
Batch Fermentation of Salt-Acclimatizing Microalga for Omega-3 Docosahexaenoic Acid Production Using Biodiesel-Derived Crude Glycerol Waste as a Low-Cost Substrate


Abstract
:1. Introduction
2. Materials and Methods
2.1. Algal Strain and Cultivation Medium
2.2. Pretreatment of Crude Glycerol
2.3. Aerobic Batch Heterotrophic Fermentation
2.4. Cell Growth and Lipid Extraction
3. Results and Discussion
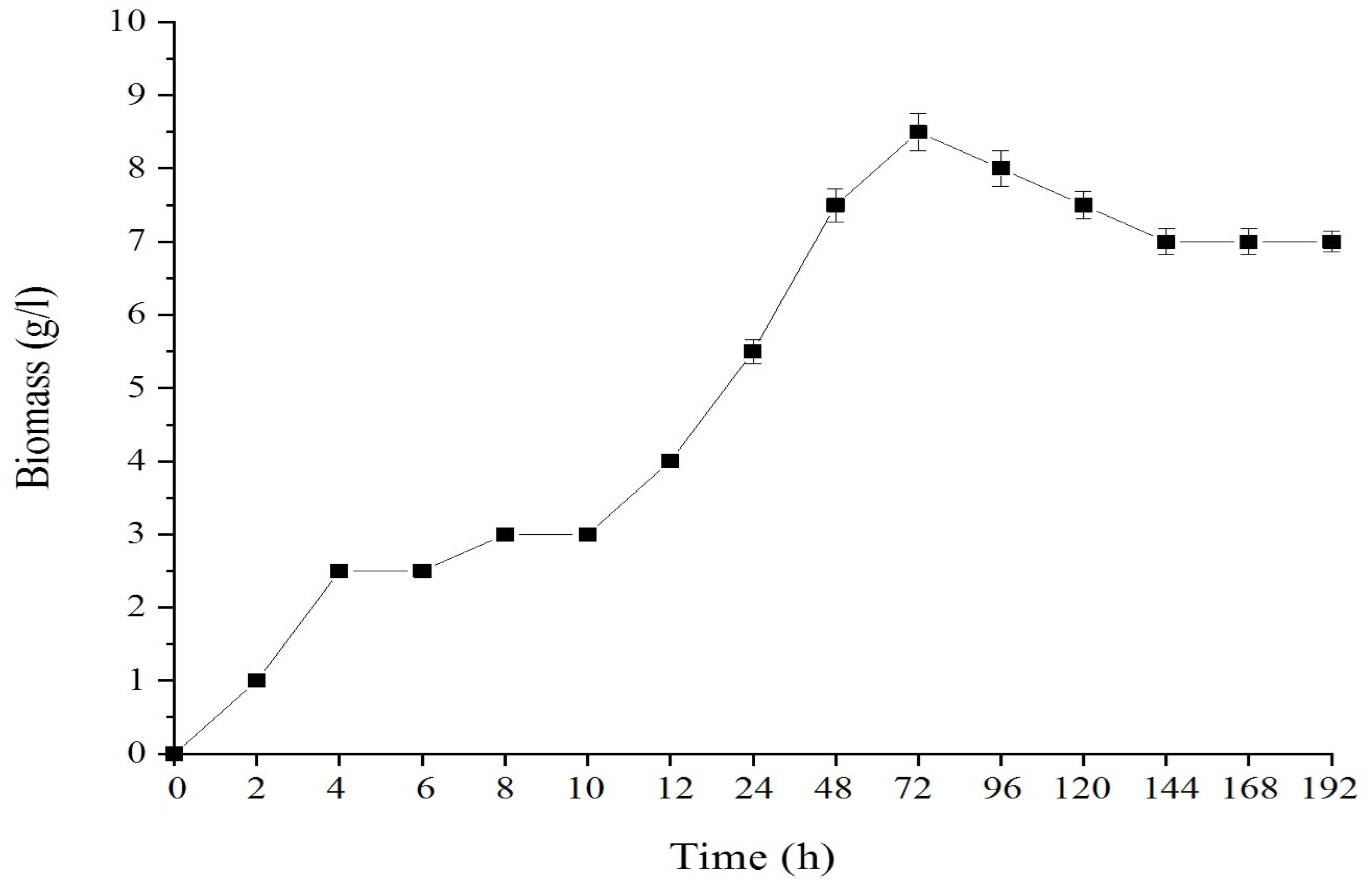
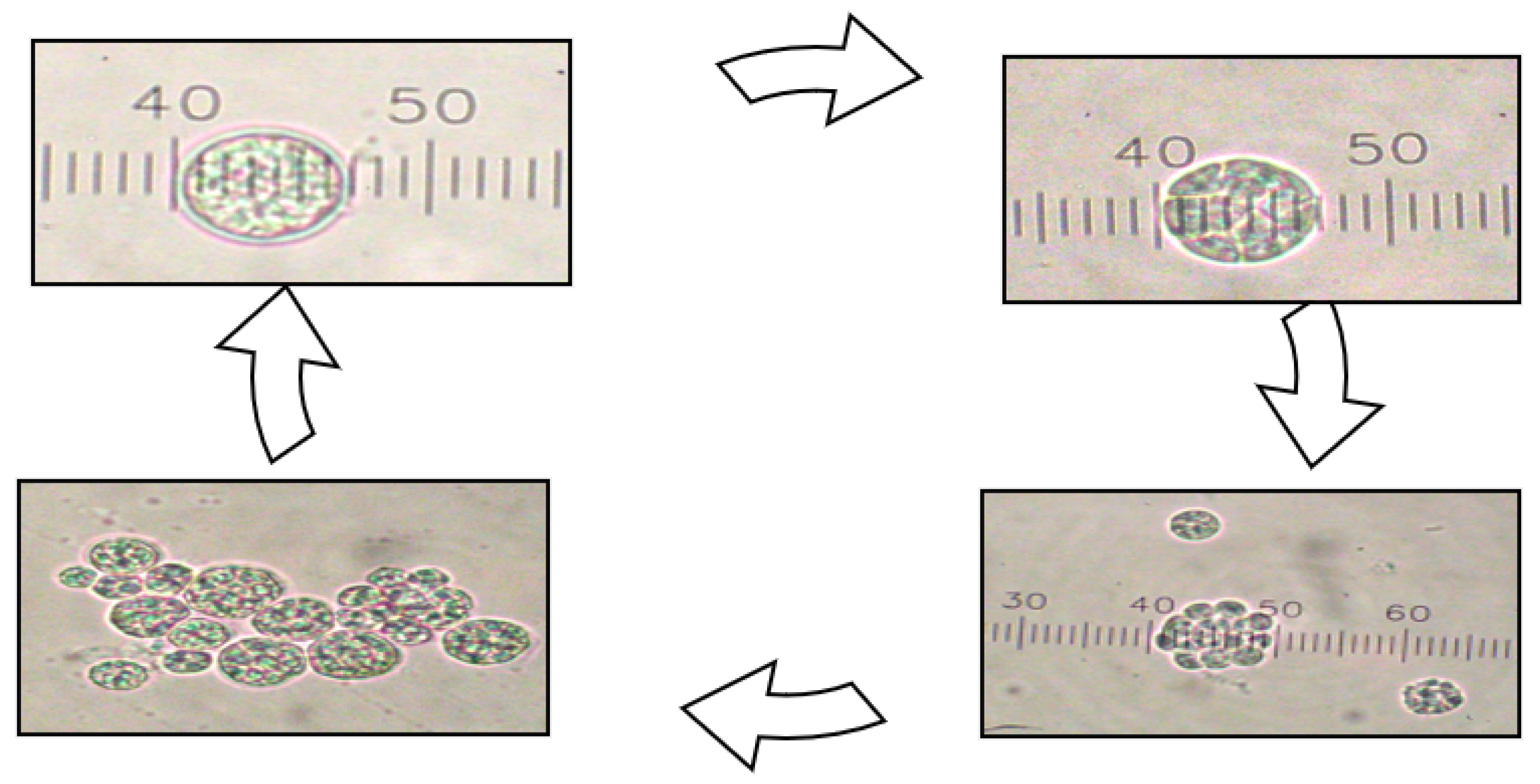
3.1. Growth Monitoring and Morphology
3.2. Characteristics of the BDWG
3.3. Effect of the BDWG Concentration on Algal Biomass and DHA Accumulation
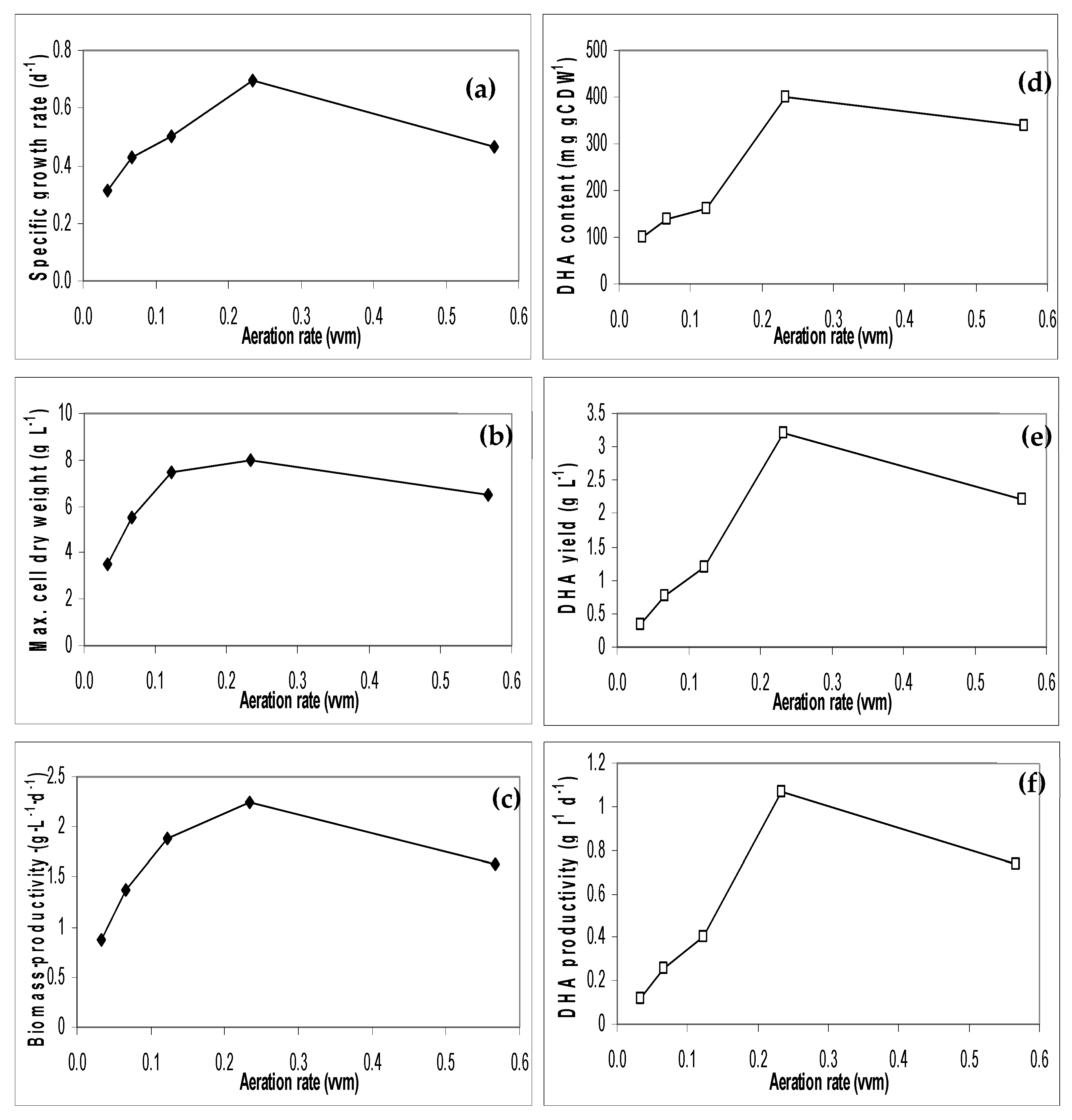
3.4. Effect of Aeration Rates on Algal Biomass and DHA Production
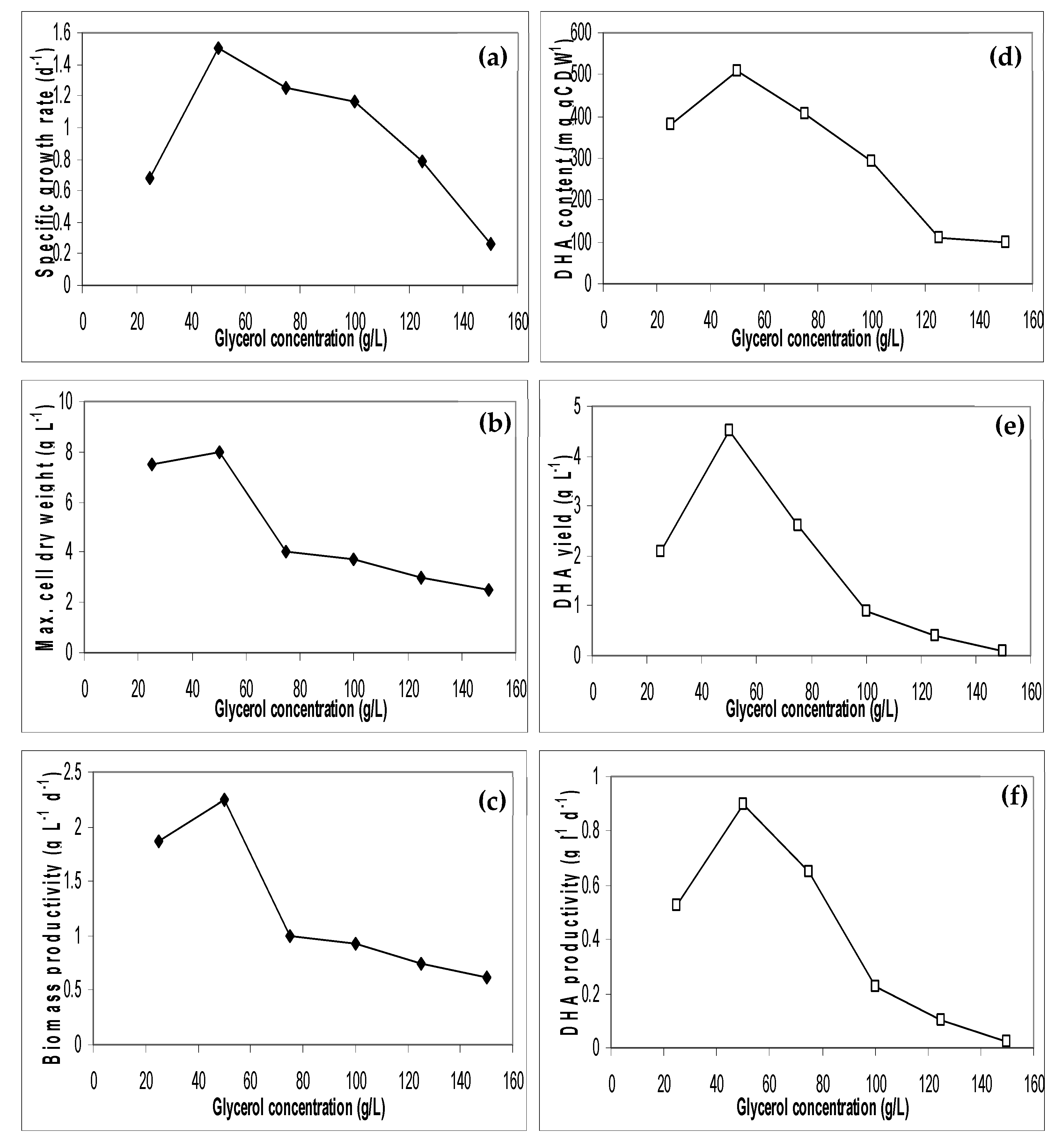
3.5. Effect of Initial Glycerol Waste Concentration
3.6. Effect of Carbon Sources
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Arachidonic acid |
| AC | Autotrophic cultivation |
| ALA | Alpha linoleic acid |
| ATCC | American Type Culture Collection |
| ATCC MYA-1381 | Schizochytrium limacinum |
| BGDW | Biodiesel-derived glycerol waste |
| CDW | Cell dry weight |
| DHA | Docohexaenoic acid |
| DGLA | Homo gamma linoleic acid |
| EPA | Eicosapeentaenoic acid |
| HC | Heterotrophic cultivation |
| FAMEs | Fatty acid methyl esters |
| FID | Flame Ionisation Detector |
| GC | Gas chromatography |
| GPY | Glucose–Peptone–Yeast |
| LA | Linoleic acid |
| MC | Mixotrophic cultivation |
| PUAFs | Polyunsaturated fatty acids |
| v/v | Volume by volume |
| v/v/m | Volume by volume by minute |
| WCO | Waste cooking oil |
| w/v | Weight by volume |
| w/w | Weight by weight |
References
