
Increased Extracellular Saponin Production after the Addition of Rutin in Truffle Liquid Fermentation and Its Antioxidant Activities

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Raw Materials
2.2. Culture Medium
2.3. Inoculation and Culture
2.4. Experimental Design
2.4.1. Nutrient Selection of the Fermentation Medium
2.4.2. Orthogonal Experimental Design for Medium Composition Optimization
2.4.3. Orthogonal Experimental Design for Culture Condition Optimization
2.5. Analytical Methods
2.5.1. Determination of Extracellular Saponin
2.5.2. Measurement of Dry Biomass
2.5.3. Test of Glucose Content
2.5.4. Determination of Antioxidant Activities
3. Results and Discussion
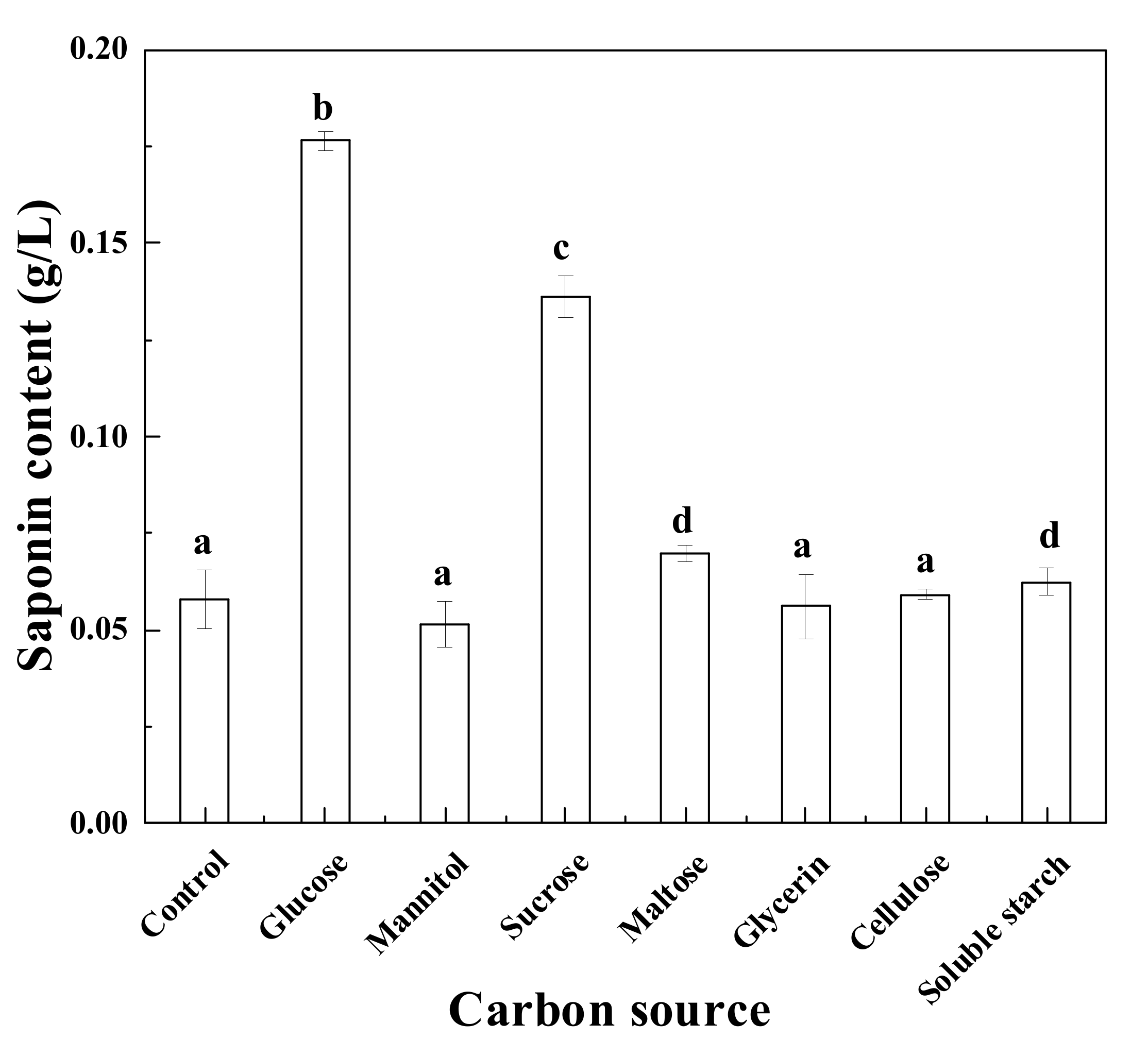
3.1. Effects of Various Carbon Sources on Saponin Production
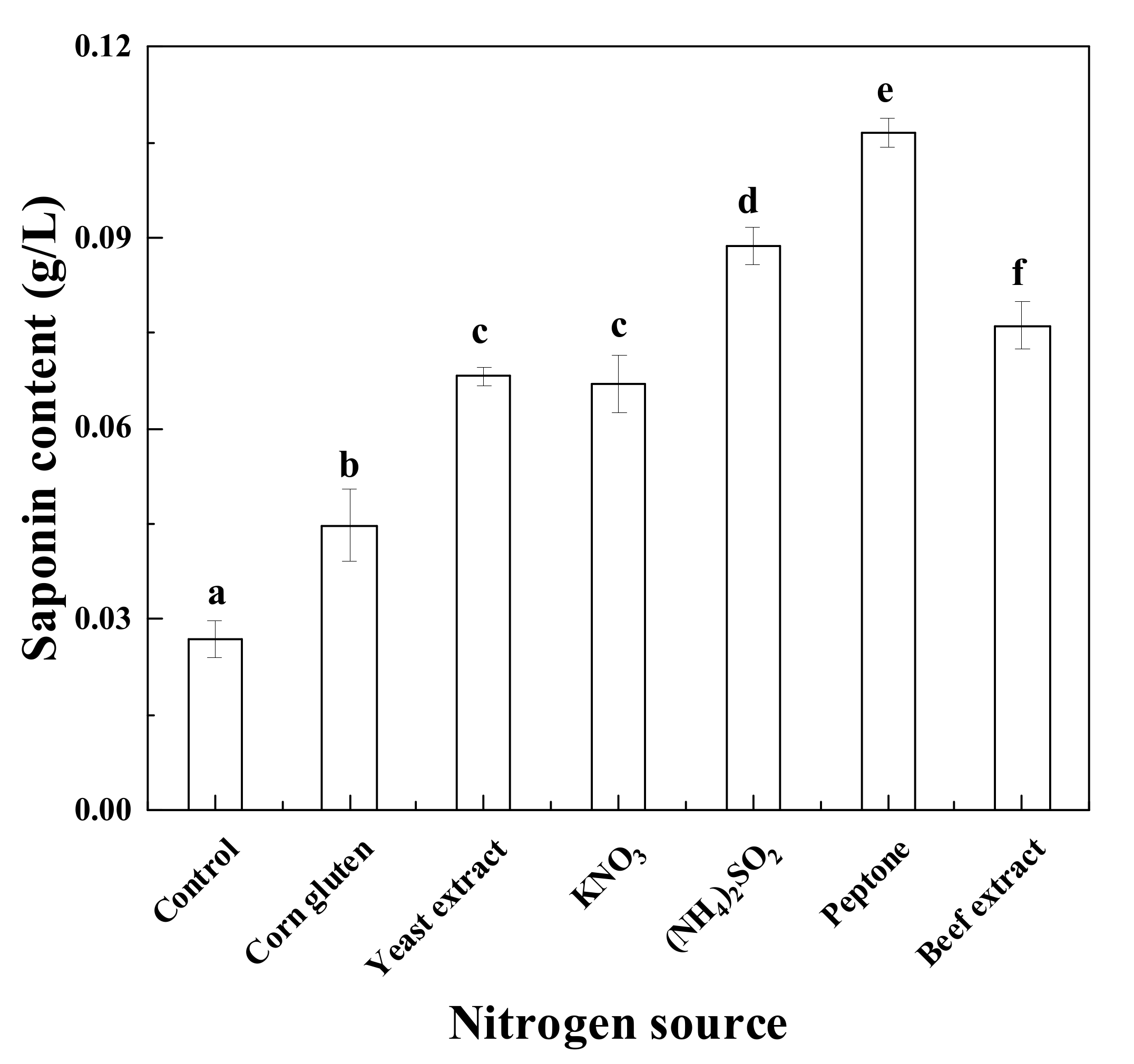
3.2. Effects of Various Nitrogen Sources on Saponin Production
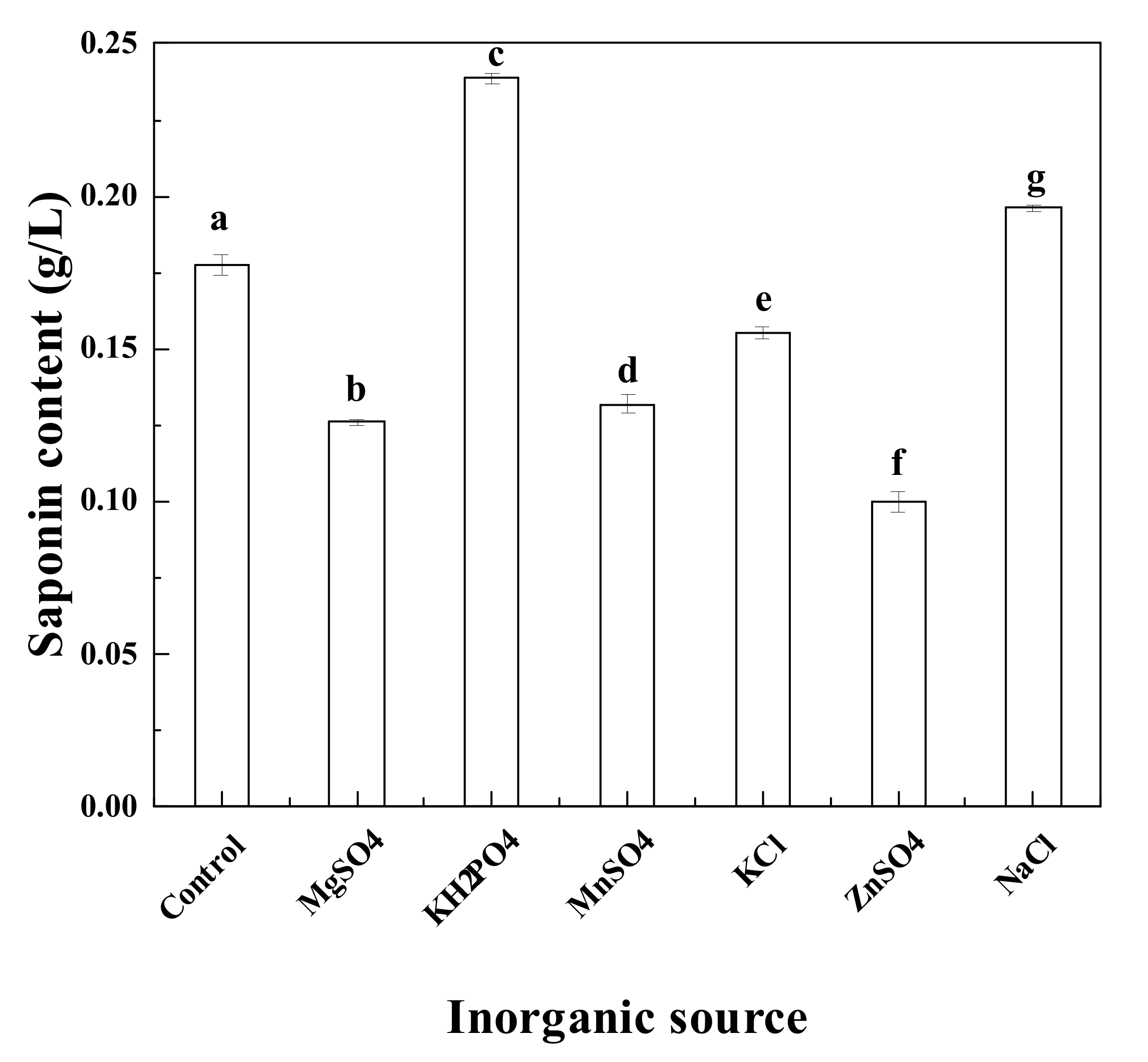
3.3. Effects of Various Inorganic Salts on Saponin Production
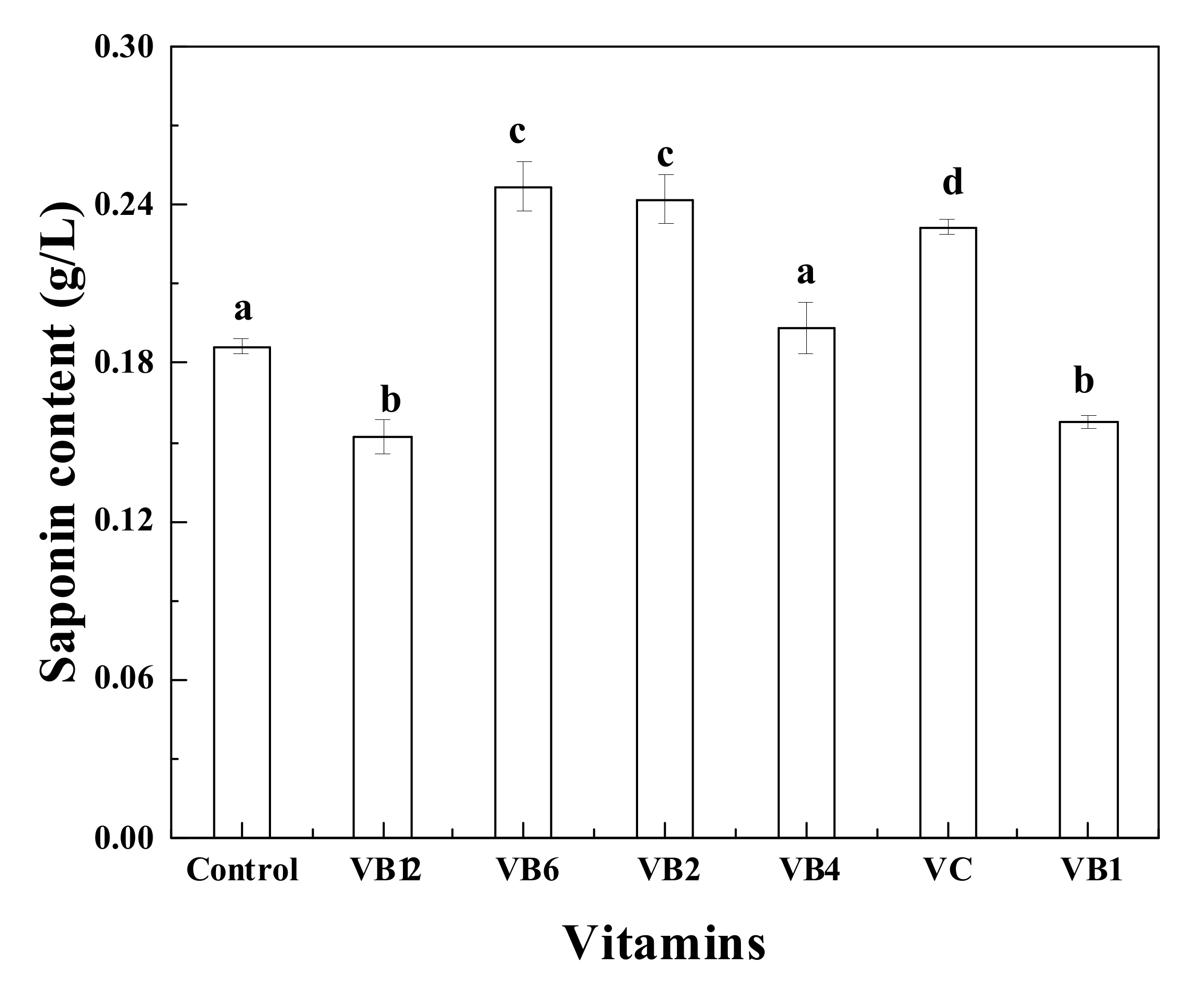
3.4. Effects of Various Vitamins on Saponin Production
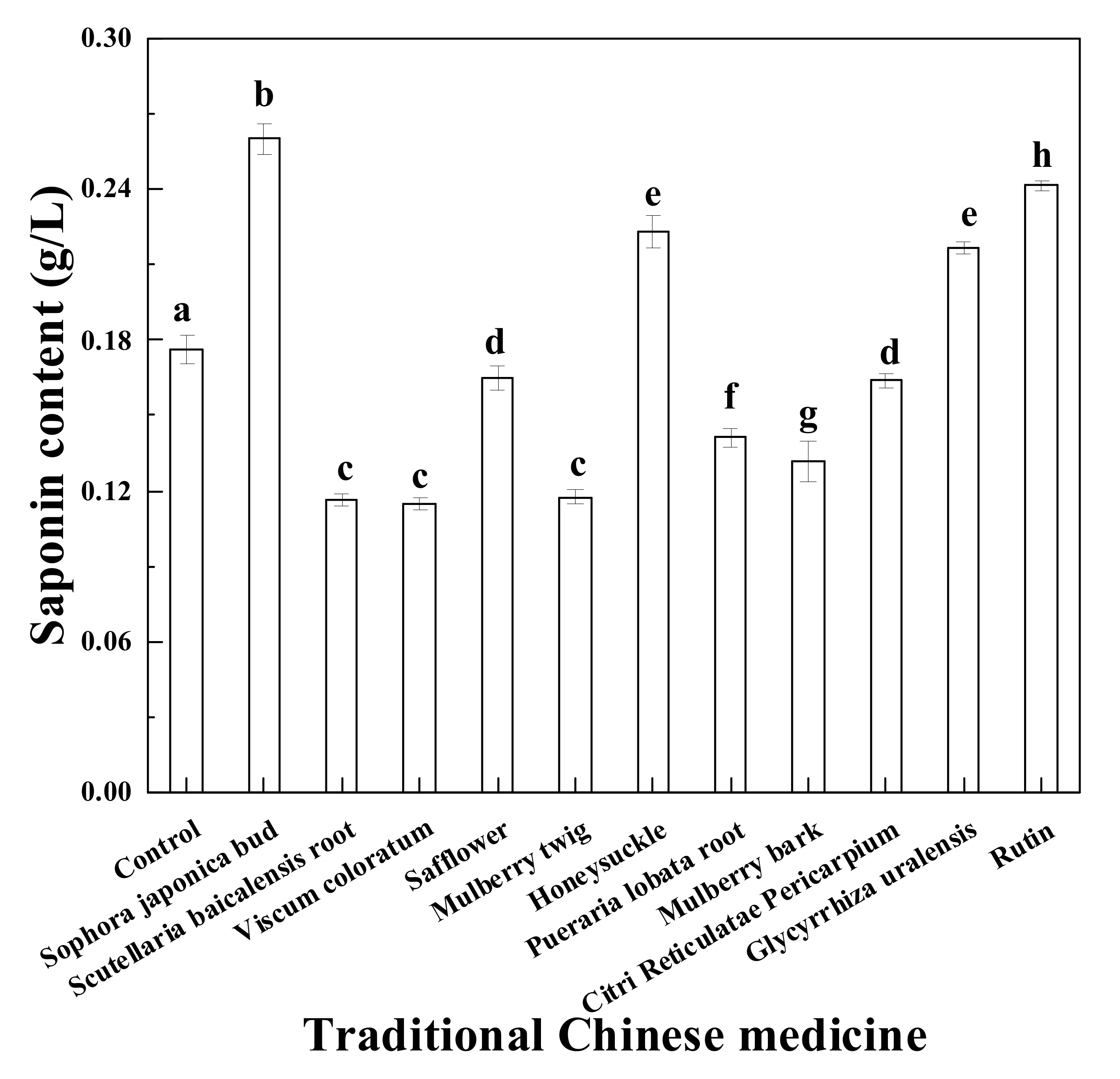
3.5. Effects of Various Traditional Chinese Medicines on Saponin Production
3.6. Analysis of the Orthogonal Results of Medium Composition Optimization
3.7. Analysis of the Orthogonal Results of Fermentation Condition Optimization
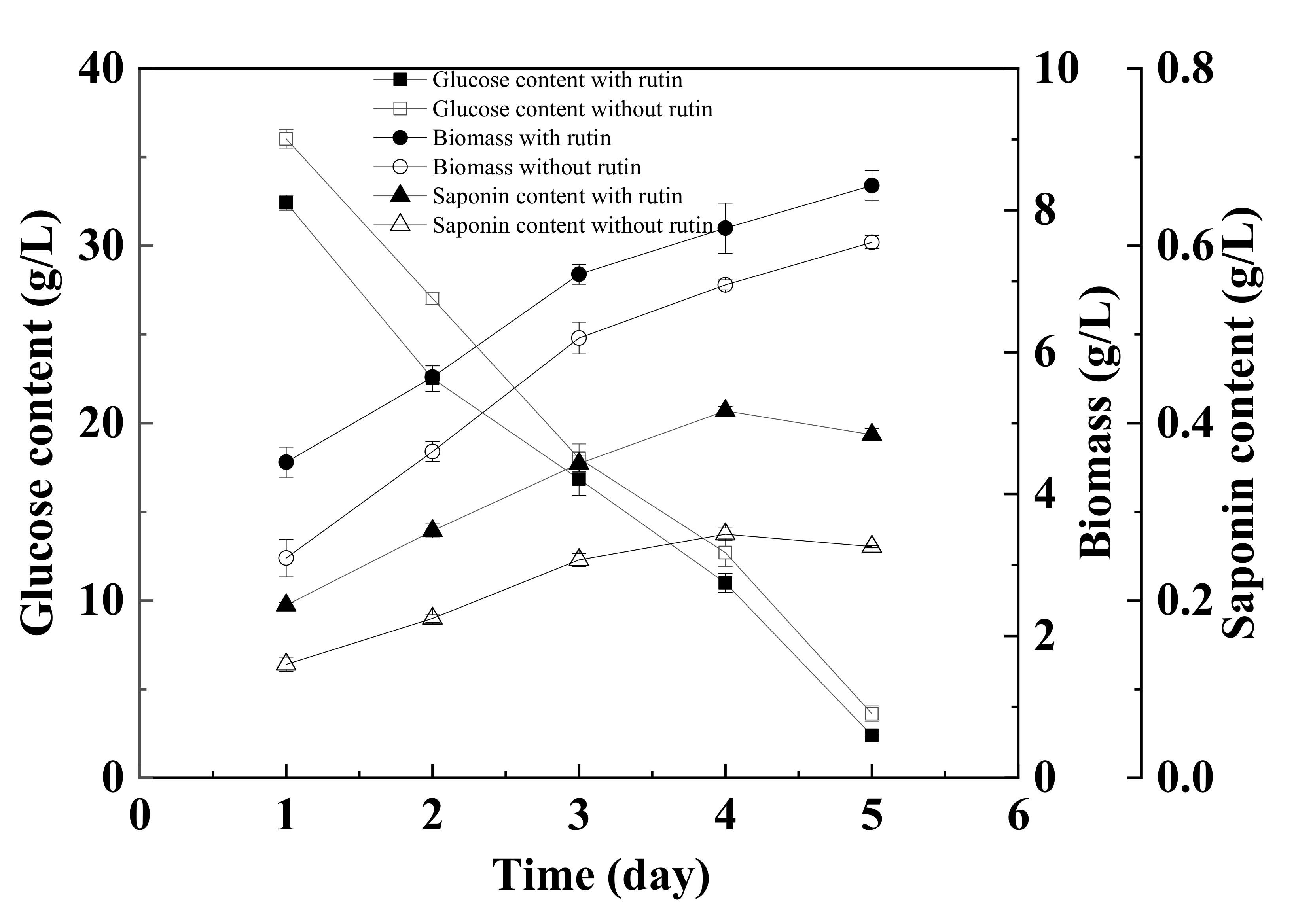
3.8. Truffle Fermentation under Optimal Medium Composition and Culture Conditions
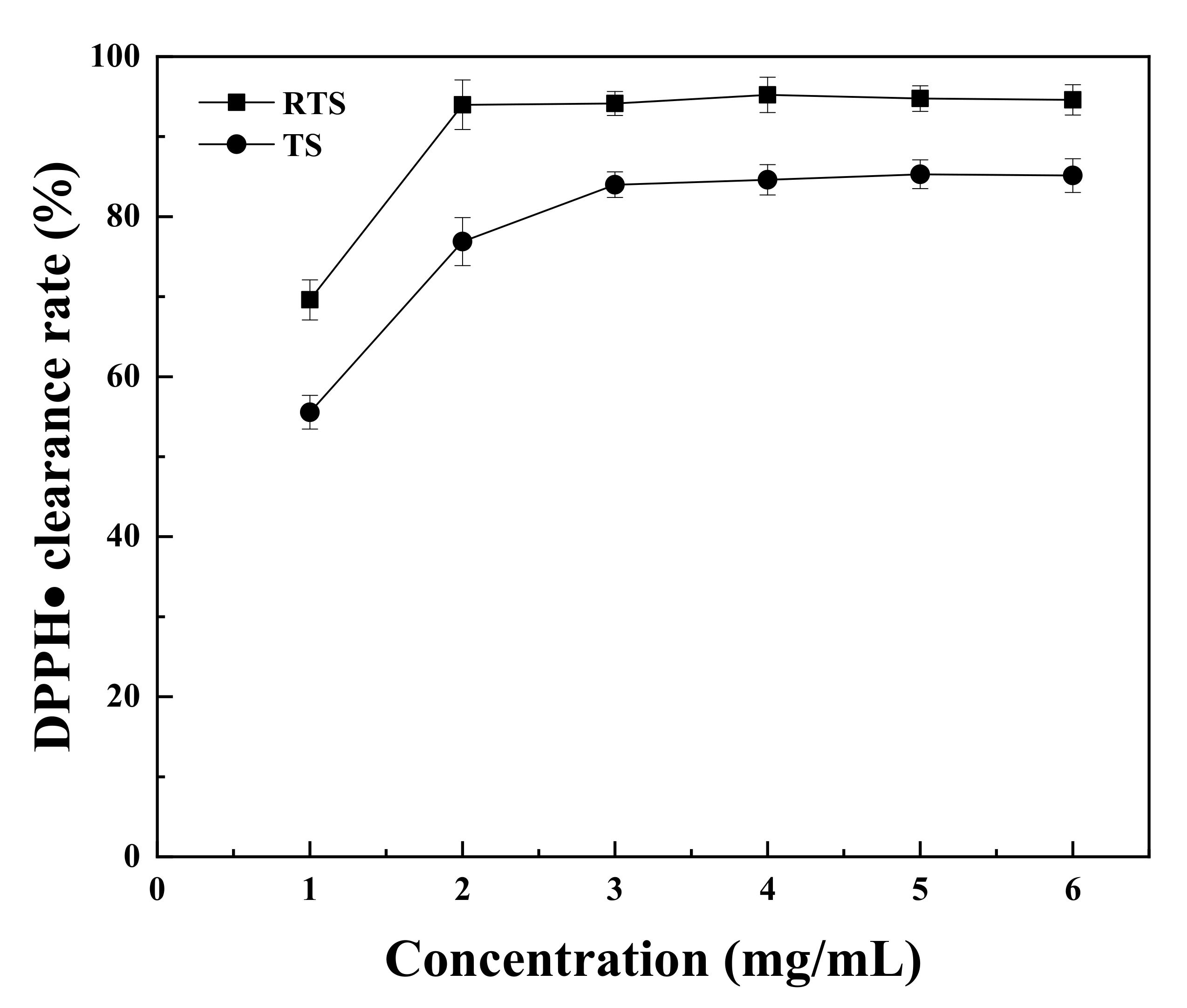
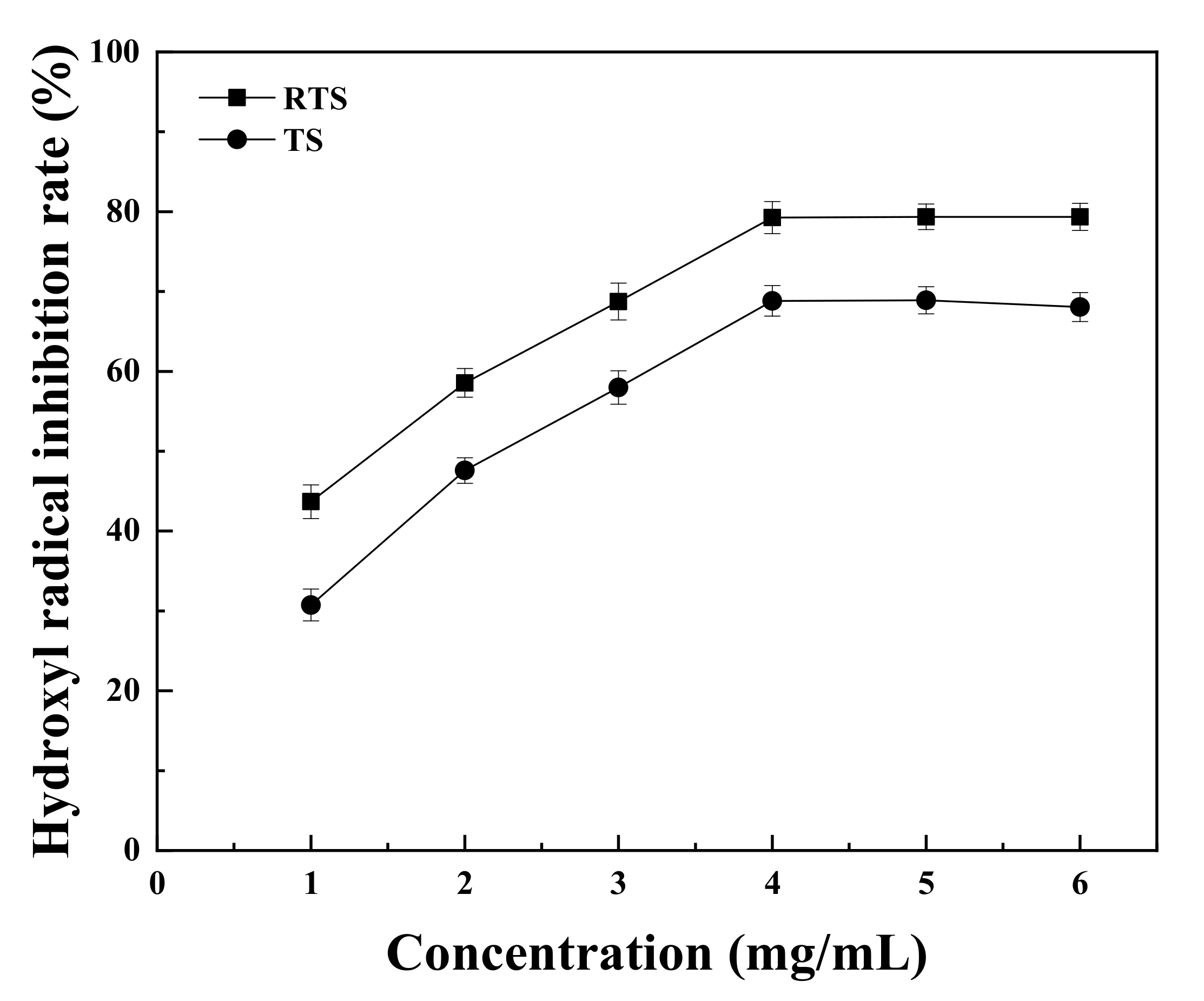
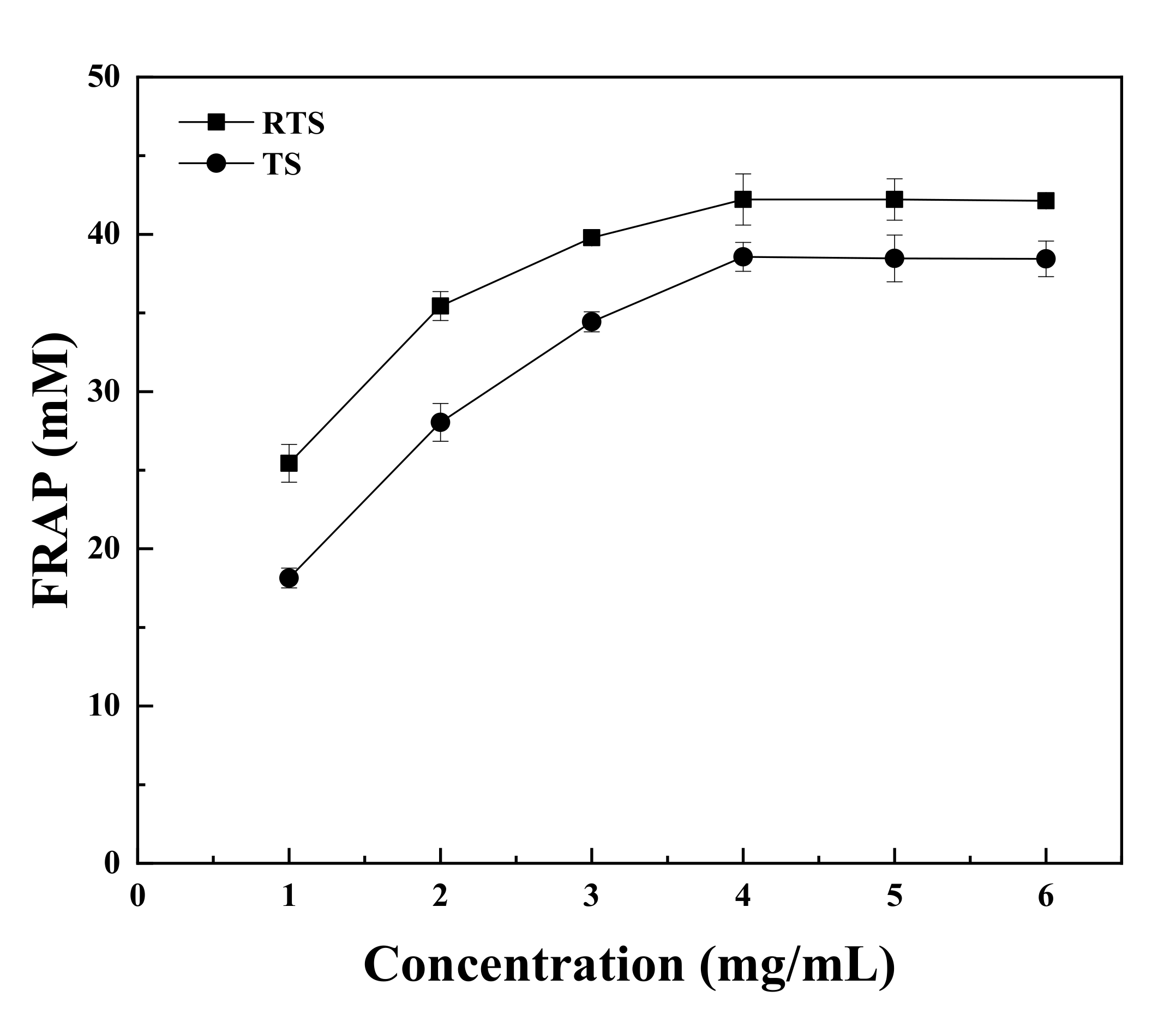
3.9. Antioxidant Activities of Trufflle Saponin
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Magedans, Y.V.D.S.; Phillips, M.A.; Fett-Neto, A.G. Production of plant bioactive triterpenoid saponins: From metabolites to genes and back. Phytochem. Rev. 2020, 6, 1–22. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Han, F.Y.; Chen, J.J.; Wang, W.; Zhang, Y.; Yao, G.D.; Song, S.J. Timosaponin AIII, a steroidal saponin, exhibits anti-tumor effect on taxol-resistant cells in vitro and in vivo. Steroids 2019, 146, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.X.; Chen, L.P.; Wang, J.J.; Wang, K.W.; Zhou, J.Y. Structure-function relationship of the saponins from the roots of Platycodon grandiflorum for hemolytic and adjuvant activity. Int. Immunopharmacol. 2011, 11, 2047–2056. [Google Scholar] [CrossRef]
- Jeepipalli, S.; Du, B.; Sabitaliyevich, U.Y.; Xu, B.J. New insights into potential nutritional effects of dietary saponins in protecting against the development of obesity. Food Chem. 2020, 318, 126474. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.G.; Park, H.M.; Yoon, K.S. Analysis of saponin composition and comparison of the antioxidant activity of various parts of the quinoa plant (Chenopodium quinoa Willd.). Food Sci. Nutr. 2020, 8, 694–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, S.M.; Haq, A.U.; Byrd, J.A.; Berhow, M.A.; Cartwright, A.L.; Bailey, C.A. Haemolytic and antimicrobial activities of saponin -rich extracts from guar meal. Food Chem. 2010, 119, 600–605. [Google Scholar] [CrossRef]
