Abstract
Studying non-Saccharomyces yeasts as bio-protectors can help find new alternatives to the chemical additive SO2 in winemaking. The present article evaluates the effect of two native yeasts, Candida oleophila and Candida boidinii, as potential bio-protectors to replace SO2 during the production of Sauvignon Blanc wine. Fermentation was conducted on simple and mixed inoculum at two concentrations, 1 × 106 and 1 × 107 cells/mL. We monitored the population of deterioration microorganisms, including lactic acid bacteria (LAB), acetic acid bacteria (AAB), and Brettanomyces bruxellensis (BB), apart from the final chemical and volatile composition of the wine. The results were compared with fermentations protected with SO2, where Candida oleophila yeast was more effective against lactic acid bacteria, whereas Candida boidinii was more effective against acetic acid bacteria and Brettanomyces bruxellensis; meanwhile, the fermentations with the initial inoculum of 1 × 107 cells/mL showed better results than those with an inoculum of 1 × 106 cells/mL. Bio-protector use did not negatively affect wine quality, equaling the effectiveness of SO2 for spoilage microorganism inhibition. This study reveals for the first time the potential of Candida oleophila and Candida boidinii yeasts as bio-protectors in microbiological wine stabilization.
1. Introduction
Sulfur dioxide (SO2) has played a key role in the wine industry due to its antimicrobial, antioxidant, and antioxidase properties, inhibiting the development of spoilage microorganisms including yeasts, lactic acid bacteria (LAB), and acetic acid bacteria (AAB) [1]; keeping the wine from browning after reacting with hydrogen peroxide [2]; and reducing wine quinones to their phenolic form [3]. However, recent decades have seen concerns raised related to its use in foods due to possible pseudo-allergic reactions associated with its consumption, including dermatitis, rashes, anaphylaxis, and even abdominal pain [4], which has led to limitations on its use in winemaking. The World Health Organization has established a maximum daily acceptable sulfur dioxide intake of 0.7 mg/kg of body weight. At the same time, the International Wine and Winery Organization (OIV) has progressively limited the maximum allowable SO2 concentrations, currently set at 150 mg/L for red wines and 200 mg/L for white wines [5,6]. Along with growing public demand for food without chemical additives [7], there is currently a significant change in enological research where bio-protection has arisen as a promising alternative that can either partially or wholly replace SO2 in the industry [8,9,10].
Bio-protection in winemaking is defined as using antagonistic microorganisms added into grapes to inhibit spoilage microorganisms and/or extend the shelf life of wine without any negative impacts on its sensorial properties [11,12]. Its action mechanism is grounded in competition for nutrients by creating an ecological niche where bio-protective yeasts can occupy a space in the grape and affect other microorganisms via nutrient limitation [13]. Bio-protection can appear in two modalities: active and passive competition strategies [14]. Active competition is performed by bacteria, which dominate the fermentation process by being stronger and presenting unique traits like killer toxins, while passive competition is performed by all microorganisms that consume nutrients over others [15].
Most current studies on bio-protection with NSY have been conducted on red grape must, with Merlot, Tempranillo, and Pinot Noir grapes using the yeasts Torulaspora delbrueckii, Metschnikowia pulcherrima, and Lachancea thermotolerans [16,17,18,19,20,21,22,23,24]. This has been because these yeasts have unique characteristics as controlling agents; M. pulcherrima has an active bio-protection capacity due to synthesizing pulcherrimine, a proven antifungal and antimicrobial agent [25]; T. delbrueckii can produce killer proteins that control the population of deterioration bacteria [26]; and L. thermotolerans has a high antioxidant capacity that can dominate fermentation in the face of high oxygen concentrations in the must [27]. T. delbrueckii and M. pulcherrima are NSY strains that are currently available commercially as bio-protectors to be used in winemaking, whether in a mixed format or in a monoculture format. However, other commercial bio-protectors are also formed with Lactobacillus Plantarum bacteria as a bio-controlling agent.
Many non-saccharomyces yeasts have not been studied as bio-protectors, despite proven antimicrobial potential in areas apart from winemaking. For instance, the NSY C. oleophila has proven potential with food, competing for nutrients and space, and leading to the cell death of spoilage microorganisms, particularly Penicillium spp. during the harvest of different fruits and vegetables [28,29,30,31]. The yeast Candida boidinii has also shown antimicrobial potential for table olive production, cutting undesirable bacteria from oil, and granting sensory advantages to the final product [32]. There is currently only one study about these yeasts in winemaking, concerning their fermentation potential and capacity to reduce ethanol in wine [33].
Given the lack of existing literature on bio-protection in Sauvignon must and the few studies carried out with Candida spp. in winemaking, and in particular regarding the species Candida oleophila and Candida boidinii; the objective of this study is to evaluate the impact of bio-protection as a replacement for SO2 in the microbiological profile of Sauvignon Blanc wine using Candida oleophila and Candida boidinii as bio-protectors under two inoculation modalities: simple and mixed, and at two initial inoculation concentrations: 1 × 106 and 1 × 107 cells/mL. Major wine spoilage microorganism populations were monitored, including lactic bacteria, acetic bacteria, and Brettanomyces bruxellensis, along with the final physio-chemical and volatile composition of the wine, comparing its antimicrobial effect and final quality with wines protected using sulfites.
2. Materials and Methods
2.1. Yeasts
The Candida boidinii (NSY 2) and Candida oleophila (NSY 9) yeasts used were obtained from the microorganism collection at the food fermentation lab in PUC/UTFSM. The yeasts were isolated from grapes gathered at the Palo Alto Winery, located in the Maule Region of Chile (35°26′ S 71°40′ W), and identified via partial rDNA 26S sequencing [34]. The yeast cultures were kept in stock format in a glycerol medium at a temperature of −80 °C, for subsequent activation on agar plates (Yeast and Mould Agar, Oxoid, Madrid, Spain), which were refrigerated at 4 °C until the moment of inoculation. The fermentation yeast chosen was the commercial strain of S. cerevisiae EC 1118™ (Lallemand, Montréal, QC, Canada) which was kept inactive under refrigeration at 4 °C until its inoculation time as per manufacturers’ instructions.
2.2. Grape Juice
We used the 2023 harvest of Vitis vinifera ‘Sauvignon Blanc’ grapes from the Los Robles farm of Viña Emiliana (34°36′ S 71°7′ W), without sulfites, stored in 2 kg bags at a temperature of −18 °C until the moment of use. This was followed by a process of removing sticks, leaves, stems, and insects attached to the fruit, and then the grapes were pressed to obtain the grape juice (22.5 °Brix, density 1.095 g/mL, pH 3.18, titratable acidity 5.6 g/L tartaric acid). The grape juice was conditioned via refrigeration at 4 °C for 20 h.
2.3. Winemaking Assays
The tests were planned with the objective of studying the bio-protective effect of the non-saccharomyces inoculation on wines without sulfites and comparing them with the effect of fermentations conducted using SO2, along with evaluating the effect of the concentration of the bio-protective inoculation. To this end, wine was made using white Sauvignon Blanc grapes in triplicate in 1 L Schott bottles using the following inoculation strategies:
- Inoculation of the bio-protective initiator Candida spp. (C. oleophila or C. boidinii) at 1 × 106 cells/mL, without adding SO2 and with inoculation at 72 h with commercial S. cerevisiae yeast at a dose of 1 × 106 cells/mL.
- Inoculation of the bio-protective initiator Candida spp. (C. oleophila or C. boidinii) at 1 × 107 cells/mL, without adding SO2 and inoculation at 72 h with commercial S. cerevisiae yeast at a dose of 1 × 106 cells/mL.
- Inoculation of the mixed bio-protective initiator C. oleophila/C. boidinii 50:50 at 1 × 107 cells/mL, without adding SO2 and inoculation at 72 h with commercial S. cerevisiae yeast at a dose of 1 × 106 cells/mL.
- Initial spontaneous fermentation was initiated by adding 40 ppm of SO2 and inoculating for 72 h with commercial S. cerevisiae at a dose of 1 × 106 cells/mL.
- Initial spontaneous fermentation without addition of SO2 and inoculation at 72 h with commercial S. cerevisiae yeast at a dose of 1 × 106 cells/mL.
2.3.1. Preparation of Initial Inoculum
The colonies of Candida spp. yeast stored on agar plates at 4 °C were passed into a tube with 35 mL of liquid Sabouraud Dextrose Broth medium (Biokar, Pantin, France), which was incubated at 28 °C for 48 h. This was followed by separation via centrifuging at 6000 rpm for 10 min, eliminating the supernatant under a lighter, and washing twice with peptone water (1%) before inoculation.
2.3.2. Experiment Montage
The grape juice was distributed in modified Schott bottles maintaining sterility in a laminar flow hood. Each fermentation was carried out in triplicate to verify experimental reproducibility. The flasks were adapted with an additional aperture, allowing for the insertion of a probe to avoid contamination and air exposure while taking samples, and letting O2 in during initial conditioning. Each Schott bottle with 1 L of grape juice was subsequently put through an airing process for 10 min via an air pump, supplying a flow of 0.940 L/min to ensure oxygen saturation in the medium. Finally, the flasks were inoculated with the bio-protective yeast (106 or 107 cells/mL) or with the control using SO2 at 40 ppm, depending on the case, and placed in the incubating shaker at 10 °C for cold pre-fermentative maceration over 72 h. Finally, the commercial yeast S. cerevisiae was inoculated at a dose of 106 cells/mL to start alcoholic fermentation.
2.3.3. Fermentation Follow-Up
For each fermentation, 12 mL samples were taken in triplicate every 24 h, measuring density, brix° degrees, and optic density (OD600). The remaining samples were stored at a freezer temperature of −18 °C to corroborate measurements if necessary. Measurements continued until all sugars in the medium were consumed, which occurs when the density reaches values below 0.994 g/mL or does not remain constant for 3 consecutive days.
2.4. Microbiological Analysis
2.4.1. Sample Serial Dilution
For the microbiological analysis, 15 mL samples were taken on 5 occasions: initial must, after 24 h of pre-fermentative maceration, after 72 h of pre-fermentative maceration, halfway through alcoholic fermentation (day 6 or 8), and at the end of alcoholic fermentation (AF). Samples were collected in sterile Falcon tubes and were serially diluted in 0.01% peptone water (Sigma Aldrich, St. Louis, MO, USA). First, 1 mL of the sample was pipetted into a sterile 9 mL peptone water tube. The tube was stirred with a “vortex” shaker for 20 s. Then, 1 mL was transferred to the next 9 mL sterile peptone water tube. The process was repeated until 7 decimal dilutions were completed according to the OIV protocol [35]. Finally, 0.1 mL of each dilution was inoculated on agar plates with different media depending on the microorganism. Additionally, for the results to be statistically valid, each dilution was inoculated in triplicate.
2.4.2. Microorganism Media
For the total yeast count, we used Yeast Glucose Chloramphenicol Agar (YCA) medium (20 g/L of glucose, 5 g/L of yeast extract, 0.1 g/L of chloramphenicol, 15 g/L of bacteriological agar, 100 mg/L of chloramphenicol, and 150 mg/L of biphenyl).