- Saman, A.-u.-R.; Wijesekara, R.G.; Nomura, N.; Sato, S.; Matsumura, M. Pre-treatment and utilization of raw glycerol from sunflower oil biodiesel for growth and 1,3-propanediol production by Clostridium butyricum. J. Chem. Technol. Biotechnol. 2008, 83, 1072–1080. [Google Scholar]
- Chi, Z.; Pyle, D.; Wen, Z.; Frear, C.; Chen, S. A laboratory study of producing docosahexaenoic acid from biodiesel-waste glycerol by microalgal fermentation. Process Biochem. 2007, 42, 1537–1545. [Google Scholar] [CrossRef]
- Chi, Z.Y.; Liu, Y.; Frear, C.; Chen, S.L. Study of a two-stage growth of DHA-producing marine algae Schizochytrium limacinum SR21 with shifting dissolved oxygen level. Appl. Microbiol. Biotechnol. 2009, 81, 1141–1148. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.W.; Dasari, M.A.; Sutterlin, W.R.; Suppes, G.J. Removal of residual catalyst from simulated biodiesel’s crude glycerol for glycerol hydro-genolysis to propylene glycol. Ind. Eng. Chem. Res. 2006, 45, 791–795. [Google Scholar] [CrossRef]
- Dasari, M.A.; Kiatsimkul, P.-P.; Sutterlin, W.R.; Suppes, G.J. Low-pressure hydrogenolysis of glycerol to propylene glycol. Appl. Catal. A Gen. 2005, 281, 225–231. [Google Scholar] [CrossRef]
- Dharmadi, Y.; Murarka, A.; Gonzalez, R. Anaerobic fermentation of glycerol by Escherichia coli: A new platform for metabolic engineering. Biotechnol. Bioeng. 2006, 94, 821–829. [Google Scholar] [CrossRef]
- Ethier, S.; Woisard, K.; Vaughan, D.; Wen, Z. Continuous culture of the microalgae Schizochytrium limacinum on biodiesel-derived crude glycerol for producing docosahexaenoic acid. Bioresour. Technol. 2011, 102, 88–93. [Google Scholar] [CrossRef]