- RamLi, N.H.; Yusup, S.; Quitain, A.T.; Johari, K.; Kueh, B.W.B. Optimization of saponin extract using microwave-assisted extraction as a sustainable biopesticide to reduce Pomacea canaliculata population in paddy cultivation. Sustain. Chem. Pharm. 2019, 11, 23–35. [Google Scholar] [CrossRef]
- Mohaddes-Kamranshahi, M.; Jafarizadeh-Malmiri, H.; Simjoo, M.; Jafarizad, A. Evaluation of the saponin green extraction from Ziziphus spina-christi leaves using hydrothermal, microwave and Bain-Marie water bath heating methods. Green Process. Synth. 2019, 8, 62–67. [Google Scholar] [CrossRef]
- Hu, J.L.; Nie, S.P.; Huang, D.F.; Li, C.; Xie, M.Y. Extraction of saponin from Camellia oleifera cake and evaluation of its antioxidant activity. Int. J. Food Sci. Technol. 2012, 47, 1676–1687. [Google Scholar] [CrossRef]
- Li, W.; Liu, Z.B.; Wang, Z.; Chen, L.; Sun, Y.S.; Hou, J.G.; Zheng, Y.N. Application of accelerated solvent extraction to the investigation of saikosaponins from the roots of Bupleurum falcatum. J. Sep Sci. 2010, 33, 1870–1876. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Guo, W.H.; Sun, X.Y.; Li, K.H.; You, X.L. A culture system for the stable and high-efficiency proliferation of adventitious roots of Panax notoginseng and ginsenoside accumulation. Ind. Crop. Prod. 2020, 157, 112882. [Google Scholar] [CrossRef]
- Długosz, M.; Markowski, M.; Pczkowski, C. Source of nitrogen as a factor limiting saponin production by hairy root and suspension cultures of Calendula officinalis L. Acta Physiol. Plant. 2018, 40, 35. [Google Scholar] [CrossRef] [Green Version]
- Bai, Y.; Zhang, L.W.; Jin, W.W.; Wei, M.; Zhou, P.P.; Zheng, G.H.; Niu, L.L.; Nie, L.; Zhang, Y.L.; Wang, H.Y.; et al. In situ high-valued utilization and transformation of sugars from Dioscorea zingiberensis C.H. Wright for clean production of diosgenin. Bioresour. Technol. 2015, 196, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Zhang, T.; Pang, X.; Wei, Y.; Liu, H.; Zhang, Y.; Ma, B.; Yu, L. Isolation of endophytic fungi from Dioscorea zingiberensis C. H. Wright and application for diosgenin production by solid-state fermentation. Appl. Microbiol. Biotechnol. 2018, 102, 5519–5532. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Wang, C.; Sun, W.; Zhou, A.; Wang, Y.; Zhang, G.; Zhou, X.; Huo, Y.; Li, C. Boosting 11-oxo-beta-amyrin and glycyrrhetinic acid synthesis in Saccharomyces cerevisiae via pairing novel oxidation and reduction system from legume plants. Metab. Eng. 2018, 45, 43–50. [Google Scholar] [CrossRef]
- Zhuang, Y.; Yang, G.Y.; Chen, X.; Liu, Q.; Zhang, X.; Deng, Z.; Feng, Y. Biosynthesis of plant-derived ginsenoside rh2 in yeast via repurposing a key promiscuous microbial enzyme. Metab. Eng. 2017, 42, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Fan, J.; Wang, C.; Feng, X.; Li, C. Enhancing oleanolic acid production in engineered Saccharomyces cerevisiae. Bioresour. Technol. 2018, 257, 339–343. [Google Scholar] [CrossRef] [Green Version]
