

Microbial Factories and Exploiting Synergies of Bioreactor Technologies to Produce Bioproducts



Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioproduct | Nutrient | Organism | Source | Process | Ref. |
|---|---|---|---|---|---|
| Ceramics | Silicon polymers Biominerals | Algae | Exoskeleton | Photobioreactors | [23] [24,25] |
| Insulators | Polyvinyl alcohol (PVA) | Bacteria | Biopolymer | Heterotrophic bioreactor | [26] |
| Liquid crystals | Biominerals | Algae, vascular plants | Biopolymer | Photobioreactor | [27] |
| Optics | 2-methyl pentanol | Bacteria | Enzymatic | Trade secret-Codexis | [28] |
| Semiconductors | Silica frustules | Algae | Exoskeleton | Photobioreactor | [29] |
| Polyvinyl alcohol (PVA) | Algae, vascular plants | Biopolymer | Photobioreactor | [27] | |
| Medical sensors | Glycerides | abfp 1 | Fat-degrade enzymes | Bioreactors | [30] |
| Polymers | [31] | ||||
| Biodiesel | Triglycerides | Algae | Fat droplets | Photobioreactor | [32] |
| Various alcohols | Butanol, propanol | Fungi (yeasts) | Differing pathways | Bioreactor | [33] |
| Ethanol | Various sugars | Fungi (yeasts) | Differing pathways | Bioreactors | [34,35] |
| Cellulosic material | Vascular plants, | Cutin | Photobioreactor | [36] | |
| Lignin | Vascular plants | Cutin | Fermenter | [37] | |
| Glycerol | algae, vascular plants | Fat droplets | Photobioreactor | [38] | |
| Cellulosic material | Fungi | Cell wall | Bioreactors | [39] | |
| Hydrogen | Organics | Bacteria Algae | Biomass Biomass | Dark fermenters | [40,41] |
| Lignins | Vascular plants | Cutin | Vacuum pyrolysis, catalyst | [42] | |
| Glycerol | Algae, vascular plants | Fat droplets | High temperature, pressure, metal catalyst | [43] | |
| Waste glycerol | Bacteria | Biomass | Microbial fuel cell | [44,45] | |
| Ethane | Organic waste | Bacteria | Biomass | Digester, single-multi-stage | [46] |
| Lipids | Algae | Fat droplets | Anaerobic digester | [47] | |
| Other oils | Lipids | Algae, | Fat droplets | Fluidized beds—pyrolysis | [9] |
| vascular plants | Pyrolysis, gasification | [48] | |||
| Solar cells | Silica frustules | Diatoms | Exoskeleton | Photobioreactor | [29] |
| Membranes | PVA hydrogels | Bacteria | Chitosan, Gelatin | Bioreactors | [49] |
| Surfactants | Biopolymers | abfp 1 | Various | Various | [50] |
| Nano- composites | Nano-carbon | Algae, vascular plants | Biopolymers | Photobioreactor | [51] [52] |
| Adhesives | Algae Bacteria Fungi | Biopolymers | Bioreactors | [53,54] [55] [56] | |
| Nanocrystals | Cellulose fibers | Bacteria, vascular plants | Cell wall | Bioreactors | [57] |
| Nanofibers | Proteins, peptides | Gluconacetobacter xylinus | Biopolymer | Bioreactors | [58] |
| Structures | Carbon fiber | Algae | Glycerol | Photobioreactor | [59] |
| Acrylic | Acrylic acid | Algae, vascular plants | Fat droplets | Photobioreactor | [38] |
| PET, PLA | Glycols | Algae, vascular plants | Fat droplets | Photobioreactors | [60,61] |
| Cellulose fibers | Vascular plants | Cell wall | Photobioreactor | [62] | |
| Polyvinyl | Ethanol | Algae, vascular plants | Fat droplets | Photobioreactor | [63] |
| Cement | Calcium, Silica | Algae Vascular plants | Exoskeletons Biomass | Photobioreactor Photobioreactor | [64] [65] |
| Nano-cement nuclei | Calcium carbonate | Algae–cocci | Exoskeletal | Photobioreactor | [66] |
2. Materials and Methods
3. Results
3.1. Results of Yeast and Algae Bioreactors
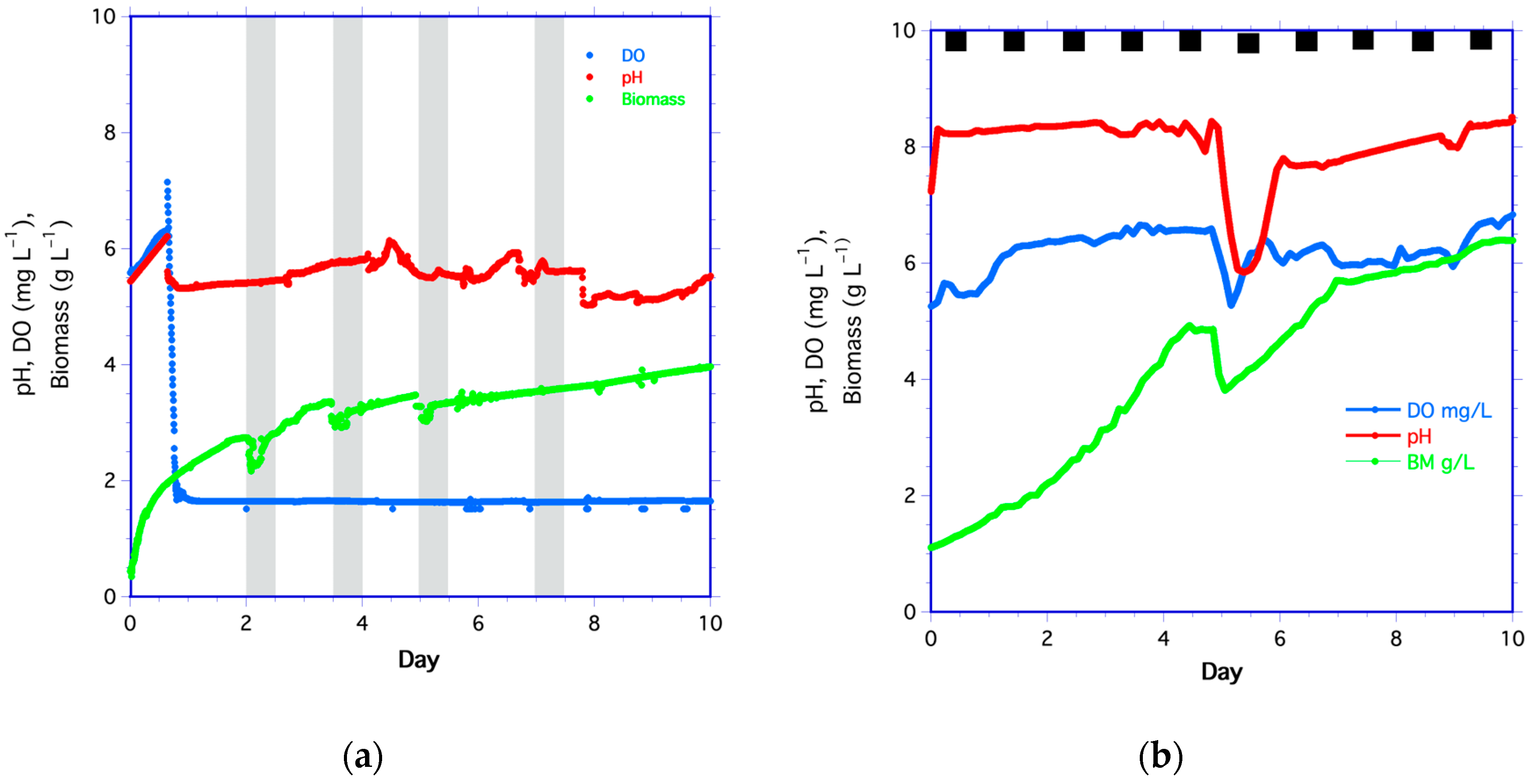
3.1.1. Yeast Time Series
3.1.2. Algae Time Series
3.1.3. Growth Rates, DO, and CO2 Rates
Yeast