- Johnson, D.T.; Taconi, K.A. The glycerin glut: Options for the value-added conversion of crude glycerol resulting from biodiesel production. Environ. Prog. 2007, 26, 338–348. [Google Scholar] [CrossRef]
- Johnson, M.B.; Wen, Z. Production of biodiesel fuel from the microalga Schizochytrium limacinum by direct transesterification of algal biomass. Energy Fuel 2009, 10, 5179–5183. [Google Scholar] [CrossRef]
- Liang, Y.; Sarkany, N.; Cui, Y.; Blackburn, J.W. Batch stage study of lipid production from crude glycerol derived from yellow grease or animal fats through microalgal fermentation. Bioresour. Technol. 2010, 101, 6745–6750. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Wu, J.-Y.; Ho, K.-P. Characterization of oxygen transfer conditions and their effects on Phaffia rhodozyma growth and carotenoid production in shake-flask cultures. Biochem. Eng. J. 2006, 27, 331–335. [Google Scholar] [CrossRef]
- Indarti, E.; Majid, M.I.A.; Hashim, R.; Chong, A. Direct FAME synthesis for rapid total lipid analysis from fish oil and cod liver oil. J. Food Compos. Anal. 2005, 18, 161–170. [Google Scholar] [CrossRef]
- Meesters, P.A.E.P.; Huijberts, G.N.M.; Eggink, G. High-cell-density cultivation of the lipid accumulating yeast Cryptococcus curvatus using glycerol as a carbon source. Appl. Microbiol. Biotechnol. 1996, 45, 575–579. [Google Scholar] [CrossRef]
- Narayan, M.S.; Manoj, G.P.; Vatchravelu, K.; Bhagyalakshmi, N.; Mahadevaswamy, M. Utilization of glycerol as carbon source on the growth, pigment and lipid production in Spirulina platensis. Int. J. Food Sci. Nutr. 2005, 56, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, S.; Aggelis, G. Lipid production by Yarrowia lipolytica growing on industrial glycerol in a single-stage continuous culture. Bioresour. Technol. 