
Improvement in Ethylene Glycol Bioconversion to Glycolic Acid by Gluconobacter oxydans Using Central Composite Experimental Design and Bioreactor Fed-Batch Strategies


 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microbial Growth and Maintenance
2.2. Selection of Culture Medium for Cell Propagation
2.3. Optimization of Cell Growth Medium
2.4. Bioconversion of Ethylene Glycol to Glycolic Acid in STR and Bubble Column Bioreactor
2.5. Analytical Methods
3. Results and Discussion
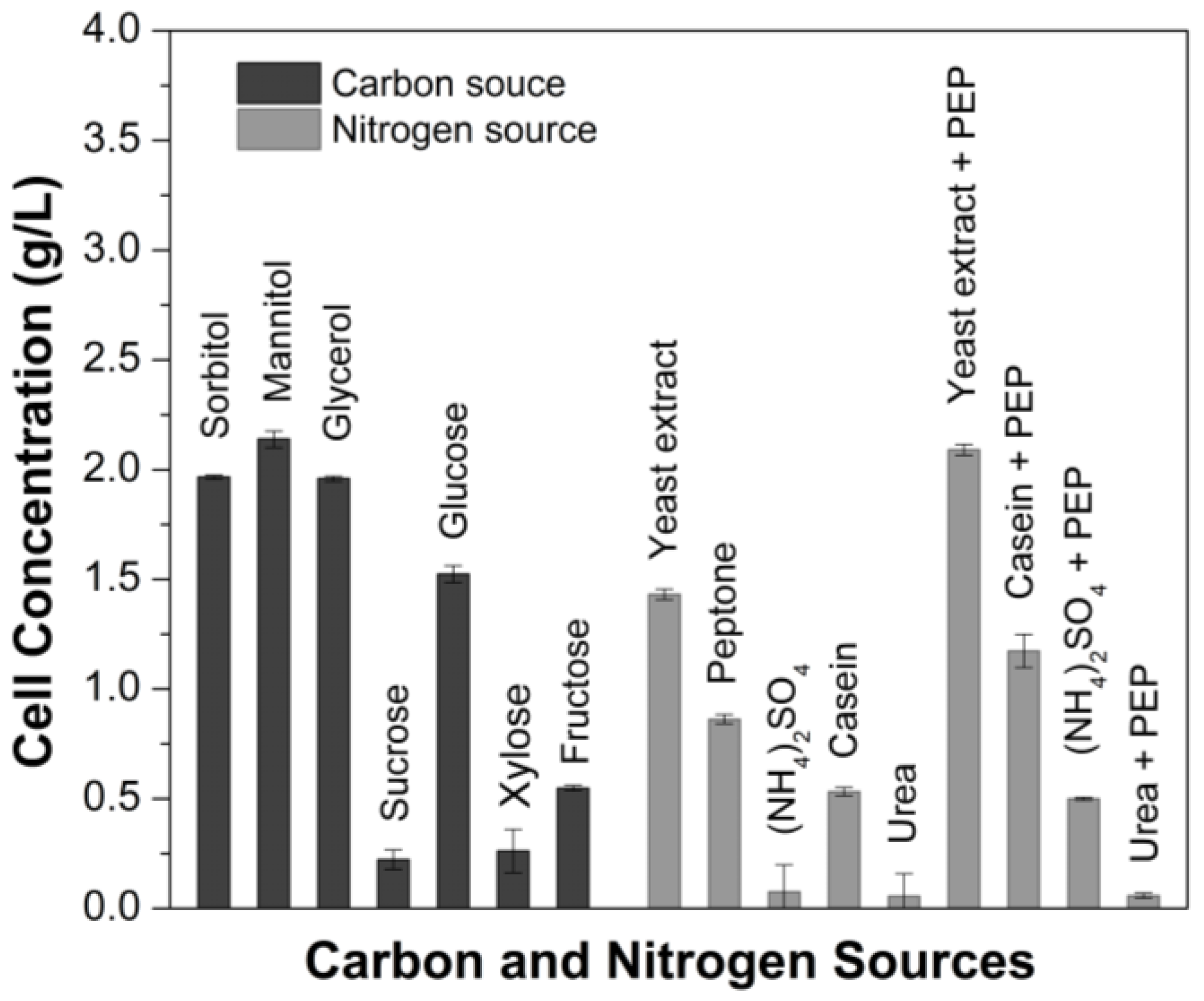
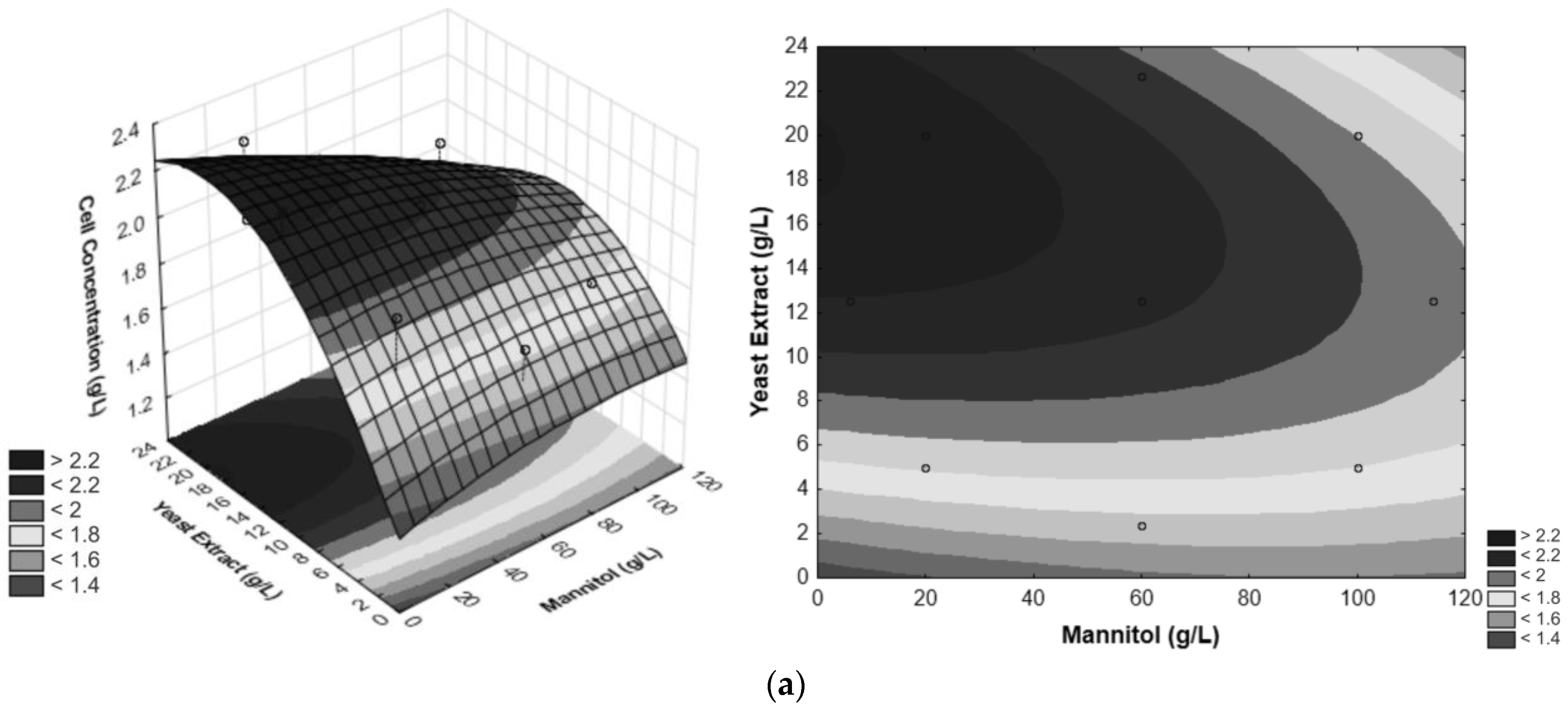
3.1. Optimization of the Growth Medium for G. oxydans
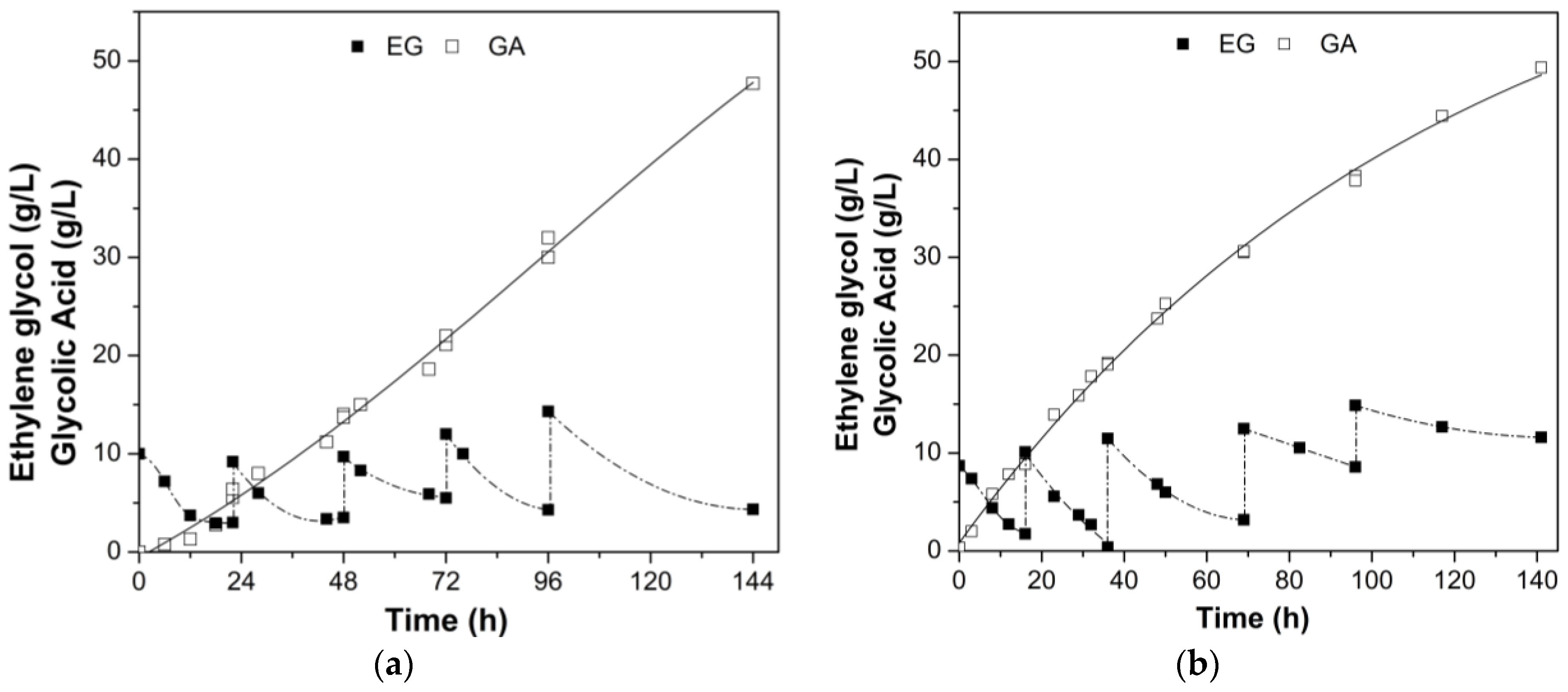
3.2. Fed-Batch Bioconversion of Ethylene Glycol to Glycolic Acid
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salusjärvi, L.; Havukainen, S.; Koivistoinen, O.; Toivari, M. Biotechnological production of glycolic acid and ethylene glycol: Current state and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 2525–2535. [Google Scholar] [CrossRef]
- Cabulong, R.B.; Lee, W.-K.; Bañares, A.B.; Ramos, K.R.M.; Nisola, G.M.; Valdehuesa, K.N.G.; Chung, W.-J. Engineering Escherichia coli for glycolic acid production from D-xylose through the Dahms pathway and glyoxylate bypass. Appl. Microbiol. Biotechnol. 2018, 102, 2179–2189. [Google Scholar] [CrossRef] [PubMed]
- Jem, K.J.; Tan, B. The development and challenges of poly (lactic acid) and poly (glycolic acid). Adv. Ind. Eng. Polym. Res. 2020, 3, 60–70. [Google Scholar] [CrossRef]
- Davy, J.M. Process for the Production of Glycolic Acid. U.S. Patent 10,640,443, 5 May 2020. [Google Scholar]
- Becker, J.; Lange, A.; Fabarius, J.; Wittmann, C. Top value platform chemicals: Bio-based production of organic acids. Curr. Opin. Biotechnol. 2015, 36, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zha, M.; Cao, J.; Yan, H.; Feng, X.; Chen, D.; Yang, C. Glycolic Acid Production from Ethylene Glycol via Sustainable Biomass Energy: Integrated Conceptual Process Design and Comparative Techno-economic–Society–Environment Analysis. ACS Sustain. Chem. Eng. 2021, 9, 10948–10962. [Google Scholar] [CrossRef]