- Caboni, P.; Scano, P.; Sanchez, S.; Garcia-Barreda, S.; Marco, P. Multi-platform metabolomic approach to discriminate ripening markers of black truffles (Tuber melanosporum). Food Chem. 2020, 319, 126573. [Google Scholar] [CrossRef]
- Patel, S.; Rauf, A.; Khan, H.; Khalid, S.; Mubarak, M.S. Potential health benefits of natural products derived from truffles: A review. Trends Food Sci. Technol. 2018, 70, 1–8. [Google Scholar] [CrossRef]
- Yeh, C.W.; Kan, S.C.; Lin, C.C.; Shieh, C.J.; Liu, Y.C. Polyhydroxylated steroids and triterpenoids from an entophytic fungus, Hypocreales sp. nchu01 isolated from Tuber magnatum. J. Taiwan Inst. Chem. Eng. 2016, 64, 22–30. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Farag, M.A.; Yosri, N.; Sabir, J.S.M.; Saeed, A.; Al-Mousawi, S.M.; Taha, W.; Musharraf, S.G.; Patel, S.; El-Seedi, H.R. Truffles: From Islamic culture to chemistry, pharmacology, and food trends in recent times. J. Taiwan Inst. Chem. Eng. 2019, 91, 193–218. [Google Scholar] [CrossRef]
- Wang, S.N.; Marcone, M.F. The biochemistry and biological properties of the world’s most expensive underground edible mushroom: Truffles. Int. Food Res. J. 2011, 44, 2567–2581. [Google Scholar] [CrossRef]
- Li, J.; Zu, Y.G.; Fu, Y.J.; Yang, Y.C.; Li, S.M.; Li, Z.N.; Wink, M. Optimization of microwave-assisted extraction of triterpene saponins from defatted residue of yellow horn (Xanthoceras sorbifolia bunge.) kernel and evaluation of its antioxidant activity. Innov. Food Sci. Emerg. Technol. 2010, 11, 637–643. [Google Scholar] [CrossRef]
- Xu, L.; Wang, F.; Zhang, Z.C.; Terry, N. Optimization of polysaccharide production from Cordyceps militaris by solid-state fermentation on rice and its antioxidant activities. Foods 2019, 8, 590. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, Y.Z.; Xu, L.; Ma, H.L. An efficient ultrasound-assisted extraction method of pea protein and its effect on protein functional properties and biological activities. LWT-Food Sci. Technol. 2020, 127, 109348. [Google Scholar] [CrossRef]
- Kumkrai, P.; Weeranantanapan, O.; Chudapongse, N. Antioxidant, α-glucosidase inhibitory activity and sub-chronic toxicity of Derris reticulata extract: Its antidiabetic potential. BMC Complement. Altern. Med. 2015, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.N.; Wang, C.L.; Lay, H.L. Effect of nutrition, vitamin, grains, and temperature on the mycelium growth and antioxidant capacity of Cordyceps militaris (strains AG-1 and PSJ-1). J. Radiat. Res. Appl. Sci. 2018, 11, 130–138. [Google Scholar] [CrossRef]