2002, 82, 43–49. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Fakasb, S.; Ficka, M.; Chevalota, I.; Galiotou-Panayotoub, M.; Komaitisb, M.; Marca, I.; Aggelisc, G. Biotechnological valorisation of raw glycerol discharged after bio-diesel (fatty acid methyl esters) manufacturing process: Production of 1,3-propanediol, citric acid and single cell oil. Biomass Bioenergy 2008, 32, 60–71. [Google Scholar] [CrossRef]
- Pyle, D.J.; Garcia, R.A.; Wen, Z. Producing Docosahexaenoic Acid (DHA)-Rich Algae from Biodiesel-Derived Crude Glycerol: Effects of Impurities on DHA Production and Algal Biomass Composition. J. Agric. Food Chem. 2008, 56, 3933–3939. [Google Scholar] [CrossRef]
- Ren, L.J.; Ji, X.J.; Huang, H.; Qu, J.; Feng, Y.; Tong, Q.Q.; Ouyang, P.K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef]
- Zhang, Y.; Ward, V.; Dennis, D.; Plechkova, N.V.; Armenta, R.; Rehmann, L. Efficient Extraction of a Docosahexaenoic Acid (DHA)-Rich Lipid Fraction from Thraustochytrium sp. Using Ionic Liquids. Materials 2018, 11, 1986. [Google Scholar] [CrossRef]
- Yang, Y.; Li, G.; Li, F.; Xu, F.; Hu, P.; Xie, Z.; Lu, X.; Ding, Y.; Wang, Z. Impact of DHA from Algal Oil on the Breast Milk DHA Levels of Lactating Women: A Randomized Controlled Trial in China. Nutrients 2022, 14, 3410. [Google Scholar] [CrossRef]
- Sittiprapaporn, P.; Bumrungpert, A.; Suyajai, P.; Stough, C. Effectiveness of fish oil-DHA supplementation for cognitive function in Thai children: A randomized, doubled-blind, two-dose, placebo-controlled clinical trial. Foods 2022, 11, 2595. [Google Scholar] [CrossRef]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Essential fatty acids in health and chronic disease. Am. J. Clin. Nutr. 1999, 70, 560S–569S. [Google Scholar] [CrossRef] [PubMed]
