Abstract
Lactic acid bacteria (LAB), due to their many advantageous features, have been utilized in food manufacturing for centuries. Spontaneous fermentation, in which LAB play a fundamental role, is one of the oldest methods of food preservation. LAB survival and viability in various food products are of great importance. During technological processes, external physicochemical stressors appear often in combinations. To ensure the survival of LAB, adjustment of optimal physicochemical conditions should be considered. LAB strains should be carefully selected for particular food matrices and the technological processes involved. The LAB’s robustness to different environmental stressors includes different defense mechanisms against stress, including the phenomenon of adaptation, and cross-protection. Recently established positive health effects and influence on human wellbeing have caused LAB to be some of the most desirable microorganisms in the food industry. A good understanding of LAB defense and adaptation mechanisms can lead to both optimization of food production and storage conditions, as well as to obtaining LAB strains with increased tolerance to stressors. Hopefully, as a result, the final food product with naturally present or added LAB can achieve outstanding quality and safety with health benefits that meet consumer expectations.
1. Introduction
Lactic acid bacteria (LAB) have been and are used in the production of a wide variety of food products. Before microorganisms were known and isolated, spontaneous lactic fermentation was applied to preserve food and obtain products with desired sensory properties. The oldest records of the production and consumption of fermented milk products come from ancient Abyssinia, Assyria, Babylon, Egypt, China, Europe, and Palestine [1,2,3]. LAB consist of several genera, e.g., Lactiplantibacillus, Lacticaseibacillus, Levilactobacillus, Leuconostoc, Pediococcus, Lactococcus—lactic streptococci—Streptococcus, Enterococcus, Carnobacterium, Weissella, and Oenococcus. Currently, a new taxonomic nomenclature is in force, according to which the previous genus of Lactobacillus has been divided into 23 new genera [4]. In general, the LAB are characterized as Gram-positive, non-sporulating, aerobic to facultatively anaerobic (aerotolerant) bacteria that produce lactic acid during fermentation. These are asporogenous rods, cocci, or coccobacilli that are oxidase, catalase, and benzidine negative, lack cytochromes, do not reduce nitrates to nitrite, and are gelatinase negative.
The LAB, based on the end product of their fermentation, are grouped as either obligatory homofermentative, obligatory heterofermentative, or facultative heterofermentative [2,5,6]. The homofermentatives produce lactic acid as the major product of the fermentation of glucose. The heterofermentatives use the phosphoketolase pathway and produce several products besides lactic acid, including carbon dioxide, acetic acid, and ethanol, from the fermentation of glucose. The obligatory homofermentatives possess the enzyme aldolase and can ferment glucose more directly to lactic acid than the heterofermentatives. The facultative heterofermentatives degrade hexoses via glycolysis and pentoses through the phosphoketolase pathway [7].
Spontaneous fermentation, in which LAB play a fundamental role, is one of the oldest methods of food preservation. LAB are commonly present in food, either entering spontaneously or being added during processing. Due to their long history of use, they have Generally Recognized As Safe (GRAS) status or Qualified Presumption of Safety (QPS) status [8,9]. In addition to preservation, LAB play an important role in shaping the sensory quality of food: odor, flavor, color, and texture.
In food processing and storage, as well as during gastrointestinal transit, LAB often encounter different environmental conditions, which can sometimes be extreme. The main stress factors include high and low temperature, acidic and alkaline pH, oxidative stress, high hydrostatic pressure, osmotic pressure, and starvation. According to Booth et al., the definition of stress covers any change in the genome, proteome, or environment that imposes either reduced growth or survival potential. Such changes lead to attempts by a cell to restore a pattern of metabolism that fits it either for survival or faster growth [10]. In addition, the viability of the LAB is affected by many biological factors, such as the type of strain, bacterial growth interactions with starter cultures and the natural microbiota of the product, sensitivity to metabolites produced by other competing bacteria, and various pathogenic and putrefactive microorganisms [11,12]. Exposure of LAB to environmental and biological stress can be harmful. Metabolism and many cell functions can be affected and, finally, cell survival may be compromised. Knowledge about how LAB combat environmental stress to survive and understanding of the adaptation mechanisms can be used in various steps of food production, processing, and storage.
The cell viability of LAB is defined as the ability to grow and form cell colonies under certain environmental conditions [13]. In general, the viability of LAB in food matrix during fermentation as well as during other manufacturing processes and their stability during transport and storage are strongly dependent on the specific bacterial strain. Identification of individual strains of LAB enables the proper selection of both starter cultures and cultures added to various food products. LAB may be added to food products as probiotic or bioprotective adjunct cultures in the form of frozen, dried, encapsulated, or liquid cultures. The use of optimal physicochemical conditions for LAB, both in production processes and during food storage, ensures the survival of LAB in high numbers, guaranteeing the expected health-promoting effect.
The food matrix structure was not found to be a dominant factor for LAB survival. It is difficult to determine conclusively to what extent the physical structure of the matrix, i.e., solid, gel, emulsion, or liquid, affects the viability of the LAB. However, more aerated matrices due to oxidative stress may adversely affect LAB viability. The content and proportions of nutrients in the matrix and physicochemical parameters such as temperature, pH, water content, water activity (aw), O2 content, and access of O2 to the food product are of key importance for LAB viability. In addition, the buffer capacity of the food matrix also plays an important role in the protection of the LAB, reducing the impact of excessive acidification [14].
In response to environmental physicochemical stress factors, LAB exhibit multiple adaptive mechanisms [15]. LAB do not produce spore forms, as, e.g., Bacillus strains do. To survive in adverse conditions, they can, among other responses, limit basic life functions, such as colony formation. The sensitivity of bacteria to stress factors depends on the stage of their development, the occurrence of previous stresses, as well as the conditions of regeneration after the stress factor [16]. Any bacterial cell is far more sensitive to any (sudden) stress in the exponential growth phase than in any (sudden or long-term) stress in the stationary phase. Cells are more resistant to stress in their stationary compared to exponential phase because they always develop/possess a generalized stress response (GSR) system, regulated mainly by RpoS to face upcoming starvation; the GSR is independent of specific stress [17,18]. On this basis, bacteria (including LAB) in most foods actually are in the stationary phase, trying to develop/maintain the least energy-consuming adaptive responses for long-term survival, rather than in the exponential growth, including all postfermented foods during ripening and/storage [19]. Overall, when response to a stress is not required, the genes involved are switched off, which never happens in the stationary phase under RpoS-regulated GSR conditions.
According to Davis, non-culturable microorganisms are classified into four groups: (1) the non-replicating state (active physiology and membrane integrity), (2) the starving state (dramatic decrease in metabolism), (3) the dormant state (low metabolic activity and inability to divide without additional recovery attempts, viable but non-culturable (VBNC)), and (4) the irreparably damaged state (progressively declining metabolism that terminates in death) [20].
Many preservation and pasteurization techniques are used in the food industry, including traditional, thermal methods, water content reduction (drying, addition of sugar or salt), and addition of various preservatives. These methods are mainly used to eliminate unfavorable bacteria to ensure the microbiological safety of foods. Unfortunately, they also inhibit the growth of LAB and even lead to inactivation of bacterial cells. In addition, adverse changes may affect the quality of food products, including the reduction of nutrients and bioactive compounds (e.g., heat-sensitive vitamins or polyphenols) [16]. Therefore, during food processing, it is necessary to take into account both the resistance and adaptability of the LAB strain to different physicochemical conditions as well as to consider the optimization of physicochemical parameters during food processing, storage, and transport.
This review article discusses the impact of physicochemical stressors confronted by LAB during food manufacturing. LAB are subjected to different environmental stresses such as temperature and pH changes and osmotic and oxidative stresses. Metabolism and many cell functions can be affected and, finally, cell survival may be compromised. The impact of the main stressors on LAB was characterized. A deep understanding of the influence of selected physicochemical stressors on LAB can be complicated by the coexistence of various stressors. The survival rate of LAB in food products is described, with particular emphasis on the selection of optimal conditions during food processing and storage. The LAB defense mechanisms against stress, including the phenomenon of adaptation and cross-protection, are presented. Some methods to reduce the occurrence and impact of stress factors on LAB including the addition of cytoprotective agents, synergistic microbial interactions, encapsulation, and new non-thermal methods of food preservation are presented. Taking into account the importance of LAB in food production and their potential health-promoting impact, knowledge of the impact of environmental and bacterial factors and understanding of defense mechanisms can lead to optimization of both food production and storage conditions, as well as to obtaining LAB strains with increased tolerance to stressors.
2. Effect of Temperature on LAB Survival
The optimal growth temperature for most LAB ranges from 30 to 45 °C [11]. Some thermophilic strains show viability at temperatures up to 60 °C. The homofermentative Lactobacillus acidophilus and Streptococcus thermophilus can grow at temperatures as high as 45 °C with optimum growth from 40–45 °C [21,22,23]. When the temperature reduces beyond the optimal range for exponential colony growth, the bacterial cells are subjected to thermal stress [24]. Even small temperature differences can have a significant impact on the LAB viability. In the study of Neffe-Skocińska et al., the L. casei LOCK900 strain added to loins has a better survival rate during the 21-day ripening of fermented pork at 20 °C (from 107 to 108 cfu/g) compared to samples aged at 16 °C (from 104 to 106 cfu/g) [25]. Therefore, the temperature in various steps of food production and storage should be appropriate for specific LAB strains. This issue is especially important for probiotic-based foods.