Algae
3.2. Results from CO2 Capture and Use by Algae
4. Discussion
CO2 and O2 Processes and Resource Synergies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khoo, C.G.; Dasan, Y.K.; Lam, M.K.; Lee, K.T. Algae biorefinery: Review on a broad spectrum of downstream processes and products. Bioresour. Technol. 2019, 292, 121964. [Google Scholar] [CrossRef]
- Bastiaens, L.; Van Roy, S.; Thomassen, G.; Elst, K. Biorefinery of algae: Technical and economic considerations. In Microalgae-Based Biofuels and Bioproducts from Feedstock Cultivation to End-Products; Gonzalez-Fernandez, C., Muñoz, R., Eds.; Woodhead Publishing Series in Energy; Woodhead: Sawston, UK, 2017; pp. 327–345. [Google Scholar]
- Rumbold, K.; van Buijsen, H.J.J.; Overkamp, K.M.; van Groenestijn, J.W.; Punt, P.J.; Werf, M.J.v.d. Microbial production host selection for converting second-generation feedstocks into bioproducts. Microb. Cell Fact. 2009, 8, 64. [Google Scholar] [CrossRef]
- Granata, T.; Egli, M. Biological Nutrients: Sustainable Materials Concept. In ESA Report 2016 (AO/1-7707/13/NL/R), T324-001QT; Volume WP 2100 + WP 2200 Milestone Report V02R01; ESA: Paris, France, 2016; pp. 1–25. [Google Scholar]
- Barbosa, M. Microalgal Photobioreactors: Scale-Up and Optimisation; Wageningen University: Wageningen, The Netherlands, 2003. [Google Scholar]
- Granata, T.; Habermacher, P.; Härri, V.; Egli, M. The influence of bio-optical properties of Emiliania huxleyi and Tetraselmis sp. on biomass and lipid production when exposed to different light spectra and intensities of an adjustable LED array and standard light sources. SN Appl. Sci. 2019, 1, 524. [Google Scholar] [CrossRef]
- Chi, Z.-M.; Liu, G.; Zhao, S.; Li, J.; Peng, Y. Marine yeasts as biocontrol agents and producers of bio-products. Appl. Microbiol. Biotechnol. 2010, 86, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- de Jong, E.; Higson, A.; Walsh, P. Bio-Based Chemicals—Value Added Products from Biorefineries; Volume Task 42, Biorefinery; IEA Bioenergy: Paris, France, 2015; p. 36. [Google Scholar]
- Harmsen, J.; Powell, J.B.; Venderbosch, R.H.; Prins, W. Fast Pyrolysis of Biomass for Energy and Chemicals: Technologies at Various Scales; John Wiley & Sons: Hoboken, NJ, USA, 2019. [Google Scholar]
- Mülhaupt, R. Green polymer chemistry and bio-based plastics: Dreams and reality. Macromol. Chem. Phys. 2013, 214, 159–174. [Google Scholar] [CrossRef]
- Rajagopal, R. Bio-Based Chemicals, Specialities and Polymers; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Au, S.H.; Shihm, S.C.C.; Wheeler, A.R. Integrated microbioreactor for culture and analysis of bacteria, algae and yeast. Biomed Microdevices 2011, 13, 41–50. [Google Scholar] [CrossRef]
- Rosero-Chasoy, G.; Rodríguez-Jasso, R.M.; Aguilar, C.N.; Buitrón, G.; Chairez, I.; Ruiz, H.A. Microbial co-culturing strategies for the production high-value compounds, a reliable framework towards sustainable biorefinery implementation—An overview. Bioresour. Technol. 2021, 321, 124458. [Google Scholar] [CrossRef]
- Sheppard, V.C.; Scheffel, A.; Poulsen, N.; Kröger, N. Live Diatom Silica Immobilization of Multimeric and Redox-Active Enzymes. Appl. Environ. Microbiol. 2012, 78, 211–218. [Google Scholar] [CrossRef]
- Alam, M.A.; Wan, C.; Tran, D.T.; Mofijur, M.; Ahmed, S.F.; Mehmood, M.A.; Shaik, F.; Vo, D.-V.N.; Xu, J. Microalgae binary culture for higher biomass production, nutrients recycling, and efficient harvesting: A review. Environ. Chem. Lett. 2022, 20, 1153–1168. [Google Scholar] [CrossRef]
