
Application of Autochthonous Yeast Saccharomyces cerevisiae XG3 in Treixadura Wines from D.O. Ribeiro (NW Spain): Effect on Wine Aroma



Abstract
:1. Introduction
2. Materials and Methods
2.1. Grapevine Cultivars and Yeast Strains
2.2. Fermentations Trials
2.3. Microbiological Control
2.4. Musts and Wines Chemical Analysis
2.5. Identification and Quantification of Wine Volatile Compounds
2.6. Sensory Evaluation
2.7. Statistical Analysis
3. Results and Discussion
3.1. Fermentations
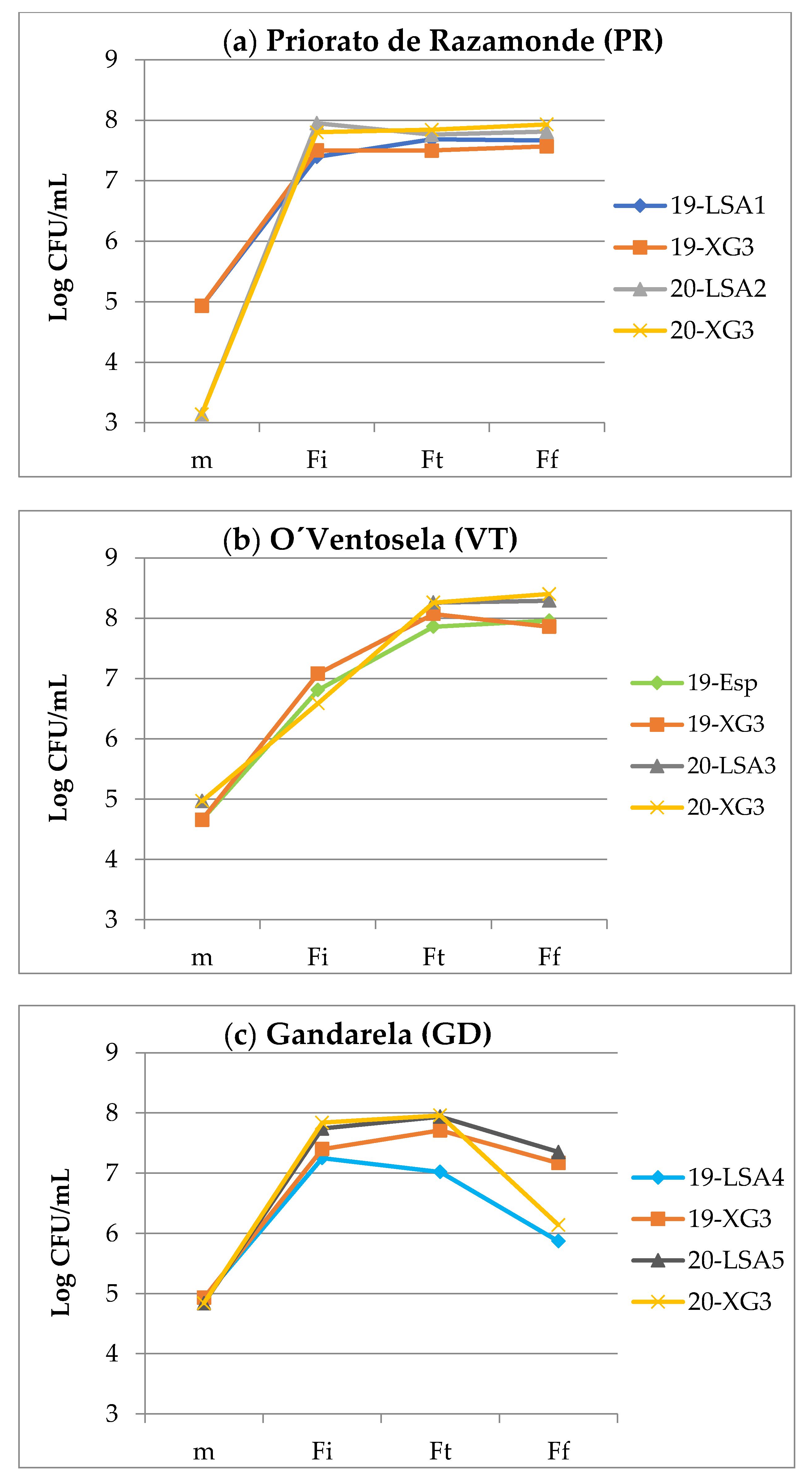
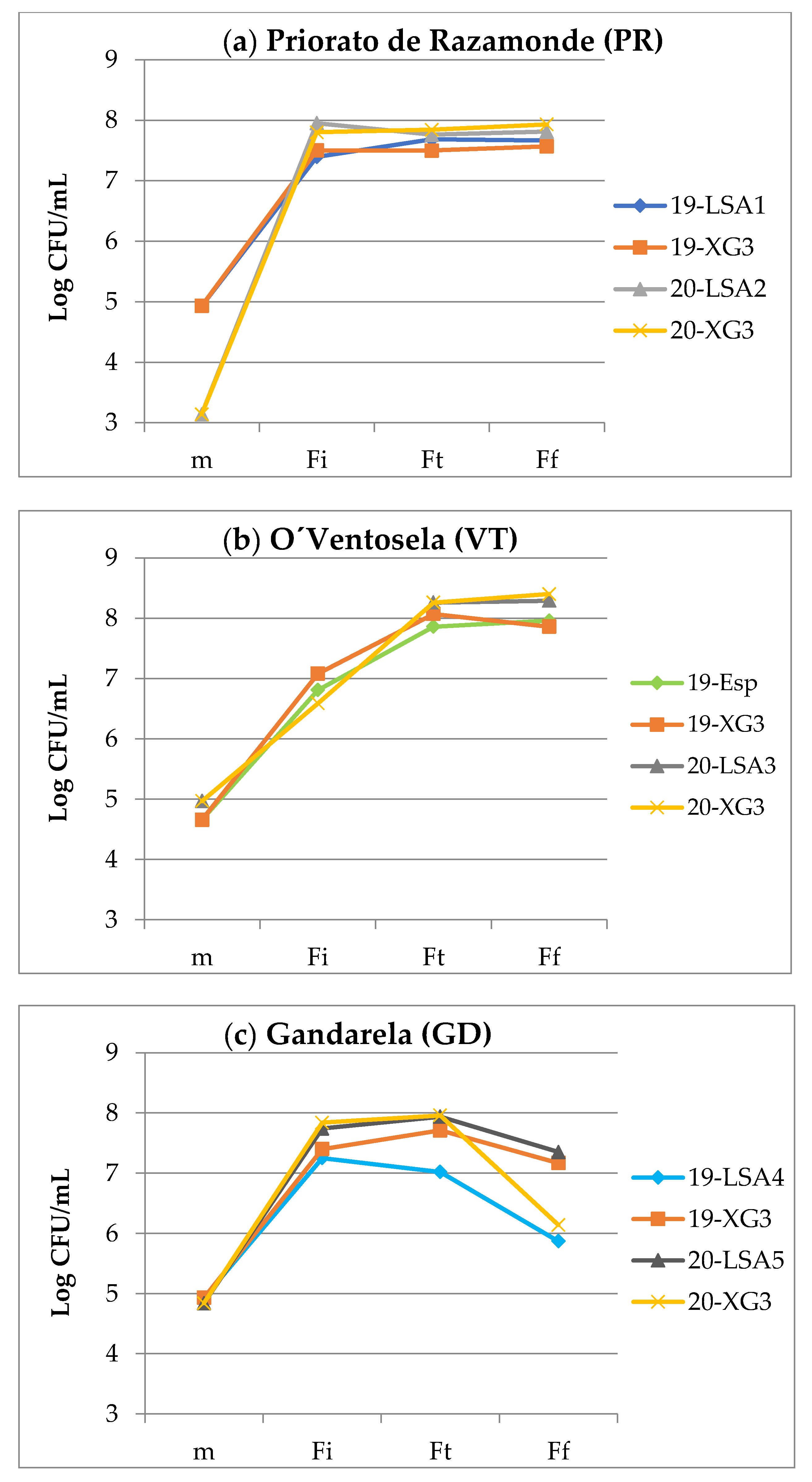
3.1.1. Fermentation Kinetics
3.1.2. Yeast Population Dynamic during Fermentation
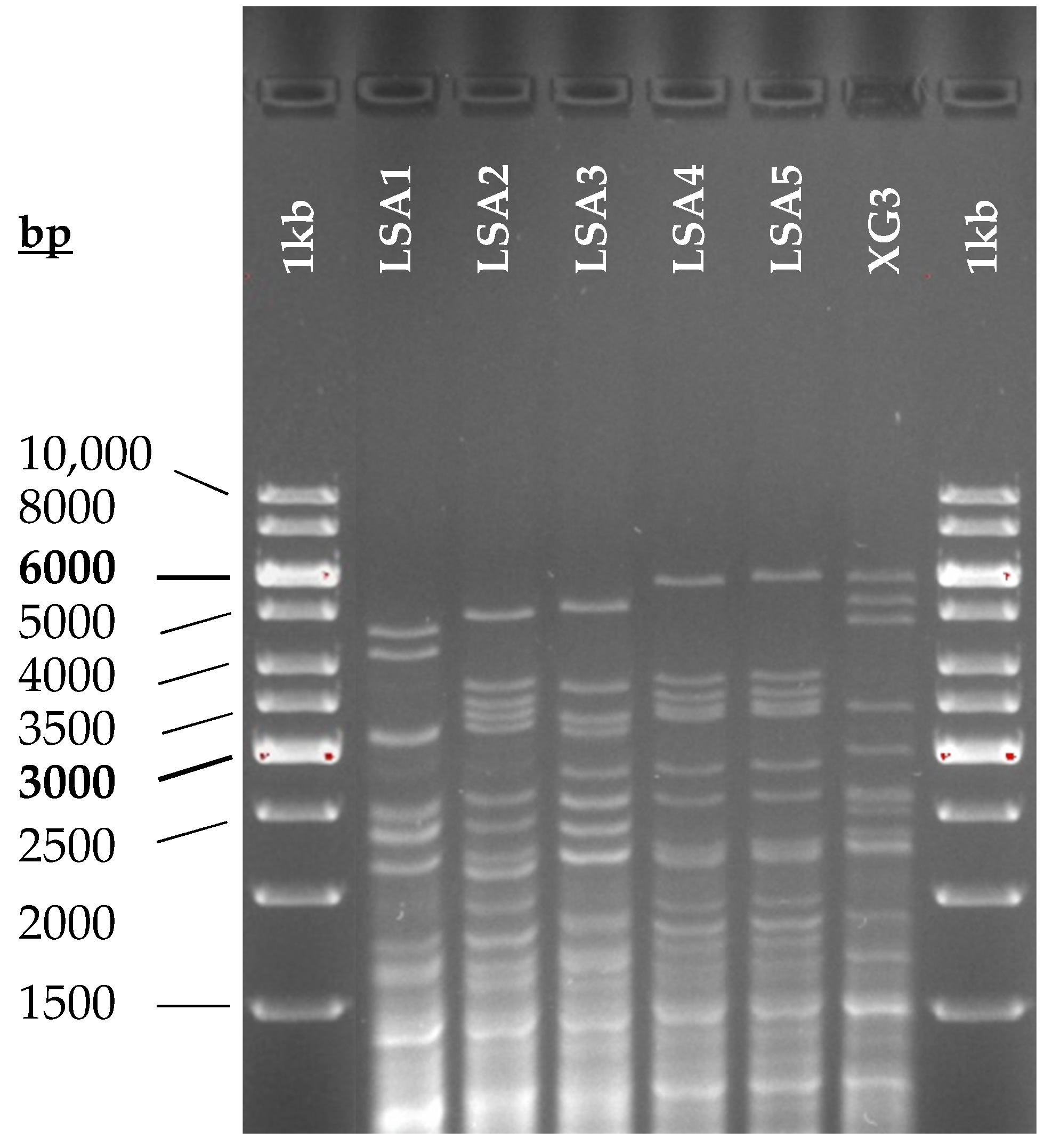
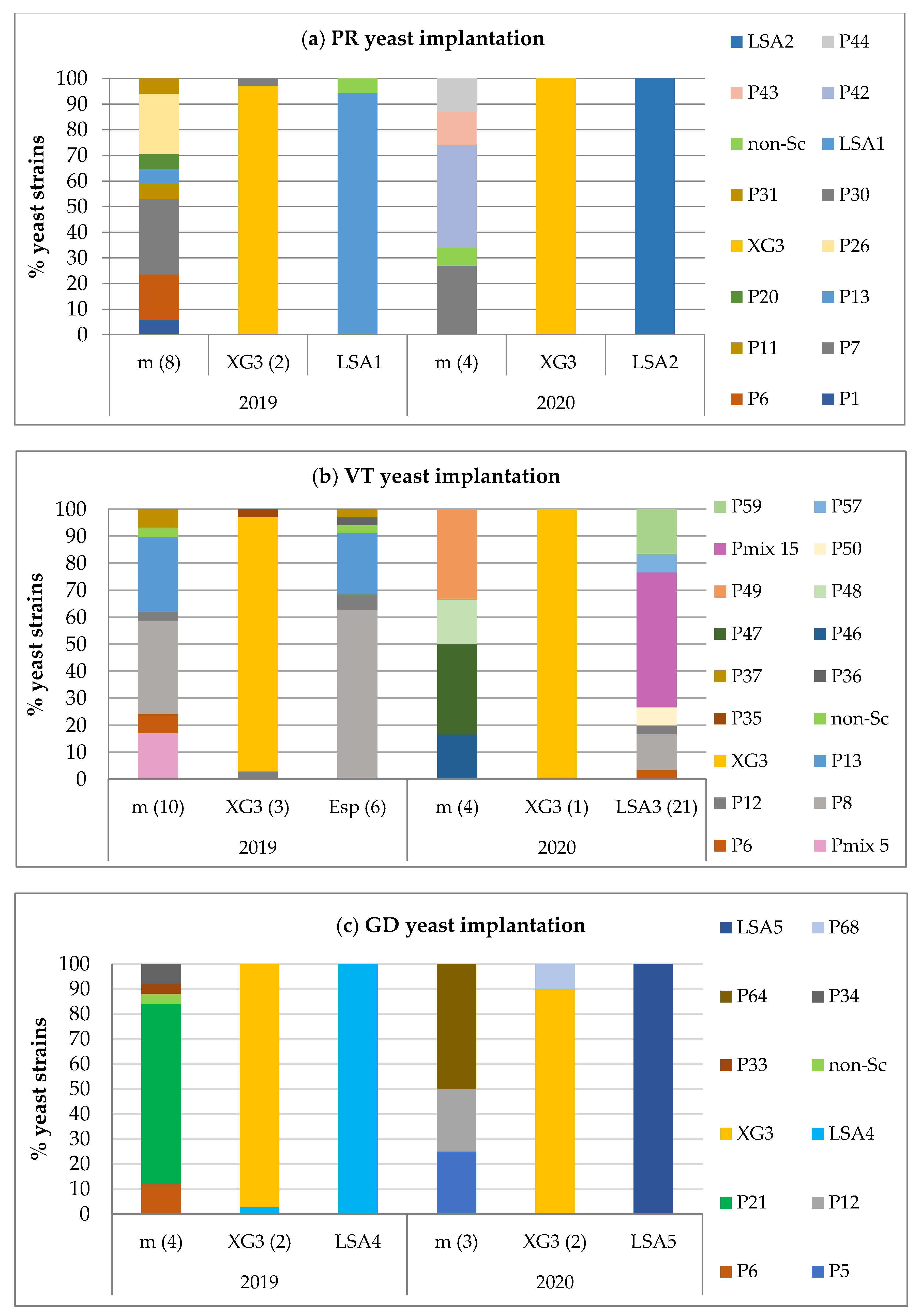
3.1.3. Yeast Implantation
3.2. Chemical Characteristics of Wines
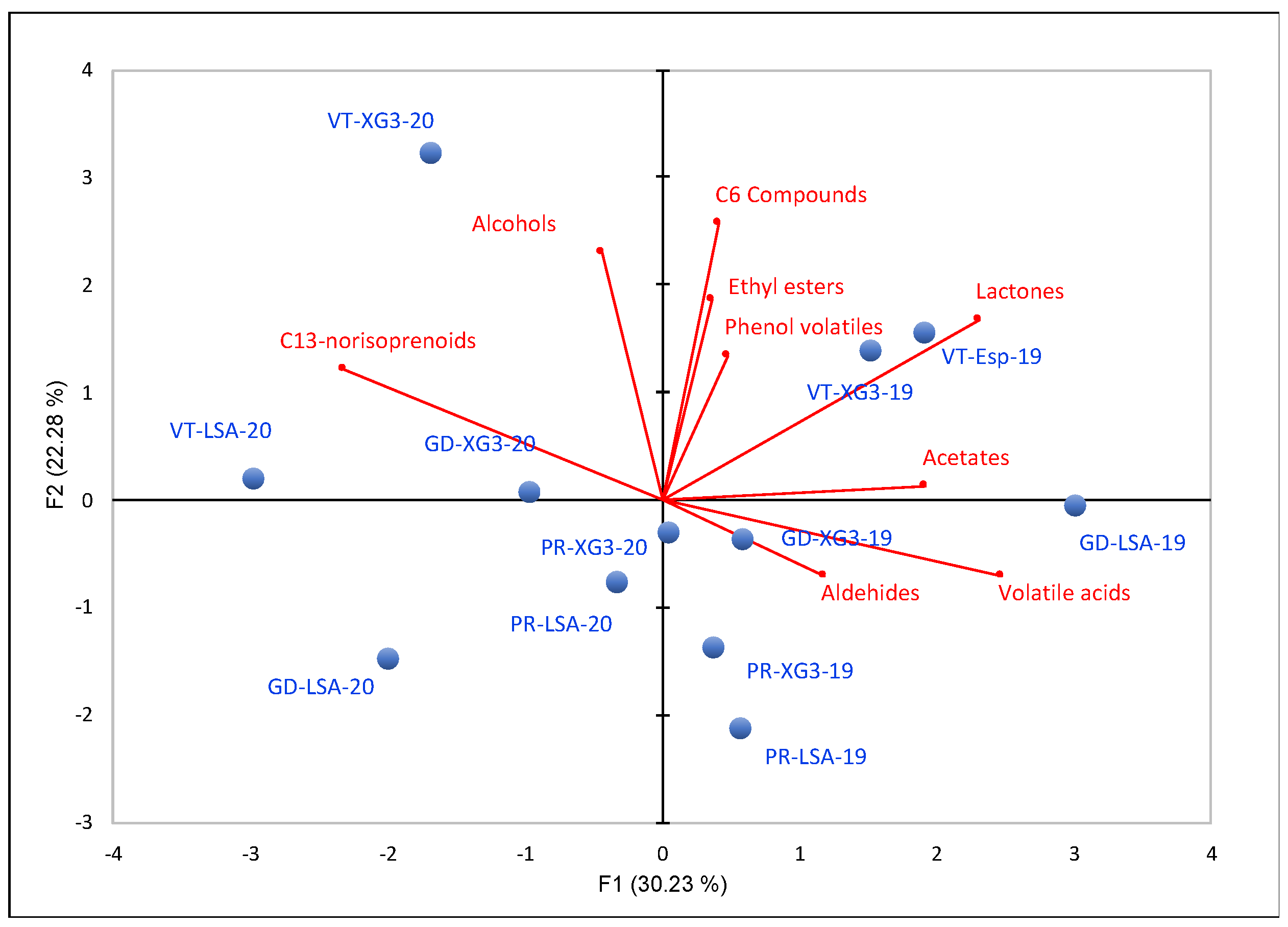
3.3. Wine Aroma Composition
