquib, Mohd, Prachi Agnihotri, Ashish Sarkar, Swati Malik, Sonia Mann, Debolina Chakraborty, Lovely Joshi, Rajesh Malhotra, and Sagarika Biswas. 2024. "Functional Significance of miR-4693-5p in Targeting HIF1α and Its Link to Rheumatoid Arthritis Pathogenesis" Non-Coding RNA 10, no. 2: 22. https://doi.org/10.3390/ncrna10020022
APA StyleSaquib, M., Agnihotri, P., Sarkar, A., Malik, S., Mann, S., Chakraborty, D., Joshi, L., Malhotra, R., & Biswas, S. (2024). Functional Significance of miR-4693-5p in Targeting HIF1α and Its Link to Rheumatoid Arthritis Pathogenesis. Non-Coding RNA, 10(2), 22. https://doi.org/10.3390/ncrna10020022