- Tyson, K.S.; Bozell, J.; Wallace, R.; Petersen, E.; Moens, L. Biomass Oil Analysis: Research Needs and Recommendations; NREL/TP-510-34796; National Renewable Energy Laboratory: Golden, CO, USA, 2004. [Google Scholar]
- Ursin, V.M. Modification of Plant Lipids for Human Health: Development of Functional Land-Based Omega-3 Fatty Acids. J. Nutr. 2003, 133, 4271–4274. [Google Scholar] [CrossRef] [PubMed]
- Bozzatello, P.; Blua, C.; Rocca, P.; Bellino, S. Mental Health in Childhood and Adolescence: The Role of Polyunsaturated Fatty Acids. Biomedicines 2021, 9, 850. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Liang, Y.; Cui, Y.; Trushenski, J.; Blackburn, J.W. Converting crude glycerol derived from yellow grease to lipids through yeast fermentation. Bioresour. Technol. 2010, 101, 7581–7586. [Google Scholar] [CrossRef]
- Wu, S.T.; Yu, S.T.; Lin, L.P. Effect of culture conditions on docosahexaenoic acid production by Schizochytrium sp. 31. Proc. Biochem. 2005, 40, 3103–3108. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Dujjanutat, P.; Muanruksa, P.; Kaewkannetra, P. Biocircular platform for third generation biodiesel production: Batch/fed batch mixotrophic cultivations of microalgae using glycerol waste as a carbon source. Biochem. Eng. J. 2021, 175, 108128. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Kaewkannetra, P. Cultivation of microalga, Chlorella vulgaris under different auto-hetero-mixo trophic growths as a raw material during biodiesel production and cost evaluation. Energy 2014, 78, 4–8. [Google Scholar] [CrossRef]