- Van Uytvanck, P.P.; Hallmark, B.; Haire, G.; Marshall, P.J.; Dennis, J.S. Impact of Biomass on Industry: Using Ethylene Derived from Bioethanol within the Polyester Value Chain. ACS Sustain. Chem. Eng. 2014, 2, 1098–1105. [Google Scholar] [CrossRef]
- Kandasamy, S.; Samudrala, S.P.; Bhattacharya, S. The route towards sustainable production of ethylene glycol from a renewable resource, biodiesel waste: A review. Catal. Sci. Technol. 2019, 9, 567–577. [Google Scholar] [CrossRef]
- Hua, X.; Cao, R.; Zhou, X.; Xu, Y. Integrated process for scalable bioproduction of glycolic acid from cell catalysis of ethylene glycol. Bioresour. Technol. 2018, 268, 402–407. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, M.; Sasaki, M.; Hidalgo, A.-R.G.D.; Nakano, M.; Shimizu, S. Glycolic Acid Production Using Ethylene Glycol-Oxidizing Microorganisms. Biosci. Biotechnol. Biochem. 2001, 65, 2265–2270. [Google Scholar] [CrossRef]
- Loder, D.J. Process for Manufacture of Glycolic Acid. U.S. Patent 2,152,852, 4 April 1939. [Google Scholar]
- Häberlein, H.H.; Häberlein, J.T.; Haberlein, M.C.; Ebmeyer, F.; Häberlein, H.; Mohn, H. 54 Process for Preparing a Particularly Pure Glycolic Acid 30 Foreign Application Priority Data. U.S. Patent US5723662A, 18 June 1997. [Google Scholar]
- Kobetz, P.; Lindsay, K. 440 1 Process for the Preparation: Of Glycolic Acid Background of the Invention 1. Field of the Invention. U.S. Patent 3,867,440, 18 February 1975. [Google Scholar]
- Chauhan, S.; DiCosimo, R.; Fallon, R.; Gavagan, J.; Payne, M. Method for Producing Glycolic Acid from Glycolonitrile Using Nitrilase. U.S. Patent USOO641 698OB1, 9 July 2002. [Google Scholar]
- DiCosimo, R.; Panova, A.; O’Keefe, D.; Thompson, J.S.; Payne, M. Enzymatic Production of Glycolic Acid. U.S. Patent US7198927B2, 3 April 2007. [Google Scholar]