Populations of B. bruxellensis were determined in a specific medium composed of 4 g/L of yeast extract, 5 g/L of digested pancreatic casein, 5 g/L of dextrose, 0.55 g/L of monopotassium phosphate, 0.425 g/L of potassium chloride, 0.125 g/L of calcium chloride, 0.125 g/L of magnesium sulfate, 0.0025 g/L of ferric chloride, 0.0025 g/L of manganese sulfate, 20 g/L of agar, 0.022 g/L of bromocresol green, 10 mL/L of cycloheximide, and 10 mL/L of chloramphenicol.
Acetic acid bacteria were counted in Agar WLD medium (4 g/L of yeast extract, 5 g/L of digested pancreatic casein, 5 g/L of dextrose, 0.55 g/L of monopotassium phosphate, 0.425 g/L of potassium chloride, 0.125 g/L of calcium chloride, 0.125 g/L of magnesium sulfate, 0.0025 g/L of ferric chloride, 0.0025 g/L of manganese sulfate, 20 g/L of agar, 0.022 g/L of bromocresol green, 200 μL of cycloheximide, and 12.5 mg/L of penicillin).
Lactic acid bacteria were counted in MRS Agar medium (20 g/L of glucose, 10 g/L of meat extract, 4 g/L of yeast extract, 2 g/L of potassium dihydrogen phosphate, 5 g/L of sodium acetate, 2 g/L of ammonium citrate, 0.2 g/L of magnesium sulfate ∙ 6 H2O, 0.05 g/L of manganese sulfate ∙ 4 H2O, 1.08 mL/L of Tween 80, 16 g/L of bacteriological agar, 10 g/L of digested pancreatic casein, and 100 mg/L of natamycin) and the non-Saccharomyces yeasts were counted in agar lysin medium (20 g/L of bacteriological agar, 2.5 g/L lysine hydrochloride, 11.75 g/L of yeast carbon base, and 100 mg/L of chloramphenicol). It is necessary to mention that each medium is specific since it contains antibiotics that inhibit the growth of other microorganisms. Chloramphenicol inhibits bacteria, cycloheximide and natamycin inhibit yeasts, and penicillin inhibits lactic acid bacteria.
2.4.3. Sample Incubation and Counting Colonies
Yeast and acetic acid bacteria plates were stored upside down under aerobic conditions for 48 h at 28 °C, while lactic acid bacteria plates were incubated anaerobically at the same time and temperature conditions using gas packs (Oxoid) to avoid the growth of acetic bacteria.
To conduct the counting, the dilution chosen must have between 10 and 300 colonies, which are isolated enough to be counted. The number of microorganisms N present in CFU/mL units is obtained from the following equation [35]:
where
- is the sum of colonies counted on a plate.
- is the number of replies, in this case 3.
- is the dilution [d = 1 when the undiluted liquid product (test sample) is retained].
- is the volume of the inoculum placed in each plate, in this case 0.1 mL.
The analysis of results will mainly be carried out during the pre-ferment stage (first 72 h), as there is minimal influence from other factors that could alter the results, such as Saccharomyces cerevisiae yeast inoculated on the third day and metabolites produced during must fermentation, such as ethanol.
2.5. Chemical Composition Analysis
The final samples of wine were chemically characterized. Ethanol volume (%vol) and tartaric acid g/L were determined via a high-pressure liquid chromatography unit (HPLC) (Agilent Technologies, Infinity 1260, Waldbronn, Germany), using a 300 × 7.8 mm Aminex HPX-87 chromatography column (BIORAD, Hercules, CA, USA), operating with a 210 nm wavelength DAD detector. The measurements were taken with a run time of 30 min, a sample injection volume of 20 μL, the mobile phase of H2SO4 with a concentration of 0.005 mMol, and a flow of 0.6 mL/min. The rest of the compounds, such as acetic acid g/L, glycerol g/L, total sugar concentration (glucose–fructose) g/L, malic acid g/L, and lactic acid g/L, were determined using a Biosystems Y-15 Enology Automatic Analysis System (Biosystems, Barcelona, Spain). Prior to the detection, Y-15 was calibrated using external standards, which were technically supported by the Biosystems Enterprise (Cumming, GA, USA).
2.6. Volatile Composition Analysis
Volatile compounds (VOCs) were identified in triplicate via the samples obtained at the end of alcoholic fermentation via mass spectrophotometry (gas chromatography) together with a mass detector with solid phase microextraction (GC-MS-SPME). To begin, 3 mL of the sample was transferred to a 20 mL gas chromatography (GC) vial with magnetic aluminum tape and a septum of Teflon/PTFE, adding 1 mL of NaCl-saturated solution. Samples were stored frozen (−20 °C) until analysis. Then, the samples were thawed and heated at 50 °C for 10 min with agitation at 300 rpm, before beginning the extraction phase for volatile compounds where it used an SPME fiber for 15 min. Following extraction, the SPME fiber was inserted in the chromatograph injector configured at 250 °C in splitless mode for 5 min. An automatic fiber (DVB-CAR-PDMS) from Supelco with serial number 57298U was used for this process. Chromatographic analysis was performed on a Thermo unit, Triplus 1310 model, using an ISQ LT mass detector with an RTX5MS (30 m × 0.25 mm ID × 0.25 μm film) (ISQ LT GC-MS, Thermo Fisher Scientific Inc., Waltham, USA) column and helium as the carrying gas at 1 mL/min. The chromatographic program began at 40 °C for 5 min, rising to 10 °C/min until reaching a temperature of 250 °C and remaining there for 10 min. The injector operates in electron impact mode at 70 eV with a source temperature of 250 °C. Mass spectra and reconstructed chromatograms (total ion current [TIC]) were obtained via automatic scanning in the mass range m/z 35–500. VOC profiles were analyzed with Chromeleon 7.3 software to integrate peaks and the NIST main library was utilized to identify compounds by comparing their mass spectral profiles with those in the 2017 NIST database. The relative abundance of a particular compound was quantified using its peak area (or the sum of individual peak areas) regarding the total [36].