- Rattanapoltee, P.; Kaewkannetra, P. Utilization of Agricultural Residues of Pineapple Peels and Sugarcane Bagasse as Cost-Saving Raw Materials in Scenedesmus acutus for Lipid Accumulation and Biodiesel Production. Appl. Biochem. Biotechnol. 2014, 173, 1495–1510. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Before Treatment | After Treatment |
|---|---|---|
| Colour | Dark brown-black | brown |
| pH | 9.98 | 6.5 |
| After mixing with culture medium | Water and soap were separated | Well mixed |
| Time (Day) | BDGW (g/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 25 (g/L) | 50 (g/L) | 75 (g/L) | 100 (g/L) | 125 (g/L) | ||||||
| Biomass (g/L) | DHA in Biomass (mg/g) | Biomass (g/L) | DHA in Biomass (mg/g) | Biomass (g/L) | DHA in Biomass (mg/g) | Biomass (g/L) | DHA in Biomass (mg/g) | Biomass (g/L) | DHA in Biomass (mg/g) | |
| 0 | 0.70 | *nd | 0.70 | *nd | 0.85 | *nd | 0.80 | *nd | 0.80 | *nd |
| 1 | 3.70 | 219.590 | 2.50 | 180.677 | 2.25 | *nd | 1.95 | *nd | 1.67 | *nd |
| 2 | 4.35 | 230.635 | 3.42 | 215.055 | 2.55 | *nd | 2.03 | *nd | 1.70 | *nd |
| 3 | 5.00 | 323.296 | 5.17 | 323.021 | 3.50 | *nd | 3.33 | *nd | 1.70 | *nd |
| 4 | 5.55 | 379.532 | 6.48 | 329.823 | 4.93 | 224.328 | 3.35 | 215.220 | 1.60 | *nd |
| 5 | 5.35 | 131.671 | 8.98 | 510.527 | 5.05 | 393.659 | 3.27 | 292.470 | 1.60 | *nd |
| 6 | 4.58 | 153.338 | 9.03 | 371.087 | 6.75 | 248.955 | 3.53 | 140.750 | 1.70 | 97.716 |
| 7 | 3.63 | 201.489 | 9.13 | 247.764 | 6.53 | 407.849 | 2.50 | 209.488 | 1.60 | 111.564 |