Food processing often requires the use of higher temperatures, e.g., in the pasteurization process, which exposes the LAB to heat stress. Heat treatment of food may result in the inactivation of probiotic bacterial cultures. Thermophilic LAB strains can tolerate temperatures up to 65 °C, but in general heat stress above 50 °C is harmful to most LAB. [26]. The extent of cell heat injuries depends on the duration and severity of the heat stress and in severe form can lead to death. Sublethal stress could initiate an adaptive protective mechanism that can lead to stress tolerance [27]. In a study by Kulkarni et al. it was found that due to gradual adaptation, the thermotolerant variant of Lactobacillus acidophilus NCFM can survive at 65 °C at pH 2.0 for 40 min. The authors concluded that the first 10 min at 65 °C caused most cells to change into a non-culturable state, and during the next 10 to 20 min of exposure, only a few cells were able to activate adaptive mechanisms [28]. Therefore, when food processing includes applying high temperatures, bacterial preconditioning should be considered because it is impractical to apply non-resistant strains. The necessity to use high temperatures, above 100 °C, e.g., during baking, cooking, and extrusion, is lethal to the cells and therefore LAB cultures should be added at later stages of food processing, after thermal operations. Extreme temperatures of up to 200 °C with short exposure times are used in spray drying as well as freeze drying down to –196 °C [28]. High temperatures are associated with osmotic and shear stress, which leads to the denaturation of proteins and damage to nucleic acids and cell membranes [29].
In response to heat stress bacterial cells induce the synthesis of proteins called heat shock proteins (HSPs) including the chaperonins (DnaK and GroEL) and cofactors (GroES) and small heat shock proteins (sHSPs). These proteins counteract the harmful effects of high temperature by improving the folding of the incipient polypeptides and by repairing thermal damage to cell structures, including the cell membrane [30]. Exposing LAB to sublethal stress before applying extreme temperatures, i.e., spray drying, can enhance bacterial viability. In the study of Broeckx et al. two methods of L. rhamnosus GG heat adaptation were evaluated: pretreatment with 50 °C for 15 min and cyclic consecutive spray drying (up to 10 spray-drying cycles). Compared to the non-pretreated samples both methods showed enhanced survival rates of L. rhamnosus GG just after spray drying and during long-term storage at room temperature. The spray-drying cycles should be limited to three consecutive cycles because the fourth resulted in a log reduction of approximately 0.6 after drying [31].
Low temperatures above 0 °C used in the fermentation, ripening, and storage of some products are generally not harmful to LAB. However, cellular metabolism decreases and enzyme activity and protein synthesis drop, which in turn can inhibit LAB cell growth. LAB can adapt to decreased temperature and with a reduced rate can grow at a temperature below 20 °C and even at 15 °C, but not at 7 °C [32]. Products such as ice cream and frozen yogurt are usually stored at −20 °C, which can lead to the formation of extracellular ice crystals as well ice crystals within the LAB cells. The ice crystals can damage the cell structures, including the destruction of cell membrane integrity and its fluidity and damage of DNA strands and RNA structure, affecting transcription and replication [33,34]. Another effect of freezing is the dehydration of bacterial cells. This creates another stress—high osmolarity—caused by high concentrations of internal solutes [26]. Finally, this injury can lead to cell death.
Membrane integrity and denaturation of macromolecules have been reported as factors that determine survival after freezing, membrane stiffening, cell dehydration, and volume reduction [35]. In response to cold stress, LAB cells synthesize cold shock proteins (CSPs) including CspL, CspP, and CspC, which alleviates cold stress and contributes to the maintenance of active growth conditions [24,36]. In response to cold stress and heat shock, there is a change in the composition of fatty acids in the cell membrane. The increase in the unsaturated fatty acid/saturated fatty acid (UFA/SFA) ratio enhances the membrane rigidity by adjusting fluidity [37]. In case of a short-term heat shock, it is not possible to rapidly change the lipid composition of cell membranes. In such a situation, heat shock proteins can bind to cell membrane lipids, offsetting the thermal effect of increasing cell membrane fluidity [35]. Wang et al. studied resistance to freezing and frozen storage at −20 °C of Lactobacillus acidophilus RD758. It was revealed that cryoadaptation was related to combined responses. Firstly, there was an increased synthesis of the stress protein ATP-dependent ClpP. Secondly, two cold-induced proteins, pyruvate kinase and a putative glycoprotein endopeptidase, and a higher synthesis of unsaturated and cyclic fatty acids (CFAs) led to a higher UFA/SFA ratio [38]. In the study of Liu et al. L. plantarum K25 isolated from Tibetan kefir grains under cold stress conditions reduced membrane fluidity by the decrease in fatty acid biosynthesis, some proteins increased their abundance, including CSP (CspA, CspB, and CspC), and carbohydrate metabolism was reduced to save energy for survival [39]. The upregulation of the proteins involved in the temperature stress response and modifications in the composition of cell membrane fats are considered to be the main adaptive responses of LAB to environmental stress related to temperature changes [40]. Adaptive mechanisms within certain ranges might efficiently counteract heat stress as well as cold stress. However, further investigations are needed for designing tailor-made LAB food manufacturing.
3. Influence of pH on LAB and Their Adaptation to Acid Stress
As a result of the fermentation of sugars contained in food products, LAB can produce various organic acids. In the biochemical transformations carried out by the LAB, the main end product of sugar fermentation is lactic acid, which lowers the pH of the food matrix. Thus, LABs that are acid-tolerant produce a bacteriostatic and even bactericidal environment harmful to many competitive microorganisms, including putrefactive and pathogenic bacteria. The LAB cell membrane like that of other acidophiles is highly impermeable to proton influx into the cytoplasm [41]. Unfortunately, due to prolonged exposure to acidic conditions or if the pH is too low the lifespan of LAB can be compromised [40]. For the growth of LAB during food production and storage, a neutral or slightly acidic pH is optimal (5.5–6.0). However, Lactobacillus strains have good tolerance to acidic environments and grow in the pH range of 3.7 and 4.3, while Bifidobacterium spp. do not tolerate a drop in pH below 4.0 [42]. Acidification of the cytosol is genotoxic and results in the denaturation of proteins. Autoacidification of the environment by LAB effectively counteracts the growth of competing microorganisms. The antibacterial activities of organic acids along with LAB-produced bacteriocin and other biologically active byproducts such ethanol, enzymes, diacetyl, and exopolysaccharides (EPSs) are employed to counteract undesired, putrefactive, and pathogenic bacteria. However, long-term exposure to acidic conditions also leads to the inhibition of LAB growth and may be lethal to cells as a result [40]. At a low pH, undissociated lactic acid and acetic acid passively diffuse across the membrane into the cell, leading to a decrease in intracellular pH (pHi) [32]. Intracellular acidification significantly reduces enzyme activity and causes permanent damage to the LAB proteins [43]. Foods with high acidity (pH < 4.6), such as fruit juices and yogurts, are challenging for LAB survival, but at the same time, due to the inactivation of undesirable microorganisms in an acidic environment, they may require milder processing methods than products with low acidity (pH > 4.6) [16]. In the study of Sheehan et al. the survival rates of Lactobacillus and Bifidobacterium in orange, pineapple, and cranberry juice were assessed. All studied strains survived at levels above 106 cfu/mL up to 12 weeks in less acidic environments in orange (pH 3.65) and pineapple juice (pH 3.40), while in acidic cranberry juice (pH 2.50), a drop of 3–4 log cycles in viable cells was recorded a few days after inoculation [44].
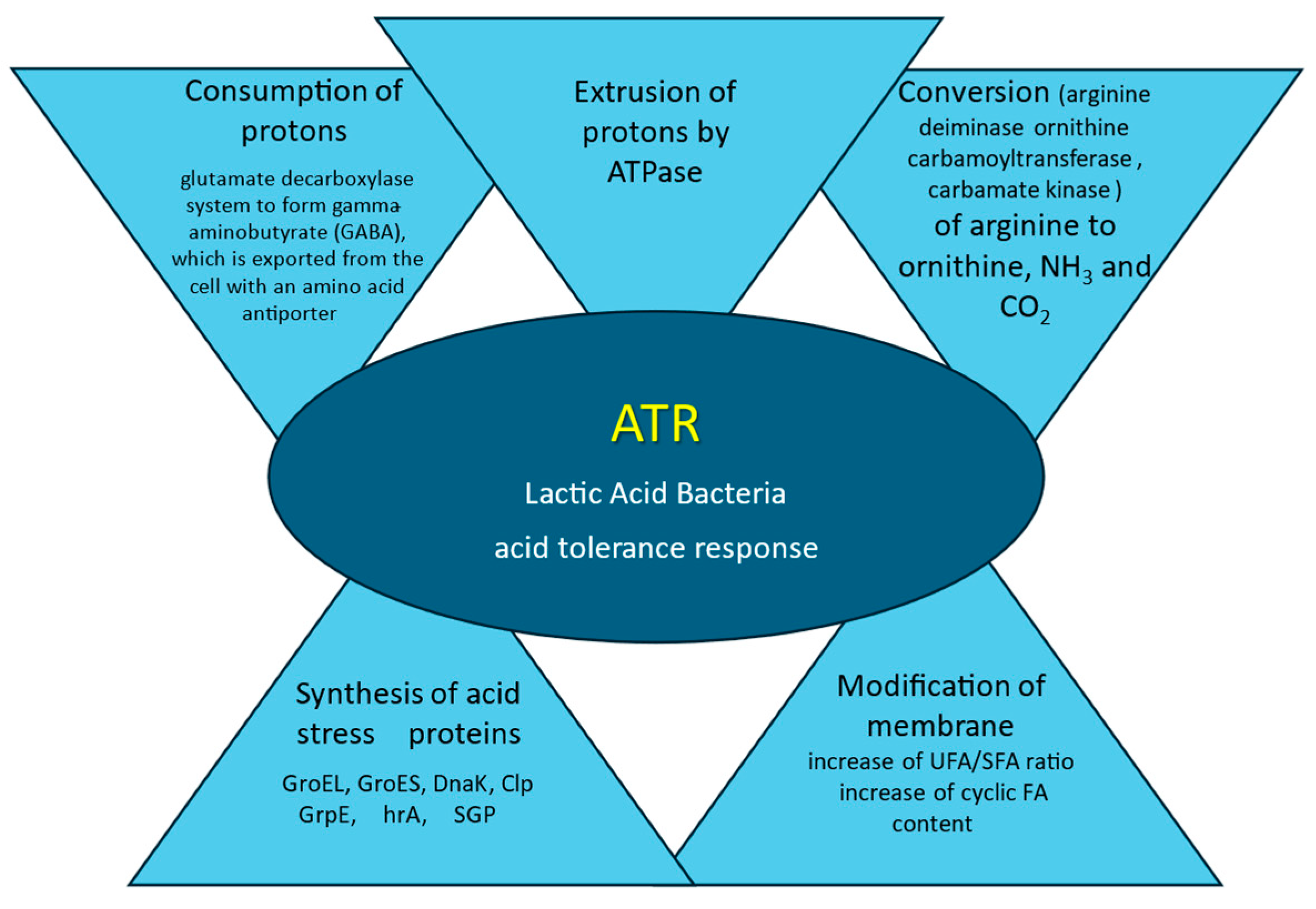
In response to the stress of low pH, bacteria have developed some resistance systems (Figure 1). To counteract acidification and survive in a mild and highly acidic environment LAB activate defense mechanisms that regulate homeostasis, maintaining an intracellular pH of about 6.0, thus also contributing to an increase in acid resistance [45]. The main protective mechanism is the removal from the cytoplasm of accumulated protons (H+) by ATPase, which results in an intracellular increase in pH [32]. Proton motive force (PMF) is a measurement of the energy state of the cell membrane generated by a charge separation between the cytoplasm and external milieu created by membrane potential and pH gradient across the membrane. The balance between the permeability of the cell wall for proton influx and the energetic and transport systems maintains the cell-appropriate PMF [41,46]. Hence, to maintain their viability by fighting acid stress bacterial cells consume large amounts of energy in the form of ATP, depleting energy resources [47,48]. When the energy is exploited to the minimum the cell metabolism is inactivated [49].