2.7. Data Analysis
2.7.1. ANOVA Analysis and Post hoc Tukey Test for Chemical Compounds
Analyses of variance (ANOVA) and Tukey tests (significance level 95%, p ≤ 0.05) were performed for the chemical compounds, treating them as dependent variables, while the fermentative strategies were the independent variables. The aim was to test the statistical significance of differences between all fermentation strategies. Both tests were conducted using Minitab Statistical 21 software.
2.7.2. PCA for Volatile Compounds
We performed principal component analysis (PCA) to simplify the volatile compound’s data dimensionality, thereby obtaining 2 principal dimensions that explain the data’s largest variation and the most amount of information. The data treatment was performed prior to analysis, given that each of the volatile compounds is associated with a number called the match factor (SI), indicating the degree of similarity between the measured chromatogram and the NIST library. Accordingly, compounds associated with an SI factor greater than 800 were considered, indicating good to excellent reliability [37]. In the samples, 27 compounds were identified; however, only 7 volatile compounds accounted for at least 80% of the accumulative abundance. Despite this, it was decided to analyze all compounds to obtain results that provide the maximum amount of information associated with the aromatic composition. The Minitab Statistical 21 software was used to calculate the correlation matrix, defining the main components as those whose eigenvalues are greater than 1 and graphing them with biplots. Finally, to evaluate whether there were significant differences in the production of each volatile compound identified between the different modalities studied, we ran a post hoc Tukey test, with a significance level of 95% (p ≤ 0.05), in the same way as with the chemical compounds.
3. Results
3.1. Kinetics of Alcoholic Fermentation (AF)
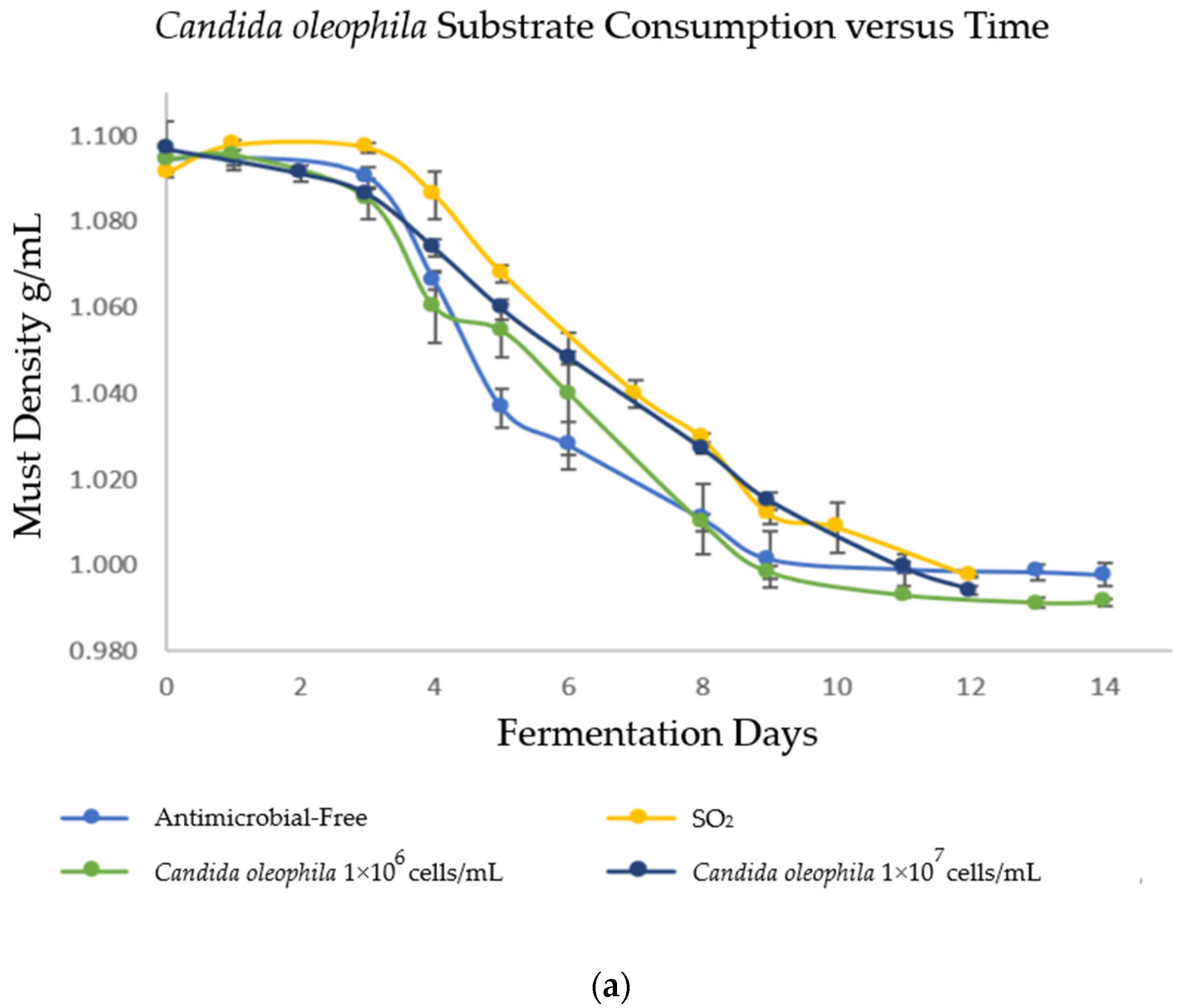
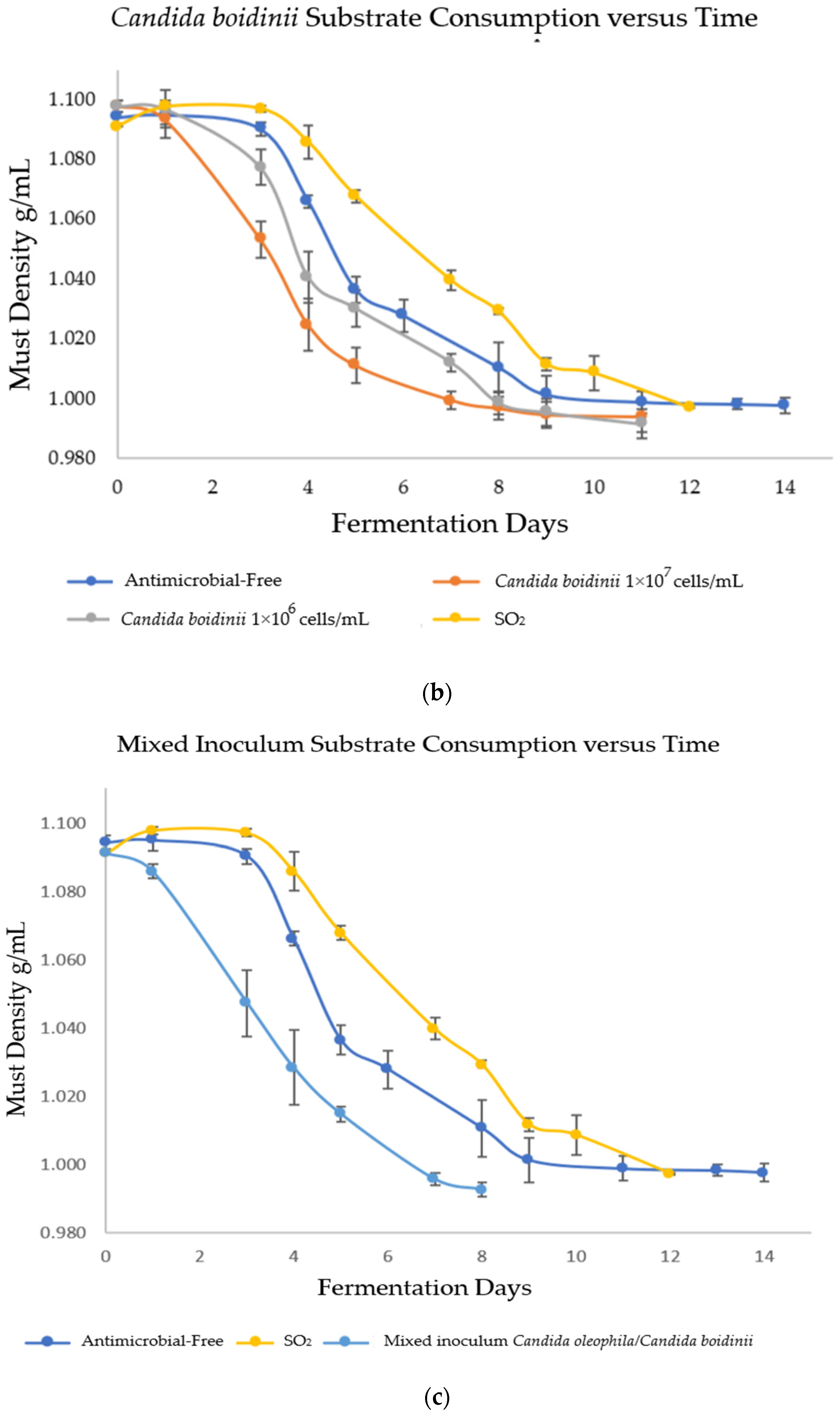
In order to evaluate the fermentation capacity of Candida spp. yeasts as bio-protectors and verify that fermentation reached its end, the must density was monitored daily during alcoholic fermentation (AF). The results are shown in Figure 1, where strategies bio-protected with C. oleophila finished fermentation on days 12 and 14 (Figure 1a) for the inoculum of 1 × 107 and 1 × 106 cells/mL, respectively; C. boidinii finished fermentation on day 11 for both inoculums (Figure 1b); and the mixed strategy of C. oleophila/C. boidinii 50:50 finished fermentation on day 8 (Figure 1c). In contrast, initially, spontaneous fermentations were performed on days 12 and 14 for the strategy with sulfites and antimicrobial-free, respectively.
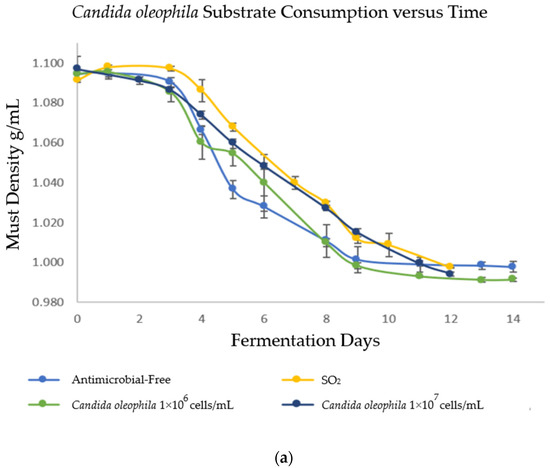
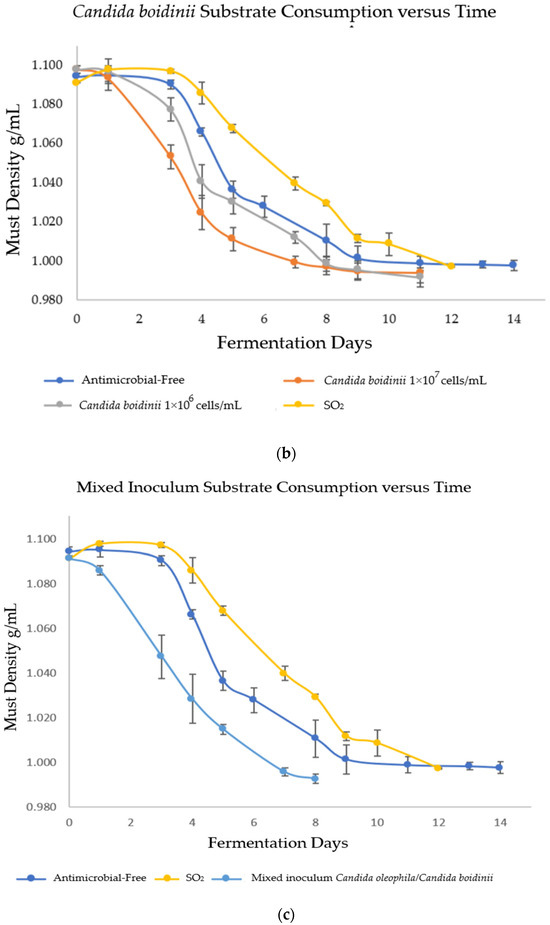
Figure 1.
Evolution of substrate consumption via measurement of density as a function of time. (a) Bio-protection with Candida oleophila; (b) bio-protection with Candida boidinii; and (c) bio-protection with C. oleophila/C. boidinii 50:50 (CO/CB). Error bars indicate standard deviation from triplicates.
3.2. Population Dynamics of Deterioration Microorganisms
Microorganism population analysis was conducted to evaluate spoilage microorganism behavior in the face of different bio-protective modalities over time. The first 72 h after bio-protector inoculation are highly relevant for analysis since they help to analyze the behavior of the spoilage microorganisms against the antimicrobial agent under study with minimal influence of external factors that can induce cell death, such as the inoculation of fermentative S. cerevisiae yeast or a rise in ethanol concentrations during fermentation. A bio-protective strategy will exhibit antimicrobial potential when it shows a lower concentration of spoilage microorganisms, whether LAB, AAB, or BB, compared to the control with SO2, and simultaneously demonstrates significant differences from the sulfite strategy via the post hoc Tukey test (p ≤ 0.05).
3.2.1. Evolution of Acetic Acid Bacteria during Fermentation
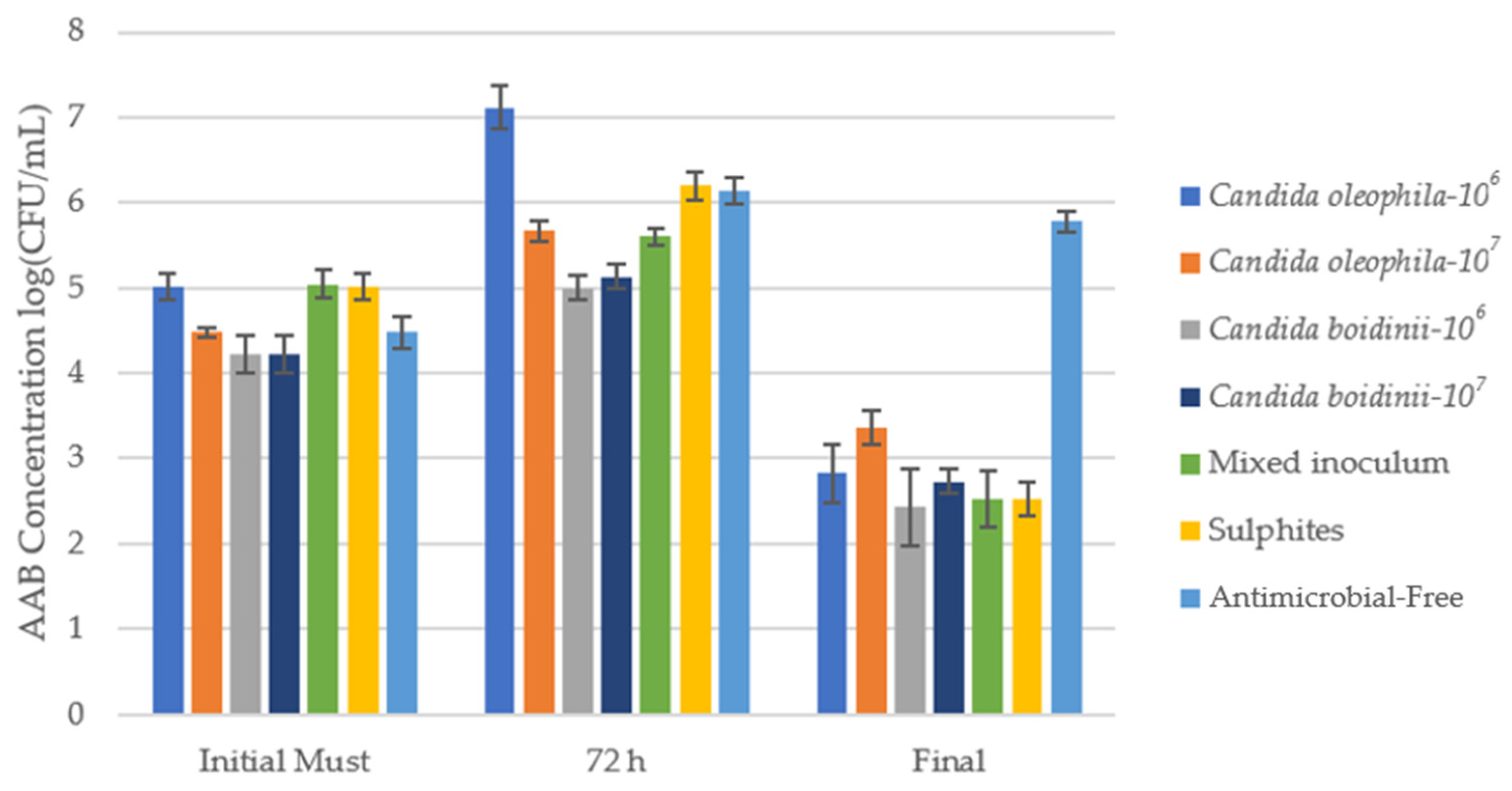
Figure 2 shows the counts of acetic acid bacteria AAB (log10(CFU/mL)) in a WLD Agar medium for an initial must after 72 h and at the end of alcoholic fermentation, and Table 1 shows the counts of AAB in CFU/mL units with the post hoc Tukey test to assess significant differences between the different modalities for each day.