| 8 | 3.78 | 154.119 | 9.45 | 357.488 | 8.15 | 203.118 | 2.32 | 205.859 | 1.60 | *nd |
| Glycerol Sources | Glycerol Concentrations Where Inhibition Occurs (g/L) | Cell DRY Weight (g/L) | DHA Production (g/L) | References |
|---|---|---|---|---|
| Soybean oil | 64–85 | 18.7–18.6 | 3.05–2.71 | Chi et al., 2007 [2] |
| Soybean oil | 40 | 7.0 | 1.5 | Ethier et al., 2011 [7] |
| 50:50 (w/w) chicken fat and soybean oil mixture | 60 | 12 | 1.6 | Ethier et al., 2011 [7] |
| Yellow grease (recycled cooking oil) | 35 | 13.3 | - | Liang et al., 2010 [10] |
| Waste cooking oil (WCO) | 50 | 8.0 | 4.8 | This study |
| Parameter | Unit | Crude Glycerol | Glucose | Commercial Glycerol |
|---|---|---|---|---|
| Maximum cell dry weight | g/L | 8.95 | 9.85 | 9.80 |
| Maximum specific growth rate | /d | 0.50 | 0.44 | 0.39 |
| Maximum biomass productivity | g/Ld | 1.79 | 1.64 | 1.63 |
| DHA content | mg/gDW | 586 | 580 | 502 |
| DHA yield | g/L | 7.10 | 7.70 | 6.70 |
| DHA productivity | g/Ld | 1.42 | 1.28 | 1.11 |
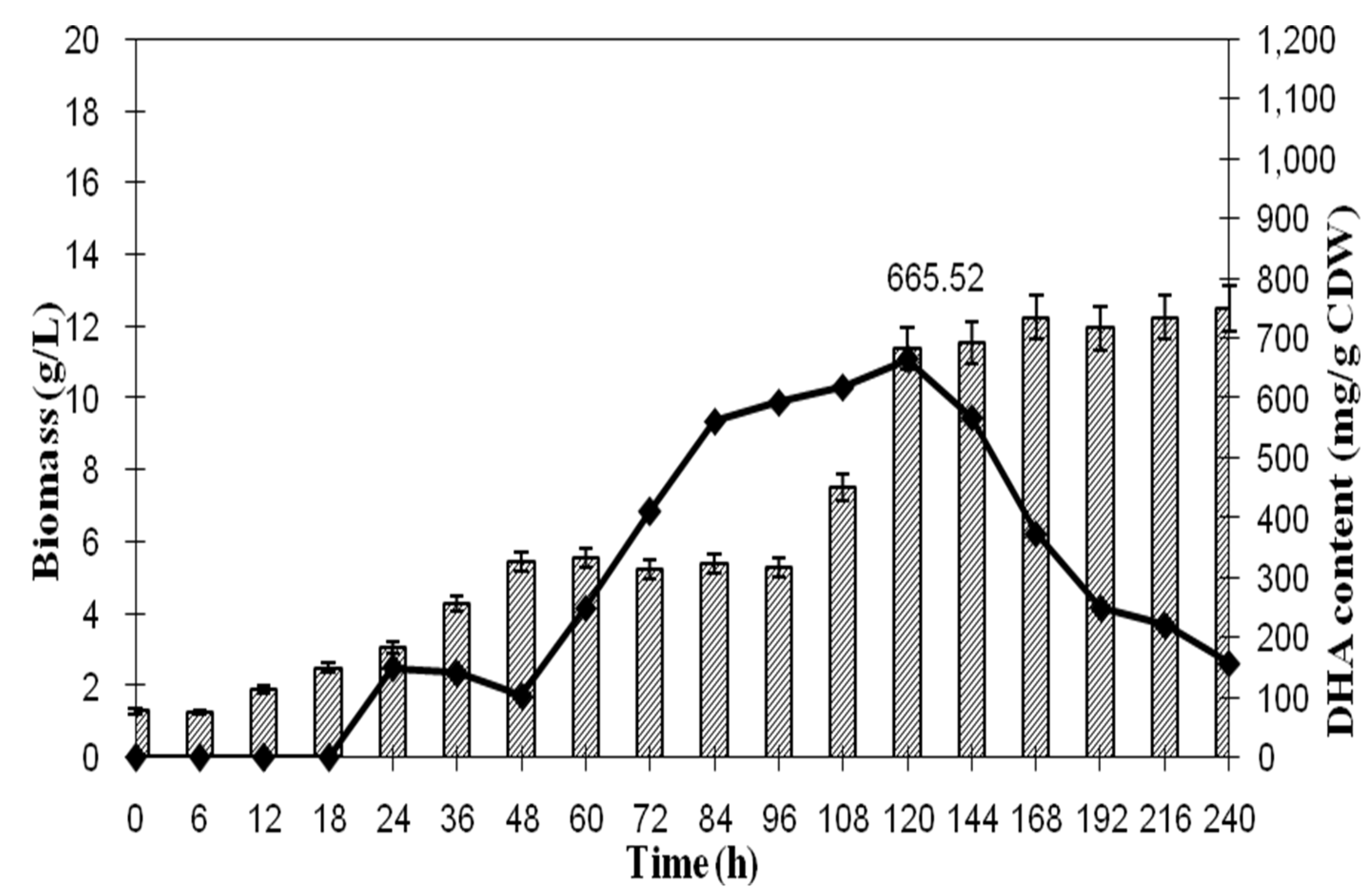
| Time (h) | Biomass (g/L) | Glycerol Conc. (g/L) | Biomass Yield (g/g-Glycerol) | Volumetric Productivity Rate of DHA | DHA | |
|---|---|---|---|---|---|---|
| DHA in Biomass (mg/g) | DHA YIELD (g/g Glycerol) | |||||
| 0 | 1.30 | 18.08 | *nd | *nd | *nd | *nd |
| 6 | 1.25 | 15.67 | 0.52 | *nd | *nd | *nd |
| 12 | 1.90 | 14.53 | 1.67 | *nd | *nd | *nd |
| 18 | 2.50 | 9.41 | 0.49 | *nd | *nd | *nd |
| 24 | 3.05 | 6.57 | 1.07 | 0.02 | 149.48 | 0.16 |
| 36 | 4.30 | 6.07 | 8.60 | 0.02 | 141.94 | 1.12 |
| 48 | 5.45 | 5.03 | 5.24 | 0.01 | 103.60 | 0.54 |
| 60 | 5.55 | 4.88 | 37.00 | 0.02 | 250.45 | 9.27 |
| 72 | 5.25 | 4.78 | 52.50 | 0.03 | 411.71 | 21.60 |