- Feng, J.; Feng, N.; Tang, Q.J.; Liu, Y.F.; Yang, Y.; Liu, F.; Zhang, J.S.; Lin, C.C. Optimization of Ganoderma lucidum polysaccharides fermentation process for large-Scale production. Appl. Biochem. Biotechnol. 2019, 189, 972–986. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Zhong, J.J.; Geng, A. Improvement of ganoderic acid production by fermentation of Ganoderma lucidum with cellulase as an elicitor. Process Biochem. 2014, 49, 1580–1586. [Google Scholar] [CrossRef]
- Liu, G.Q.; Wang, X.L.; Han, W.J.; Lin, Q.L. Improving the fermentation production of the individual key triterpene ganoderic acid me by the medicinal fungus Ganoderma lucidum in submerged culture. Molecules. 2012, 17, 12575–12586. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.C.; Wang, J.; Liu, L.; Sun, Q.S.; Shi, W.J.; Liu, X.C.; Wang, F. The optimum conditions and mechanism for increasing exo-polysaccharide production of Truffles melanosporum by dioscorea saponins. LWT-Food Sci. Technol. 2019, 107, 331–339. [Google Scholar] [CrossRef]
- Zhu, Y.; Ni, R.; Huang, R. Process optimization for the production of diosgenin with Trichoderma reesei. Bioprocess. Biosyst. Eng. 2010, 33, 647–655. [Google Scholar] [CrossRef]
- Xu, C.; Yu, J.; Zhao, S.; Wu, S.; He, P.; Jia, X.; Liu, Y.; Mao, D. Effect of carbon source on production, characterization and bioactivity of exopolysaccharide produced by Phellinus vaninii ljup. An. Acad. Bras. Cienc. 2017, 89, 2033–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Lin, Y.Q.; Yang, C.; Ying, Z.H.; Jiang, X.L. Production of liquid spawn of an edible mushroom, sparassis latifolia by submerged fermentation and mycelial growth on pine wood sawdust. Sci. Hortic. 2016, 209, 22–30. [Google Scholar] [CrossRef]
- Cheng, D.Q.; Ding, Z.S.; Lin, M.A.; Pan, P.L.; Chen, Y.T. Isolation and fermentation culture of fungi from cordyceps soofifera. Am. J. Chin. Med. 2006, 29, 99–101. [Google Scholar]
- Meng, F.; Jia, S.; Bo, Y.; Wu, L.; Yi, W. Screening and optimisation of fermentation media of Marasmius androsaceus using response surface methodology. Oxid. Commun. 2015, 38, 1029–1038. [Google Scholar]
- Tammela, P.; Laitinen, L.; Galkin, A.; Wennberg, T.; Heczko, R.; Vuorela, H. Permeability characteristics and membrane affinity of flavonoids and alkyl gallates in caco-2 cells and in phospholipid vesicles. Arch. Biochem. Biophys. 2014, 425, 193–199. [Google Scholar] [CrossRef]
- Kuang, Y.; Lin, Y.; Li, K.; Song, W.; Ji, S.; Qiao, X. Screening of hepatoprotective compounds from licorice against carbon tetrachloride and acetaminophen induced HepG2 cells injury. Phytomedicine 2017, 34, 59–66. [Google Scholar] [CrossRef]
- Huang, Z.B.; Zhang, L.J.; Wang, Y.L. Effects of rutin and its derivatives on citrinin production by Monascus aurantiacus Li AS3.4384 in liquid fermentation using different types of media. Food Chem. 2019, 284, 205–212. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Shen, W.L.; Liu, D.; Li, J.S. Enhanced production of mycelial biomass and ganoderic acid in submerged culture of Ganoderma applanatum ACCC-52297 elicited by feeding rutin. Afr. J. Microbiol. Res. 2011, 5, 3452–3461. [Google Scholar] [CrossRef]