- Cotellessa, C.; Peris, K.; Chimenti, S. Glycolic acid and its use in dermatology. J. Eur. Acad. Dermatol. Venereol. 1995, 5, 215–217. [Google Scholar] [CrossRef]
- Ginjupalli, K.; Shavi, G.V.; Averineni, R.K.; Bhat, M.; Udupa, N.; Upadhya, P.N. Poly(α-hydroxy acid) based polymers: A review on material and degradation aspects. Polym. Degrad. Stab. 2017, 144, 520–535. [Google Scholar] [CrossRef]
- Wei, G.; Yang, X.; Gan, T.; Zhou, W.; Lin, J.; Wei, D. High cell density fermentation of Gluconobacter oxydans DSM 2003 for glycolic acid production. J. Ind. Microbiol. Biotechnol. 2009, 36, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hua, X.; Zhou, X.; Xu, Y.; Zhang, W. Continuous co-production of biomass and bio-oxidized metabolite (sorbose) using Gluconobacter oxydans in a high-oxygen tension bioreactor. Bioresour. Technol. 2019, 277, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.; Goto, M.; Noda, S.; Matsutani, M.; Hodoya, Y.; Kataoka, N.; Adachi, O.; Matsushita, K.; Yakushi, T. The 5-Ketofructose Reductase of Gluconobacter sp. Strain CHM43 Is a Novel Class in the Shikimate Dehydrogenase Family. J. Bacteriol. 2021, 203, e0055820. [Google Scholar] [CrossRef] [PubMed]
- Moghadami, F.; Fooladi, J.; Hosseini, R. Introducing a thermotolerant Gluconobacter japonicus strain, potentially useful for coenzyme Q10 production. Folia Microbiol. 2019, 64, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Zahid, N.; Schweiger, P.; Galinski, E.; Deppenmeier, U. Identification of mannitol as compatible solute in Gluconobacter oxydans. Appl. Microbiol. Biotechnol. 2015, 99, 5511–5521. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Zeng, W.; Yu, S.; Li, J.; Zhou, J. Rapid Enabling of Gluconobacter oxydans Resistance to High D-Sorbitol Concentration and High Temperature by Microdroplet-Aided Adaptive Evolution. Front. Bioeng. Biotechnol. 2021, 9, 731247. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Yang, X.; Zhou, W.; Lin, J.; Wei, D. Adsorptive bioconversion of ethylene glycol to glycolic acid by Gluconobacter oxydans DSM 2003. Biochem. Eng. J. 2009, 47, 127–131. [Google Scholar] [CrossRef]