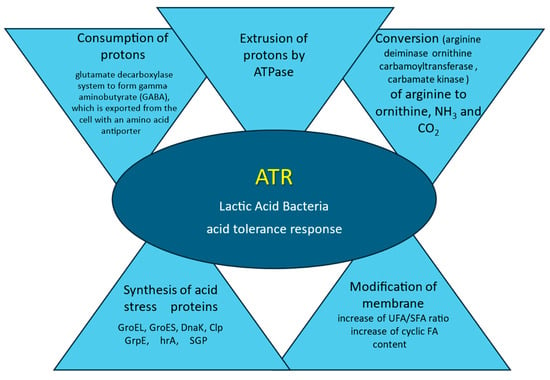
Figure 1.
Mechanism of LAB adaptation to acid stress. According to: de Angelis and Gobetti, 2004 [32], Fonseca et al., 2019 [35], Papadimitriou et al., 2016 [40], Filho et al., 2017 [50], Wu et al., 2014 [51].
The next important mechanism that causes an increase in pH is the enzymatic (arginine deiminase) conversion of arginine to ornithine, ammonia, and CO2. Another intracellular mechanism is the biochemical consumption of protons in the glutamate decarboxylase system to form gamma-aminobutyrate (GABA), which is exported from the cell with an amino acid antiporter [50]. In response to acid stress, LAB cells induce the synthesis of acid stress proteins and chaperones such as GroEL, GroES, DnaK, Clp GrpE, hrA, and SGP. These proteins enhance the protection and regeneration processes, therefore improving bacterial survival rates [51]. To prevent the flow of protons into the cells, the membranes of the LAB cells are modified. The ratio of total SFA to UFA is reduced and membrane fluidity decreases. In defense of proton influx, the modifications of the membrane are accompanied by changes in the expression of the corresponding genes [52].
The complex adaptation strategy includes the above-mentioned mechanisms and synthesis of defending proteins that protect against acid stress from a system that allows the LAB to adapt to a low pH, known as the acid tolerance response (ATR) [53]. In the acidophile microorganism genome a great amount of genes responsible for DNA and protein regeneration are present, reflecting acidic-related injuries that require robust repairs [41]. The results of studies on the influence of pH changes on the fatty acid composition of cell membranes indicate that adaptation to variable pH is strain-dependent [54]. The main reaction to low pH is a decrease in the UFA/SFA ratio and an increase in the cyclic FA content [35].
Many studies investigated the issue of LAB viability and adaptation to acid stress, but few studies investigated the influence of alkaline environments on LAB metabolism and survival. Lactobacilli are generally not alkali-tolerant, however, several alkali-tolerant or alkalotrophic LAB, such as the genera Enterococcus and Carnobacterium, have the ability to grow at pH 8.5 or even higher values [55]. The fermentation of alkaline materials such as Spanish-style green olives and Chinese noodle dough (L. plantarum NRIC 0380) starts with a high pH [56,57]. Sawatari and Yokota studied the maximum pH for growth for 34 strains of Lactobacilli. L. casei NRIC 1917 and L. paracasei subsp. tolerans NRIC 1940 showed the highest alkali tolerance that allows growth up to pH 8.9, while other studied strains showed tolerance at maximum pH of 6.7 and 7.1. According to the authors’ suggestions, a Na+/H+ antiporter and glycolysis reactions could contribute to the survival of bacterial strains in an alkaline environment [58].
4. Influence of Oxygen
LABs are anaerobic or relatively anaerobic bacteria, lacking catalase and superoxide dismutase activity, and susceptible to adverse effects and oxygen toxicity. The exposure to oxidative stress for LAB is harmful and their range depends on the species and strain. It can lead to irreversible damage to the bacterial cell. Generally, LAB tend to be more tolerant to oxygen than bifidobacteria [13,59]. Several LAB strains can metabolize oxygen in reactions of flavin oxidases [60].
The extent of the effect of oxygen on bacterial cells may be related to the type of nutritional matrix (dissolved oxygen), the fragmentation of the matrix (aeration), as well as to the access of atmospheric oxygen during storage of the product (penetration through the packaging material). Oxidative stress due to the presence of oxygen causes intracellular accumulation of toxic aerobic metabolites such as superoxide anion. Oxygen exerts a toxic effect on anaerobic LAB, acting toxically through peroxides and free radicals called reactive oxygen species (ROS) containing at least one oxygen atom and one or more unpaired electrons. This group of oxygen byproducts includes oxygen free radicals, e.g., superoxide anion radical, hydroxyl radical, hydroperoxyl radical, singlet oxygen, reactive nitrogen species (RNS), and reactive sulfur species. As a result of the biochemical oxidation reaction, ROS can damage nucleic acid, leading to its fragmentation, cellular lipids within the plasma membrane, and proteins with the reduction of ATP and depletion of the energy resources [14,61].
Oxidative stress induces adaptive responses in anaerobes. An increase in resistance to oxidative stress in LAB may result in increased viability and longer survival in food products. LAB generate ROS-removing enzymes, including catalases that degrade H2O2, superoxide dismutase converting superoxide radicals to oxygen and water, the NADH oxidase/NADH peroxidase system, pyruvate oxidase (POX), and lactate oxidase that blocks oxygen and ROS accumulation [60,62,63,64]. Talwalkar and Kailasapathy (2004) demonstrated an adaptive response of L. acidophilus and Bifidobacterium spp. strains to increasing concentrations of dissolved oxygen in yogurt (0, 60, 150, and 210 ppm), which made it possible to maintain the viability of the bacteria during 35 days of storage [65]. Nguyen et al. showed that progressive exposure to H2O2 resulted in improvements in the tolerance of a Lacticaseibacillus rhamnosus strain to oxidative stress, which was previously unbearable. The authors found that an increase in cell antioxidant activity was correlated with an increase in mRNA expression for genes responsible for oxidative stress tolerance and bacterial morphological changes [66]. ROS are powerful bacterial stressors but non-lethal oxidative stress as shown above can generate adaptive mechanisms. On the other hand, some LAB strains can produce hydrogen peroxide as an antibacterial antimicrobial.
A high fat content in the food matrix promotes the oxidation reaction, which may influence the occurrence of oxidative stress adversely affecting the survival of LAB. In the study of Klu et al., the survival of Lacticaseibacillus rhamnosus GG as influenced by storage conditions and product matrices was assessed. They report the lower survivability of L. rhamnosus GG at 37 °C in inoculated full fat peanut butter (fat content 50.10 ± 1.16%), reaching an undetectable level at 19 weeks of storage, while in reduced fat butter (fat content 39.90 ± 0.62%) the bacterial count decreased to 3.35 log CFU/g. During 48 weeks of storage at 4 °C, there was no significant difference in counts of L. rhamnosus GG between full fat and reduced fat butter, while at 25 °C significant differences appeared at 27 and 40 weeks of storage. The authors concluded that the survival of L. rhamnosus GG was influenced by storage temperature, storage time, and fat content. It can be explained by the increased rate of lipid oxidation resulting in the formation of ROS including primary and secondary oxidative compounds such as peroxyl, alkoxyl radicals, aliphatic aldehydes, ketones, and alcohols [67].
Certain vitamins, such as vitamins C and E, which help to remove oxidative oxygen, may play a role in reducing the effects of oxidative stress on LAB cells. Therefore, another way to oxidative stress protection can be the addition of oxygen scavengers or antioxidants. In the research of Homayouni et al. the impact of the addition of 0.05% L-cysteine and L-ascorbic on the growth rate of four probiotic bacteria in ice cream in simulated conditions (Lactobacillus acidophilus, Lacticaseibacillus casei, Bifidobacterium bifidum, and Bifidobacterium longum) was assessed. The results disclosed differences in studied strains with the highest growth rate of Lacticaseibacillus casei. The impact of additives also showed distinctions with better biomass production of L. casei with L-cysteine compared to L-ascorbic acid. It can be concluded that the effect of oxygen scavenging on the LAB growth rate is strain-dependent [68].