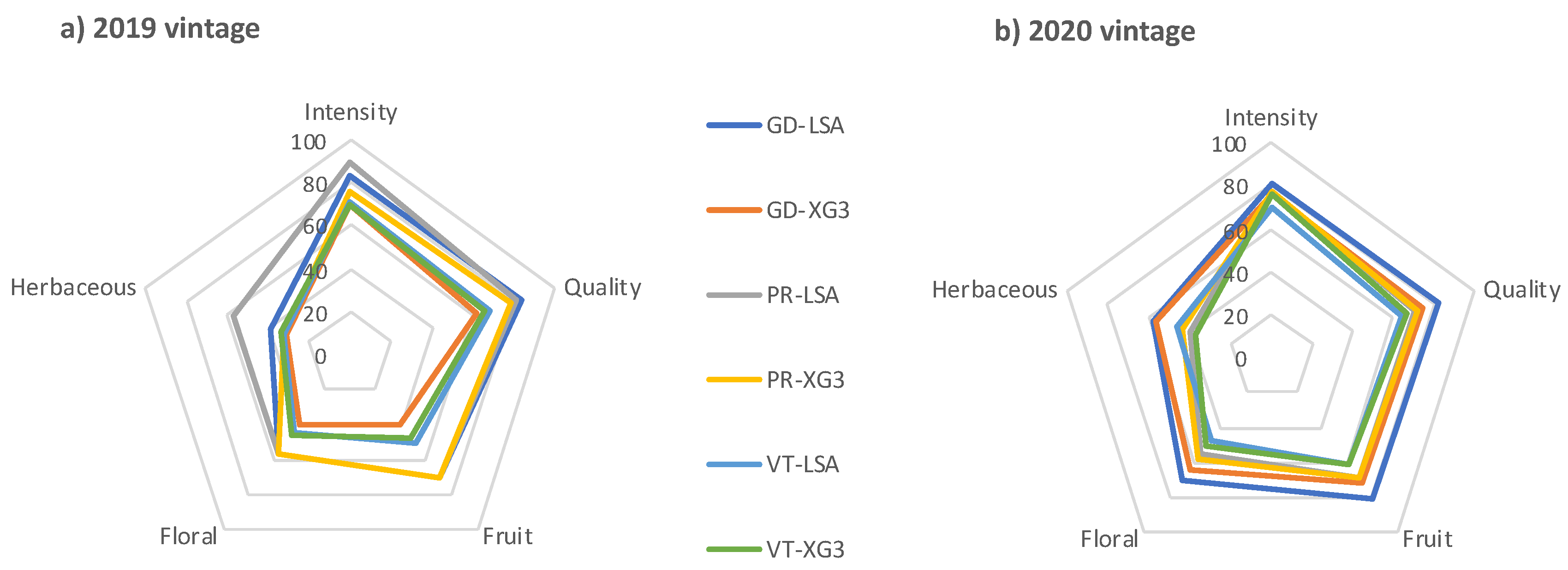
3.4. Sensory Evaluation of Treixadura Wines
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its importance to wine aroma—A review. S. Afr. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Vilanova, M.; Masneuf-Pomarède, I.; Dubourdieu, D. Influence of Saccharomyces cerevisiae strains on general composition and sensory properties of white wines made from Vitis vinifera cv. Albariño. Food Technol. Biotechnol. 2005, 43, 79–83. [Google Scholar]
- Dubourdieu, D.; Tominaga, T.; Masneuf, I.; Peyrot des Gachons, C.; Murat, M.L. The Role of Yeasts in Grape Flavor Development during Fermentation: The Example of Sauvignon blanc. Am. J. Enol. Vitic. 2006, 57, 81–88. [Google Scholar]
- Fleet, G.H.; Heard, G.M. Yeasts—Growth during fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [Green Version]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in