- Dong, G.L.; Lee, A.Y.; Kim, K.T.; Cho, E.J.; Lee, S. Novel dammarane-type triterpene saponins from Panax ginseng root. Chem. Pharm. Bull. 2016, 63, 927–934. [Google Scholar] [CrossRef]
- Xiong, S.L.; Hou, D.B.; Huang, N.; Li, A.L. Preparation and biological activity of saponin from Ophiopogon japonicus. Afr. J. Pharm. Pharmacol. 2016, 6, 1964–1970. [Google Scholar] [CrossRef]
- He, N.W.; Zhao, Y.; Ling, G.; Shang, J.; Yang, X.B. Antioxidant, antiproliferative, and pro-apoptotic activities of a saponin extract derived from the roots of Panax notoginseng (burk.) f.h. chen. J. Med. Food. 2012, 15, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Miao, Y.; Huang, L.; Li, J.; Sun, H.; Zhao, Y. Antioxidant activities of saponins extracted from Radix trichosanthis: An in vivo and in vitro evaluation. BMC Complement. Altern. Med. 2014, 14, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odeyemi, S.; Afolayan, A.; Bradley, G. In vitro anti-inflammatory and free radical scavenging activities of crude saponins extracted from Albuca bracteata jacq. bulb. Afr. J. Tradit. Complement. Altern. Med. 2015, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Sikandar, A.; Zhang, M.Y.; Wang, Y.Y.; Zhu, X.F.; Liu, X.Y.; Fan, H.Y.; Xuan, Y.H.; Chen, L.J.; Duan, Y.X. Mycochemical screening and analysis, antioxidant activity, and biochemical composition of fermentation strain snef1216 (Penicillium chrysogenum). J. Anal. Methods Chem. 2020, 2020, 3073906. [Google Scholar] [CrossRef]
- Parthasarathi, S.; Hong, S.C.; Oh, M.H.; Park, Y.S.; Pyo, M.K. Anti-oxidant and hepatoprotective effect of white ginsengs in H2O2-treated HepG2 cells. Nat. Prod. Rep. 2015, 21, 210–218. [Google Scholar]
- Jahangir, M.; Ilyas, S.A.; Mazhar, F.; Abbasi, M.A.; Aziz-Ur-Rehman.; Usman, M.F. Antimicrobial and antioxidant potential of Astragalus psilocentros. Asian J. Chem. 2013, 25, 175–180. [Google Scholar] [CrossRef]
- Li, Y.L.; Xin, X.M.; Chang, Z.Y.; Shi, R.J.; Miao, Z.M.; Ding, J.; Hao, G.P. The endophytic fungi of Salvia miltiorrhiza bge.f. alba are a potential source of natural antioxidants. Bot Stud. 2015, 56, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satish, A.; Reddy, P.V.; Sairam, S.; Ahmed, F.; Urooj, A. Antioxidative effect and DNA protecting property of Moringa oleifera root extracts. J. Herb. Med. 2014, 20, 209–220. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Level | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| F1 (g/L) | 20 | 30 | 40 | 50 | 60 |
| F2 (g/L) | 2 | 3 | 4 | 5 | 6 |
| F3 (g/100 mL) | 0.2 | 0.4 | 0.6 | 0.8 | 1 |
| F4 (g/L) | 0.2 | 0.7 | 1.2 | 1.7 | 2.2 |
| F5 (g/L) | 0.2 | 0.7 | 1.2 | 1.7 | 2.2 |
| F6 (g/L) | 0.02 | 0.04 | 0.06 | 0.08 | 0.10 |
| F7 (g/L) | 0.02 | 0.04 | 0.06 | 0.08 | 0.10 |
| F8 (g/L) | 0.02 | 0.04 | 0.06 | 0.08 | 0.10 |
| RUN | F1 | F2 | F3 | F4 | F5 | F6 | F7 | F8 | ETSC |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 30 | 6 | 0.4 | 0.7 | 0.2 | 0.08 | 0.1 | 0.06 | 0.183 |