- Hua, X.; Du, G.; Xu, Y. Cost-practical of glycolic acid bioproduction by immobilized whole-cell catalysis accompanied with compressed oxygen supplied to enhance mass transfer. Bioresour. Technol. 2019, 283, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.-J.; Zhou, J.; Zhu, D.; Cai, B.; Lin, J.-P.; Hua, Q.; Wei, D.-Z. Functions of membrane-bound alcohol dehydrogenase and aldehyde dehydrogenase in the bio-oxidation of alcohols in Gluconobacter oxydans DSM 2003. Biotechnol. Bioprocess Eng. 2012, 17, 1156–1164. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, L.; Mao, X.; Lin, J.; Wei, D. Enhancement of cell growth and glycolic acid production by overexpression of membrane-bound alcohol dehydrogenase in Gluconobacter oxydans DSM 2003. J. Biotechnol. 2016, 237, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, Y.; Wang, Y.; Gu, J.; Lu, X.; Liao, X.; Shi, J.; Kim, C.H.; Lye, G.; Baganz, F.; et al. Ethylene glycol and glycolic acid production from xylonic acid by Enterobacter cloacae. Microb. Cell Fact. 2020, 19, 89. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Yao, Y.; Yang, Y.; Zhang, Z.; Gu, J.; Mojovic, L.; Knezevic-Jugovic, Z.; Baganz, F.; Lye, G.; Shi, J.; et al. Ethylene glycol and glycolic acid production by wild-type Escherichia coli. Biotechnol. Appl. Biochem. 2021, 68, 744–755. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Ma, Z.; Yang, L.; Ma, J. Enhanced Bioconversion of Ethylene Glycol to Glycolic Acid by a Newly Isolated Burkholderia sp. EG13. Appl. Biochem. Biotechnol. 2014, 174, 1572–1580. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Mao, Y.; Zhang, X. Metabolic engineering of E. coli for efficient production of glycolic acid from glucose. Biochem. Eng. J. 2015, 103, 256–262. [Google Scholar] [CrossRef]