| 84 | 5.40 | 3.92 | 6.28 | 0.04 | 561.70 | 3.52 |
| 96 | 5.30 | 3.49 | 12.23 | 0.03 | 593.86 | 7.33 |
| 108 | 7.50 | 2.97 | 14.42 | 0.04 | 619.47 | 8.94 |
| 120 | 11.40 | 1.76 | 9.42 | 0.06 | 665.52 | 6.27 |
| 144 | 11.05 | nd | 6.28 | 0.05 | 568.18 | 3.78 |
| 168 | 12.25 | nd | nd | 0.03 | 373.67 | *nd |
| 192 | 11.95 | nd | nd | 0.02 | 250.65 | *nd |
| 216 | 12.75 | nd | nd | 0.01 | 212.54 | *nd |
| 240 | 13.00 | nd | nd | 0.01 | 151.95 | *nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanamool, V.; Enmak, P.; Kaewkannetra, P. Batch Fermentation of Salt-Acclimatizing Microalga for Omega-3 Docosahexaenoic Acid Production Using Biodiesel-Derived Crude Glycerol Waste as a Low-Cost Substrate. Fermentation 2024, 10, 86. https://doi.org/10.3390/fermentation10020086
Tanamool V, Enmak P, Kaewkannetra P. Batch Fermentation of Salt-Acclimatizing Microalga for Omega-3 Docosahexaenoic Acid Production Using Biodiesel-Derived Crude Glycerol Waste as a Low-Cost Substrate. Fermentation. 2024; 10(2):86. https://doi.org/10.3390/fermentation10020086
Chicago/Turabian StyleTanamool, Varavut, Prayoon Enmak, and Pakawadee Kaewkannetra. 2024. "Batch Fermentation of Salt-Acclimatizing Microalga for Omega-3 Docosahexaenoic Acid Production Using Biodiesel-Derived Crude Glycerol Waste as a Low-Cost Substrate" Fermentation 10, no. 2: 86. https://doi.org/10.3390/fermentation10020086
APA StyleTanamool, V., Enmak, P., & Kaewkannetra, P. (2024). Batch Fermentation of Salt-Acclimatizing Microalga for Omega-3 Docosahexaenoic Acid Production Using Biodiesel-Derived Crude Glycerol Waste as a Low-Cost Substrate. Fermentation, 10(2), 86. https://doi.org/10.3390/fermentation10020086