alcoholic fermentation processes: Role of physiological fitness and microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef]
- Callejon, R.M.; Clavijo, A.; Ortigueira, P.; Troncoso, A.M.; Paneque, P.; Morales, M.L. Volatile and sensory profile of organic red wines produced by different selected autochthonous and commercial Saccharomyces cerevisiae strains. Anal. Chim. Acta 2010, 660, 68–75. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Grieco, F.; Tufariello, M.; Quarta, A.; Mita, G.; Spano, G.; Grieco, F. Autochthonous fermentation starters for the industrial production of Negroamaro wines. J. Ind. Microbiol. Biotechnol. 2012, 39, 81–92. [Google Scholar] [CrossRef]
- Blanco, P.; Mirás-Avalos, J.M.; Suárez, V.; Orriols, I. Inoculation of Treixadura musts with autochthonous Saccharomyces cerevisiae strains: Fermentative performance and influence on the wine characteristics. Food Sci. Technol. Int. 2013, 19, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Mirás-Avalos, J.M.; Pereira, E.; Orriols, I. Fermentative aroma compounds and sensory profiles of Godello and Albariño wines as influenced by Saccharomyces cerevisiae yeast strains. J. Sci. Food Agric. 2013, 93, 2849–2857. [Google Scholar] [CrossRef]
- Bouzas-Cid, Y.; Falqué, E.; Orriols, I.; Mirás-Avalos, J.M. Effects of irrigation over three years on the amino acid composition of Treixadura (Vitis vinifera L.) musts and wines, and on the aromatic composition and sensory profiles of its wines. Food Chem. 2018, 240, 707–716. [Google Scholar] [CrossRef]
- Falqué, E.; Fernández, E.; Dubourdieu, D. Volatile components of Loureira, Dona Branca, and Treixadura wines. J. Agric. Food Chem. 2002, 50, 538–543. [Google Scholar] [CrossRef]
- Vilanova, M.; Oliveira, J.M. El Potencial Aromático de Las Variedades de Vid Cultivadas en Galicia; Xunta de Galicia: Santiago de Compostela, Spain, 2017; pp. 66–67. [Google Scholar]
- Querol, A.; Barrio, E.; Huerta, T.; Ramón, D. Molecular Monitoring of Wine Fermentations Conducted by Active Dry Yeast Strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- Blanco, P.; Mirás-Avalos, J.M.; Orriols, I. Effect of must characteristics on the diversity of Saccharomyces strains and their prevalence in spontaneous fermentations. J. Appl. Microbiol. 2012, 112, 936–944. [Google Scholar] [CrossRef]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- OIV: International Organisation of Vine and Wine. Compendium of International Methods of Wine and Must Analysis. Vol 1 y 2. Paris, France. 2018. Available online: http://www.oiv.int (accessed on 7 May 2020).