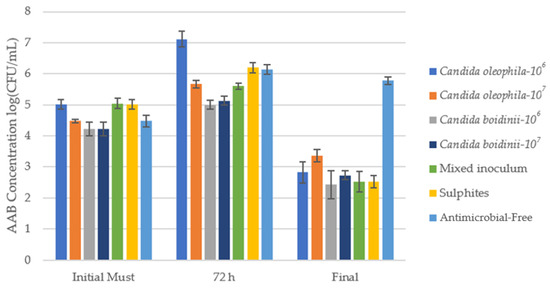
Figure 2.
Cell growth media of acetic acid bacteria (AAB) log10(CFU/mL) identified via culture on a plate with WLD agar medium for the bio-protected strategies, control with sulfites (SO2), and antimicrobial-free strategies at 3 times: initial must, day 3 (72 h), and at the end of fermentation. Error bars indicate standard deviation from triplicates.

Table 1.
Numeration of the different populations during the winemaking process for AAB at different moments across the fermentation: Initial Must, Day 1, Day 3, and at the end of the fermentation with post hoc Tukey test analysis (significance level, p ≤ 0.05), where different letters indicate significant differences between various strategies. * The treatments’ antimicrobial effectiveness ranking was as follows, according to control capacity in the first 72 h: Candida oleophila-106 = Candida boidinii-107 > Candida boidinii/Candida oleophila-107 > Candida oleophila-107 > Antimicrobial-free > SO2 > Candida oleophila-106.
In the initial must, the bacterial population stood between 1.67 × 104 CFU/mL and 1.10 × 105 CFU/mL, a value that rose during the first 72 h. At this point, fermentation with the bio-protector Candida boidinii achieved the lowest concentrations of AAB, with values of 1.00 × 105 and 1.37 × 105 CFU/mL for the inoculum of 1 × 106 and 1 × 107 cells/mL, respectively, without significant differences between both modalities (Table 1). On the other hand, the bio-protected strategy with the mixed inoculum C. oleophila/C. boidinii maintained an ABB concentration of 4.00 × 105 CFU/mL, remaining below the sulfite control. Overall, by day 3, bio-protected strategies with Candida boidinii controlled AAB populations more effectively than the control, exhibiting significantly lower concentrations than the traditional sulfite strategy (Table 1).
Over the course of fermentation, a decline in AAB concentrations in all strategies apart from the antimicrobial-free mode, which at the end of fermentation presented a higher spoilage microorganism concentration than the initial must (Figure 2). The bio-protected strategy that showed the best results was Candida boidinii, which managed to reduce AAB populations by at least 62 times compared to the initial must. Additionally, in all samples, the Candida boidinii 106 strategy reached concentrations equal to or greater than the Candida boidinii 107 strategy, so no dependency on the initial inoculum concentration was observed in the case of AAB.
3.2.2. Evolution of Lactic Acid Bacteria (LAB) during Fermentation
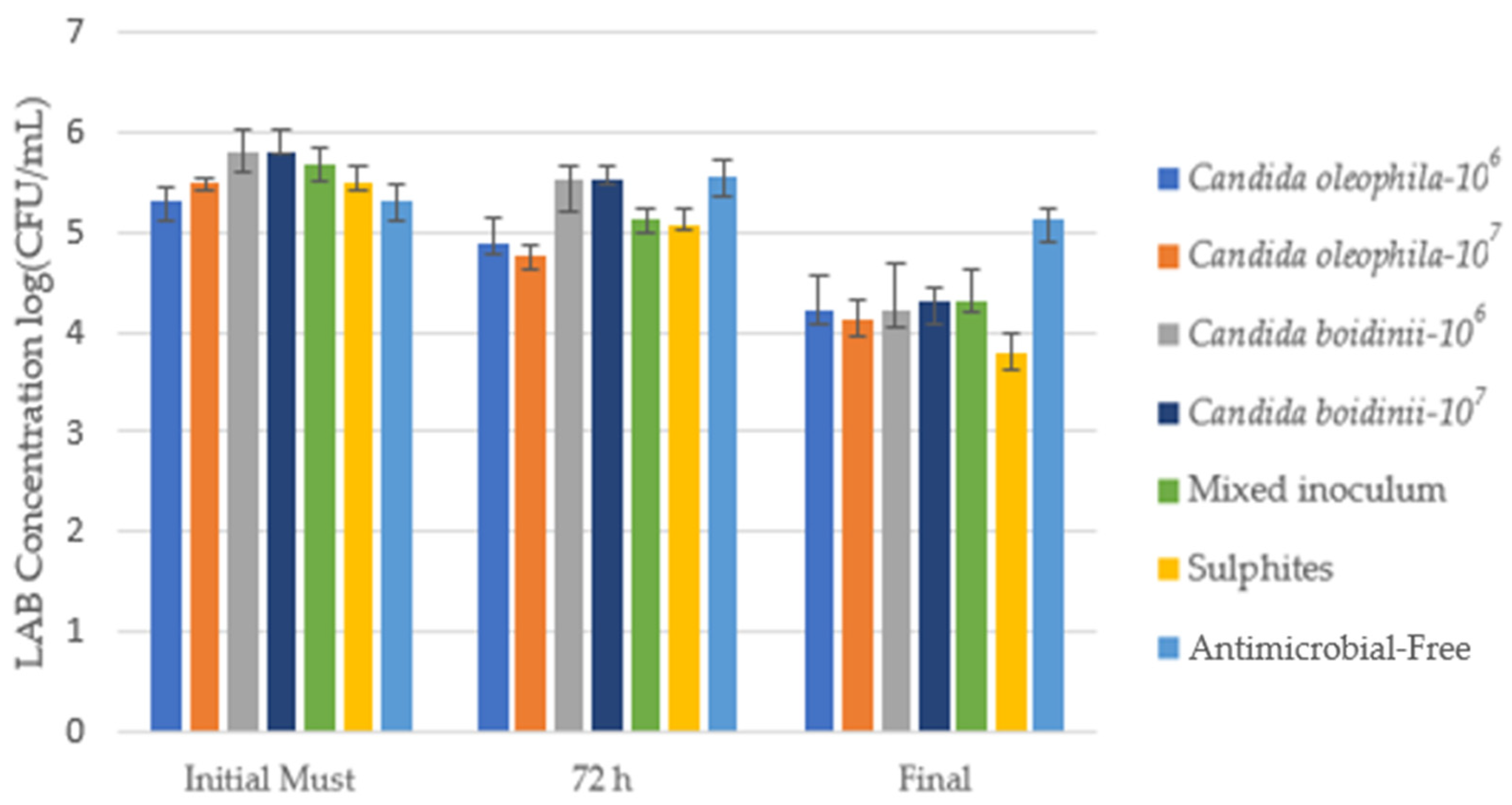
Figure 3 shows the counts of acetic acid bacteria AAB (log10(CFU/mL)) in an MRS Agar medium for the initial must, after 72 h, and at the end of alcoholic fermentation. Table 2 shows the counts of LAB in CFU/mL units with the post hoc Tukey test to assess significant differences between the different modalities for each day.
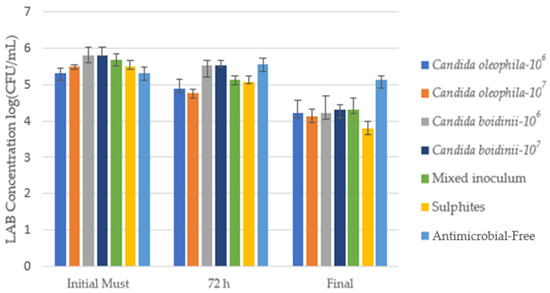
Figure 3.
Cell growth media of lactic acid bacteria (LAB) log10(CFU/mL) identified via culture on a plate with MRS agar medium for the bio-protected strategies, control with sulfites (SO2), and antimicrobial-free strategies at 3 times: Initial must, day 3 (72 h), and at the end of fermentation. Error bars indicate standard deviation from triplicates.
Table 2.
Numeration of the different populations during the winemaking process for LAB in different moments across the fermentation: Initial Must, Day 1, Day 3, and at the end of the fermentation with post hoc Tukey test analysis (significance level, p ≤ 0.05), where different letters indicate significant differences between various strategies. * The treatments’ antimicrobial effectiveness ranking was as follows, according to control capacity in the first 72 h: Candida oleophila-107 > Candida oleophila-106 > SO2 > Candida boidinii/Candida oleophila-107 > Candida boidinii-106 > Candida boidinii-107 > Antimicrobial-free.
In the initial must, the bacterial population was between 2.00 × 105 CFU/mL and 6.33 × 105 CFU/mL, a concentration which fell during the first 24 h in all strategies, apart from antimicrobial-free, whose concentration rose and continued growing until fermentation was well advanced. The Candida oleophila 107 strategy achieved a lower concentration than the sulfites for the first 24 h, while Candida oleophila 106 behaved identically to the sulfites control modality, reaching concentrations of 2.00 × 105 CFU/mL at 24 h (Table 2).
At 72 h, the Candida boidinii yeast reached the highest lactic acid bacteria (LAB) concentrations, second only to the antimicrobial-free modality (Figure 3). Candida oleophila presented the lowest LAB concentrations, reaching concentrations of 7.67 × 104 CFU/mL and 5.67 × 104 CFU/mL for Candida oleophila 106 and Candida oleophila 107, respectively (Table 2), remaining below the sulfite control fermentation. With the mixed Candida oleophila/Candida boidinii 50:50 inoculum, we observed a spoilage microorganism concentration above that obtained with Candida oleophila but below that obtained with Candida boidinii, reaching a concentration of 1.33 × 105 CFU/m. Additionally, this strategy exhibited behavior similar to that of sulfites, without significant differences (Table 2).
During fermentation, a considerable LAB concentration drop was observed in all strategies, except for the antimicrobial-free modality, which on the final day presented a concentration slightly below that obtained in the initial must (Table 2). The sulfite strategy was able to cut LAB concentration to 6.00 × 103 CFU/mL, being the strategy that showed the lowest final concentrations of this spoilage microorganism along with the bio-protected strategy with Candida oleophila 107, without observing significant differences between both modalities (Table 2). At the end of fermentation, the Candida oleophila 107 strategy managed to reduce LAB populations by at least 23 times compared to the initial must.