| 2 | 30 | 2 | 0.2 | 0.2 | 0.7 | 0.04 | 0.06 | 0.02 | 0.105 |
| 3 | 30 | 2 | 1 | 0.2 | 1.2 | 0.08 | 0.04 | 0.1 | 0.042 |
| 4 | 50 | 5 | 0.2 | 1.2 | 0.2 | 0.08 | 0.06 | 0.02 | 0.144 |
| 5 | 60 | 3 | 0.2 | 0.2 | 0.7 | 0.06 | 0.1 | 0.02 | 0.102 |
| 6 | 20 | 2 | 0.2 | 0.2 | 0.2 | 0.02 | 0.02 | 0.02 | 0.040 |
| 7 | 20 | 3 | 0.4 | 2.2 | 0.7 | 0.08 | 0.02 | 0.08 | 0.084 |
| 8 | 20 | 3 | 0.8 | 0.7 | 1.2 | 0.02 | 0.06 | 0.06 | 0.037 |
| 9 | 40 | 3 | 1 | 0.7 | 0.2 | 0.02 | 0.02 | 0.02 | 0.049 |
| 10 | 30 | 6 | 0.2 | 1.7 | 0.7 | 0.02 | 0.04 | 0.04 | 0.126 |
| 11 | 20 | 3 | 0.2 | 0.2 | 0.2 | 0.04 | 0.04 | 0.08 | 0.042 |
| 12 | 30 | 2 | 0.6 | 0.7 | 2.2 | 0.06 | 0.06 | 0.08 | 0.101 |
| 13 | 20 | 2 | 0.6 | 0.7 | 0.2 | 0.04 | 0.1 | 0.04 | 0.032 |
| 14 | 30 | 3 | 1 | 1.7 | 2.2 | 0.04 | 0.08 | 0.02 | 0.042 |
| 15 | 30 | 3 | 0.4 | 0.2 | 0.2 | 0.02 | 0.02 | 0.04 | 0.079 |
| 16 | 50 | 4 | 0.4 | 0.2 | 2.2 | 0.02 | 0.1 | 0.1 | 0.125 |
| 17 | 30 | 4 | 0.2 | 1.2 | 0.7 | 0.04 | 0.02 | 0.06 | 0.190 |
| 18 | 50 | 6 | 1 | 0.7 | 0.7 | 0.04 | 0.02 | 0.08 | 0.142 |
| 19 | 20 | 2 | 0.4 | 0.7 | 0.7 | 0.02 | 0.04 | 0.02 | 0.046 |
| 20 | 40 | 5 | 0.2 | 1.7 | 1.2 | 0.02 | 0.1 | 0.08 | 0.123 |
| 21 | 60 | 6 | 0.6 | 0.2 | 1.2 | 0.1 | 0.02 | 0.02 | 0.176 |
| 22 | 60 | 4 | 1 | 2.2 | 0.2 | 0.02 | 0.06 | 0.04 | 0.104 |
| 23 | 20 | 6 | 0.8 | 1.2 | 2.2 | 0.02 | 0.04 | 0.02 | 0.109 |
| 24 | 50 | 2 | 0.4 | 2.2 | 1.2 | 0.04 | 0.04 | 0.02 | 0.086 |
| 25 | 60 | 3 | 0.2 | 0.7 | 2.2 | 0.08 | 0.04 | 0.04 | 0.070 |
| 26 | 40 | 2 | 0.8 | 0.2 | 0.7 | 0.08 | 0.08 | 0.04 | 0.054 |
| 27 | 20 | 2 | 1 | 1.2 | 0.7 | 0.1 | 0.1 | 0.04 | 0.055 |
| 28 | 40 | 2 | 0.2 | 2.2 | 2.2 | 0.1 | 0.02 | 0.06 | 0.071 |
| 29 | 50 | 3 | 0.2 | 0.7 | 1.7 | 0.1 | 0.04 | 0.04 | 0.103 |
| 30 | 30 | 5 | 0.6 | 2.2 | 0.7 | 0.02 | 0.04 | 0.04 | 0.109 |
| 31 | 40 | 6 | 0.4 | 0.2 | 1.7 | 0.04 | 0.06 | 0.04 | 0.148 |
| 32 | 20 | 3 | 0.4 | 1.7 | 0.7 | 0.1 | 0.06 | 0.1 | 0.055 |
| 33 | 20 | 4 | 0.6 | 1.7 | 1.7 | 0.08 | 0.02 | 0.02 | 0.072 |
| 34 | 30 | 5 | 0.4 | 0.7 | 0.2 | 0.1 | 0.08 | 0.02 | 0.166 |
| 35 | 30 | 4 | 0.8 | 0.2 | 0.2 | 0.1 | 0.04 | 0.08 | 0.115 |
| 36 | 30 | 3 | 0.4 | 1.2 | 1.2 | 0.06 | 0.02 | 0.04 | 0.076 |
| 37 | 40 | 3 | 0.6 | 1.2 | 0.2 | 0.04 | 0.04 | 0.1 | 0.139 |
| 38 | 60 | 2 | 0.4 | 1.2 | 1.7 | 0.02 | 0.08 | 0.08 | 0.074 |
| 39 | 60 | 5 | 0.8 | 0.7 | 0.7 | 0.04 | 0.02 | 0.1 | 0.156 |
| 40 | 30 | 2 | 0.2 | 0.7 | 1.7 | 0.02 | 0.02 | 0.1 | 0.043 |
| 41 | 50 | 3 | 0.6 | 0.2 | 0.7 | 0.02 | 0.08 | 0.06 | 0.137 |
| 42 | 60 | 2 | 0.4 | 1.7 | 0.2 | 0.04 | 0.04 | 0.06 | 0.061 |
| 43 | 20 | 6 | 0.2 | 2.2 | 0.2 | 0.06 | 0.08 | 0.1 | 0.135 |
| 44 | 20 | 5 | 0.4 | 0.2 | 2.2 | 0.04 | 0.02 | 0.04 | 0.069 |
| 45 | 30 | 3 | 0.8 | 2.2 | 1.7 | 0.04 | 0.1 | 0.02 | 0.088 |
| 46 | 40 | 4 | 0.4 | 0.7 | 0.7 | 0.06 | 0.04 | 0.02 | 0.162 |
| 47 | 20 | 5 | 1 | 0.2 | 1.7 | 0.06 | 0.04 | 0.06 | 0.063 |
| 48 | 50 | 2 | 0.8 | 1.7 | 0.2 | 0.06 | 0.02 | 0.04 | 0.097 |
| 49 | 20 | 4 | 0.2 | 0.7 | 1.2 | 0.04 | 0.08 | 0.04 | 0.067 |
