
Exploring the Potential Lipid-Lowering and Weight-Reducing Mechanisms of FH06 Fermented Beverages Based on Non-Targeted Metabolomics and Network Pharmacology

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of FH06
2.2. UHPLC-QE-MS Conditions
2.3. FH06 Compound Screening and Target Prediction
2.4. Disease Target Prediction
2.5. PPI Network Construction Analysis of Effective Target Proteins
2.6. Effective Target-GO Enrichment and KEGG Pathway Analysis
2.7. Metabolite–Target–Pathway Network Construction Analysis
3. Results and Analysis
3.1. FH06 Compound Screening and Target Prediction
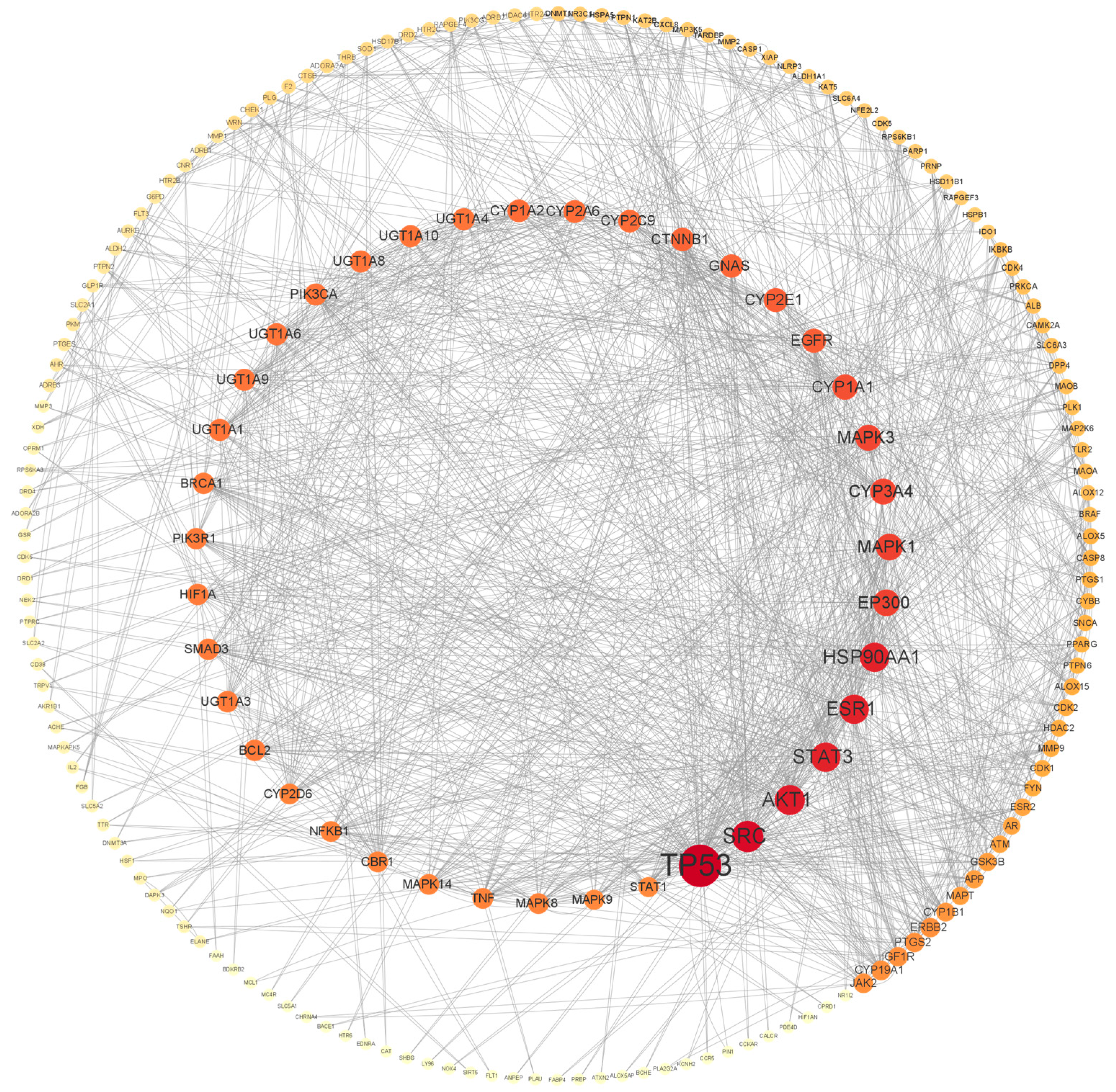
3.2. Construction of the PPI Network of the FH06 Target Protein
3.3. GO Enrichment Analysis of the Effect of FH06 on Therapeutic Targets for Obesity
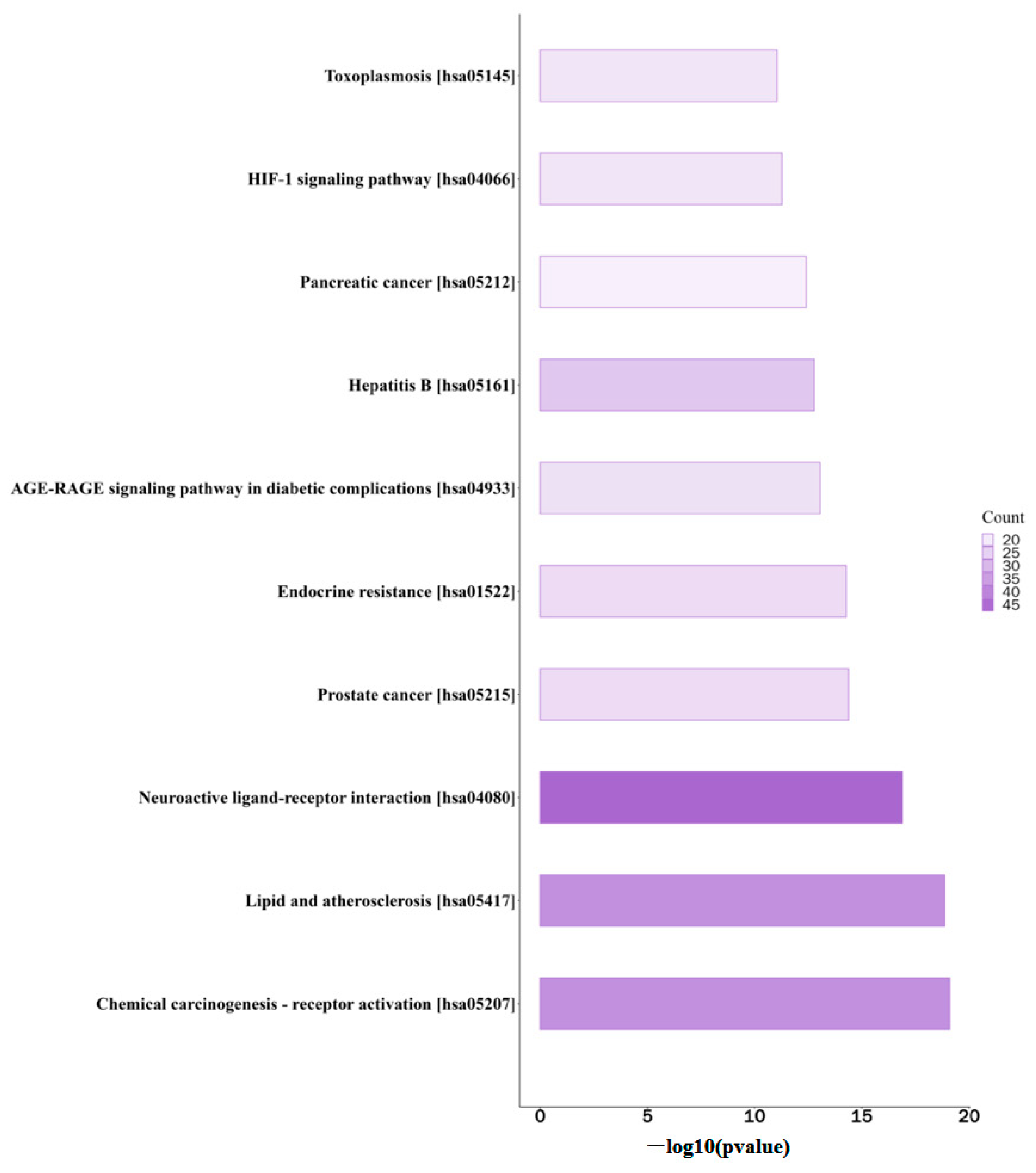
3.4. KEGG Analysis of the Effective Compounds of FH06 That Act on Therapeutic Targets for Obesity
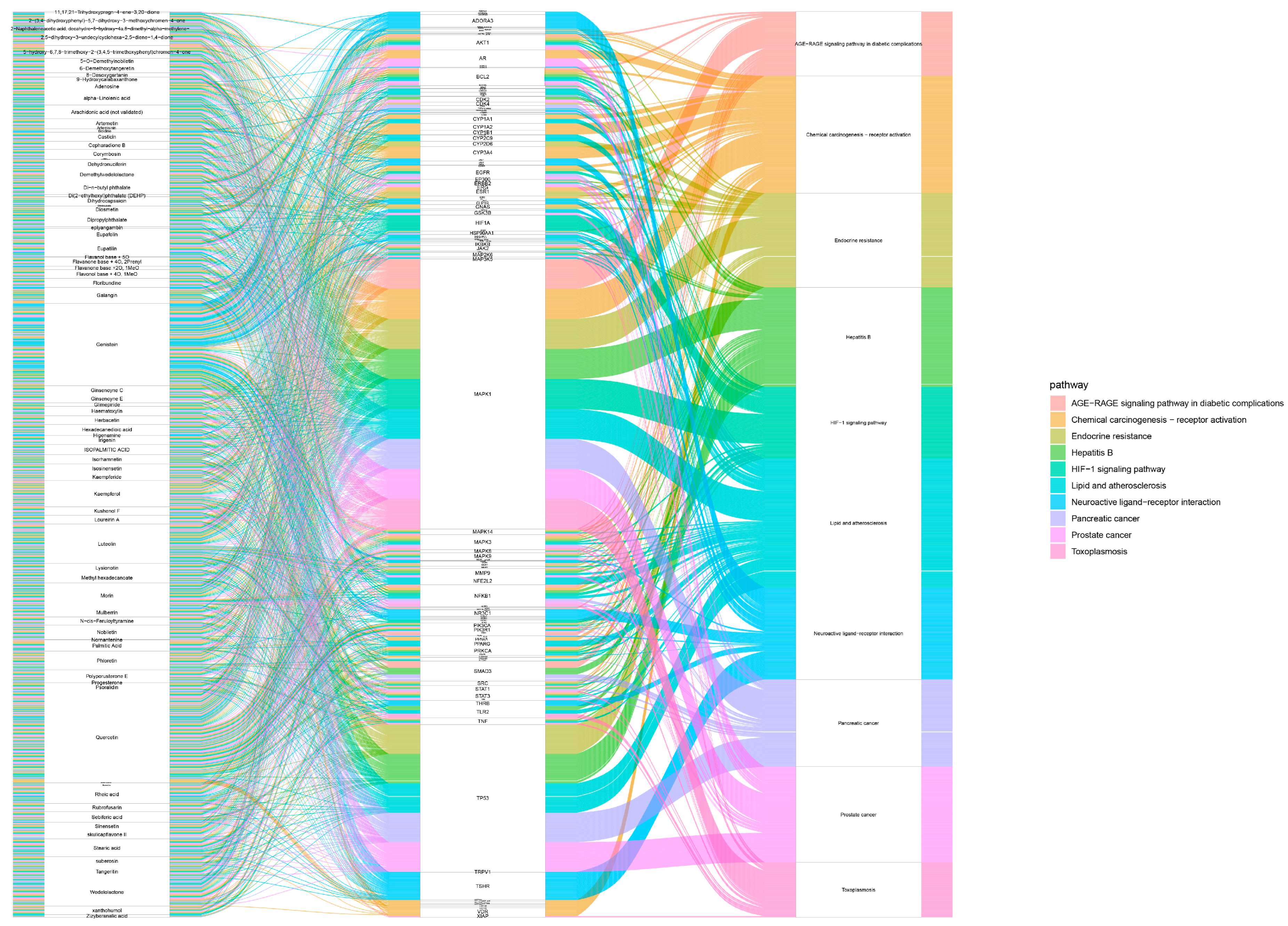
3.5. Construction of the FH06 Effective Compound–Target–Pathway Network
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Ha, S.; Lau, H.C.; Yu, J. Excess body weight: Novel insights into its roles in obesity comorbidities. Semin. Cancer Biol. 2023, 92, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y. Expert consensus on drug treatment management path for patients with coronary atherosclerotic heart disease. Clin. Medicat. J. 2023, 21, 1–18. [Google Scholar]
- Ding, J.W.; Chi, X.Y.; Zhang, Y.; Wang, L.L.; Jiang, J.D.; Lin, Y. The biological principle of “food and medicine homology”. Acta Pharm. Sin. 2024, 1–22. [Google Scholar] [CrossRef]
- Yan, X.-T.; Zhang, W.; Zhang, Y.; Zhang, Z.; Chen, D.; Wang, W.; Ma, W.; Qu, H.; Qian, J.-Y.; Gu, R. In Vitro Anti-Obesity Effect of Shenheling Extract (SHLE) Fermented with Lactobacillus fermentum grx08. Foods 2022, 11, 1221. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Ye, Y.; Xiao, L.; Duan, X.; Zhang, Y.; Zhang, H. Anticancer effects of ginsenoside Rg3 (Review). Int. J. Mol. Med. 2017, 39, 507–518. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.T.; Zhang, Z.; Wang, Y.; Zhang, W.; Zhang, L.; Liu, Y.; Chen, D.; Wang, W.; Ma, W.; Qian, J.Y.; et al. Antioxidant capacity, flavor and physicochemical properties of FH06 functional beverage fermented by lactic acid bacteria: A promising method to improve antioxidant activity and flavor of plant functional beverage. Appl. Biol. Chem. 2023, 66, 7. [Google Scholar] [CrossRef]
- Su, S.B. Problems and Countermeasures in the Development of Network Pharmacology Research. Chin. J. Pharmacol. Toxicol. 2023, 37, 5. [Google Scholar]
- Wei, T.T.; Chandy, M.; Nishiga, M.; Zhang, A.; Kumar, K.K.; Thomas, D.; Manhas, A.; Rhee, S.; Justesen, J.M.; Chen, I.Y.; et al. Cannabinoid receptor 1 antagonist genistein attenuates marijuana-induced vascular inflammation. Cell 2022, 185, 1676–1693.e1623. [Google Scholar] [CrossRef] [PubMed]
- Jung, C.H.; Cho, I.; Ahn, J.; Jeon, T.I.; Ha, T.Y. Quercetin reduces high-fat diet-induced fat accumulation in the liver by regulating lipid metabolism genes. Phytother. Res. 2013, 27, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Ji, G.; Cao, A.; Shi, J.; Shi, H.; Xie, J.; Wu, D. Study on the effect and mechanism of flavanone on proliferation and apoptosis of human gastric cancer AGS cells. China J. Chin. Mater. Medica 2011, 36, 790–794. [Google Scholar]
- Kang, S.I.; Shin, H.S.; Ko, H.C.; Kim, S.J. Effects of sinensetin on lipid metabolism in mature 3T3-L1 adipocytes. Phytother. Res. 2013, 27, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Androutsopoulos, V.P.; Ruparelia, K.; Arroo, R.R.; Tsatsakis, A.M.; Spandidos, D.A. CYP1-mediated antiproliferative activity of dietary flavonoids in MDA-MB-468 breast cancer cells. Toxicology 2009, 264, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Torres-Villarreal, D.; Camacho, A.; Castro, H.; Ortiz-Lopez, R.; de la Garza, A.L. Anti-obesity effects of kaempferol by inhibiting adipogenesis and increasing lipolysis in 3T3-L1 cells. J. Physiol. Biochem. 2019, 75, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y. Study on anti-aging and antioxidant effects of isoflavones. China Surfactant Deterg. Cosmet. 2024, 54, 313–319. [Google Scholar]