- Liao, B.Q.; Allen, D.G.; Droppo, I.; Leppard, G.; Liss, S. Surface properties of sludge and their role in bioflocculation and settleability. Water Res. 2001, 35, 339–350. [Google Scholar] [CrossRef]
- Sial, A.; Zhang, B.; Zhang, A.; Liu, K.; Imtiaz, S.A.; Yashir, N. Microalgal–Bacterial Synergistic Interactions and Their Potential Influence in Wastewater Treatment: A Review. BioEnergy Res. 2021, 14, 723–738. [Google Scholar] [CrossRef]
- Saravanan, A.; Kumar, P.S.; Varjani, S.; Jeevanantham, S.; Yaashikaa, P.R.; Thamarai, P.; Abirami, B.; George, C.S. A review on algal-bacterial symbiotic system for effective treatment of wastewater. Chemosphere 2021, 271, 129540. [Google Scholar] [CrossRef]
- Sun, L.; Ma, J.; Li, L.; Tian, Y.; Zhang, Z.; Liao, H.; Li, J.; Tang, W.; He, D. Exploring the essential factors of performance improvement in sludge membrane bioreactor technology coupled with symbiotic algae. Water Res. 2020, 181, 115843. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, W.; Zeng, W.; Yan, Z.; Jia, B.; Li, G.; Liang, H. Organic carbon promotes algae proliferation in membrane-aeration based bacteria-algae symbiosis system (MA-BA). Water Res. 2020, 176, 115736. [Google Scholar] [CrossRef]
- Swain, M.R.; Mishra, J.; Thatoi, H. Bioethanol production from sweet potato (Ipomoea batatas L.) flour using co-culture of Trichoderma sp. and Saccharomyces cerevisiae in solid-state fermentation. Braz. Arch. Biol. Technol. 2013, 56, 171–179. [Google Scholar] [CrossRef]
- Tong, C.Y.; Honsa, K.; Derek, C.J.C. A review on microalgal-bacterial co-culture: The multifaceted role of beneficial bacteria towards enhancement of microalgal metabolite production. Environ. Res. 2023, 228, 15872. [Google Scholar] [CrossRef]
- Rogato, A.; De Tommasi, E. Physical, Chemical, and Genetic Techniques forDiatom Frustule Modification: Applications in Nano technology. Appl. Sci. 2020, 10, 8738. [Google Scholar] [CrossRef]
- Renne, G. Biomineralization. In S&TR; Lawrence Livermore National Lab.: Livermore, CA, USA, 2006; pp. 1–7. [Google Scholar]
- Launey, M.E.; Munch, E.; Alsem, D.H.; Saiz, E.; Tomsia, A.P.; Ritchie, R.O. A novel biomimetic approach to the design of high-performance ceramic–metal composites. J. R. Soc. Interface 2009, 7, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Bennerman, A.D.; Liu, J.; Wan, W. A ‘degradable’ poly(vinyl alcohol) iron oxide nanoparticle hydrogel. Acta Biomater. 2017, 58, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Bettinger, C.J.; Bao, Z. Biomaterials-based organic electronic devices. Polym. Int. 2010, 59, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Adrio, J.; Demain, A. Microbial Enzymes: Tools for Biotechnological Processes. Biomolecules 2014, 4, 117–139. [Google Scholar] [CrossRef]
- Bradbury, J. Nature’s Nanotechnologists: Unveiling the Secrets of Diatoms. PLoS Biol. 2004, 2, e306. [Google Scholar] [CrossRef]
- Bornscheuer, U.T. Chapter 1—Enzymes in Lipid Modification: An Overview. In Lipid Modification by Enzymes and Engineered Microbes; Volume Enzymes in Lipid Modification: An Overview; AOCS Press: Champaign, IL, USA, 2018; pp. 1–9. [Google Scholar]
- Muskovich, M.; Bettinger, C.J. Biomaterials-Based Electronics: Polymers and Interfaces for Biology and Medicine. Adv. Healthc. Mater. 2012, 1, 248–266. [Google Scholar] [CrossRef]
- Wijffels, R.H.; Barbosa, M.J.; Eppink, M.H.M. Microalgae for the production of bulk chemicals and biofuels. Biofuels Bioprod. Biorefin. 2010, 4, 287–295. [Google Scholar] [CrossRef]