| Factor | Level | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |
| F1(v/v, %) | 4 | 7 | 10 | 13 | 16 |
| F2 (°C) | 20 | 22 | 24 | 26 | 28 |
| F3 (rpm) | 70 | 100 | 130 | 160 | 190 |
| F4 (mL) | 50 | 70 | 90 | 110 | 130 |
| F5 | 5.0 | 5.5 | 6.0 | 6.5 | 7.0 |
| F6 (h) | 96 | 120 | 144 | 168 | 192 |
| RUN | F1 | F2 | F3 | F4 | F5 | F6 | ETSC |
|---|---|---|---|---|---|---|---|
| 1 | 4 | 24 | 100 | 110 | 6 | 5 | 0.147 |
| 2 | 16 | 24 | 190 | 50 | 7 | 7 | 0.102 |
| 3 | 4 | 20 | 70 | 50 | 5 | 4 | 0.219 |
| 4 | 7 | 28 | 190 | 130 | 6 | 4 | 0.144 |
| 5 | 4 | 26 | 190 | 90 | 6.5 | 8 | 0.092 |
| 6 | 13 | 24 | 130 | 90 | 5.5 | 4 | 0.255 |
| 7 | 16 | 26 | 130 | 130 | 5 | 5 | 0.196 |
| 8 | 13 | 20 | 100 | 130 | 7 | 8 | 0.201 |
| 9 | 7 | 24 | 160 | 70 | 5 | 8 | 0.088 |
| 10 | 10 | 28 | 100 | 90 | 5 | 7 | 0.096 |
| 11 | 10 | 24 | 70 | 130 | 6.5 | 6 | 0.226 |
| 12 | 4 | 28 | 130 | 70 | 7 | 6 | 0.125 |
| 13 | 13 | 28 | 160 | 50 | 6.5 | 5 | 0.126 |
| 14 | 16 | 28 | 70 | 110 | 5.5 | 8 | 0.148 |
| 15 | 16 | 22 | 100 | 70 | 6.5 | 4 | 0.181 |
| 16 | 13 | 22 | 190 | 110 | 5 | 6 | 0.115 |
| 17 | 7 | 22 | 70 | 90 | 7 | 5 | 0.210 |
| 18 | 16 | 20 | 160 | 90 | 6 | 6 | 0.117 |
| 19 | 10 | 26 | 160 | 110 | 7 | 4 | 0.291 |
| 20 | 4 | 22 | 160 | 130 | 5.5 | 7 | 0.096 |
| 21 | 13 | 26 | 70 | 70 | 6 | 7 | 0.163 |
| 22 | 10 | 22 | 130 | 50 | 6 | 8 | 0.096 |
| 23 | 10 | 20 | 190 | 70 | 5.5 | 5 | 0.104 |
| 24 | 7 | 20 | 130 | 110 | 6.5 | 7 | 0.115 |
| 25 | 7 | 26 | 100 | 50 | 5.5 | 6 | 0.091 |
| Item | Coefficient | t | Significance |
|---|---|---|---|
| (Constant) | −15.222 | –2.768 | 0.009 |
| F12 | –0.008 | –3.384 | 0.002 |
| F1 | 0.696 | 3.973 | 0.000 |
| F22 | –0.041 | –0.186 | 0.853 |
| F2 | 2.372 | 1.353 | 0.185 |
| F32 | –12.521 | –2.257 | 0.031 |
| F3 | 12.082 | 1.837 | 0.075 |
| F42 | –0.731 | –0.823 | 0.417 |
| F4 | 1.674 | 0.793 | 0.433 |
| F52 | –0.448 | –0.504 | 0.617 |
| F5 | –0.050 | –0.024 | 0.981 |
| F62 | –394.311 | –0.711 | 0.482 |
| F6 | 60.102 | 0.990 | 0.330 |
| F72 | 237.132 | 0.427 | 0.672 |
| F7 | –20.234 | –0.308 | 0.760 |
| F82 | 304.787 | 0.549 | 0.587 |
| F8 | –29.144 | –0.443 | 0.661 |
| Model: R2 = 0.777; Adj-R2 = 0.665; F-value = 6.959; p = 0.000 | |||
| Item | Coefficient | t | Significance |
|---|---|---|---|
| (Constant) | 79.136 | 1.062 | 0.309 |
| F12 | –0.045 | –1.125 | 0.283 |
| F1 | 1.132 | 1.391 | 0.189 |
| F22 | –0.134 | –1.492 | 0.162 |
| F2 | 6.354 | 1.467 | 0.168 |
| F32 | 0.00006566 | 0.164 | 0.873 |
| F3 | –0.071 | –0.678 | 0.511 |
| F42 | –0.00009911 | –0.110 | 0.914 |
| F4 | 0.0079 | 0.484 | 0.637 |
| F52 | 2.949 | 2.045 | 0.063 |
| F5 | –33.483 | –1.932 | 0.077 |
| F62 | 1.042 | 2.891 | 0.014 |
| F6 | –14.784 | –3.401 | 0.005 |
| Model: R2 = 0.862; Adj-R2 = 0.725; F-value = 6.266; p = 0.002 | |||
| Source | Preparation Method | Composition | Concentration (mg/mL) | DPPH· (%) | ·OH (%) | FRAP (mM) | Reference |
|---|---|---|---|---|---|---|---|