- Li, Q.Q.; Li, Z.L.; Liu, H.; Huang, L.J.; Wang, H.; Yang, Z.Y.; Chen, G.Y.; Zhang, C.X. The effect and mechanism of prickly pear fruit polysaccharides on proliferation and apoptosis of lung adenocarcinoma A549 cells. Tradit. Chin. Drug Res. Clin. Pharmacol. 2024, 35, 484–492. [Google Scholar] [CrossRef]
- Maji, A.K.; Mahapatra, S.; Banerji, P.; Banerjee, D. Immunomodulatory effect of Wedelia chinensis and demethylwedelolactone by interfering with various inflammatory mediators. Orient. Pharm. Exp. Med. 2015, 15, 23–31. [Google Scholar] [CrossRef]
- Gimlihan, S.; Baimantimin, N.; Ayniwal, W.-B.; Baidan, Y.; Nuribia, B.; Muhaddis, N.; Zhou, W.-T. Network pharmacological study on the mechanism of action of the active ingredients of the anti-inflammatory oral solution of Pueraria Mirifica. J. Shenyang Pharm. Univ. 2020, 37, 1117–1124+1143. [Google Scholar] [CrossRef]
- Goldstein, I.; Ezra, O.; Rivlin, N.; Molchadsky, A.; Madar, S.; Goldfinger, N.; Rotter, V. p53, a novel regulator of lipid metabolism pathways. J. Hepatol. 2012, 56, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Liu, L.; Zheng, F.; Ma, Y.; Zhang, J. The potent role of Src kinase-regulating glucose metabolism in cancer. Biochem. Pharmacol. 2022, 206, 115333. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Xiao, Y.; Yang, P.; Wang, R. Pan-cancer Analysis Reveals SRC May Link Lipid Metabolism and Macrophages. Iran. J. Biotechnol. 2023, 21, e3325. [Google Scholar] [CrossRef] [PubMed]
- Hua, T.N.M.; Kim, M.K.; Vo, V.T.A.; Choi, J.W.; Choi, J.H.; Kim, H.W.; Cha, S.K.; Park, K.S.; Jeong, Y. Inhibition of oncogenic Src induces FABP4-mediated lipolysis via PPARγ activation exerting cancer growth suppression. EBioMedicine 2019, 41, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Xia, L.; Goldberg, H.J.; Lee, K.W.; Shah, A.; Stavar, L.; Masson, E.A.; Momen, A.; Shikatani, E.A.; John, R.; et al. Inhibition of Src kinase blocks high glucose-induced EGFR transactivation and collagen synthesis in mesangial cells and prevents diabetic nephropathy in mice. Diabetes 2013, 62, 3874–3886. [Google Scholar] [CrossRef]