3.2.3. Evolution of Brettanomyces bruxellensis Yeasts during Fermentation
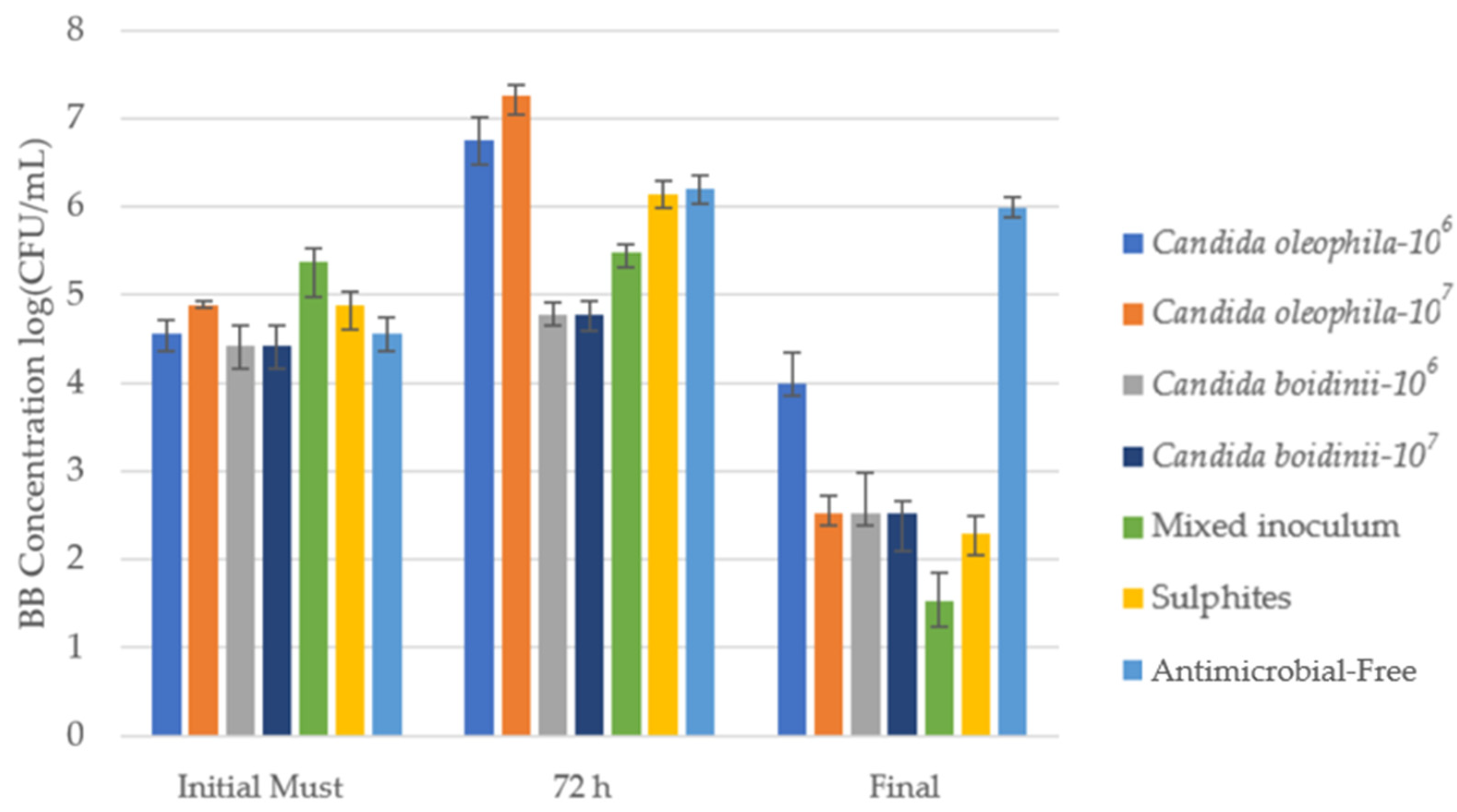
Figure 4 shows the counts of B. bruxellensis BB (log10(CFU/mL)) in specific Agar media for the initial must, after 72 h, and at the end of alcoholic fermentation. Table 3 shows the counts of BB in CFU/mL units with the post hoc Tukey test to assess significant differences between the different modalities for each day.
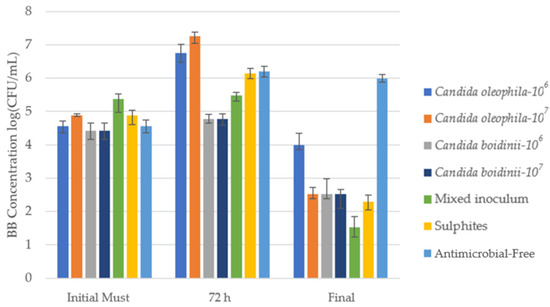
Figure 4.
Cell growth media of Brettanomyces bruxellensis (BB) log10 (CFU/mL) identified via culture on a plate with specific agar medium for the bio-protected strategies, control with sulfites (SO2) and antimicrobial-free strategies at 3 times: initial must, day 3 (72 h), and at the end of fermentation. Error bars indicate standard deviation from triplicates.
Table 3.
Numeration of the different populations during the winemaking process for Brettanomyces bruxellensis in different moments across the fermentation: Initial Must, Day 1, Day 3, and at the end of the fermentation with post hoc Tukey test analysis (significance level, p ≤ 0.05), where different letters indicate significant differences between various strategies. * The treatments’ antimicrobial effectiveness ranking was as follows, according to control capacity in the first 72 h Candida boidinii-107 > Candida boidinii/Candida oleophila-107 > Candida boidinii-106 = SO2 > Antimicrobial-free > Candida oleophila-106 > Candida oleophila-107.
In the initial must, the bacterial population stood between 2.67 × 104 CFU/mL and 2.33 × 105 CFU/mL, a concentration that rose during the first 24 h in most strategies except the fermentation with the mixed C. oleophila/C. boidinii 50:50 inoculum, where the concentration fell compared with the initial must (Table 3).
At 72 h, we can observe how in most studied fermentations, the BB concentration continued rising (Table 3), while the Candida boidinii 107 concentration remained almost constant between 24 h and 72 h, with 6.00 × 104 CFU/mL, presenting a lower concentration than the sulfites control and the remaining strategies.
During fermentation, a considerable drop in B. bruxellensis concentrations occurred in all strategies except in the antimicrobial-free strategy, which at the end of fermentation presented a higher concentration than the initial must (9.67 × 105 CFU/mL), a phenomenon observed with all spoilage microorganisms (Table 1, Table 2 and Table 3). The mixed strategy offered the greatest antimicrobial protection at the end of fermentation, reducing BB populations by at least 7000 times compared to the initial must, while the C. boidinii strategies did not present significant differences with the sulfite control (Table 3).
3.3. Chemical Compound Analysis via Biosystem Y15 and HPLC
A chemical composition analysis was performed (Table 4) to estimate the concentration of some compounds of interest in order to verify that the final wine quality was not negatively affected using Candida spp. bio-protectors to replace sulfites. The analyses were only performed for fermentations with an initial inoculum of 1 × 107 cells/mL since in all fermentations, the spoilage microorganisms were controlled as well as or better than fermentations with the inoculum of 1 × 106 cells/mL.
Table 4.
Chemical analysis of the must for each modality under study with their respective standard deviations and ANOVA–Tukey tests (significance level 95%, p ≤ 0.05), where different letters indicate significant differences between various strategies and p value ≤ 0.05 indicates significant differences in at least one of the strategies.
The strategy with Candida oleophila was characterized by a greater concentration of acetic acid (0.27 ± 0.03 g/L), significantly different from the other protected strategies, and close to the antimicrobial-free fermentation. Candida Boidinii was the only strategy that did not present significant differences from the fermentation with sulfites in acetic acid production. However, it was characterized by a greater ethanol concentration (12.78 ± 0.15 g/L), significantly different from the other strategies. In turn, the mixed CO/CB strategy presented the lowest ethanol levels (11.70 ± 0.15 g/L), unlike the sulfite control and the other fermentations. With regard to the production of the metabolite tartaric acid, no bio-protected strategy presented significant differences with the control except for the antimicrobial-free strategy, which was characterized by low concentrations of this compound (1.33 ± 0.01 g/L); however, it presented ethanol concentrations like the sulfite strategy.
No significant differences were obtained between the bio-protected strategies and sulfites in the production of L-malic acid, glycerol, L-lactic acid, and residual sugar.
3.4. Volatile Compound Analysis via GC-MS-SPME
At the end of alcoholic fermentation, volatile compounds were analyzed (Table 5) using GC-MS-SPME for each implemented strategy. In total, we obtained 27 volatile compounds and only 7 volatile compounds of greater abundance: Phenylethyl Alcohol; Hexanoic acid, ethyl ester; Octanoic acid, ethyl ester; Decanoic acid, ethyl ester; Ethyl 9-decenoate; Dodecanoic acid, ethyl ester; and Acetic acid, hexyl ester. These compounds exceeded 3% abundance in at least one of the strategies.
Table 5.
The concentration of volatile compounds (% of abundance) in bio-protection strategies with C. oleophila (CO), C. boidinii (CB), mixed inoculum C. oleophila/C. boidinii (CO/CB), antimicrobial-free, and SO2. Data are average values of two biological replicates ± standard deviation, while the letters a, b, c, d, and e represent significantly different statistical groups (post hoc Tukey, p < 0.05).
The bio-protection strategy with C. oleophila led to high concentrations of phenylethyl alcohol (21.20 ± 2.33%) and the lowest concentrations of esters, except for the hexanoic acid–ethyl ester compound, whose concentration presented no significant differences between modalities, and where the C. oleophila strategy presented an abundance of 23.60 ± 4.59%. In turn, the strategies with Candida Boidinii and the mixed inoculum C. boidinii/C. oleophila were characterized by a high production of the compound ethyl-9-decenoate (13.61 ± 0.62% and 6.14 ± 0.15%, respectively), by contrast with the sulfites control (2.72 ± 0.36%) and the strategy antimicrobial-free (1.57 ± 0.22%), characterized by low concentrations of this compound. We also observed higher concentrations of dodecanoic acid–ethyl ester for the C. boidinii application (3.90 ± 0.38%) and mixed strategy (4.71 ± 0.34%), compared to the control with sulfites (2.42 ± 0.31%). Another notable aspect is the high production of the octanoic acid–ethyl ester compound. All strategies apart from C. oleophila present values above 30% abundance for this compound, which is highly relevant for the volatile composition of the final wine product obtained. On the other hand, the wines with C. boidinii in simple or mixed inoculum contained higher concentrations of acids (hexanoic acid and octanoic acid). Although significant, these differences were not relevant in terms of enology or contribution to organoleptic components (<5% of the total amount).