- Srivastava, R.K.; Wan, J.U.N. Yeast Species Mediated Bioprocesses and Bio-Products for Biotechnological Application. J. Biotechnol. Biomed. Sci. 2019, 2, 1–11. [Google Scholar] [CrossRef]
- Wu, J.; Elliston, A.; Le Gall, G.; Colquhoun, I.J.; Collins, S.R.A.; Dicks, J.; Roberts, I.N.; Waldron, K.W. Yeast diversity in relation to the production of fuels and chemicals. Sci. Rep. 2017, 7, 14259. [Google Scholar] [CrossRef] [PubMed]
- Kavscek, M.; Strazar, M.; Curk, T.; Natter, K.; Petrovic, U. Yeast as a cell factory: Current state and perspectives. Microb. Cell Fact. 2015, 14, 94. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.G.; Xiao, Y.; Xia, X.X.; Zhao, X.Q.; Peng, L.; Srinophakun, P.; Bai, F.W. Cellulosic ethanol production: Progress, challenges and strategies for solutions. Biotechnol. Adv. 2019, 37, 491–504. [Google Scholar] [CrossRef]
- Clark, J.H.; Deswarte, F.E.I.; Farmer, T.J. The integration of green chemistry into future biorefineries. Biofuels Bioprod. Biorefin. 2009, 3, 72–90. [Google Scholar] [CrossRef]
- Van Haveren, J.; Scott, E.L.; Sanders, J. Bulk chemicals from biomass. Biofuels Bioprod. Biorefin. 2008, 2, 41–57. [Google Scholar] [CrossRef]
- Grigoriev, I.V.; Cullen, D.; Hibbett, D.; Goodwin, S.B.; Jeffries, T.W.; Kubicek, C.P.; Kuske, C.; Magnuson, J.K.; Martin, V.; Spatafora, J.; et al. Fueling the Future with Fungal Genomics; Contract No. DE-AC02-05CH11231; U.S. Department of Energy: Washington, DC, USA, 2011.
- Khanna, N.; Das, D. Biohydrogen production by dark fermentation. Interdiscip. Rev. Energy Environ. 2012, 2, 363–472. [Google Scholar] [CrossRef]
- Benemann, J.; Rocheleu, R. Biohydrogen Production. In Hydrogen Program, Final Summary Report 1996–2000; U.S. Department of Energy: Washington, DC, USA, 2000. [Google Scholar]
- Mohanty, P.; Pant, K.K.; Mittal, R. Hydrogen generation from biomass materials: Challenges and opportunities. WIREs Energy Environ. 2015, 4, 139–155. [Google Scholar] [CrossRef]
- Panagiotopoulou, P.; Papadopoulou, C.; Matralis, H.; Verykios, X. Production of renewable hydrogen by reformation of biofuels. WIREs Energy Environ. 2014, 3, 231–253. [Google Scholar] [CrossRef]
- Pant, D.; Van Bogaert, G.; Diels, L.; Vanbroekhoven, K. A review of the substrates used in microbial fuel cells (MFCs) for sustainable energy production. Bioresour. Technol. 2010, 101, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Dolfin, E.; Scott, J.R.; Edwards, S.R.; Jones, C.; Curtis, T.P. Production of hydrogen from domestic wastewater in a pilot-scale microbial electrolysis cell. Appl. Microbiol. Biotechnol. 2013, 97, 6979–6989. [Google Scholar]
- Monnet, F. An Introduction to Anaerobic Digestion of Organic Wastes; Remade Scotland: Crieff, UK, 2003. [Google Scholar]
- Uggetti, E.; Sialve, B.; Trably, E.; Steyer, J.P. Integrating microalgae production with anaerobic digestion: A biorefinery approach. Biofuels Bioprod. Biorefin. 2014, 8, 516–529. [Google Scholar] [CrossRef]
- Kar, Y.; Şen, N. Catalytic pyrolysis of the oily seeds of Styrax officinalis L. for bio-fuels and valuable industrial chemicals. Environ. Prog. Sustain. Energy 2012, 31, 619–627. [Google Scholar] [CrossRef]
- Wan, W.; Bannerman, A.D.; Yang, L.; Mak, H. Poly(Vinyl Alcohol) Cryogels for Biomedical Applications. In Advances in Polymer Science; Künkel, A., Battagliarin, G., Winnacker, M., Rieger, B., Coates, G., Eds.; Springer Link: Berlin/Heidelberg, Germany, 2014; Volume 263, pp. 283–321. [Google Scholar]
- Wool, R.; Sun, S.X. Affordable Resins and Adhesives from Optimized Soybean Varieties; DEFC07-01ID14217 DOE Final Technical Report 2003; U.S. Department of Energy: Washington, DC, USA, 2004.