5. Influence of Osmotic Stress and Water Activity
In food manufacturing and storage LAB often encounter different environmental solute concentrations, resulting in cellular osmotic stress. The traditional method of food preservation is the reduction of water content. As a result of drying or the addition of water-binding solutes, the amount of free water in the food is significantly reduced. The bacterial cell wall is responsible for the shape of the cell and plays a key role in maintaining osmotic pressure. Both too high (hypoosmotic stress) and low water content (hyperosmotic stress) in food products are not favorable to the growth rate of LAB. The relationship between water content and water activity (aw) is non-linear and depends on the interaction of food components with water. The lower limit of aw for bacterial growth is about 0.91 [16]. The effective ion concentration in cytoplasm is defined as the ionic strength. With the activity of specific ion channels and solute transporters, it maintains the cell volume. Another factor is macromolecule crowding. Bacterial proteins along with ribosomal RNA are space-consuming and therefore reduce cell volume [69]. When cells are under environmental osmotic stress osmoregulatory transporters are activated and ionic concentration changes as well as ionic strength [70,71]. Dehydration and the associated osmotic stress lead to a decrease in the bacterial cell volume, an increase in ionic strength and crowding, and a decrease in cell turgor.
In response to osmotic stress, various protective mechanisms are activated to maintain or restore internal physicochemical conditions [70]. The primary response of LAB to extracellular hyperosmotic stress is the absorption of potassium which, according to Sleator and Hill, is the signal for intracellular accumulation of specific osmoprotective solutes [72]. The secondary response is the synthesis and/or uptake of compounds called osmoprotectants, such as glycine betaine, glycine, choline, proline, carnitine, dimethylsulfonioacetate, and some amino acids. They retain water in the cell, maintaining intracellular osmotic balance and turgor pressure, cell volume, and electrolyte concentration, as well as stabilizing the enzyme function [72]. Due to the limited biosynthesis capacity, osmoprotectants are taken up through the cell membrane from the environment, contrary to the gradient [40,72]. Organic osmolytes are classified into three chemical categories: zwitterionic solutes (free polar amino acids: betaine, ecotine, hydroxyecotine), non-charged solutes (carbohydrates, carboxamine, acetylated neutral glutamine dipeptide), and anionic solutes (β-glutamate, β-hydroxybutyrates, negatively charged carbohydrates) [73]. In the case of hypoosmotic shock, osmoprotectants are rapidly removed from the cell [32].
Matrices with high aw, e.g., fruit juices, promote the overall growth of microorganisms, including undesirable putrefactive bacteria and competing microorganisms. On the other hand, in matrices with low water content, e.g., dry fermented meat, LAB cells are damaged and their number decreases [14]. A particular challenge for LAB cells is freeze drying, which is used, among others, for the production of starter cultures. In freeze drying, as a result of the sudden removal of extracellular water, there are: an increase in the ratio of cell surface area to cell volume, loss of pressure, deformation of the cell membrane, and decrease in cell turgor. These changes can eventually lead to cell death [74]. Increased contact of the cell membrane with the surrounding air also poses an increased risk of damage by the reactive oxygen species. The effect of induced osmotic stress may be a decrease in growth rate, a decrease in metabolic activity, and finally increased cell mortality during irrigation [35]. In the study of Teixeira et al. a non-linear relationship was found in the viability of spray-dried strains of Lactobacillus delbrueckii ssp. bulgaricus with water activity, showing the highest survival rates at medium and low aw values in the range of 0.11 to 0.23 [75]. The cell damage is related to the lack of osmotic equilibrium with the extracellular medium. The adaptation of LAB to osmolality and food matrix changes is reflected by the appropriate cell turgor. However, the measurement of turgor pressure is difficult, which makes it difficult to assess the effect of the osmotic pressure on cells.
6. Additives
Food ingredients, including additives, can positively or negatively affect the lifespan of LAB. Sweeteners, salts, aromatic compounds, flavors, colorants, and preservatives can affect both the growth and viability of LAB. In the study of Vinderola et al. the growth of 24 strains of lactic acid starter bacteria in liquid media containing different substances, such as salts (NaCl and KCl), sugars (sucrose and lactose), sweeteners (acesulfame and aspartame), aroma compounds (diacetyl, acetaldehyde, and acetoin), antibiotics (nisin, natamycin), and commercially available, natural colorings, flavoring agents, flavoring–coloring agents, and preservatives (lysozyme), was assessed. Most of these additives at the concentrations used for industrial manufacturing did not influence the LAB growth. Commercially available additives like flavoring–coloring agents (strawberry, vanilla, and peach) were shown to inhibit the growth of LAB. This effect was strain-dependent, e.g., complete growth inhibition was observed for L. acidophilus A3 and A9 [76].
In the case of meat products, including fermented ones, the influence of curing agents, such as nitrate and/or nitrite with salt, on LAB ranges from no impact and growth delay to complete growth inhibition. The membrane permeability of ions is regulated by membrane Na+/H+ pH-dependent antiporters and some proteins maintaining the intracellular ion balance and osmotic pressure [77,78]. The acclimatization to salt stress is a complex mechanism. In the study of Li et al. on L. plantarum it was found that under salt stress the cell wall composition alters by upregulated synthesis of proteins responsible for amino acid metabolism (metS, GshAB, GshR3, PepN, GshR4, and serA) and, consecutively, carbohydrate metabolism (I526_2330, Gpd, and Gnd) and peptidoglycan hydrolysis (N876_0118940) [79]. A fermented meat environment containing NaCl is harmful to many undesirable microorganisms, while the growth of LAB is strain-dependent and in the case of salt-tolerant strains is promoted [80]. In the study of Łaszkiewicz et al. poultry meat cured with sodium nitrite did not have a negative effect on Levilactobacillus brevis KL5 [81]. In the study of Gandhi et al. L. casei was found to have better salt resistance than L. acidophilus and B. longum and for food products containing up to 5% NaCl [82]. Arihara et al. studied sensitivity to sodium chloride and sodium nitrite in L. gasseri. Using UV irradiation they generated salt-tolerant mutant strains and demonstrated satisfactory growth of mutant strain L. gasseri 1131-M8 in meat containing 3.3% sodium chloride and 200 ppm sodium nitrite [83].
Among the factors supporting the viability of LAB, we can distinguish those that stimulate the growth of bacteria (e.g., sugars, amino acids, fatty acids, whey protein) and cytoprotective agents, including oxygen scavengers or antioxidants (e.g., inulin, oligofructose, glycerol sorbitol, whey protein, vitamins, and other antioxidants) and fats, e.g., dihydrosterculic acid [11,50]. In this context, supplementing the food matrix with prebiotics that promote LAB growth seems to be of significant importance [84].
To reduce the adverse impact of oxygen on the lifespan of LAB, additives acting as absorbers or antioxidants can be successfully used. Such substances increase the survival rate of LAB in food products, as exemplified by the results of the L. acidophilus survival study in yogurt with the addition of ascorbic acid (vitamin C). In the study of Ghandi et al. the presence of ascorbic acid was found to improve both the survival and the maintenance of fermentation activity of the dried bacterial strains during storage [85]. Antioxidants naturally present in nutritional matrices, e.g., in fruit juices and pulps, significantly increase the survival rate of LAB, but the effect is strain-dependent. An example is the study by Senadira et al. which found an increased survival rate of added B. animalis ssp. lactis BB-12 in yogurts with added fruit pulps compared to control hs, whereas the differences of counts of yogurt bacteria Streptococcus thermophiles and Lactobacillus delbrueckii ssp. bulgaricus were insignificant [86].
7. Methods to Reduce the Occurrence and Impact of Stress Factors on LAB
Adding LAB into the food matrix is a technological and microbiological challenge due to unfavorable and even extreme environmental conditions during food manufacturing and storage. To reduce the impact of stress factors on LAB, various methods of protecting LAB colonies have been introduced, such as edible coatings and microencapsulation. Additional protection may be provided by the addition of cytoprotective agents, including those with thermo- or cryoprotective properties, as well as oxygen scavengers or antioxidants. Protective measures against oxidative stress for LAB also include the use of packaging materials that inhibit the passage of atmospheric oxygen into the product [59,87]. To adjust optimal physicochemical parameters of food manufacturing, new food processing techniques are being introduced in the food industry, such as non-thermal techniques like high hydrostatic process (HHP), high-pressure carbon dioxide, radiation treatments, pulsed electric field (PEF), ultrasonic (US), irradiation, hydrodynamic cavitation, and cold plasma (CP) [88,89]. HHP is a promising method of inactivation of pathogenic microbes in a wide range of foods (meat, dairy, fruits) but technical problems limit its use [90]. PEF is mostly used in the preservation of liquid foods. This method reduces microbial load and inhibits spoilage enzyme activity, increasing the shelf-life of food products [91]. US is an emerging technology that is applied to reduce the microbial load in food products. Although it has many applications in food processing, further research is needed to implement this method in food preservation [92]. Gamma irradiation is an effective technique for food preservation. It provides good results in food preservation but technological obstacles as well as a lack of consumer acceptance limit the use of this method [93]. CP is an effective method of decontamination of microbial load in food. It has a great potential and can be applied in various products [94]. These methods offer food preservation and safety at near-ambient temperature. This has a positive effect on the viability of probiotic bacteria, nutritional quality, and food sensory attributes.
New technologies offer biodegradable and edible biopolymers with antimicrobial properties such as chitosan or polylactic acid (PLA). Natural biopolymers have many properties, including antimicrobial effect, mechanical strength, biodegradability, and low toxicity. They are also suitable for bacteriocins (nisin, pediosin, lactisin) antioxidants, antimicrobial compounds (polyphenols, tannins, flavonoids, gallic acid, silver nanoparticles), and ROS scavenger incorporation [95]. This allows keeping the food environment optimal for LAB. This combined antimicrobial packaging technology allows milder processing of packaged food, limiting physicochemical stress.
Entrapment of probiotic cells in edible coatings and films is a promising approach of providing LAB to food that can overcome the harmful influence of physicochemical stresses associated with food processing. The materials used for edible film/coating production are biopolymers, proteins, lipids, or composites [96]. The wide application of edible coatings in the food industry is limited by technological problems but innovative nanotechnology can bring new possibilities [97].
Another method of prevention against environmental stressors of LAB is encapsulation. This technique may improve the survival of LAB in food subjected to adverse conditions. Nowadays, different encapsulation techniques, such as extrusion, emulsion, spray drying, spray chilling, and fluidization, are available [98]. Encapsulation of probiotic strains is successfully used in dairy food products but there is a growing number of reports of meat products [99].