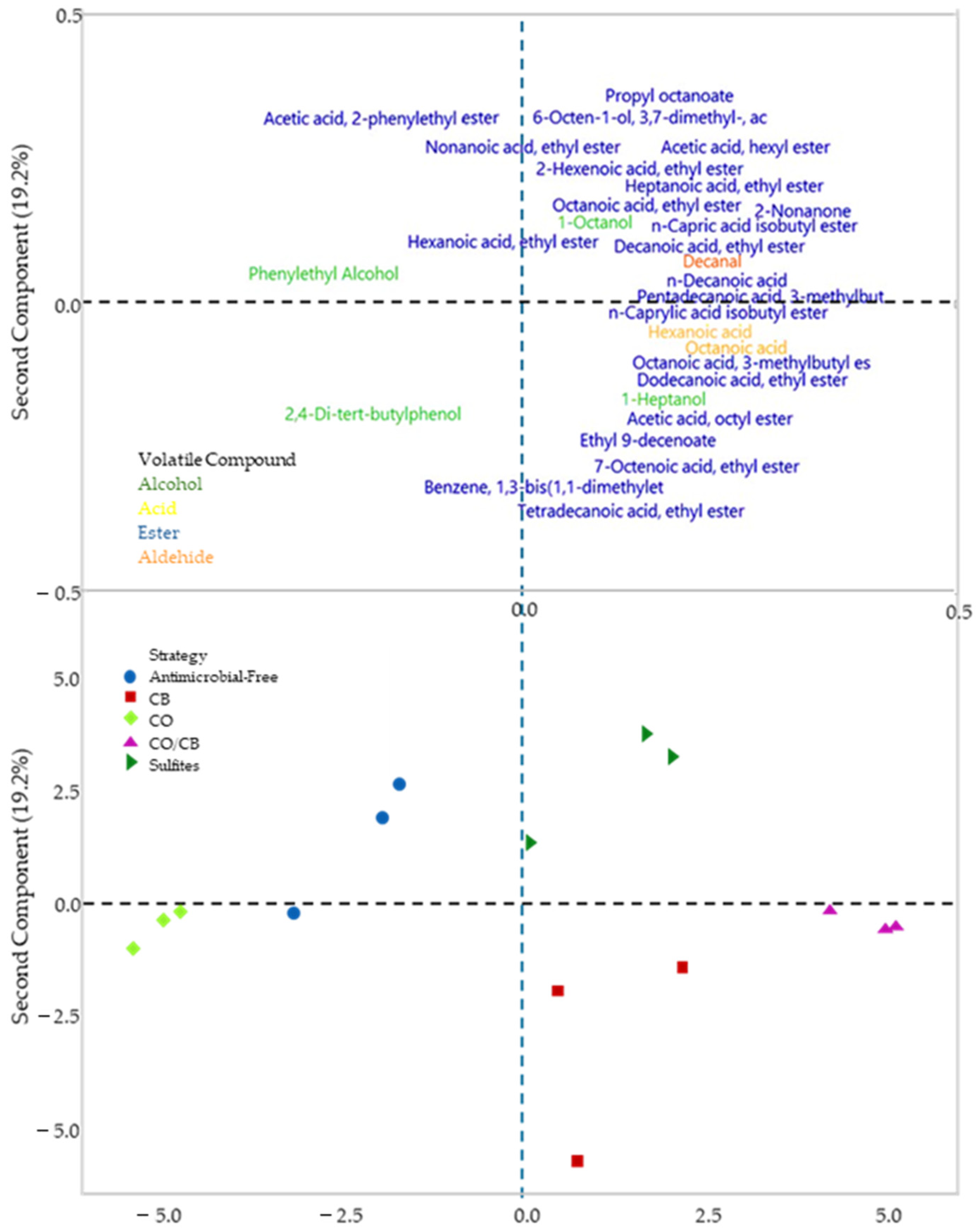
A principal component analysis (PCA) was generated in the form of biplots (Figure 5), where the first principal component represents 43.6% of the variance and separates the bio-protected strategy with C. oleophila and antimicrobial-free strategy (left) from the other strategies (right), while the second principal component represents 19.2% of the variance and separates the control strategies (SO2 and antimicrobial-free) from the bio-protectives strategies. The C. oleophila strategy was correlated to phenylethyl alcohol and 2,4-di-tert-buthylphenol content, the sulfite strategy with acetic acid 2-phenylethyl ester, and the C. boidinii and mixed strategies with acids content, like hexanoic acid and octanoic acid; in addition, it correlated positively with most of the esters, the majority group within the analysis.
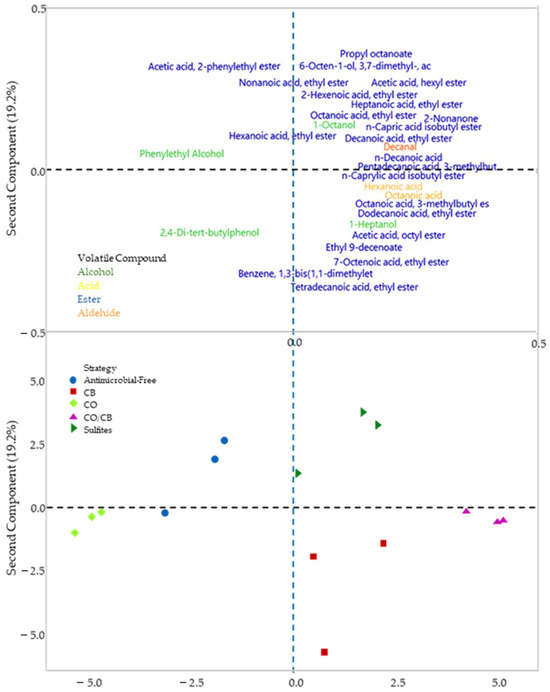
Figure 5.
Double projection graph for different bio-protection strategies in monoculture fermentation using different native yeasts compared to the control with SO2 and fermentation without antimicrobials. CO (Candida oleophila), CB (Candida boidinii), CO/CB (Candida oleophila and Candida boidinii 50/50), Antimicrobial-Free, and sulfites. The PCA analysis was conducted with Minitab Statistical software 2021.
4. Discussion
Non-Saccharomyces yeasts, particularly Candida oleophila and Candida boidinii, have gained little attention in enological research, unlike other commercial strains including Torulaspora delbrueckii, Metschnikowia pulcherrima, and Lachancea Thermotolerans [16,17,18,19,20,21,22,23,24]. Actually, only one study reported on the fermentative potential of C. oleophila and C. boidinii and their capacity to reduce ethanol during Sauvignon Blanc wine production [33]. However, its antimicrobial qualities have not yet been studied in wine fermentation, despite the potential demonstrated in other food production areas. C. oleophila has strong antifungal action, particularly for Penicillium spp. in post-harvest studies [28,29,30,31], while C. boidinii has been studied for olive production processes, with proven effects against the spoilage microorganisms Enterobacteriácea, Coliform, and Shigella [32].
In this context, we chose to select the non-saccharomyces yeasts Candida oleophila and Candida boidinii to carry out this study, focusing on the microbiological protection of the must compared with the traditional antimicrobial additive SO2. The final goal is to provide results showing that the C. oleophila and C. boidinii yeasts present bio-controlling potential against spoilage microorganisms which can help totally or partially replace sulfites, without negatively affecting wine quality.
It is important to mention that despite sharing the same genus name, the yeasts C. oleophila and C. boidinii do not constitute what would be considered a closely related species pair because they do not share the same taxonomic classification. It is likely that C. oleophila will be transferred to the genus Kurtzmaniella, while C. boidinii is currently classified as a member of the Ogataea/Candida clade.
4.1. Candida spp. Yeasts Produce Antimicrobial Effects in Grape Must
The results indicated that the bio-protective capacity of a yeast against a spoilage microorganism depends on the yeast species used, the inoculum concentration, and the spoilage microorganism under study.
In the case of lactic acid bacteria, C. oleophila yeast (Table 2) showed bio-protective behavior greater than that generated by C. boidinii and by CO/CB across fermentation, and a better antimicrobial capacity than sulfites during pre-fermentative stages (<72 h), followed by protection comparable with the control until the end of fermentation. These results align with Escribano-Viana [16], where a mixed inoculation of T. delbrueckii/L. thermotolerans on LAB production in red wine must had a slightly better controlling effect than sulfites, followed by equaling the effect of SO2 at the end of fermentation. In turn, bio-protection rose after increasing the initial inoculum concentration from 106 cells/mL to 107 cells/mL, showing that lactic acid bacteria were negatively affected by greater initial concentrations of bio-protective Candida oleophila.
Currently, no studies indicate a reduction of lactic acid bacteria when using Candida spp. strains, although Candida oleophila has been associated with controlling post-harvest rot diseases in fruits, mainly related to fungal control [28,29,30,31]. In fact, yeasts including M. Pulcherrima and L. thermotolerans; two strains that are highly studied and proven bio-controllers against LAB [16,18,19,20], are also used as antimicrobial agents for post-harvest diseases [38], meaning that there could be a relation between both potentials. However, further study is needed to support this.
For acetic acid bacteria, Candida boidinii yeast (Table 1) presented greater antimicrobial behavior than that of Candida oleophila across fermentation, as well as being better than the strategy with sulfites and CO/CB in pre-fermentative stages (<72 h). At the end of fermentation, AAB concentrations were similar and without significant differences from sulfites and CO/CB strategies, probably due to the anaerobic conditions arising after inoculation with S. cerevisiae on the third day [16], since AAB only developed in the presence of oxygen [21]. In turn, both Candida boidinii 1 × 106 and Candida boidinii 1 × 107 strategies presented concentrations without significant differences, meaning that no direct relation was observed between the concentration of the initial inoculum and the bio-protective effect against AAB with Candida boidinii. However, when increasing C. oleophila concentrations from 1 × 106 to 1 × 107 cells/mL, there was a strong reduction in AAB concentration, from 6.33 × 104 to 3.33 × 104 CFU/mL in the sample at 72 h, with an observable dependence on the initial inoculum concentration. It would be interesting to use larger C. oleophila concentrations in future studies, since although there was a strong drop in AAB concentration at the end of the fermentation, the results did not exceed the protective effect generated by either C. boidinii or sulfites.
In turn, Brettanomyces bruxellensis could be effectively controlled with the Candida boidinii 1 × 107 and mixed inoculum strategies (Table 3 and Figure 4), showing greater efficacy than Candida oleophila until the end of fermentation. When compared with the sulfites control, the Candida boidinii 1 × 107 strategy showed better antimicrobial behavior than sulfites in pre-fermentative stages, while the mixed CO/CB strategy showed such behavior until the end of fermentation, despite initially presenting higher Brettanomyces bruxellensis concentrations. In this case, C. boidinii presented a bio-protective behavior dependent on the initial inoculum concentration, where higher concentrations correlated with greater antimicrobial effects against B. bruxellensis.
The cause of the controlling effect presented by C. boidinii may be the rapid fermentative power of this yeast, which can finish fermentation more quickly than the other modalities whether with simple inoculation or in mixed inoculation CO/CB (Figure 1b,c), leading to swiftly exhausting nutrients in the medium and producing ethanol. In fact, the Candida boidinii 1 × 107 strategy presented an ethanol production of 12.78 ± 0.15 % v/v (Table 4), a significantly higher concentration than that obtained by the sulfites control and the other strategies. This behavior was unexpected since the literature reports that non-saccharomyces yeasts in general, and Candida boidinii in particular, are yeasts that can reduce ethanol levels in wine [33,39]. However, this may be due to the aforementioned lack of control over LAB (Table 2), given that the presence of hetero-fermentative LAB can increase CO2 and ethanol concentrations in wine [40]. The rapid fermentation caused by C. boidinii yeast compared with C. oleophila had already been reported in the literature [33], where sequential fermentation with the yeasts C. boidinii–S. cerevisiae finished fermentation by day 8, while the C. oleophila–S. cerevisiae duo finished by day 15. The importance of this lies in the fact that ethanol is considered an active bio-controlling compound [15] since most microorganisms are inhibited by high concentrations of this compound. Strategies that present a rapid fall in density (C. boidinii 1 × 106, C. boidinii 1 × 107, and mixed inoculum) (Figure 1b,c), produce ethanol earlier, altering microorganisms susceptible to this metabolite. The ones affected in this case are AAB and B. bruxellensis yeast.