- Pereira, B.; Li, Z.-J.; De Mey, M.; Lim, C.G.; Zhang, H.; Hoeltgen, C.; Stephanopoulos, G. Efficient utilization of pentoses for bioproduction of the renewable two-carbon compounds ethylene glycol and glycolate. Metab. Eng. 2016, 34, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Ma, N.; Zhu, K.; Mao, Y.; Wei, X.; Zhao, Y. Balancing the carbon flux distributions between the TCA cycle and glyoxylate shunt to produce glycolate at high yield and titer in Escherichia coli. Metab. Eng. 2018, 46, 28–34. [Google Scholar] [CrossRef]
- Hua, X.; Zhou, X.; Xu, Y. Improving techno-economics of bioproduct glycolic acid by successive recycled-cell catalysis of ethylene glycol with Gluconobacter oxydans. Bioprocess. Biosyst. Eng. 2018, 41, 1555–1559. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Hua, X.; Zhou, X.; Xu, B.; Wang, H.; Xu, Y. A roundabout strategy for high-purity glycolic acid biopreparation via a resting cell bio-oxidation catalysis of ethylene glycol. Green Chem. 2022, 24, 5142–5150. [Google Scholar] [CrossRef]
- Handojo, L.; Wardani, A.K.; Regina, D.; Bella, C.; Kresnowati, M.T.A.P.; Wenten, I.G. Electro-membrane processes for organic acid recovery. RSC Adv. 2019, 9, 7854–7869. [Google Scholar] [CrossRef] [PubMed]
- Lapponi, M.J.; Méndez, M.B.; Trelles, J.A.; Rivero, C.W. Cell immobilization strategies for biotransformations. Curr. Opin. Green Sustain. Chem. 2022, 33, 100565. [Google Scholar] [CrossRef]
- He, Y.-C.; Xu, J.-H.; Su, J.-H.; Zhou, L. Bioproduction of Glycolic Acid from Glycolonitrile with a New Bacterial Isolate of Alcaligenes sp. ECU0401. Appl. Biochem. Biotechnol. 2010, 160, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Han, J.; Xu, Y. Electrodialytic bioproduction of xylonic acid in a bioreactor of supplied-oxygen intensification by using immobilized whole-cell Gluconobacter oxydans as biocatalyst. Bioresour. Technol. 2019, 282, 378–383. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Medium | Composition (g/L) | Reference |
|---|---|---|