Various microorganisms including LAB present in the nutritional matrix exhibit interactions that can be neutral, positive, or even harmful [100,101]. Symbiotic relationships of LAB have been observed in dairy products for many years. Pete and Lokema described two kinds of interaction between L. bulgaricus and S. thermophilus: stimulation of growth, followed by growth inhibition caused by excessive production of lactic acid by L. bulgaricus [102]. According to Jankov and Stoyanov the interactions between L. bulgaricus and S. thermophilus depend on the ratios and bacterial strains [103]. Another example of microbial interaction is the synergistic growth-stimulating effect between strains of Lactobacillus acidophilus and Bifidobacterium bifidum [104]. Another example of a synergistic effect was reported by Ge et al. in the study of S. thermophilus and L. bulgaricus during the fermentation and storage of yogurt. The S. thermophilus CICC 6038 quickly metabolizes lactose and produces acids, creating the acidic environment to which L. bulgaricus is better adapted. Therefore, L. bulgaricus dominates and has a higher survival rate at the end of storage in cocultured yogurt [105]. The synthesis of exopolysaccharides by S. thermophilus is an additional mechanism which makes the strain more resistant to harsh conditions [106]. This indicates that the right selection of strains and their combination are important for optimizing the viability of LAB in the nutritional matrix.
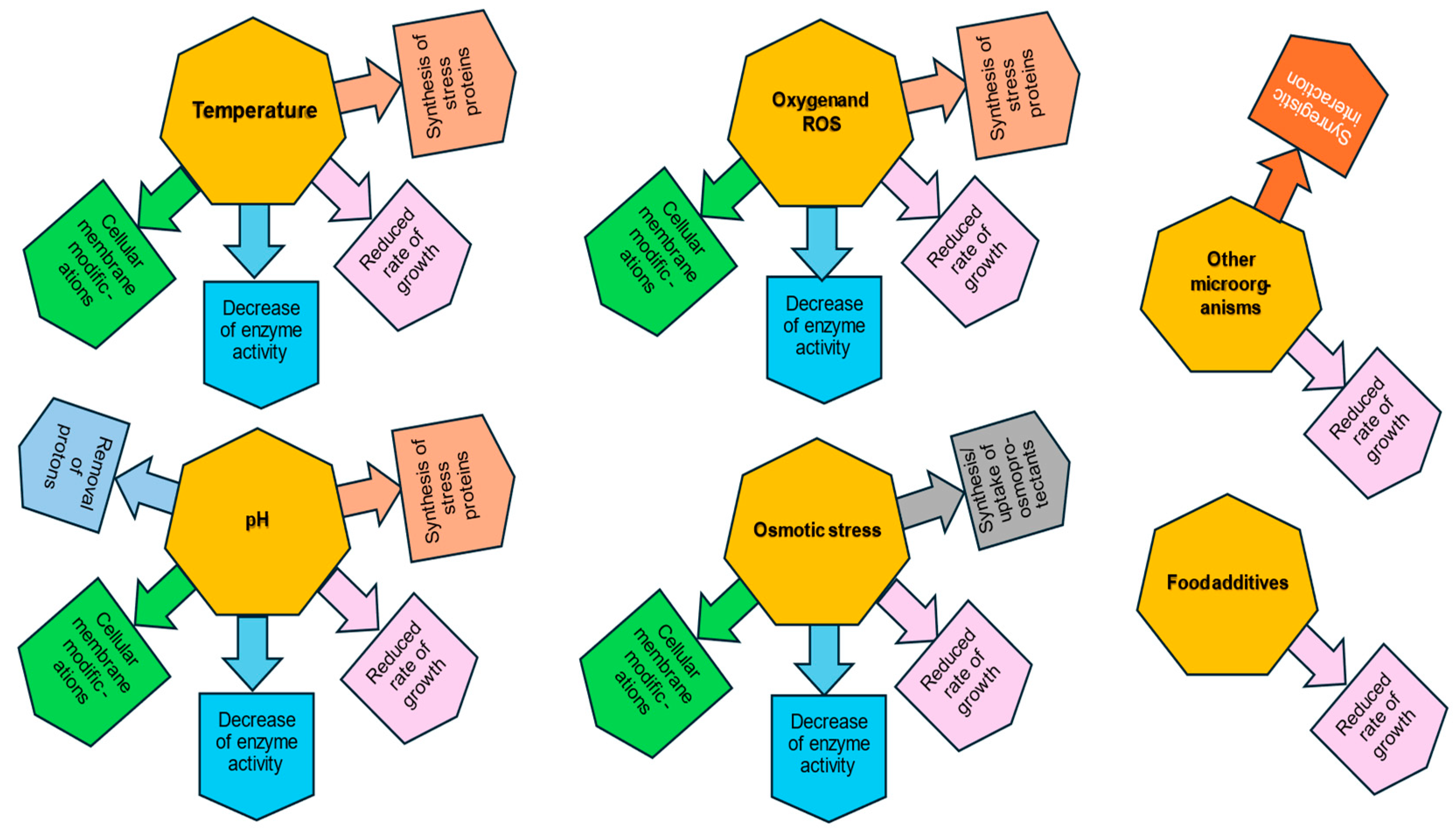
An important issue leading to the reduction of the adverse impact of stress factors on LAB is the phenomenon of stress adaptation (Figure 2). Upon an extended exposure to sublethal stress inducing a long milder adaptation under stationary conditions, mutants of high permanent resistance (and of increased virulence in the case of pathogens) may arise, unlike the gene expression of defensive mechanisms in the exponential phase temporarily, which does not result in mutations. Therefore, LAB adapt more progressively and thus much better to lower pH and increasing acid concentration during their exponential growth in a fermenting food product (e.g., cheese) than when they are suddenly exposed to any acidic treatment (intervention) in the food (dairy) industry. Stress adaptation consists of increasing resistance to a specific stress by applying moderate stress beforehand. In this way, the cells develop protection against the next, more intense stress [30]. The stress resistance following adaptation is variable and bacterial strain-dependent [107]. Increased tolerance to one stress factor also contributes to better tolerance of other types of stress, which is referred to as “cross-protection” [108]. Stress-induced expression of certain cell genes determines effective acclimatization to new environmental conditions. In the study by Maus and Ingham, exposure to moderate stress repeatedly applied to cultures of B. longum and B. lactis resulted in an increase in tolerance to cold and low pH [109]. Chen et al. demonstrated the cross-protection adaptation of L. kefiranofaciens. The acclimatization to various sublethal environmental stresses (37 and 52 °C, 20 and −20 °C, pH 5.0 and 3.0, and salt 0.05 and 0.2%) resulted in enhanced resistance with improved viability of adapted cells compared with non-adapted cells [108]. In another study Ma et al. showed that the preadaptation of L. plantarum KLDS 1.0628 to sublethal levels of heat, oxidative stress, acid, bile salt, and osmotic stress decreased the SFA/USFA ratio in the cell membrane which resulted in cross-protection against multiple stresses [110]. According to the different steps of food manufacturing and processes, Fonseca et al. distinguish two types of stress adaptive response. The first type, active at the time of food processing, involves modification of the lipid composition of the bacterial membrane, changes in proteins, and regulation of gene expression, which helps bacteria to respond to stress occurring during the first stages of production. The second type during stabilization and food storage is a passive reaction associated with subsequent stabilization processes (membrane phase transition, lipid oxidation, protein denaturation, modification of ionic strength, and viscosity) [35].
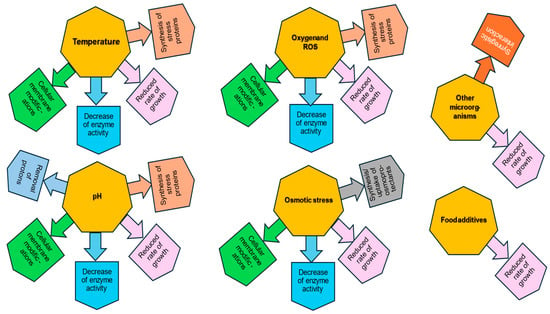
Figure 2.
Lactic acid bacteria stress response and adaptation to different types of environmental stressors. According to Wang and Zhong, 2024 [26], Fiocco et al., 2020 [30], de Angelis and Gobbetti, 2004 [32], Papadimitriou et al., 2016 [40], Filho et al., 2017 [50], Wu et al., 2014 [51], Talwalkar and Kailasapathy, 2004 [65], Nguyen et al., 2024 [66], Wang et al., Homayouni et al., 2008 [68], Sleator and Hill, 2001 [72], Vinderola et al., 2002 [76], Maus and Ingham, 2003 [109].
Genome sequencing technologies enable fast and precise physiological bacterial strain characterization and allow the assessment of niche/matrix-specific stress adaptation [111]. In the study of Bachman et al. the adaptation of an L. lactis strain was investigated by propagating it for 1000 generations in milk. Genome and phenotypic analyses revealed major changes associated with improved growth in milk [112]. The traditional process of evolutionary selection of best-fitting bacterial strains to environmental stress nowadays can be implemented by rational strain modification methods of genetic engineering and synthetic biology. Therefore, LAB strains with recombinant DNA and improved characteristics and performance adjusted for food applications could be available. However, regulatory issues along with consumers’ acceptance should be considered [113].
8. Conclusions and Future Trends
The environment of the food matrix as well as external factors during processing and storage, such as temperature, relative humidity, gaseous environment, or the presence of competing microorganisms, can support or reduce the lifespan of LAB. The physicochemical conditions are interrelated and parallelly influence LAB. To adjust to different environmental changes, LAB are equipped with defense and adaptation mechanisms which are in many cases strain-specific features. Therefore, to support or maintain LAB survival rates in related food products, it is of great value to limit the impact of various stresses by creating optimal physicochemical conditions during manufacturing and storage of food products. On the other hand, understanding LAB stress response and adaptation mechanisms is of great practical significance for choosing appropriate suitable LAB strains as well as for artisanal or large-scale industrial production.