In summary, the strategies with Candida oleophila were more effective at suppressing LAB than Candida boidinii strategies, whereas the latter was more effective against AAB and B. bruxellensis than Candida oleophila strategies.
4.2. Final Wine Quality Is Comparable to That Obtained with SO2
Quality wines were obtained with the experimental conditions used, without significant differences between the bio-protected strategies and sulfite in producing L-malic acid, L-lactic acid, glycerol, tartaric acid, and residual sugar (Table 4). All strategies were characterized by low L-lactic acid concentrations and the same L-malic acid concentration, despite differences in the final population of lactic acid bacteria (Table 2 and Figure 3). Indeed, the antimicrobial-free strategy was expected to present a higher concentration of lactic acid, given the lack of control obtained over LAB (Table 2). However, the reason for this could be explained by the inhibition of malolactic fermentation (MLF), a process responsible for transforming malic acid into lactic acid due to the presence of certain compounds such as medium-chain fatty acids, organic acids, and peptides [41], which could have been present in the final wines, preventing the production of lactic acid. On the other hand, the desired temperature for MLF to occur should be around 25 °C; in fact, it is known that temperatures between 12 and 20 °C, such as those used in this research, inhibit the metabolic activity of LAB, preventing the production of their metabolites [42]. Additionally, another inhibitory compound is alcohol, where concentrations between 12 and 15% v/v alter the bacterial cell membrane, disrupting their metabolism; indeed, the strategies with C. boidinii and antimicrobial-free presented higher alcohol concentrations (Table 4), potentially causing the inactivation of lactic bacteria that prevented differences in the final chemical profile [42]. Lastly, the type of strain must be considered, given that Oenococcus oeni is the main bacterial species responsible for carrying out this biochemical stage, due to its ability to tolerate the harsh physicochemical properties of the wine once fermentation is complete [43]; therefore, the presence of other LAB species may have prevented malic acid from transforming into lactic acid. However, to test this hypothesis, metabolomic-level analyses such as quantitative PCR would be required.
All strategies also presented a low final concentration of residual sugars (glucose and fructose), meaning that the final wines were dry and fermentative yeast action was unaffected by bio-protectors. The bio-protection modality with Candida oleophila showed significantly high acetic acid concentrations (0.267 g/L) compared with the sulfite strategy. These findings align with the microbiological analysis, where C. oleophila did not achieve effective bio-control against AAB (Table 1 and Figure 2), the main producers of this metabolite. However, the maximum allowable level in wines was not exceeded (1.2 g/L) [44], preventing negative quality impacts.
With regard to volatile composition, we obtained wines with desirable organoleptic properties mainly associated with floral and fruity character. Candida oleophila presented high concentrations of phenethyl alcohol (2-phenylethanol) associated with rose scents, significantly differing from the other strategies that stood out due to the production of ethyl esters from linear fatty acids (Table 5). This may be due to high concentrations of Brettanomyces bruxellensis, a spoilage microorganisms present in musts protected with C. oleophila (Figure 4) and which can produce phenylethyl alcohol through the Ehrlich pathway [45]. However, studies have only been conducted on the fermentation of carrot pomace, and not on wine. There is current research about the potential for Candida albicans to produce phenylethyl alcohol [46], so Candida oleophila may also produce this compound; however, there are no studies in this regard. Another alternative could be related to lipids released by the early cellular death of Candida oleophila (Table A1), which repress the acetyl transferase enzymes (ATF), decreasing esters’ synthesis [47].
Another relevant aspect is the volatile profile obtained using the Candida boidinii strategy (Table 5), which produced high concentrations of ethyl-9-decenoate (13.61% abundance), associated with fruity peach aromas and significantly differing from the other strategies. This may be due to the presence of LAB which are not effectively controlled by the bio-protector Candida boidinii. Wang et al. [48], in a study on the effect of initiating cultures on aromatic compounds and wine microbiota, showed that wines with a greater population of Leuconostoc and Lactobacillus bacteria were positively correlated with ethyl 9-decenoate. However, no direct relation has yet been established between lactic bacteria and the production pathways of this compound. The study was also conducted on Cabernet Sauvignon rather than Sauvignon Blanc, although it could aid future studies.
The initially spontaneous strategies (sulfites and antimicrobial-free) and mixed strategy were characterized by the presence of fatty acid esters, including ethyl hexanoate, ethyl octanoate, and ethyl decanoate. These results align with the literature reports on Sauvignon Blanc wines fermented with Saccharomyces Cerevisiae [49]. In fact, a study evaluating the effect of SO2 on Sauvignon Blanc must showed that the ethyl esters ethyl hexanoate, ethyl octanoate, and ethyl decanoate were produced in greater quantities in wines after adding sulfites compared to those without sulfites [49].
In general, the volatile profile was not negatively affected by bio-protectors. In fact, they added aromatic complexity to the wine, particularly Candida boidinii and Candida oleophila, which showed positive differences from the control. In future studies, it would be interesting to complement the results with genetic identification techniques, including a quantitative PCR and MALDI-TOF MS, which would help identify microorganism species and verify the proper implantation of the bio-protected strain.
5. Conclusions
This is the first study reporting the use of non-saccharomyces yeasts Candida oleophila and Candida boidinii as bio-protectors to replace sulfites in winemaking at a laboratory scale. The results obtained indicate that the yeasts studied can control microflora that deteriorate Sauvignon Blanc must with effectiveness depending on the initial concentration used and the target spoilage microorganisms. C. oleophila had a better antimicrobial effect than SO2 against LAB in pre-fermentative states, while C. boidinii was effective against AAB and Brettanomyces bruxellensis. The mixed inoculum of C. oleophila/C. boidinii could effectively control Brettanomyces bruxellensis. The initial inoculation of 1 × 107 cells/mL presented greater antimicrobial protection than those with a concentration of 1 × 106 cells/mL against LAB and B. bruxellensis. This study showed that these yeasts can exert an antimicrobial activity similar to SO2, without negatively altering the organoleptic properties of the wine.
Author Contributions
Conceptualization C.V., A.D. and A.U.; methodology, C.V., A.D. and A.U.; validation, C.V. and C.M.-A.; formal analysis, C.V., C.M.-A., A.D. and A.U.; investigation, C.V.; resources, A.U.; data curation, C.V. and A.U.; writing—original draft preparation, C.V.; writing—review and editing, C.V., C.M.-A., A.D. and A.U.; visualization, C.V., C.M.-A. and A.U.; supervision, C.M.-A. and A.U.; project administration, C.V., C.M.-A. and A.U.; funding acquisition, C.V. and A.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a Maintenance scholarship from Universidad Técnica Federico Santa María.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data and intellectual property belong to Universidad Técnica Federico Santa María.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Table A1.
Percentages of abundance (%) for non-Saccharomyces (NS) and Saccharomyces (S) yeasts compared with the total of yeasts for all strategies evaluated during experiments and at different times across fermentation (initial must, day 1, day 3, and final day).
Table A1.
Percentages of abundance (%) for non-Saccharomyces (NS) and Saccharomyces (S) yeasts compared with the total of yeasts for all strategies evaluated during experiments and at different times across fermentation (initial must, day 1, day 3, and final day).
| Strategy | C. oleophila 106 | C. oleophila 107 | C. boidinii 106 | C. boidinii 107 | Mixed Inoculum | SO2 | Antimicrobial-Free | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Day | NS | S | NS | S | NS | S | NS | S | NS | S | NS | S | NS | S |
| Must | 90.14 | 9.86 | 88.00 | 12.00 | 78.95 | 21.05 | 78.95 | 21.05 | 92.31 | 7.69 | 88.00 | 12.00 | 90.14 | 9.86 |
| 1 | 94.74 | 5.26 | 95.24 | 4.76 | 84.21 | 15.79 | 89.66 | 10.34 | 97.78 | 2.22 | 75.86 | 24.14 | 95.83 | 4.17 |
| 3 | 77.14 | 22.86 | 78.69 | 21.31 | 60.61 | 39.39 | 70.00 | 30.00 | 87.50 | 12.50 | 16.67 | 83.33 | 19.23 | 80.77 |
| Final | 6.67 | 93.33 | 7.89 | 92.11 | 9.40 | 90.60 | 4.40 | 95.60 | 11.25 | 88.75 | 3.00 | 97.00 | 12.50 | 87.50 |
References
- Fia, G.; Menghini, S.; Mari, E.; Proserpio, C.; Pagliarini, E.; Granchi, L. Replacement of SO2 with an Unripe Grape Extract and Chitosan during Oak Aging: Case Study of a Sangiovese Wine. Antioxidants 2023, 12, 365. [Google Scholar] [CrossRef]
- Oliveira, C.M.; Ferreira, A.C.; De Freitas, V.; Silva, A.M. Oxidation mechanisms occurring in wines. Food Res. Int. 2011, 44, 1115–1126. [Google Scholar] [CrossRef]
- Jagatić, A.M.; Biloš, J.; Kozina, B.; Tomaz, I.; Preiner, D.; Jeromel, A. Effect of Different Reducing Agents on Aromatic Compounds, Antioxidant and Chromatic Properties of Sauvignon Blanc Wine. Foods 2020, 9, 996. [Google Scholar] [CrossRef]
- Vally, H.; Misso, N.L.; Madan, V. Clinical effects of sulfite additives. Clin. Exp. Allergy 2009, 39, 1643–1651. [Google Scholar] [CrossRef]
- JECFA. Safety Evaluation of Certain Food Additives and Contaminants; World Health Organization: Geneva, Switzerland, 2006.