- Eshaghi, F.S.; Ghazizadeh, H.; Kazami-Nooreini, S.; Timar, A.; Esmaeily, H.; Mehramiz, M.; Avan, A.; Ghayour-Mobarhan, M. Association of a genetic variant in AKT1 gene with features of the metabolic syndrome. Genes. Dis. 2019, 6, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Albury-Warren, T.M.; Pandey, V.; Spinel, L.P.; Masternak, M.M.; Altomare, D.A. Prediabetes linked to excess glucagon in transgenic mice with pancreatic active AKT1. J. Endocrinol. 2016, 228, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Rotllan, N.; Wanschel, A.C.; Fernández-Hernando, A.; Salerno, A.G.; Offermanns, S.; Sessa, W.C.; Fernández-Hernando, C. Genetic Evidence Supports a Major Role for Akt1 in VSMCs During Atherogenesis. Circ. Res. 2015, 116, 1744–1752. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Zou, T.; Shen, X.; Nelson, P.J.; Li, J.; Wu, C.; Yang, J.; Zheng, Y.; Bruns, C.; Zhao, Y.; et al. Lipid metabolism in cancer progression and therapeutic strategies. MedComm 2021, 2, 27–59. [Google Scholar] [CrossRef] [PubMed]
- Huo, N.; Cong, R.; Sun, Z.J.; Li, W.C.; Zhu, X.; Xue, C.Y.; Chen, Z.; Ma, L.Y.; Chu, Z.; Han, Y.C.; et al. STAT3/LINC00671 axis regulates papillary thyroid tumor growth and metastasis via LDHA-mediated glycolysis. Cell Death Dis. 2021, 12, 799. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Song, Z.; Han, Q.; Zhao, H.; Pan, Z.; Lei, Z.; Zhang, J. Targeted inhibition of STAT3 induces immunogenic cell death of hepatocellular carcinoma cells via glycolysis. Mol. Oncol. 2022, 16, 2861–2880. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.M.; Brown, K.A.; Iyengar, N.M. Targeting obesity-related dysfunction in hormonally driven cancers. Br. J. Cancer 2021, 125, 495–509. [Google Scholar] [CrossRef] [PubMed]
- WU, J.-M. Exploring the Effects of Astragalus and Pueraria Mirifica Root Preparation on Glycolipid Metabolism in Skeletal Muscle of Diabetic Rats Based on PI3K/Akt and APN/AMPK Signaling Pathways. Bachelor’s Thesis, Liaoning University of Chinese Medicine, Shenyang, China, 2020. [Google Scholar]