During recent years, several attempts have been made to achieve better survival of LAB in food products. Among them, the most promising are the encapsulation method and the addition of different protectants. Nowadays, there is a growing interest in minimally processed foods with fewer or no added preservatives. Therefore, to ensure microbiological food safety new methods of food processing have been introduced, including high-pressure carbon dioxide, pulsed electric fields, ultrasound, irradiation, or cold plasma. While new food processing operations are promising, they create new combinations of environmental stress with unknown impacts on LAB growth rate. Moreover, emerging technologies have to overcome some barriers for implementation in the food industry, including high costs and consumers’ acceptance.
In the context of the interaction of various physicochemical factors, the key issue is how to optimize food manufacturing to achieve food products containing beneficial LAB strains of exceptional quality and safety. Despite the advances, further studies are needed to better understand the impact of specific factors and their combinations on LAB viability.
Author Contributions
B.S.: conceptualization, methodology, software, validation, formal analysis, resources, data curation, writing—original draft preparation, writing—review and editing, A.S.: investigation, writing—review and editing, M.T.: investigation, writing—review and editing, D.K.-K.: visualization, supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- El-Gendy, S.M.; Shaker, M. Fermented foods of Egypt and the Middle East. J. Food Prot. 1983, 46, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Tamang, J.P. Diversity of fermented foods. In Fermented Foods and Beverages of the World, 1st ed.; Tamang, J.P., Kailasapathy, K., Eds.; CRC Press: New York, NY, USA, 2010; pp. 41–84. [Google Scholar]
- Sionek, B.; Szydłowska, A.; Küçükgöz, K.; Kołożyn-Krajewska, D. Traditional and New Microorganisms in Lactic Acid Fermentation of Food. Fermentation 2023, 9, 1019. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.; Mattarell, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food. Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- Zheng, J.; Ruan, L.; Sun, M.; Gänzle, M.G. A genomic view of Lactobacilli and Pediococci demonstrates that phylogeny matches ecology and physiology. Appl. Environ. Microbiol. 2015, 81, 7233–7243. [Google Scholar] [CrossRef] [PubMed]
- Duar, R.M.; Lin, X.B.; Zheng, J.; Martino, M.E.; Grenier, T.; Pérez-Muñoz, M.E.; Leulier, F.; Gänzle, M.; Walter, J. Lifestyles in transition: Evolution and natural history of the genus Lactobacillus. FEMS Microbiol. Rev. 2017, 41, 27–48. [Google Scholar] [CrossRef] [PubMed]
- Generally Recognized as Safe (GRAS)|FDA. Available online: https://www.fda.gov/food/food-ingredients-packaging/generally-recognized-safe-gras (accessed on 20 April 2024).
- Qualified Presumption of Safety (QPS)|EFSA. Available online: https://efsa.europa.eu/en/topic/qualified-presumption-safety-qps (accessed on 20 April 2024).
- Booth, I.R. Stress and the single cell: Intrapopulation diversity is a mechanism to ensure survival upon exposure to stress. Int. J. Food Microbiol. 2002, 78, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Terpou, A.; Papadaki, A.; Lappa, I.K.; Kachrimanidou, V.; Bosnea, L.A.; Kopsahelis, N. Probiotics in Food Systems: Significance and Emerging Strategies towards Improved Viability and Delivery of Enhanced Beneficial Value. Nutrients 2019, 11, 1591. [Google Scholar] [CrossRef]
- Hossain, M.I.; Sadekuzzaman, M.; Ha, S.D. Probiotics as potential alternative biocontrol agents in the agriculture and food industries: A review. Food Res. Int. 2017, 100, 63–73. [Google Scholar] [CrossRef]
- Mbye, M.; Baig, M.A.; AbuQamar, S.F.; El-Tarabily, K.A.; Obaid, R.S.; Osaili, T.M.; Al-Nabulsi, A.A.; Turner, M.S.; Shah, N.P.; Ayyash, M.M. Updates on understanding of probiotic lactic acid bacteria responses to environmental stresses and highlights on proteomic analyses. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1110–1124. [Google Scholar] [CrossRef]
- Gomand, F.; Borges, F.; Burgain, J.; Guerin, J.; Revol-Junelles, A.M.; Gaiani, C. Food matrix design for effective lactic acid bacteria delivery. Ann. Rev. Food Sci. Technol. 2019, 10, 285–310. [Google Scholar] [CrossRef] [PubMed]
- Kathiriya, M.R.; Vekariya, Y.V.; Hati, S. Understanding the Probiotic Bacterial Responses Against Various Stresses in Food Matrix and Gastrointestinal Tract: A Review. Probiotics Antimicrob. Proteins 2023, 15, 1032–1048. [Google Scholar] [CrossRef] [PubMed]
- Lima, V.; Pinto, C.A.; Saraiva, J.A. The dependence of microbial inactivation by emergent nonthermal processing technologies on pH and water activity. Innov. Food Sci. Emerg. Technol. 2023, 89, 103460. [Google Scholar] [CrossRef]
- Brauer, A.M.; Shi, H.; Levin, P.A.; Huang, K.C. Physiological and regulatory convergence between osmotic and nutrient stress responses in microbes. Curr. Opin. Cell Biol. 2023, 81, 102170. [Google Scholar] [CrossRef] [PubMed]
- Sazykin, I.S.; Sazykina, M.A. The role of oxidative stress in genome destabilization and adaptive evolution of bacteria. Gene 2023, 857, 147170. [Google Scholar] [CrossRef] [PubMed]
- Rees, C.E.D.; Dodd, C.E.R.; Gibson, P.T.; Booth, I.R.; Stewart, G.S.A.B. The significance of bacteria in stationary phase to food microbiology. Int. J. Food Microbiol. 1995, 28, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Davis, C. Enumeration of probiotic strains: Review of culture-dependent and alternative techniques to quantify viable bacteria. J. Microbiol. Method. 2014, 103, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Ozogul, F.; Yazgan, H.; Ozogul, Y. Lactic acid bacteria: Lactobacillus acidophilus. In Encyclopedia of Dairy Sciences, 3rd ed.; Mc Sweeney, P.L., McNamara, J.P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 187–197. [Google Scholar]
- Oberg, T.S.; McMahon, D.J.; Culumber, M.D.; McAuliffe, O.; Oberg, C.J. Invited review: Review of taxonomic changes in dairy-related lactobacilli. J. Dairy Sci. 2022, 105, 2750–2770. [Google Scholar] [CrossRef]
- Delcour, J.; Ferain, T.; Hols, P. Advances in the genetics of thermophilic lactic acid bacteria. Curr. Opin. Biotechnol. 2000, 11, 497–504. [Google Scholar] [CrossRef]
- Song, S.; Bae, D.-W.; Lim, K.; Griffiths, M.W.; Oh, S. Cold stress improves the ability of Lactobacillus plantarum L67 to survive freezing. Intern. J. Food Microbiol. 2014, 191, 135–143. [Google Scholar] [CrossRef]
- Neffe-Skocinska, K.; Gierejkiewicz, M.; Kołozyn-Krajewska, D. Optimization of fermentation conditions for dry-aged sirloins with probiotic bacteria added. Food Sci. Technol. Qual. Zywn. Nauk. Technol. Jaskosc 2011, 6, 36–46. [Google Scholar] [CrossRef]
- Wang, A.; Zhong, Q. Drying of probiotics to enhance the viability during preparation, storage, food application, and digestion. A review. Compr. Rev. Food Sci. Food Saf. 2024, 23, e13287. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Haslbeck, M.; Buchner, J. The Heat Shock Response: Life on the Verge of Death. Mol. Cell 2010, 40, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Haq, S.F.; Samant, S.; Sukumaran, S. Adaptation of Lactobacillus acidophilus to thermal stress yields a thermotolerant variant which also exhibits improved survival at pH 2. Probiot. Antimicrob. Proteins 2018, 10, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Ananta, E.; Knorr, D. Evidence on the Role of Protein Biosynthesis in the Induction of Heat Tolerance of Lactobacillus Rhamnosus GG by Pressure Pre-Treatment. Int. J. Food Microbiol. 2004, 96, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Fiocco, D.; Longo, A.; Arena, M.P.; Russo, P.; Spano, G.; Capozzi, V. How probiotics face food stress: They get by with a little help. Crit. Rev. Food Sci. Nutr. 2020, 60, 1552–1580. [Google Scholar] [CrossRef] [PubMed]
- Broeckx, G.; Kiekens, S.; Jokicevic, K.; Byl, E.; Henkens, T.; Vandenheuvel, D.; Claes, I.; Lebeer, S.; Kiekens, F. Heat-pretreated Lactobacillus rhamnosus GG shows enhanced survival capacity after spray drying. Dry. Technol. 2022, 40, 3602–3613. [Google Scholar] [CrossRef]
- de Angelis, M.; Gobbetti, M. Environmental stress responses in Lactobacillus. A review. Proteomics 2004, 4, 106–122. [Google Scholar] [CrossRef]
- Gaucher, F.; Bonnassie, S.; Rabah, H.; Marchand, P.; Blanc, P.; Jeantet, R.; Jan, G. Review: Adaptation of Beneficial Propionibacteria, Lactobacilli, and Bifidobacteria Improves Tolerance Toward Technological and Digestive Stresses. Front. Microbiol. 2019, 10, 841. [Google Scholar] [CrossRef]