- Coelho, E.; Lemos, M.; Genisheva, Z.; Domingues, L.; Vilanova, M.; Oliveira, J.M. Simple and quick LLME/GC-MS methodology to quantify minor volatile compounds in alcoholic beverages. Molecules 2020, 25, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ISO Norm 8589. Sensory Analysis: General Guidance for the Design of Test Rooms; International Organization for Standardization: Geneva, Switzerland, 1988. [Google Scholar]
- Lawless, H.T.; Heymann, H. Sensory evaluation of food. In Principle and Practices; Kluwer Academic/Plenum Publishers: Boston, MA, USA, 1998. [Google Scholar]
- ISO Norm 3591. Sensory analysis. In Apparatus Wine Tasting Glass; International Organization for Standardization: Geneva, Switzerland, 1977. [Google Scholar]
- Castrillo Cachón, D. Estudio de la Diversidad de Levaduras en uva de Cultivo ecológico y Convencional en Galicia: Patrones Biogeográficos e Influencia en las Características Químicas y Sensoriales del vino. Ph.D. Thesis, Universidad de León, León, Spain, 2018. [Google Scholar]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans Population of in Monoculture and Mixed Fermentations: Impact on Wine Characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. [Google Scholar] [CrossRef]
- Gil-Diaz, M.; Valero, E.; Cabellos, J.M.; García, M.; Arroyo, T. The impact of active dry yeasts in commercial wineries from the Denomination of Origen “Vinos de Madrid”, Spain. 3 Biotech 2019, 9, 382. [Google Scholar] [CrossRef]
- Delteil, D. Aspectos prácticos del levadurado en condiciones mediterráneas.Técnica de inoculación y relación entre la población seleccionada y la población indígena. Rev. F. d’Oenol. 2001, 189, 14–19. [Google Scholar]
- Barrajón, N.; Arévalo-Villena, M.; Rodríguez-Aragón, L.J.; Briones, A. Ecological study of wine yeast in inoculated vats from La Mancha region. Food Control 2009, 20, 778–783. [Google Scholar] [CrossRef]
- Lange, J.N.; Faasse, E.; Tantikachornkiat, M.; Gustafsson, F.S.; Halvorsen, L.C.; Kluftinger, A.; Ledderhof, D.; Durall, D.M. Implantation and persistence of yeast inoculum in Pinot Noir fermentations at three Canadian wineries. Int. J. Food Microbiol. 2014, 180, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Dry, P.R.; Kutyna, D.R.; Francis, I.L.; Henschke, P.A.; Curtin, C.D.; Chambers, P.J. Strategies for reducing alcohol concentration in wine. Aust. J. Grape Wine Res. 2015, 21, 670–679. [Google Scholar] [CrossRef]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef] [PubMed]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeast. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, C.; Mendes-Faia, A.; Mendes-Ferreira, A. The nitrogen source impacts major volatile compounds release by Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microbiol. 2012, 160, 87–93. [Google Scholar] [CrossRef]
- Amerine, M.A.; Ough, C.S. Alcohols. In Methods for Wine and Must Analysis; John Wiley and Sons, Inc.: New York, NY, USA, 1980; pp. 74–127. [Google Scholar]
- Rapp, A.; Mandery, H. Wine aroma. Experentia 1986, 42, 873–884. [Google Scholar] [CrossRef]
- Fukuda, K.; Yamamoto, N.; Kiyokawa, Y.; Yanagiuchi, T.; Wakai, Y.; Kitamoto, K.; Inoue, Y.; Kimura, A. Balance of activities of alcohol acetyltransferase and esterase in Saccharomyces cerevisiae is important for production of isoamyl acetate. Appl. Environ. Microbiol. 1998, 64, 4076–4078. [Google Scholar] [CrossRef] [Green Version]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N.; Lavigne, V. Synthesis of Volatile Phenols by Saccharomyces cerevisiae in Wines. J. Sci. Food Agric. 1993, 62, 191–202. [Google Scholar] [CrossRef]