- Zhao, J.-L.; Zeng, L.-F.; Liang, G.-H.; Huang, H.-T.; Yang, W.-Y.; Luo, M.-H.; Pan, J.-K.; Li, J.-H.; Liu, J. Advances in the mechanism of osteoporosis treatment by active ingredients of traditional Chinese medicine based on signaling pathway. Chin. Herb. Med. 2020, 51, 6084–6094. [Google Scholar]
- Cong, J.-F.; Fan, Y.; Li, X.; Liu, Q.; Liu, L. Effects of effective components of Astragalus and Pueraria Mirifica Tang on PI3K/Akt signaling pathway in liver of diabetic rats based on analytical factorial design. J. Tradit. Chin. Med. 2020, 35, 815–820. [Google Scholar] [CrossRef]
- Ma, H.; Gao, N.-N.; Chen, Y.-Y.; Xiao, Y.; Pan, Y.-L. Effects of Glucotonic Drink on PI3K-AKT pathway in pancreatic tissues of rats with type 2 diabetes mellitus. J. Guizhou Med. Univ. 2020, 45, 50–55. [Google Scholar] [CrossRef]
- Jiang, Z.-Y.; Xiong, H.-L.; Yao, J.-J. Effects of glucose and vitamin C on ACC, FAS and CPTⅠactivities of yolk sac pups of silver crucian carp (Carassius auratus auratus L.) in Puan. J. Zool. 2014, 49, 904–912. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Li, W.-X.; Zhang, H.; Zhang, M.-L.; Meng, X.-L.; Jia, W.-H.; Niu, L.; Wang, Y.; Tang, J.-F. Lipid-regulating effect of Schisandra chinensis-Glycyrrhiza glabra combination and its effect on triglyceride synthesis pathway. Chin. J. Pharmacol. 2021, 37, 136–142. [Google Scholar]
- Song, S.-Z.; Wu, J.-P.; Gao, L.-S.; Li, H. Progress of peroxisome proliferator-activated receptor gamma signaling pathway in the regulation of lipid metabolism. Chin. J. Anim. Nutr. 2020, 32, 1473–1483. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Id | Compound | OB | DL |
|---|---|---|---|---|
| 1 | M375.249T650.399 | Glyceryl linolenate | 100.00 | 0.81 |
| 2 | M280.133T192.065 | Remerine | 100.00 | 0.37 |
| 3 | M373.128T336.748 | Sinensetin | 100.00 | 0.75 |
| 4 | M373.128T305.926 | Isosinensetin | 100.00 | 0.67 |
| 5 | M441.371T206.852 | Sebiferic acid | 100.00 | 0.92 |
| 6 | M343.117T343.045 | 6-Demethoxytangeretin | 100.00 | 0.58 |
| 7 | M301.034T93.965 | Demethylwedelolactone | 54.37 | 0.30 |
| 8 | M282.149T143.561 | Floribundine | 96.37 | 0.33 |