- International Organisation of Vine and Wine (OIV). Molecular sulfur dioxide. In Compendium of International Methods of Must and Wine Analysis; OIV: Paris, France, 2019. [Google Scholar]
- Nieto-Villegas, R.; Bernabéu, R.; Rabadán, A. The impact of chemophobia on wine consumer preferences explored through the case of sulfites. J. Agric. Food Res. 2023, 14, 100692. [Google Scholar] [CrossRef]
- Ma, Y.; Yu, K.; Wang, N.; Xiao, X.; Leng, Y.; Fan, J.; Du, Y.; Wang, S. Sulfur dioxide-free wine with polyphenols promotes lipid metabolism via the Nrf2 pathway and gut microbiota modulation. Food Chem. X 2024, 21, 101079. [Google Scholar] [CrossRef]
- Delso, C.; Berzosa, A.; Sanz, J.; Álvarez, I.; Raso, J. Pulsed electric field processing as an alternative to sulfites (SO2) for controlling saccharomyces cerevisiae involved in the fermentation of Chardonnay white wine. Food Res. Int. 2023, 165, 112525. [Google Scholar] [CrossRef]
- Nogueira, D.P.; Jiménez-Moreno, N.; Esparza, I.; Moler, J.A.; Ferreira-Santos, P.; Sagües, A.; Teixeira, J.A.; Ancín-Azpilicueta, C. Evaluation of grape stems and grape stem extracts for sulfur dioxide replacement during grape wine production. Curr. Res. Food Sci. 2023, 6, 100453. [Google Scholar] [CrossRef]
- Hosseini, A.; Koushesh, M.; Watkins, C.B. Microbial antagonists to biologically control postharvest decay and preserve fruit quality. Crit. Rev. Food Sci. Nutr. 2023, 1–13. [Google Scholar] [CrossRef]
- Alexandre, H.; Puyo, M.; Tourdot-Maréchal, R. Bio-protection in Winemaking. In New Advances in Saccharomyces; Morata, A., Loira, I., González, C., Escott, C., Eds.; IntechOpen: London, UK, 2023. [Google Scholar]
- Leyva, M.; Mounier, J.; Valence, F.; Coton, M.; Thierry, A.; Coton, E. Antifungal microbial agents for food biopreservation—A review. Microorganisms 2017, 5, 37. [Google Scholar] [CrossRef]
- Bauer, M.A.; Kainz, K.; Carmona-Gutierrez, D.; Madeo, F. Microbial wars: Competition in ecological niches and within the microbiome. Microb. Cell 2018, 5, 215–219. [Google Scholar] [CrossRef] [PubMed]
- Di Gianvito, P.; Englezos, V.; Rantsiou, K.; Cocolin, L. Bio-protection strategies in winemaking. Int. J. Food Microbiol. 2022, 364, 109532. [Google Scholar] [CrossRef] [PubMed]
- Escribano-Viana, R.; González-Arenzana, L.; Garijo, P.; Fernández, L.; López, R.; Santamaría, P.; Gutiérrez, A.R. Bio-protective Effect of a Torulaspora delbrueckii/Lachancea thermotolerans-Mixed Inoculum in Red Winemaking. Fermentation 2022, 8, 337. [Google Scholar] [CrossRef]
- Simonin, S.; Alexandre, H.; Nikolantonaki, M.; Coelho, C.; Tourdot-Maréchal, R. Inoculation of Torulaspora delbrueckii as a bio-protection agent in winemaking. Food Res. Int. 2018, 107, 451–461. [Google Scholar] [CrossRef] [PubMed]
- Simonin, S.; Roullier-Gall, C.; Ballester, J.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Peyron, D.; Alexandre, H.; Tourdot-Maréchal, R. Bio-protection as an alternative to sulfites: Impact on chemical and microbial characteristics of red wines. Front. Microbiol. 2020, 11, 1308. [Google Scholar] [CrossRef] [PubMed]
- Windholtz, S.; Redon, P.; Lacampagne, S.; Farris, L.; Lytra, G.; Cameleyre, M.; Barbe, J.C.; Coulon, J.; Thibon, C.; Masneuf-Pomarede, I. Non-Saccharomyces yeasts as bio-protection in the composition of red wine and in the reduction of sulfur dioxide. LWT 2021, 149, 111781. [Google Scholar] [CrossRef]
- Windholtz, S.; Dutilh, L.; Lucas, M.; Maupeu, J.; Vallet-Courbin, A.; Farris, L.; Coulon, J.; Masneuf-Pomarède, I. Population dynamics and yeast diversity in early winemaking stages without sulfites revealed by three complementary approaches. Appl. Sci. 2021, 11, 2494. [Google Scholar] [CrossRef]
- Windholtz, S.; Nioi, C.; Coulon, J.; Masneuf-Pomarede, I. Bio-protection by non-Saccharomyces yeasts in oenology: Evaluation of O2 consumption and impact on acetic acid bacteria. Int. J. Food Microbiol. 2023, 405, 110338. [Google Scholar] [CrossRef]
- Lebleux, M.; Alexandre, H.; Romanet, R.; Ballester, J.; David-Vaizant, V.; Adrian, M.; Tourdot-Maréchal, R.; Rouiller-Gall, C. Must protection, sulfites versus bio-protection: A metabolomic study. Food Res. Int. 2023, 173, 113383. [Google Scholar] [CrossRef]
- Rubio-Bretón, P.; Gonzalo-Diago, A.; Iribarren, M.; Garde-Cerdán, T.; Pérez-Álvarez, E.P. Bio-protection as a tool to free additives winemaking: Effect on sensorial, anthocyanic and aromatic profile of young red wines. LWT 2018, 98, 458–464. [Google Scholar] [CrossRef]
- Puyo, M.; Simonin, S.; Bach, B.; Klein, G.; Alexandre, H.; Tourdot-Maréchal, R. Bio-protection in oenology by Metschnikowia pulcherrima: From field results to scientific inquiry. Front. Microbiol. 2023, 14, 1252973. [Google Scholar] [CrossRef]
- Kántor, A.; Hutková, J.; Petrová, J.; Hleba, L.; Kacániová, M. Antimicrobial activity of pulcherrimin pigment produced by Metschnikowia pulcherrima against various yeast species. J. Microbiol. Biotechnol. Food Sci. 2015, 5, 282. [Google Scholar] [CrossRef]
- Villalba, M.L.; Sáez, J.S.; Del Monaco, S.; Lopes, C.A.; Sangorrín, M.P. TdKT, a new killer toxin produced by Torulaspora delbrueckii effective against wine spoilage yeasts. Int. J. Food Microbiol. 2016, 217, 94–100. [Google Scholar] [CrossRef]
- Shekhawat, K.; Bauer, F.F.; Setati, M.E. Impact of oxygenation on the performance of three non-Saccharomyces yeasts in co-fermentation with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 2479–2491. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Zhang, R.; Xiong, B. Management of postharvest diseases of kiwifruit with a combination of the biocontrol yeast Candida oleophila and an oligogalacturonide. Biol. Control 2021, 156, 104549. [Google Scholar] [CrossRef]
- Ballet, N.; Souche, J.L.; Vandekerckove, P. Efficacy of Candida oleophila, strain O, in preventing postharvest diseases of fruits. In Proceedings of the III International Symposium on Postharvest Pathology: Using Science to Increase Food Availability, Bari, Italy, 7 November 2015. [Google Scholar]
- Droby, S.; Cohen, L.; Daus, A.; Weiss, B.; Horev, B.; Chalutz, E.; Katz, H.; Keren-Tzur, M.; Shachnai, A. Commercial testing of Aspire: A yeast preparation for the biological control of postharvest decay of citrus. Biol. Control 1998, 12, 97–101. [Google Scholar] [CrossRef]
- Sui, Y.; Wisniewski, M.; Droby, S.; Piombo, E.; Wu, X.; Yue, J. Genome sequence, assembly, and characterization of the antagonistic yeast Candida oleophila used as a biocontrol agent against post-harvest diseases. Front. Microbiol. 2020, 11, 295. [Google Scholar] [CrossRef]
- Bonatsou, S.; Panagou, E.Z. Fermentation of cv. kalamata natural black olives with potential multifunctional yeast starters. Foods 2022, 11, 3106. [Google Scholar] [CrossRef] [PubMed]
- Benavides, S.; Franco, W.; Ceppi De Lecco, C.; Durán, A.; Urtubia, A. Evaluation of Indigenous Candida oleophila and Candida boidinii in Monoculture and Sequential Fermentations: Impact on Ethanol Reduction and Chemical Profile in Chilean Sauvignon Blanc Wines. J. Fungi 2022, 8, 259. [Google Scholar] [CrossRef]
- Franco, W.; Benavides, S.; Valencia, P.; Ramírez, C.; Urtubia, A. Native Yeasts and Lactic Acid Bacteria Isolated from Spontaneous Fermentation of Seven Grape Cultivars from the Maule Region (Chile). Foods 2021, 10, 1737. [Google Scholar] [CrossRef]
- International Organization of Vine and Wine (OIV). Microbiological Analysis of Wines and Musts-Detection, Differentiation and Counting of Microorganisms; OIV: Paris, France, 2009. [Google Scholar]
- Perestrelo, R.; Barros, A.S.; Câmara, J.S.; Rocha, S.M. In-depth search focused on furans, lactones, volatile phenols, and acetals as potential age markers of Madeira wines by comprehensive two-dimensional gas chromatography with time-of-flight mass spectrometry combined with solid phase microextraction. J. Agric. Food Chem. 2011, 59, 3186–3204. [Google Scholar] [CrossRef] [PubMed]
- Gujar, A.; Anderson, T.; Cavagnino, D.; Patel, A. Comparative analysis of mass spectral matching for confident compound identification using the Advanced Electron Ionization source for GC-MS. Thermoscientific 2018, 10598, 1–7. [Google Scholar]
- Bosqueiro, A.; Bizarria, R.; Rosa-Magri, M.M. Biocontrol of post-harvest tomato rot caused by Alternaria arborescens using Torulaspora indica. Biocontrol Sci. Technol. 2023, 33, 115–132. [Google Scholar] [CrossRef]
- Xynas, B.; Barnes, C. Yeast or water: Producing wine with lower alcohol levels in a warming climate: A review. J. Sci. Food Agric. 2023, 103, 3249–3260. [Google Scholar] [CrossRef] [PubMed]
- König, H.; Fröhlich, J. Lactic acid bacteria. In Biology of Microorganisms on Grapes, in Must and in Wine; König, H., Unden, G., Fröhlich, J., Eds.; Springer: Cham, Germany, 2017; pp. 3–41. [Google Scholar]
- Balmaseda, A.; Bordons, A.; Reguant, C.; Bautista-Gallego, J. Non-Saccharomyces in wine: Effect upon Oenococcus oeni and malolactic fermentation. Front. Microbiol. 2018, 9, 352041. [Google Scholar] [CrossRef] [PubMed]
- Gil-Sánchez, I.; Suáldea, B.B.; Moreno-Arribas, M.V. Malolactic Fermentation. Red Wine Technology; Academic Press: Cambridge, MA, USA, 2019; pp. 85–98. [Google Scholar] [CrossRef]
- Bartowsky, E.J. Oenococcus oeni and malolactic fermentation–moving into the molecular arena. Aust. J. Grape Wine Res. 2005, 11, 174–187. [Google Scholar] [CrossRef]
- International Organization of Vine and Wine. Maximum acceptable Limits. In International Code of Oenological Practices; OIV: Paris, France, 2015. [Google Scholar]
- Lindsay, M.A.; Granucci, N.; Greenwood, D.R.; Villas-Boas, S.G. Fermentative Production of Volatile Metabolites Using Brettanomyces bruxellensis from Fruit and Vegetable By-Products. Fermentation 2022, 8, 457. [Google Scholar] [CrossRef]
- Ghosh, S.; Kebaara, B.W.; Atkin, A.L.; Nickerson, K.W. Regulation of aromatic alcohol production in Candida albicans. Appl. Environ. Microbiol. 2008, 74, 7211–7218. [Google Scholar] [CrossRef]
- Fujii, T.; Kobayashi, O.; Yoshimoto, H.; Furukawa, S.; Tamai, Y. Effect of aeration and unsaturated fatty acids on expression of the Saccharomyces cerevisiae alcohol acetyltransferase gene. Appl. Environ. Microbiol. 1997, 63, 910–915. [Google Scholar] [CrossRef]
- Wang, M.; Wang, J.; Chen, J.; Philipp, C.; Zhao, X.; Wang, J.; Liu, Y.; Suo, R. Effect of commercial yeast starter cultures on Cabernet Sauvignon wine aroma compounds and microbiota. Foods 2022, 11, 1725. [Google Scholar] [CrossRef]
- Coetzee, C.; Johannes du Toit, W. A comprehensive review on Sauvignon blanc aroma with a focus on certain positive volatile thiols. Food Res. Int. 2012, 45, 287–298. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).