- Carvalho, A.S.; Silva, J.; Ho, P.; Teixeira, P.; Malcata, F.X.; Gibbs, P. Relevant factors for the preparation of freeze-dried lactic acid bacteria. Int. Dairy J. 2004, 14, 835–847. [Google Scholar] [CrossRef]
- Fonseca, F.; Pénicaud, C.; Tymczyszyn, E.E.; Gómez-Zavaglia, A.; Passot, S. Factors influencing the membrane fluidity and the impact on production of lactic acid bacteria starters. A review. Appl. Microbiol. Biotech. 2019, 103, 6867–6883. [Google Scholar] [CrossRef] [PubMed]
- Barria, C.; Malecki, M.; Arraiano, C.M. Bacterial adaptation to cold. Microbiology 2013, 159, 2437–2443. [Google Scholar] [CrossRef] [PubMed]
- Haddaji, N.; Mahdhi, A.K.; Ismaiil, M.B.; Bakhrouf, A. Effect of environmental stress on cell surface and membrane fatty acids of Lactobacillus plantarum. Arch. Microbiol. 2017, 199, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Delettre, J.; Guillot, A.; Corrieu, G.; Béal, C. Influence of cooling temperature and duration on cold adaptation of Lactobacillus acidophilus RD758. Cryobiology 2005, 50, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Ma, Y.; Zheng, Y.; Zhao, W.; Zhao, X.; Luo, T.; Zhang, J.; Yang, Z. Cold-Stress Response of Probiotic Lactobacillus plantarum K25 by iTRAQ Proteomic Analysis. J. Microbiol. Biotechnol. 2020, 30, 187–195. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Alegría, Á.; Bron, P.A.; de Angelis, M.; Gobbetti, M.; Kleerebezem, M.; Lemos, J.A.; Linares, D.M.; Ross, P.; Stanton, C.; et al. Stress Physiology of Lactic Acid Bacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 837–890. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, M.K.; Giri, S.K. Probiotic functional foods: Survival of probiotics during processing and storage. J. Fun. Foods 2014, 9, 225–241. [Google Scholar] [CrossRef]
- Gao, X.; Kong, J.; Zhu, H.; Mao, B.; Zhao, J. Lactobacillus, Bifidobacterium and Lactococcus response to environmental stress: Mechanisms and application of cross-protection to improve resistance against freeze-drying. J. Appl. Microbiol. 2021, 132, 802–821. [Google Scholar] [CrossRef]
- Sheehan, V.M.; Ross, P.G.; Fitzgerald, F. Assessing the acid tolerance and the technological robustness of probiotic cultures for fortification in fruit juices. Innov. Food Sci. Emerg. Technol. 2007, 8, 279–284. [Google Scholar] [CrossRef]
- Schumacher, K.; Brameyer, S.; Jung, K. Bacterial acid stress response: From cellular changes to antibiotic tolerance and phenotypic heterogeneity. Curr. Opin. Microbiol. 2023, 75, 102367. [Google Scholar] [CrossRef] [PubMed]
- Guan, N.Z.; Liu, L. Microbial response to acid stress: Mechanisms and applications. Appl. Microbiol. Biotechnol. 2020, 104, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Cotter, P.D.; Hill, C. Surviving the Acid Test: Responses of Gram-Positive Bacteria to Low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y. F1F0-ATPase functions under markedly acidic conditions in bacteria. In Regulation of Ca2+-ATPases, V-ATPases and F-ATPases; Chacraborti, S., Dhalla, N.S., Eds.; Adavances in Biochemistry in Health and Desease; Springer: Cham, Switzerland, 2016; pp. 459–468. [Google Scholar]
- Mendonça, A.A.; de Paula Pinto-Neto, W.; da Paixão, G.A.; Santos, D.d.S.; De Morais, M.A., Jr.; De Souza, R.B. Journey of the Probiotic Bacteria: Survival of the Fittest. Microorganisms 2023, 11, 95. [Google Scholar] [CrossRef] [PubMed]
- de Mesquita, A.R.; da Mota Silveira, L.P.; da Cruz Filho, I.J.; de Lima, V.F.; da Mota Silveira Filho, V.; Araujo, A.A.; da Silva, T.L.; de Freitas Araújo, K.; da Silva Macedo, L. Metabolism and physiology of Lactobacilli: A review. J. Environ. Anal. Progress 2017, 2, 125–136. [Google Scholar] [CrossRef]
- Wu, C.; He, G.; Zhang, J. Physiological and proteomic analysis of Lactobacillus case in response to acid adaptation. J. Ind. Microbiol. Biotechnol. 2014, 41, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Pan, M.; Wan, C.; Shah, N.P.; Tao, X.; Wei, H. Physiological and transcriptional responses and cross protection of Lactobacillus plantarum ZDY2013 under acid stress. J. Dairy Sci. 2016, 99, 1002–1010. [Google Scholar] [CrossRef]
- Lorca, G.L.; Raya, R.R.; Taranto, M.P.; De Valdez, G.F. Characterization of the protein-synthesis dependent adaptive acid tolerance response in Lactobacillus acidophilus. J. Mol. Microbiol. Biotechnol. 2002, 4, 525–532. [Google Scholar] [PubMed]
- Wu, C.; Zhang, J.; Wang, M.; Du, G.; Chen, J. Lactobacillus casei combats acid stress by maintaining cell membrane functionality. J. Ind. Microbiol. Biotech. 2012, 39, 1031–1039. [Google Scholar] [CrossRef]
- Ishikawa, M.; Kodama, K.; Yasuda, H.; Okamoto-Kainuma, A.; Koizumi, K.; Yamasato, K. Presence of halophilic and alkaliphilic lactic acid bacteria in various cheeses. Lett. Appl Microbiol. 2006, 44, 308–313. [Google Scholar] [CrossRef]
- Sánchez, A.-H.; Rejano, L.; Montaño, A.; de Castro, A. Utilization at high pH of starter cultures of lactobacilli for Spanish-style green olive fermentation. Int. J. Food Microbiol. 2001, 67, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Sawatari, Y.; Sugiyama, H.; Suzuki, Y.; Hanaoka, A.; Saito, K.; Yamauchi, H.; Okada, S.; Yokota, A. Development of fermented instant Chinese noodle using Lactobacillus plantarum. Food Microbiol. 2005, 22, 539–546. [Google Scholar] [CrossRef]
- Sawatari, Y.; Yokota, A. Diversity and mechanisms of alkali tolerance in lactobacilli. Appl. Environ. Microbiol. 2007, 73, 3909–3915. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Li, X.; Zhang, G.; Sadiq, F.A.; Simal-Gandara, J.; Xiao, J.; Sang, Y. Probiotics in the dairy industry-Advances and opportunities. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3937–3982. [Google Scholar] [CrossRef]
- Zotta, T.; Parente, E.; Ricciardi, A. Aerobic metabolism in the genus Lactobacillus: Impact on stress response and potential applications in the food industry. J. App. Microbiol. 2017, 122, 857–869. [Google Scholar] [CrossRef]
- Feng, T.; Wang, J. Oxidative stress tolerance and antioxidant capacity of lactic acid bacteria as probiotic: A systematic review. Gut Microbes 2020, 12, 1801944. [Google Scholar] [CrossRef]
- Liu, Q.; Hang, X.; Liu, X.; Tan, J.; Li, D.; Yang, H. Cloning and heterologous expression of the manganese superoxide dismutase gene from Lactobacillus casei Lc18. Ann. Microbiol. 2012, 62, 129–137. [Google Scholar] [CrossRef]
- Quatravaux, S.; Remize, F.; Bryckaert, E.; Colavizza, D.; Guzzo, J. Examination of Lactobacillus plantarum lactate metabolism side effects in relation to the modulation of aeration parameters. J. Appl. Microbiol. 2006, 101, 903–912. [Google Scholar] [CrossRef]
- Barre, O.; Mourlane, F.; Solioz, M. Copper induction of lactate oxidase of Lactococcus lactis: A novel metal stress response. J. Bacteriol. 2007, 189, 5947–5954. [Google Scholar] [CrossRef]
- Talwalkar, A.; Kailasapathy, K. Oxidative stress adaptation of probiotic bacteria. Milchwissenschaft 2004, 59, 140–143. [Google Scholar]
- Nguyen, A.V.; Yaghoobi, M.; Zhang, S.; Li, P.; Li, Q.; Dogan, B.; Ahnrud, G.P.; Flock, G.; Marek, P.; Simpson, K.W.; et al. Adaptive laboratory evolution of probiotics toward oxidative stress using a microfluidic-based platform. Small 2024. [Google Scholar] [CrossRef]
- Klu, Y.A.K.; Williams, J.H.; Phillips, R.D.; Chen, J. Survival of Lactobacillus rhamnosus GG as influenced by storage conditions and product matrixes. J. Food Sci. 2012, 77, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Homayouni, A.; Ehsani, M.R.; Azizi, A.; Razavi, S.H.; Yarmand, M.S. Growth and survival of some probiotic strains in simulated ice cream conditions. J. Appl. Sci. 2008, 8, 379–382. [Google Scholar] [CrossRef]
- Liu, B.; Hasrat, Z.; Poolman, B.; Boersma, A.J. Decreased effective macromolecular crowding in Escherichia coli adapted to hyperosmotic stress. J. Bacteriol. 2019, 201, e00708-18. [Google Scholar] [CrossRef]
- Poolman, B. Physicochemical homeostasis in bacteria. FEMS Microbiol Rev. 2023, 47, fuad033. [Google Scholar] [CrossRef] [PubMed]
- Syeda, R.; Qiu, Z.; Dubin, A.E.; Murthy, S.E.; Florendo, M.N.; Mason, D.E.; Mathur, J.; Cahalan, S.M.; Peters, E.C.; Montal, M.; et al. LRRC8 proteins form volume- regulated anion channels that sense ionic strength. Cell 2016, 164, 499. [Google Scholar] [CrossRef] [PubMed]
- Sleator, R.D.; Hill, C. Bacterial osmoadaptation: The role of osmolytes in bacterial stress and virulence. FEMS Microbiol. Rev. 2001, 26, 49–71. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [PubMed]
- Wendel, U. Assessing viability and stress tolerance of probiotics. A review. Front. Microbiol. 2022, 12, 818468. [Google Scholar] [CrossRef]
- Teixeira, P.C.; Castro, M.H.; Malcata, F.X.; Kirby, R.M. Survival of Lactobacillus delbrueckii ssp. bulgaricus following spray-drying. J. Dairy Sci. 1995, 78, 1025–1031. [Google Scholar] [CrossRef]