| 9 | M303.050T64.081 | Herbacetin | 54.18 | 0.27 |
| 10 | M295.226T315.850 | Isopalmitic acid | 87.27 | 0.69 |
| 11 | M301.072T152.445 | Hematoxylin | 61.51 | 0.32 |
| 12 | M287.055T126.555 | Kaempferol | 64.44 | 0.24 |
| 13 | M389.124T299.923 | Artemetin | 100.00 | 0.77 |
| 14 | M409.163T399.221 | 9-Hydroxycalabaxanthone | 100.00 | 0.97 |
| 15 | M283.152T143.561 | Artemisinin | 96.33 | 0.28 |
| 16 | M421.163T63.588 | Mulberrin | 89.55 | 0.88 |
| 17 | M489.214T300.675 | Glimepiride | 76.66 | 0.72 |
| 18 | M423.180T68.924 | Flavanone base + 4O, 2Prenyl | 85.94 | 0.89 |
| 19 | M403.138T328.764 | Nobiletin | 100.00 | 0.80 |
| 20 | M395.109T306.267 | Tangeritin | 100.00 | 0.70 |
| 21 | M299.056T261.149 | Diosmetin | 74.04 | 0.33 |
| 22 | M301.071T260.804 | Kaempferide | 78.85 | 0.32 |
| 23 | M314.138T191.030 | N-cis-Feruloyltyramine | 87.65 | 0.64 |
| 24 | M279.159T605.154 | Di-n-butyl phthalate | 100.00 | 0.36 |
| 25 | M273.076T128.611 | Rubrofusarin | 91.08 | 0.23 |
| 26 | M287.055T202.797 | Luteolin | 61.50 | 0.25 |
| 27 | M255.102T37.307 | 5-Methoxyflavanone | 100.00 | 0.22 |
| 28 | M287.091T224.607 | Flavanone base +2O, 1MeO | 88.64 | 0.28 |
| 29 | M391.285T805.080 | Di(2-ethylhexyl)phthalate (DEHP) | 100.00 | 0.82 |
| 30 | M347.219T557.971 | Reichstein’s substance S | 80.36 | 0.66 |
| 31 | M469.181T109.966 | Epiyangambin | 100.00 | 0.99 |
| 32 | M315.255T507.075 | Methyl hexadecanoate | 100.00 | 0.72 |
| 33 | M389.123T459.238 | 5-O-Demethylnobiletin | 100.00 | 0.73 |
| 34 | M357.098T301.842 | Corymbosin | 100.00 | 0.64 |
| 35 | M293.179T626.901 | 2,5-dihydroxy-3-undecylcyclohexa-2,5-diene-1,4-dione | 81.20 | 0.66 |
| 36 | M287.128T267.648 | Loureirin A | 100.00 | 0.36 |
| 37 | M419.134T465.131 | 5-hydroxy-6,7,8-trimethoxy-2-(3,4,5-trimethoxyphenyl)chromen-4-one | 100.00 | 0.88 |
| 38 | M373.093T374.031 | Casticin | 93.05 | 0.67 |
| 39 | M308.220T494.010 | Dihydrocapsaicin | 100.00 | 0.75 |
| 40 | M305.068T43.711 | Flavanol base + 5O | 35.45 | 0.31 |
| 41 | M315.232T750.032 | Progesterone | 100.00 | 0.52 |
| 42 | M251.125T252.620 | Dipropyl phthalate | 100.00 | 0.25 |