| A | Sorbitol—80 Yeast Extract—20 (NH4)2SO4—5 KH2PO4—2 MgSO4·7H2O—5 | Wei et al. [18] |
| B | Mannitol—25 Yeast Extract—5 Peptone—3 | CCT (mannitol medium) |
| C | Mannitol—25 Yeast Extract—5 Peptone—3 (NH4)2SO4—5 KH2PO4—2 MgSO4·7H2O—5 | modified mannitol medium |
| D | Glucose—50 Yeast Extract—5 (NH4)2SO4—5 KH2PO4—1 K2HPO4—2 MgSO4·7H2O—0.5 | Zhou et al. [19] |
| E | Glycerol—20 Glucose—5 Yeast Extract—10 | Nguyen et al. [20] |
| F | Fructose—40 Yeast Extract—3 Peptone—3 |
| a | b | ||
|---|---|---|---|
| Carbon Sources | Growing Medium with Best Result | Nitrogen Sources | Medium with Best Carbon Source |
| Sorbitol Mannitol Glycerol Sucrose Glucose Xylose Fructose | Carbon source (25 g/L) + Yeast Extract (5 g/L) Peptone (3 g/L) | Yeast Extract (YE) Peptone (PEP) Ammonium Sulfate (NH) Casein (C) Urea (U) C + PEP YE + PEP NH + PEP U + PEP | Nitrogen source (5 g/L) + Mannitol (25 g/L) |
| Experiments | Independent Variables | ||
|---|---|---|---|
| Mannitol (g/L) | YE (g/L) | PEP (g/L) | |
| 1 | 20 (−1) | 5 (−1) | 2 (−1) |
| 2 | 20 (−1) | 5 (−1) | 10 (1) |
| 3 | 20 (−1) | 20 (−1) | 2 (−1) |
| 4 | 20 (−1) | 20 (1) | 10 (1) |
| 5 | 100 (1) | 5 (−1) | 2 (−1) |
| 6 | 100 (1) | 5 (−1) | 10 (1) |
| 7 | 100 (1) | 20 (1) | 2 (−1) |
| 8 | 100 (1) | 20 (1) | 10 (1) |
| 9 | 5.87 (−1.35) | 12.5 (0) | 6 (0) |
| 10 | 114.12 (1.35) | 12.5 (0) | 6 (0) |
| 11 | 60 (0) | 2.35 (−1.35) | 6 (0) |
| 12 | 60 (0) | 22.65 (1.35) | 6 (0) |
| 13 | 60 (0) | 12.5 (0) | 0.59 (−1.35) |
| 14 | 60 (0) | 12.5 (0) | 11.41 (1.35) |
| 15 (C) | 60 (0) | 12.5 (0) | 6 (0) |
| 16 (C) | 60 (0) | 12.5 (0) | 6 (0) |
| 17 (C) | 60 (0) | 12.5 (0) | 6 (0) |
| Bioreactor Type | Time (h) | GA (g/L) | QP (g/L∙h) | YP/S (g/g) | PSR (%) | BE (%) |
|---|---|---|---|---|---|---|
| BC | 144 | 47.7 | 0.33 | 0.938 | 92.1 | 76.6 |
| STR | 140 | 49.4 | 0.35 | 1.078 | 81.5 | 88.0 |
| Microorganism | Substrate | GA (g/L) | QP (g/L∙h) | YP/S | BE (%) | Reference |
|---|---|---|---|---|---|---|