- Baker, S.; Harini, B.P.; Rakshith, D.; Satish, S. Marine microbes: Invisible nanofactories. J. Pharm. Res. 2013, 6, 383–388. [Google Scholar] [CrossRef]
- Li, C.; Mezzenga, R. The interplay between carbon nanomaterials and amyloid fibrils in bio-nanotechnology. Nanoscale 2013, 5, 6207–6218. [Google Scholar] [CrossRef]
- Bitton, R.; Bianco-Peled, H. Novel biomimetic adhesives based on algae glue. Macromol. Biosci. 2008, 8, 393–400. [Google Scholar] [CrossRef]
- Molino, P.J.; Chiovitti, A.; Higgins, M.J.; Dugdale, T.M.; Wetherbee, R. Diatom Adhesives: Molecular and Mechanical Properties; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Bamford, N.C.; Howe, P.L. Adhesive Bacterial Exopolysaccharides; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Epstein, L.; Nicholson, R. Adhesion and Adhesives of Fungi and Oomycetes; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Peng, B.L.; Dhar, N.; Liu, H.L.; Tam, K.C. Chemistry and applications of nanocrystalline cellulose and its derivatives: A nanotechnology perspective. Can. J. Chem. Eng. 2011, 89, 1191–1206. [Google Scholar] [CrossRef]
- Spaic, M.; Small, D.P.; Cook, J.R.; Wan, W. Characterization of anionic and cationic functionalized bacterial cellulose nanofibres for controlled release applications. Cellulose 2014, 21, 1529–1540. [Google Scholar] [CrossRef]
- Mathijsen, D. Algae to sustainably produce carbon fiber and simultaneously take CO2 out of the atmosphere. Reinf. Plast. 2020, 64, 50–53. [Google Scholar] [CrossRef]
- Shen, L.; Worrell, E.; Patel, M.K. Comparing life cycle energy and GHG emissions of bio-based PET, recycled PET, PLA, and man-made cellulosics. Biorefining 2012, 6, 625–639. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Er, A.C.; Aiyub, K.; Mohd Yasin, N.H.; Ngan, S.L.; Chew, K.W.; Khoo, K.S.; Ling, T.C.; Juan, J.C.; Ma, Z.; et al. Current status and perspectives of algae-based bioplastics: A reviewed potential for sustainability. Algal Res. 2023, 71, 103078. [Google Scholar] [CrossRef]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Alvarenga, R.A.F.; Dewulf, J.; De Meester, S.; Wathelet, A.; Villers, J.; Thommeret, R.; Hruska, Z. Life cycle assessment of bioethanol-based PVC. Biofuels Bioprod. Biorefin. 2013, 7, 386–395. [Google Scholar] [CrossRef]
- Parirokh, M.; Torabinejad, M. 10—Calcium Silicate–Based Cements. In Mineral Trioxide Aggregate: Properties and Clinical Applications; Torabinejad, M., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2014; p. 342. [Google Scholar]
- Rieger, J.; Kellermeier, M.; Nicoleau, L. Die Bildung von Nanopartikeln und Nanostrukturen—CaCO3, Zement und Polymere aus Sicht der Industrie. Angew. Chem. 2014, 126, 12586–12603. [Google Scholar] [CrossRef]
- Lu, X. Polymer/Calcium Carbonare Nanocomponents; Woodhead Publishing: Sawston, UK, 2006. [Google Scholar]
- Pettinari, C.; Tombesi, A. Metal–organic frameworks for carbon dioxide capture. MRS Energy Sustain. 2020, 7, 35. [Google Scholar] [CrossRef]
- Sevilla, M.; Falco, C.; Titirici, M.-M.; Fuertes, A.B. High-performance CO2 sorbents from algae. J. Mater. Chem. A 2019, 7, 17466. [Google Scholar] [CrossRef]
- Osman, A.I.; Hefny, M.; Maksoud, M.I.A.; Elgarahy, A.M.; Rooney, D.W. Recent advances in carbon capture storage and utilization technologies: A review. Environ. Chem Lett. 2021, 19, 797–849. [Google Scholar] [CrossRef]
- IPCC. Special Report on Carbon Dioxide Capture and Storage; Metz, B., Davidson, O., de Coninck, H.C., Loos, M., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2021; p. 450. [Google Scholar]
- Song, C.; Liu, Q.; Qi, Y.; Chen, G.; Song, Y.; Kansha, Y.; Kitamura, Y. Absorption-microalgae hybrid CO2 capture and biotransformation strategy—A review. Int. J. Greenhouse Gas Control 2019, 88, 109–111. [Google Scholar] [CrossRef]