| 43 | M317.066T102.739 | 2-(3,4-dihydroxyphenyl)-5,7-dihydroxy-3-methoxychromen-4-one | 65.16 | 0.32 |
| 44 | M315.052T321.661 | Eupafolin | 66.17 | 0.37 |
| 45 | M337.105T475.812 | Psoralidin | 91.75 | 0.63 |
| 46 | M315.052T267.813 | Isorhamnetin | 65.76 | 0.34 |
| 47 | M268.104T53.746 | Adenosine | 42.37 | 0.20 |
| 48 | M301.036T247.331 | Quercetin | 52.23 | 0.27 |
| 49 | M313.035T211.574 | Wedelolactone | 68.16 | 0.38 |
| 50 | M279.232T477.337 | Alpha-linolenic acid | 87.30 | 0.46 |
| 51 | M343.083T359.527 | Eupatilin | 93.25 | 0.54 |
| 52 | M272.128T93.913 | Higenamine | 66.38 | 0.23 |
| 53 | M275.162T261.545 | 2-Naphthaleneacetic acid, decahydro-8-hydroxy-4a,8-dimethyl-alpha-methylene- | 84.87 | 0.18 |
| 54 | M277.179T577.922 | Ginsenoyne C | 94.40 | 0.61 |
| 55 | M330.133T154.081 | N-trans-Feruloyloctopamine | 80.75 | 0.59 |
| 56 | M461.324T753.044 | Polyporusterone E | 91.69 | 0.84 |
| 57 | M326.139T125.820 | Nornantenine | 100.00 | 0.57 |
| 58 | M379.158T751.393 | 8-Desoxygartanin | 100.00 | 0.75 |
| 59 | M423.180T118.178 | Kushenol F | 90.75 | 0.93 |
| 60 | M431.211T588.064 | Grandisin | 100.00 | 0.92 |
| 61 | M328.154T117.982 | Boldine | 83.23 | 0.51 |
| 62 | M313.274T752.302 | 2,3-dihydroxypropyl hexadecanoate | 100.00 | 0.58 |
| 63 | M333.061T100.848 | Flavonol base + 4O, 1MeO | 54.06 | 0.38 |
| 64 | M327.159T169.368 | Dehydrodiisoeugenol | 100.00 | 0.62 |
| 65 | M375.107T236.493 | Skullcapflavone II | 93.75 | 0.56 |
| 66 | M259.167T418.190 | Ginsenoyne E | 100.00 | 0.58 |
| 67 | M343.082T336.916 | Lysionotin | 96.66 | 0.54 |
| 68 | M255.233T20.106 | Palmitic acid | 87.40 | 0.72 |
| 69 | M294.149T131.469 | Dehydronuciferine | 100.00 | 0.38 |
| 70 | M262.144T577.956 | Suberosin | 100.00 | 0.20 |
| 71 | M269.046T918.356 | Genistein | 76.33 | 0.22 |
| 72 | M301.036T313.918 | Morin | 53.80 | 0.27 |
| 73 | M340.154T255.885 | Crebanine | 100.00 | 0.63 |
| 74 | M283.025T337.240 | Rheic acid | 47.04 | 0.20 |
| 75 | M283.265T26.412 | Stearic acid | 91.98 | 0.68 |
| 76 | M275.092T162.583 | Phloretin | 72.81 | 0.25 |
| 77 | M359.078T271.495 | Irigenin | 83.91 | 0.60 |
| 78 | M271.060T246.155 | Galangin | 77.70 | 0.20 |