| Enterobacter cloacae * | xylonic acid | 13 | 0.9 | 0.99 mol/mol | - | Zhang et al. [28] |
| Escherichia coli W3110 | xylonic acid | 10.3 | 0.7 | 0.5 mol/mol | - | Lu et al. [29] |
| Gluconobacter oxydans NL71 | ethylene glycol | 93.5 | 2 | 0.94 g/g | 94.2% | Hua et al. [25] |
| Burkholderia sp. EG13 | ethylene glycol | 60 | 0.5 | - | 98.8% | Gao et al. [30] |
| Escherichia coli BW25113 * | glucose | 56.4 | 0.47 | 0.52 g/g | 61.2% | Deng et al. [31] |
| Gluconobacter oxydans DSM 2003 * | ethylene glycol | 73.3 | 1.63 | - | 93.5% | Zhang et al. [27] |
| Escherichia coli K-12 MG1655 | xylose | 40 | 0.58 | 0.63 g/g | - | Pereira et al. [32] |
| Escherichia coli MG1655 * | glucose | 65.5 | 0.85 | 0.76 g/g | 92.9% | Deng et al. [33] |
| Gluconobacter oxydans CCT 0552 | ethylene glycol | 94.2 | 0.41 | 1.19 g/g | 97.3% | This work |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, I.M.T.S.; de Souza, E.F., Jr.; Brenes, R.G.R.; Grieco, M.A.B.; Bojorge Ramirez, N.I.; Pereira, N., Jr. Improvement in Ethylene Glycol Bioconversion to Glycolic Acid by Gluconobacter oxydans Using Central Composite Experimental Design and Bioreactor Fed-Batch Strategies. Fermentation 2024, 10, 320. https://doi.org/10.3390/fermentation10060320
Santos IMTS, de Souza EF Jr., Brenes RGR, Grieco MAB, Bojorge Ramirez NI, Pereira N Jr. Improvement in Ethylene Glycol Bioconversion to Glycolic Acid by Gluconobacter oxydans Using Central Composite Experimental Design and Bioreactor Fed-Batch Strategies. Fermentation. 2024; 10(6):320. https://doi.org/10.3390/fermentation10060320
Chicago/Turabian StyleSantos, Isabella Maria Tenório Soares, Evanildo F. de Souza, Jr., Ricardo Gonzalo Ramirez Brenes, Maria Angela Bernardes Grieco, Ninoska Isabel Bojorge Ramirez, and Nei Pereira, Jr. 2024. "Improvement in Ethylene Glycol Bioconversion to Glycolic Acid by Gluconobacter oxydans Using Central Composite Experimental Design and Bioreactor Fed-Batch Strategies" Fermentation 10, no. 6: 320. https://doi.org/10.3390/fermentation10060320