- Raganati, F.; Miccio, F.; Ammendola, P. Adsorption of Carbon Dioxide for Post-combustion Capture: A Review. Energy Fuels 2021, 35, 12845–12868. [Google Scholar] [CrossRef]
- Cardias, B.B.; de Morais, M.G.; Costa, J.A.V. CO2 conversion by the integration of biological and chemical methods: Spirulina sp. LEB 18 cultivation with diethanolamine and potassium carbonate addition. Bioresour. Technol. 2018, 267, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Granata, T.; Follonier, C.; Burkhardt, C.; Rattenbacher, B. Methods for Oxygenation of Continuous Cultures of Brewer’s Yeast, Saccharomyces cerevisiae. Fermentation 2021, 7, 282. [Google Scholar] [CrossRef]
- Guillard, R. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W., Chanley, M., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Granata, T.; Rattenbacher, B.; John, G. Micro-Bioreactors in Space: Case Study of a Yeast (Saccharomyces cerevisiae) Bioreactor with a Non-Invasive Monitoring Method. Front. Space Technol. 2022, 2, 773814. [Google Scholar] [CrossRef]
- Snoeyink, V.L.; Jenkis, D. Water Chemistry; Johm Wiley & Sons: Hoboken, NJ, USA, 1980; p. 463. [Google Scholar]
- Ibrahim, M.H.; El-Naas, M.H.; Zhang, Z.; Van der Bruggen, B. CO2 Capture Using Hollow Fiber Membranes: A Review of Membrane Wetting. Energy Fuels 2018, 32, 963–978. [Google Scholar] [CrossRef]
- Montoya, P. Membrane Gas Exchange; MedArray, Inc.: Ann Arbor, MI, USA, 2010; pp. 1–7. [Google Scholar]
- Hafeez, S.; Safdar, T.; Pallari, E.; Manos, G.; Aristodemou, E.; Zhang, Z.; Al-Salem, S.M.; Achilleas Constantinou, A. CO2 capture using membrane contactors: A systematic literature review. Front. Chem. Sci. Eng. 2021, 15, 720–754. [Google Scholar] [CrossRef]
- Rosenfeld, E.; Beauvoit, B.; Rigoulet, M.; Salmon, J.M. Non-respiratory oxygen consumption pathways in anaerobically-grown Saccharomyces cerevisiae: Evidence and partial characterization. Yeast 2002, 19, 1299–1321. [Google Scholar] [CrossRef] [PubMed]
- Abiusi, F.; Sampietro, G.; Marturano, G.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; Tredici, M. Growth, photosynthetic efficiency, and biochemical composition of Tetraselmis suecica F&M-M33 grown with LEDs of different colors. Biotechnol. Bioeng. 2013, 111, 956–964. [Google Scholar] [PubMed]
- Pereira, H.; Páramo, J.; Silva, J.; Marques, A.; Barros, A.; Maurício, D.; Santos, T.; Schulze, P.; Barros, R.; Gouveia, L.; et al. Scale-up and large-scale production of Tetraselmis sp. CTP4 (Chlorophyta) for CO2 mitigation: From an agar plate to 100-m3 industrial photobioreactors. Sci. Rep. 2018, 8, 5112. [Google Scholar] [CrossRef] [PubMed]
- Rafay, R.; Uratani, J.M.; Hernandez, H.H.; Rodríguez, J. Growth and Nitrate Uptake in Nannochloropsis gaditana and Tetraselmis chuii Cultures Grown in Sequential Batch Reactors. Front. Mar. Sci. 2020, 7, 77. [Google Scholar] [CrossRef]
- Lee, W.-K.; Ryu, Y.-K.; Choi, W.-Y.; Kim, T.; Park, A.; Lee, Y.-J.; Jeong, Y.; Lee, C.-G.; Kang, D.-H. Year-Round Cultivation of Tetraselmis sp. for Essential Lipid Production in a Semi-Open Raceway System. Mar. Drugs 2021, 19, 314. [Google Scholar] [CrossRef] [PubMed]
- Nanninga, H.; Tyrrell, E. Importance of light for the formation of algal blooms by Emiliania huxleyi. Mar. Ecol. Prog. Ser. 1996, 136, 195–203. [Google Scholar] [CrossRef]
- Kim, H.-W.; Cheng, J.; Rittmann, B.E. Direct membrane-carbonation photobioreactor producing photoautotrophic biomass via carbon dioxide transfer and nutrient removal. Bioresour. Technol. 2016, 204, 32–37. [Google Scholar] [CrossRef]
- Lai, J.; Yusoff, F.M.; Shariff, M. Large-scale culture of a tropical marine microalga Chaetoceros calcitrans Paulsen Takano 1968 at different temperatures using annular photobioreactors. Pak. J. Biol. Sci. 2012, 15, 635–640. [Google Scholar] [CrossRef]