| 79 | M322.107T398.936 | Cepharadione B | 93.08 | 0.42 |
| 80 | M305.247T310.008 | Arachidonic acid (not validated) | 90.26 | 0.48 |
| 81 | M359.149T221.852 | Dihydrocubebin | 100.00 | 0.61 |
| 82 | M353.143T742.445 | Xanthohumol | 100.00 | 0.69 |
| 83 | M363.214T474.995 | 11,17,21-Trihydroxypregn-4-ene-3,20-dione | 73.69 | 0.68 |
| 84 | M453.337T279.747 | Zizyberanalic acid | 84.02 | 0.85 |
| 85 | M285.208T343.820 | Hexadecanedioic acid | 65.23 | 0.74 |
| Node | Degree |
|---|---|
| Genistein | 130 |
| Quercetin | 124 |
| Luteolin | 68 |
| Kaempferol | 59 |
| Rheic acid | 35 |
| Morin | 33 |
| Galangin | 33 |
| Phloretin | 29 |
| Wedelolactone | 27 |
| Alpha-linolenic acid | 27 |
| 2,5-dihydroxy-3-undecylcyclohexa-2,5-diene-1,4-dione | 27 |
| Hematoxylin | 21 |
| Arachidonic acid (not validated) | 21 |
| Diosmetin | 17 |
| Stearic acid | 16 |
| Palmitic acid | 16 |
| Nobiletin | 16 |
| 2-(3,4-dihydroxyphenyl)-5,7-dihydroxy-3-methoxychromen-4-one | 16 |
| Isorhamnetin | 15 |
| Eupafolin | 15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Wang, T.; Wang, J.; Liu, R.; Cui, Y.; Wu, X.; Dai, R.; Wu, Y.; Nie, X.; Yan, X.; et al. Exploring the Potential Lipid-Lowering and Weight-Reducing Mechanisms of FH06 Fermented Beverages Based on Non-Targeted Metabolomics and Network Pharmacology. Fermentation 2024, 10, 294. https://doi.org/10.3390/fermentation10060294
Wang H, Wang T, Wang J, Liu R, Cui Y, Wu X, Dai R, Wu Y, Nie X, Yan X, et al. Exploring the Potential Lipid-Lowering and Weight-Reducing Mechanisms of FH06 Fermented Beverages Based on Non-Targeted Metabolomics and Network Pharmacology. Fermentation. 2024; 10(6):294. https://doi.org/10.3390/fermentation10060294
Chicago/Turabian StyleWang, Haoming, Ting Wang, Jinghan Wang, Ronghan Liu, Yingying Cui, Xiurong Wu, Rui Dai, Yanglin Wu, Xiangzhen Nie, Xiantao Yan, and et al. 2024. "Exploring the Potential Lipid-Lowering and Weight-Reducing Mechanisms of FH06 Fermented Beverages Based on Non-Targeted Metabolomics and Network Pharmacology" Fermentation 10, no. 6: 294. https://doi.org/10.3390/fermentation10060294