| Tuber melanosporum broth | Extraction | TS mixture | 3 | 84.00 | 57.97 | 26.44 | This study |
| Tuber melanosporum broth | Extraction | RTS mixture | 3 | 94.13 | 68.75 | 30.28 | This study |
| Panax ginseng root | Extraction and purification | Ginsenoside-Rg18 | 0.048 | 8.76 | 87.70 | -- | [42] |
| 6-acetyl ginsenoside-Rg3 | 0.057 | 4.58 | 87.55 | -- | |||
| Ginsenoside-Rs11 | 0.057 | 7.37 | 90.74 | -- | |||
| Ginsenoside-Re7 | 0.049 | 2.19 | 89.48 | -- | |||
| Ophiopogon japonicus | Extraction and purification | Saponin mixture | 3 | 94.62 | 83 | -- | [43] |
| Panax notoginseng | Extraction | Saponin mixture | 5 | 84.7 | 50 | -- | [44] |
| Radix trichosanthis | Extraction | Saponin mixture | 3 | 70 | 3 | -- | [45] |
| Albuca bracteata Jacq. Bulb | Extraction | Saponin mixture | 4 | 87 | -- | [46] | |
| Penicillium chrysogenum broth | Extraction | Snef1216 | 0.5 | 63.86 | -- | [47] | |
| White ginseng | Extraction | Saponin mixture | 1 | 65 | 18.3 | -- | [48] |
| Panax ginseng root | Extraction | Saponin mixture AR-5R | 0.2 | -- | -- | 17.42 | [42] |
| Saponin mixture AR-4M | 0.2 | -- | -- | 13.91 | |||
| Astragalus psilocentros | Extraction | Astragaloside mixture | 2.5 | -- | 43.76 | [49] | |
| Alternaria alternata broth | Extraction | Saponin mixture SaF-2 | 0.5 | 83.25% | -- | 1.65 | [50] |
| Fusarium proliferatum broth | Extraction | Saponin mixture SaR-2 | 0.5 | 90.14% | -- | 1.68 | [50] |
| Schizophyllum commune broth | Extraction | Saponin mixture SaR-3 | 0.5 | -- | -- | 0.58 | [50] |
| Trametes hirsuta broth | Extraction | Saponin mixture SaR-6 | 0.5 | -- | -- | 0.51 | [50] |
| Salvia miltiorrhiza Bge root | Extraction | Saponin mixture | 0.5 | 80.23% | -- | 1.34 | [50] |
| Moringa oleifera root | Extraction | Saponin mixture | 0.37 | -- | -- | 0.87 | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, L.; Su, Q.; Zhang, Z.; Guan, G.; Huang, D.; Terry, N.; Wang, F. Increased Extracellular Saponin Production after the Addition of Rutin in Truffle Liquid Fermentation and Its Antioxidant Activities. Fermentation 2021, 7, 103. https://doi.org/10.3390/fermentation7030103
Xu L, Su Q, Zhang Z, Guan G, Huang D, Terry N, Wang F. Increased Extracellular Saponin Production after the Addition of Rutin in Truffle Liquid Fermentation and Its Antioxidant Activities. Fermentation. 2021; 7(3):103. https://doi.org/10.3390/fermentation7030103
Chicago/Turabian StyleXu, Ling, Qihui Su, Zhicai Zhang, Guoqiang Guan, Daming Huang, Norman Terry, and Feng Wang. 2021. "Increased Extracellular Saponin Production after the Addition of Rutin in Truffle Liquid Fermentation and Its Antioxidant Activities" Fermentation 7, no. 3: 103. https://doi.org/10.3390/fermentation7030103
APA StyleXu, L., Su, Q., Zhang, Z., Guan, G., Huang, D., Terry, N., & Wang, F. (2021). Increased Extracellular Saponin Production after the Addition of Rutin in Truffle Liquid Fermentation and Its Antioxidant Activities. Fermentation, 7(3), 103. https://doi.org/10.3390/fermentation7030103