- Granata, T. Dependency of Microalgal Production on Biomass and the Relationship to Yield and Bioreactor Scale-up for Biofuels: A Statistical Analysis of 60+ Years of Algal Bioreactor Data. Bioenergy Res. 2017, 10, 267–287. [Google Scholar] [CrossRef]
- Scott, J.P.R.; Green, D.A.; Weerts, G.; Cheuvront, S.N. Body size and its implications upon resource utilization during human space exploration missions. Nat. Sci. Rep. 2020, 10, 13836. [Google Scholar] [CrossRef]
- Verbelen, P.J.; Depraetere, S.A.; Winderickx, J.; Delvaux, F.R.; Delvaux, F. The influence of yeast oxygenation prior to brewery fermentation on yeast metabolism and the oxidative stress response. FEMS Yeast Res. 2009, 9, 226–239. [Google Scholar] [CrossRef]
- Gomar-Alba, M.; Morcillo-Parra, M.A.; Olmo, M.L. Response of yeast cells to high glucose involves molecular and physiological differences when compared to other osmostress conditions. FEMS Yeast Res. 2015, 15, fov039. [Google Scholar] [CrossRef]
- Vieira, E.D.; Andrietta, M.-d.-G.; Andrietta, S.R. Yeast biomass production: A new approach in glucose-limited feeding strategy. Braz. J. Microbiol. 2013, 44, 551–558. [Google Scholar] [CrossRef]
- Herrero, E.; Ros, J.; Bellí, G.; Cabiscol, E. Redox control and oxidative stress in yeast cells. Biochim. Biophys. Acta 2008, 1780, 1217–1235. [Google Scholar] [CrossRef]
- Pancha, I.; Chokshi, K.; George, B.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Nitrogen stress triggered biochemical and morphological changes in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2014, 156, 146–154. [Google Scholar] [CrossRef]
- Takagi, H. Metabolic regulatory mechanisms and physiological roles of functional amino acids and their applications in yeast. Biosci. Biotechnol. Biochem. 2019, 83, 1449–1462. [Google Scholar] [CrossRef]
- Fernandez, E.; Blanch, W.M.; Marañonm, E.; Holligan, P. High rates of lipid biosynthesis in cultured, mesocosm and coastal populations of the coccolithophore Emiliania huxleyi. Mar. Ecol. Prog. Ser. 1994, 114, 13–22. [Google Scholar] [CrossRef]
- Jusoh, M.; Loh, S.H.; Chuah, T.S.; Aziz, A.; Cha, T.S. Indole-3-acetic acid (IAA) induced changes in oil content, fatty acid profiles and expression of four fatty acid biosynthetic genes in Chlorella vulgaris at early stationary growth phase. Phytochemistry 2015, 111, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. 2008, 54, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Llorente, B.; Williams, T.C.; Goold, H.D.; Pretorius, I.S.; Paulsen, I.T. Harnessing bioengineered microbes as a versatile platform for space nutrition. Nat. Commun. 2022, 13, 6177. [Google Scholar] [CrossRef] [PubMed]
- Nutts, R.U. Reinheitsgebot; Duke, W.-I., Ed.; Munich, 1516; Oxford University Press: Oxford, UK, 2013. [Google Scholar]


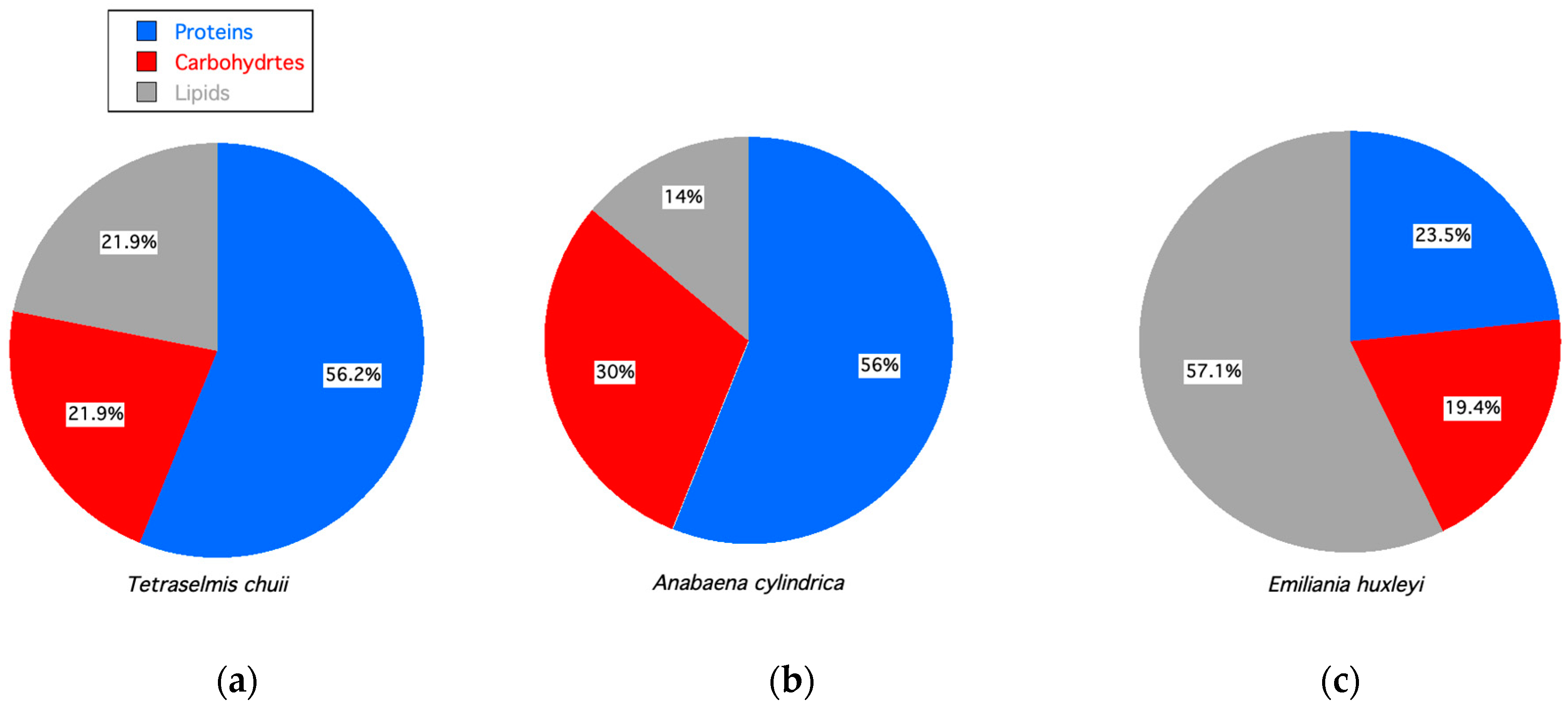
| Products | Bionutrient | Application | Ref. |
|---|---|---|---|
| Biomass | Cell wall, vacuole contents, nucleic acids | Health food, functional food feed additive, aquaculture soil conditioner | [5] |
| Colorants, antioxidants | Xanthophylls-astaxanthin, canthaxanthin β-carotene, vitamins C and E Fucoxanthin, lutein | Food, feed additives Cosmetics | [5] [6] |
| Fatty acids- | AA-Arachidonic acid, EPA -Eicosapentaenoic acid, DHA -Docosahexaenoic acid, GCA-γ-linolenic acid LA -Linoleic acid | Food additives | [5] |
| Enzymes | Superoxide dismutase-SOD Phosphoglycerate kinase-PGK Luciferase and Luciferin Restriction enzymes | Health food, research, medicine | [5] [7] |
| Polymers | Polysaccharides, starch, poly-β-hydroxybutyric acid-PHB | Food additive, cosmetics medicine | [5] |
| Special products | Peptides, toxins, isotopes, amino acids (proline, arginine, aspartic acid) | Research, medicine, fertilizers | [5] |
| Other bionutrients | Vinyl polymers, polyesters, polyamides, polyurethanes, and synthetic rubbers vinyl polymers, polyesters, polyamides, polyurethanes, and synthetic rubbers | Industry | [8,9] |