- Vinderola, C.G.; Costa, G.A.; Regenhardt, S.; Reinheimer, J.A. Influence of compounds associated with fermented dairy products on the growth of lactic acid starter and probiotic bacteria. Int. Dairy J. 2002, 12, 579–589. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Boekhorst, J.; Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef]
- Padan, E.; Bibi, E.; Ito, M.; Krulwich, T. Alkaline pH homeostasis in bacteria: New insights. Biochim. Biophys. Acta (BBA)-Biomembr. 2005, 1717, 67–88. [Google Scholar] [CrossRef]
- Li, M.; Wang, Q.; Song, X.; Guo, J.; Wu, J.; Wu, R. iTRAQ-based proteomic analysis of responses of Lactobacillus plantarum FS5-5 to salt tolerance. Ann. Microbiol. 2019, 69, 377–394. [Google Scholar] [CrossRef]
- Kołożyn-Krajewska, D.; Dolatowski, Z.J. Probiotic meat products and human nutrition. Process Biochem. 2012, 47, 1761–1772. [Google Scholar] [CrossRef]
- Łaszkiewicz, B.; Szymański, P.; Kołożyn-Krajewska, D. The effect of selected lactic acid bacterial strains on the technological and microbiological quality of mechanically separated poultry meat cured with a reduced amount of sodium nitrite. Poult. Sci. 2021, 100, 263–272. [Google Scholar] [CrossRef]
- Gandhi, A.; Shah, N.P. Effect of salt on cell viability and membrane integrity of Lactobacillus acidophilus, Lactobacillus casei and Bifidobacterium longum as observed by flow cytometry. Food Microbiol. 2015, 49, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Arihara, K.; Itoh, M. UV-induced Lactobacillus gasseri mutants resisting sodium chloride and sodium nitrite for meat fermentation. Int. J. Food Microbiol. 2000, 56, 227–230. [Google Scholar] [CrossRef] [PubMed]
- Champagne, C.P.; da Cruz, A.G.; Daga, M. Strategies to improve the functionality of probiotics in supplements and foods, current opinion. Food Sci. 2018, 22, 160–166. [Google Scholar]
- Ghandi, A.; Powell, I.B.; Broome, M.; Adhikari, B. Survival, fermentation activity and storage stability of spray dried Lactococcus lactis produced via different atomization regimes. J. Food Eng. 2013, 115, 83–90. [Google Scholar] [CrossRef]
- Senadira, S.S.; Prasanna, P.H.P.; Jayawardene, N.W.I.A.; Gunasekara, D.C.S.; Senadira, P.; Chandrasekara, A. Antioxidant, physicochemical, microbiological, and sensory properties of probiotic yoghurt incorporated with various Annona species pulp. Helium 2018, 11, E00955. [Google Scholar] [CrossRef] [PubMed]
- da Cruz, A.G.; Faria, J.D.A.F.; van Dender, A.G.F. Packaging System and Probiotic Dairy Foods. Food Res. Int. 2007, 40, 951–956. [Google Scholar] [CrossRef]
- Asaithambi, N.; Singh, S.K.; Singha, P. Current status of non-thermal processing of probiotic foods: A review. J. Food Eng. 2021, 303, 110567. [Google Scholar] [CrossRef]
- Almada-Érix, C.N.; Almada, C.N.; Souza Pedrosa, G.T.; Dos Santos, P.; Schmiele, M.; Clerici, M.T.P.S.; Martinez, J.; Lollo, P.C.; Magnani, M.; Sant’Ana, A.S. Quantifying the impact of eight unit operations on the survival of eight Bacillus strains with claimed probiotic properties. Food Res. Int. 2021, 142, 110–191. [Google Scholar] [CrossRef] [PubMed]
- Rendueles, E.; Omer, M.K.; Alvseike, O.; Alonso-Calleja, C.; Capita, R.; Prieto, M. Microbiological food safety assessment of high hydrostatic pressure processing: A review. LWT-Food Sci. Technol. 2011, 44, 1251–1260. [Google Scholar] [CrossRef]
- Niu, D.; Zeng, X.-A.; Ren, E.-F.; Xu, F.-Y.; Li, J.; Wang, M.-S.; Wang, R. Review of the application of pulsed electric fields (PEF) technology for food processing in China. Food Res. Int. 2020, 137, 109715. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, H.B.; Annapure, U.S.; Deshmukh, R.R. Non-thermal Technologies for Food Processing. Front. Nutr. 2021, 8, 657090. [Google Scholar] [CrossRef] [PubMed]
- Castell-Perez, M.E.; Moreira, R.G. Irradiation and consumers acceptance. In Innovative Food Processing Technologies; Knoerzer, K., Juliano, P., Smithers, G., Eds.; Elsevier: Cambridge, UK, 2021; pp. 122–135. [Google Scholar]
- Allai, F.M.; Azad, Z.A.; Mir, N.A.; Gul, K. Recent advances in non-thermal processing technologies for enhancing shelf life and improving food safety. Appl. Food Res. 2023, 3, 100258. [Google Scholar] [CrossRef]
- Gumienna, M.; Górna, B. Antimicrobial Food Packaging with Biodegradable Polymers and Bacteriocins. Molecules 2021, 26, 3735. [Google Scholar] [CrossRef]
- Galus, S.; Kadzińska, J. Food applications of emulsion-based edible films and coatings. Trends Food Sci. Technol. 2015, 45, 273–283. [Google Scholar] [CrossRef]
- Torres-Giner, S.; Perez-Masia, R.; Lagaron, J.M. A review on electrospun polymer nanostructures as advanced bioactive platforms. Polym. Eng. Sci. 2016, 56, 500–527. [Google Scholar] [CrossRef]
- Rodrigues, F.J.; Cedran, M.F.; Bicas, J.L.; Sato, H.H. Encapsulated probiotic cells: Relevant techniques, natural sources as encapsulating materials and food applications—A narrative review. Food Res. Int. 2020, 137, 109682. [Google Scholar] [CrossRef] [PubMed]
- Cavalheiro, C.P.; Ruiz-Capillas, C.; Herrero, A.M.; Jiménez-Colmenero, F.; Menezes, C.R.; Fries, L.L.M. Application of probiotic delivery systems in meat products. Trends Food Sci. Technol. 2015, 46, 120–131. [Google Scholar] [CrossRef]
- Yang, S.; Bai, M.; Kwok, L.Y.; Zhong, Z.; Sun, Z. The intricate symbiotic relationship between lactic acid bacterial starters in the milk fermentation ecosystem. Crit. Rev. Food Sci Nutr. 2023, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Aryana, K.J.; Olson, D.W. A 100-Year Review: Yogurt and other cultured dairy products. J Dairy Sci. 2017, 100, 9987–10013. [Google Scholar] [CrossRef] [PubMed]
- Pette, J.W.; Lolkema, H. Yoghurt. 1. Symbiosis and antibiosis in mixed cultures of Lactobacillus bulgaricus and Streptococcus thermophilus. Nederlandsch Melk-en Zuiveltijdschr. 1950, 4, 197–208. [Google Scholar]
- Jankov, J.; Stoyanov, I.V. Study on the thermophilic lactobacilli in milk for yoghurt production. In Proceedings of the XVII International Dairy Congress, Section F5, Munich, Germany, 25 August 1966; pp. 677–680. [Google Scholar]
- Kneifel, W.; Jaros, D.; Erhard, F. Microflora and acidification properties of yogurt and yogurt-related products fermented with commercially available starter cultures. Internat. J. Food Microbiol. 1993, 18, 179–189. [Google Scholar] [CrossRef]
- Ge, Y.; Yu, X.; Zhao, X.; Liu, C.; Li, T.; Mu, S.; Zhang, L.; Chen, Z.; Zhang, Z.; Song, Z.; et al. Fermentation characteristics and post-acidification of yogurt by Streptococcus thermophilus CICC 6038 and Lactobacillus delbrueckii ssp. Bulgaricus CICC 6047 at optimal inoculum ratio. J. Dairy Sci. 2023, 107, 123–140. [Google Scholar] [CrossRef] [PubMed]
- Taj, R.; Masud, T.; Sohail, A.; Sammi, S.; Naz, R.; Khanal, B.K.S.; Nawaz, M.A. In vitro screening of EPS-producing Streptococcus thermophilus strains for their probiotic potential from Dahi. Food Sci. Nutr. 2022, 10, 2347–2359. [Google Scholar] [CrossRef]
- Ricciardi, A.; Parente, E.; Guidone, A.; Ianniello, R.G.; Zotta, T.; Abu Sayem, S.M.; Varcamonti, M. Genotypic diversity of stress response in Lactobacillus plantarum, Lactobacillus paraplantarum and Lactobacillus pentosus. Int. J. Food Microbiol. 2012, 157, 278–285. [Google Scholar] [CrossRef]
- Chen, M.J.; Tang, H.Y.; Chiang, M.L. Effects of heat, cold, acid and bile salt adaptations on the stress tolerance and protein expression of kefir-isolated probiotic Lactobacillus kefiranofaciens M1. Food Microbiol. 2017, 66, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Maus, J.E.; Ingham, S. Employment of stressful conditions during culture production to enhance subsequent cold- and acid-tolerance of bifidobacteria. J. Appl. Microb. 2003, 95, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xu, C.; Liu, F.; Hou, J.; Shao, H.; Yu, W. Stress adaptation and cross-protection of Lactobacillus plantarum KLDS 1.0628. Cyta-J. Food 2021, 19, 72–80. [Google Scholar] [CrossRef]
- Bolotin, A.; Wincker, P.; Mauger, S.; Jaillon, O.; Malarme, K.; Weissenbach, J.; Ehrlih, S.D.; Sorokin, A. The complete genome sequence of the lactic acid bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 2001, 11, 731–753. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, H.; Starrenburg, M.J.C.; Molenaar, D.; Kleerebezem, M.; Vlieg, J. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution. Genome Res. 2012, 22, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hung, S.; Chan, J.; Chen, J.; Solem, C.; Jensen, P.R. Systems Biology—A Guide for Understanding and Developing Improved Strains of Lactic Acid Bacteria. Front. Microbiol. 2019, 10, 876. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).