- Winterhalter, P.; Sefton, M.A.; Williams, P.J. Two dimensional GC-DCCC analysis of the monoterpenes, norisoprenoids and shikimate derived metabolites from Riesling wine. J. Agric. Food Chem. 1990, 38, 1041–1048. [Google Scholar] [CrossRef]
- Versini, G. Sull’aroma del vino ‘traminer aromatico’ o ‘Gewiirztraminer’. Vignevini 1985, 12, 57–65. [Google Scholar]
- Oliveira, J.M.; Faria, M.; Sá, F.; Barros, F.; Araújo, I.M. C6-alcohols as varietal markers for assessment of wine origin. Anal. Chim. Acta 2006, 563, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.I.; Ebeler, S.E.; Heymann, H.; Boss, P.K.; Solomon, P.S.; Trengove, R.D. Interactions between Wine Volatile Compounds and Grape and Wine Matrix Components Influence Aroma Compound Headspace Partitioning. J. Agric. Food Chem. 2009, 57, 10313–10322. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.; Koppel, K. Associations of Volatile Compounds with Sensory Aroma and Flavor: The Complex Nature of Flavor. Molecules 2013, 18, 4887–4905. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Winery (Code) | 2019 | 2020 | ||
|---|---|---|---|---|
| XG3 | LSA | XG3 | LSA | |
| Priorato de Razamonde (PR) | 1000 L | 5000 L (LSA1) | 1000 L | 10,000 L (LSA2) |
| O’Ventosela (VT) | 5000 L | 10,000 L (Esp) | 10,000 L | 40,000 L (LSA3) |
| Gandarela (GD) | 4000 L | 5000 L (LSA4) | 2800 L | 5000 L (LSA5) |
| 2019 | 2020 | |||||
|---|---|---|---|---|---|---|
| Parameter | PR | VT | GD | PR | VT | GD |
| °Brix | 21.5 | 21.2 | 23.2 | 21.4 | 22.6 | 19.6 |
| Sugars (g/L) | 209.0 | 205.5 | 228.5 | 207.8 | 221.6 | 187.4 |
| Total acidity (g tartaric acid/L) | 5.7 | 5.0 | 4.7 | 5.6 | 4.8 | 5.9 |
| pH | 3.51 | 3.69 | -- | 3.56 | -- | 3.57 |
| Malic acid (g/L) | 3.1 | 3.2 | 2.5 | 3.3 | 2.8 | 3.6 |
| Tartaric acid (g/L) | 3.3 | -- | 4.2 | 3.4 | 3.5 | 4.2 |
| α-Amino nitrogen (mg/L) | 152.2 | 150.8 | 134.8 | 152.7 | 133.4 | 147.1 |
| Ammonia (mg/L) | 92.3 | 83.70 | 123.1 | 88.2 | 67.2 | 120.5 |
| Total sulphur dioxide (mg/L) | 70 | 45 | 99 | 76 | 50 | 92 |
| Yeast population (log CFU/mL) * | 4.93 | 4.66 | 4.93 | 3.14 | 4.97 | 4.84 |
| Priorato Razamonde | O’Ventosela | Gandarela | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | PR-LSA | PR-XG3 | VT-LSA § | VT-XG3 | GD-LSA | GD-XG3 | ||||||
| 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | 2019 | 2020 | |
| TA * (g/L) | 6.1 | 5.5 | 6.8 | 6.3 | 4.5 | 4.6 | 4.7 | 5.5 | 5.6 | 4.9 | 5.9 | 4.4 |
| VA ** (g/L) | 0.27 | 0.30 | 0.26 | 0.28 | 0.25 | 0.19 | 0.26 | 0.18 | 0.20 | 0.26 | 0.16 | 0.20 |
| Lactic acid (g/L) | 0.1 | 0.1 | 0.1 | 0.1 | 0.1 | 1.0 | 0.1 | 0.2 | 0.2 | 0.2 | 0.1 | 0.3 |
| Malic acid (g/L) | 2.6 | 2.7 | 2.4 | 2.5 | 3.0 | 1.7 | 2.7 | 3.0 | 2.3 | 2.8 | 2.6 | 2.9 |
| Tartaric acid (g/L) | 2.5 | 2.2 | 3.3 | 3.1 | 1.8 | 2.8 | 1.9 | 2.1 | 2.6 | 1.6 | 2.4 | 1.4 |
| Glucose + fructose (g/L) | 3.2 | 0.2 | 3.9 | 0.7 | 0.2 | 0.2 | 0.2 | 0.2 | 0.4 | 0.2 | 0.2 | 0.2 |
| Glycerol (g/L) | 4.3 | 5.0 | 5.0 | 5.1 | 4.0 | 4.8 | 5.0 | 5.7 | 6.3 | 6.5 | 6.2 | 5.5 |
| Alcohol content (%) v/v)) (%vol.) | 13.1 | 13.2 | 12.8 | 13.0 | 13.2 | 12.8 | 13.0 | 12.6 | 12.2 | 14.0 | 12.9 | 12.9 |