| Saturated and unsaturated fats | Biofuel/Transport | [10] | |
| Sugars starches | Industry | [11] |
| Biological Process | Energy Sources | Inputs | Synthesis—Outputs | Organisms |
|---|---|---|---|---|
| Autotrophic | ||||
| Photo-lithotrophic | Light | CO2, H2O, H2S, H2, nutrients | Carbohydrate (sugars, starches) lipids/oil (fuel), enzymes—various pathways | Vascular plants, Algae, Cyanobacteria, Chlorobiaceae, Chromaticaceae |
| Heterotrophic | ||||
| Chemo- organo- heterotrophic | Organic Oxidation | CO2 and H2 from Organics (Glucose, Pyruvate) | Carbohydrates/sugars/starch, lipids oil (fuel), enzymes—various pathways, H2, CH4, ethanol/lactic acids | Majority of Bacteria, Fungi, Methanogens, Fermenters |
| Periods | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|
| Time (Days) | 1–2 | 2.5–3.5 | 4–5 | 5.5–7.5 | 8–10 |
| Yeast | |||||
| μ (d−1) | 0.21 | 0.19 | 0.07 | 0.04 | 0.05 |
| r2 | 0.975 | 0.944 | 0.977 | 0.948 | 0.995 |
| OUR (g (L d−1)) | 8.6 | 8.6 | 8.6 | 8.6 | 8.6 |
| Λ (mg g−1) | 0.65 ± 0.04 | 0.53 ± 0.02 | 0.49 ± 0.11 | 0.47 ± 0.1 | 0.43 ± 0.01 |
| Algae | 1–2.5 | 2.5–4.4 | 4.4–4.8 | 5–7 | 7–10 |
| μ (d−1) | 0.32 | 0.34 | N.A. | 0.20 | 0.05 |
| r2 | 0.983 | 0.995 | - | 0.995 | 0.971 |
| P (mg (L d−1) | 3.4 | 3.4 | - | 3.4 | 3.4 |
| Λ (mg g−1) | 3.08 ± 0.39 | 1.83 ± 0.28 | - | 1.39 ± 0.09 | 1.04 ± 0.02 |
| Yeast Time Series | pH | CO2 gas (atm) | [CO2 aq] (M) | [HCO3−] (M) | [CO32−] (M) | Ctotal (M) | TA 1 (M) |
|---|---|---|---|---|---|---|---|
| Mean | 5.52 | 3.2 × 10−4 | 8.12 × 10−5 | 1.20 × 10−5 | 1.26 × 10−9 | 9.32 × 10−5 | 1.20 × 102 |
| +SD | 5.76 | 4.40 × 10−4 | 7.44 × 10−5 | 1.91 × 10−5 | 2.00 × 10−9 | 9.35 × 10−5 | 1.91 × 10−2 |
| −SD | 5.28 | 2.26 × 10−3 | 1.19 × 10−8 | 1.73 × 10−7 | 5.59 × 10−10 | 1.85 × 10−7 | 1.74 × 10−4 |
| Algae Time Series | |||||||
| Mean | 7.75 | 2.40 × 10−3 | 4.08 × 10−11 | 1.7 × 10−7 | 5.59 × 10−10 | 1.73 × 10−7 | 1.74 × 10−4 |
| +SD | 8.5 | 2.20 × 10−3 | 6.76 × 10−7 | 9.53 × 10−5 | 1.00 × 10−8 | 9.60 × 10−5 | 9.50 × 10−2 |
| −SD | 6.93 | 2.60 × 10−3 | 4.40 × 10−5 | 1.67 × 10−4 | 1.75 × 10−8 | 2.10 × 10−4 | 1.67 × 10−1 |
| Algae CCU | |||||||
| 5.50 | 1.0 × 10−1 | 3.38 × 10−3 | 4.77 × 10−4 | 5.01 × 10−8 | 3.86 × 10−3 | 3.86 × 10−3 | |
| 6.52 | 2.60 × 10−2 | 8.80 × 10−4 | 1.30 × 10−3 | 1.36 × 10−7 | 2.18 × 10−3 | 2.18 × 10−3 | |
| 6.70 | 1.60 × 10−2 | 5.41 × 10−4 | 1.21 × 10−3 | 1.27 × 10−7 | 1.75 × 10−3 | 1.75 × 10−3 | |
| 7.00 | 2.20 × 10−3 | 7.44 × 10−5 | 3.32 × 10−4 | 3.48 × 10−8 | 4.06 × 10−4 | 4.06 × 10−2 | |
| 7.11 | 1.60 × 10−3 | 5.41 × 10−5 | 3.11 × 10−4 | 3.26 × 10−8 | 3.65 × 10−4 | 3.65 × 10−2 | |
| 7.50 | 1.50 × 10−3 | 5.07 × 10−5 | 7.15 × 10−4 | 7.51 × 10−8 | 7.66 × 10−4 | 7.66 × 10−2 | |
| 8.50 | 2.20 × 10−4 | 7.44 × 10−6 | 1.05 × 10−3 | 1.10 × 10−7 | 1.06 × 10−3 | 1.06 × 10−3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Granata, T.; Rattenbacher, B.; Kehl, F.; Egli, M. Microbial Factories and Exploiting Synergies of Bioreactor Technologies to Produce Bioproducts. Fermentation 2024, 10, 135. https://doi.org/10.3390/fermentation10030135
Granata T, Rattenbacher B, Kehl F, Egli M. Microbial Factories and Exploiting Synergies of Bioreactor Technologies to Produce Bioproducts. Fermentation. 2024; 10(3):135. https://doi.org/10.3390/fermentation10030135
Chicago/Turabian StyleGranata, Tim, Bernd Rattenbacher, Florian Kehl, and Marcel Egli. 2024. "Microbial Factories and Exploiting Synergies of Bioreactor Technologies to Produce Bioproducts" Fermentation 10, no. 3: 135. https://doi.org/10.3390/fermentation10030135
APA StyleGranata, T., Rattenbacher, B., Kehl, F., & Egli, M. (2024). Microbial Factories and Exploiting Synergies of Bioreactor Technologies to Produce Bioproducts. Fermentation, 10(3), 135. https://doi.org/10.3390/fermentation10030135