| pH | 3.32 | 3.50 | 3.24 | 3.31 | 3.71 | 3.54 | 3.67 | 3.65 | 3.67 | 3.82 | 3.59 | 3.84 |
| Free SO2 (mg/L) | 43 | 44 | 42 | 36 | 10 | 10 | 10 | 10 | 12 | 10 | 10 | 12 |
| Total SO2 (mg/L) | 92 | 125 | 93 | 96 | 69 | 118 | 52 | 83 | 92 | 48 | 69 | 68 |
| Priorato de Razamonde | O’Ventosela | Gandarela | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Chemical Group | Year | PR-LSA | PR-XG3 | Sig. | VT-LSA | VT-XG3 | Sig. | GD-LSA | GD-XG3 | Sig. |
| Higher Alcohols | 2019 | 23,947 | 34,727 | ns | 31,145 | 61,864 | ns | 45,665 | 59,500 | * |
| 2020 | 41,824 | 34,218 | ns | 35,865 | 86,599 | * | 46,669 | 43,471 | ns | |
| C6 compounds | 2019 | 537 | 704 | ns | 1451 | 1151 | ns | 825 | 1049 | ** |
| 2020 | 734 | 561 | ns | 1210 | 1163 | ns | 410 | 706 | * | |
| Acetates | 2019 | 13,708 | 8468 | ** | 6647 | 19,068 | ** | 17,913 | 10,106 | *** |
| 2020 | 10,860 | 10,150 | ns | 1693 | 10,595 | ** | 8405 | 10,704 | * | |
| Ethyl esters | 2019 | 2676 | 2778 | ns | 5186 | 3443 | ns | 3853 | 2286 | *** |
| 2020 | 4236 | 4292 | ns | 3337 | 4150 | ns | 3065 | 4305 | ** | |
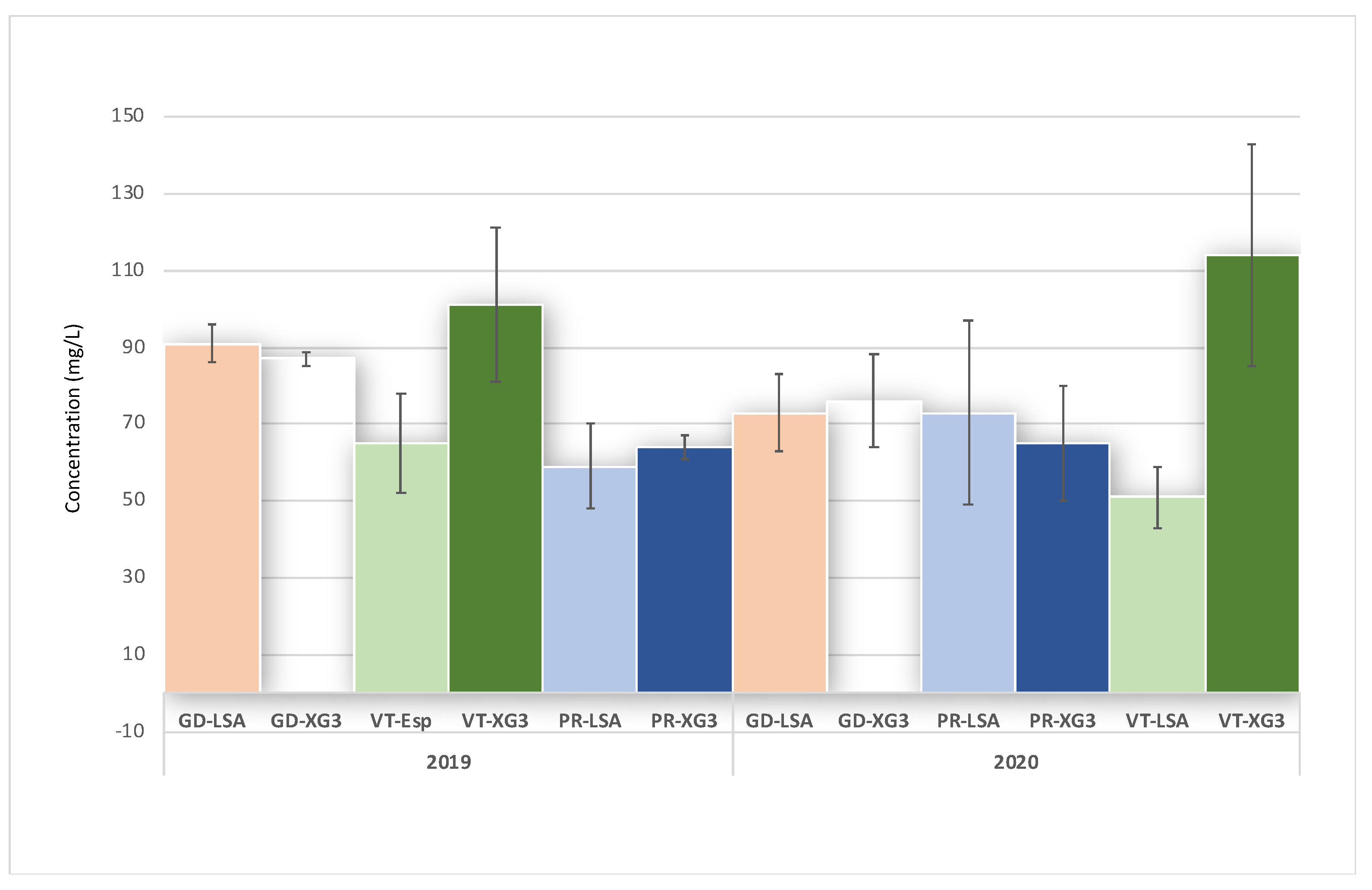
| Volatile acids | 2019 | 17,732 | 17,365 | ns | 20,684 | 15,346 | ns | 22,355 | 13,773 | *** |
| 2020 | 15,168 | 15,230 | ns | 9113 | 11,329 | ns | 14,682 | 15,918 | ns | |
| Volatile Phenols | 2019 | 392 | 345 | ns | 254 | 402 | ns | 379 | 395 | ns |
| 2020 | 163 | 673 | ** | 194 | 643 | * | 133 | 334 | ** | |
| Lactones | 2019 | 47 | 73 | * | 157 | 126 | ns | 131 | 76 | ** |
| 2020 | 42 | 51 | ns | 38 | 72 | ns | 25 | 49 | ns | |
| Aldehydes | 2019 | 24 | 32 | ns | nd | nd | - | 76 | 139 | ** |
| 2020 | nd | nd | - | nd | nd | - | nd | nd | - | |
| C13-norisoprenoids | 2019 | nd | nd | - | nd | nd | - | nd | nd | - |
| 2020 | nd | nd | - | 99 | 127 | ns | 92 | 93 | ns | |
| Year | Wines | Intensity | Quality | Fruit | Floral | Herbaceous |
|---|---|---|---|---|---|---|
| 2019 | GD-LSA | 7.0 ab | 7.0 a | 6.1 a | 5.3 | 4.8 |
| GD-XG3 | 4.9 c | 3.8 b | 3.5 ab | 3.7 | 3.2 | |
| PR-LSA | 8.0 a | 6.7 a | 6.1 a | 4.1 | 5.4 | |
| PR-XG3 | 5.7 bc | 6.1 ab | 5.0 ab | 4.1 | 3.4 | |
| VT-LSA | 5.0 bc | 4.7 ab | 3.3 b | 4.5 | 3.4 | |
| VT-XG3 | 4.8 c | 4.2 b | 3.7 ab | 4.7 | 3.6 | |
| 2020 | GD-LSA | 6.6 a | 6.8 a | 6.5 a | 4.9 a | 3.3 |
| GD-XG3 | 5.7 ab | 5.6 ab | 5.2 ab | 4.1 ab | 3.2 | |
| PR-LSA | 6.1 ab | 5.0 ab | 4.8 ab | 3.7 ab | 2.9 | |
| PR-XG3 | 6.1 ab | 5.2 ab | 4.8 ab | 4.0 ab | 2.8 | |
| VT-LSA | 4.9 b | 4.1 b | 3.7 b | 2.2 b | 3.1 | |
| VT-XG3 | 5.7 ab | 4.5 b | 4.5 ab | 3.1 ab | 2.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, P.; Vázquez-Alén, M.; Garde-Cerdán, T.; Vilanova, M. Application of Autochthonous Yeast Saccharomyces cerevisiae XG3 in Treixadura Wines from D.O. Ribeiro (NW Spain): Effect on Wine Aroma. Fermentation 2021, 7, 31. https://doi.org/10.3390/fermentation7010031
Blanco P, Vázquez-Alén M, Garde-Cerdán T, Vilanova M. Application of Autochthonous Yeast Saccharomyces cerevisiae XG3 in Treixadura Wines from D.O. Ribeiro (NW Spain): Effect on Wine Aroma. Fermentation. 2021; 7(1):31. https://doi.org/10.3390/fermentation7010031
Chicago/Turabian StyleBlanco, Pilar, María Vázquez-Alén, Teresa Garde-Cerdán, and Mar Vilanova. 2021. "Application of Autochthonous Yeast Saccharomyces cerevisiae XG3 in Treixadura Wines from D.O. Ribeiro (NW Spain): Effect on Wine Aroma" Fermentation 7, no. 1: 31. https://doi.org/10.3390/fermentation7010031