

Improved Sugar Recovery of Alkaline Pre-Treated Pineapple Leaf Fibres via Enzymatic Hydrolysis and Its Enzymatic Kinetics



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation Process
2.2. Alkaline Pre-Treatment Process
2.3. Optimisation of Enzymatic Hydrolysis Operating Parameters
2.4. Kinetics Study of Enzymatic Reactions
2.5. Quantification of Total Reducing Sugar (TRS)
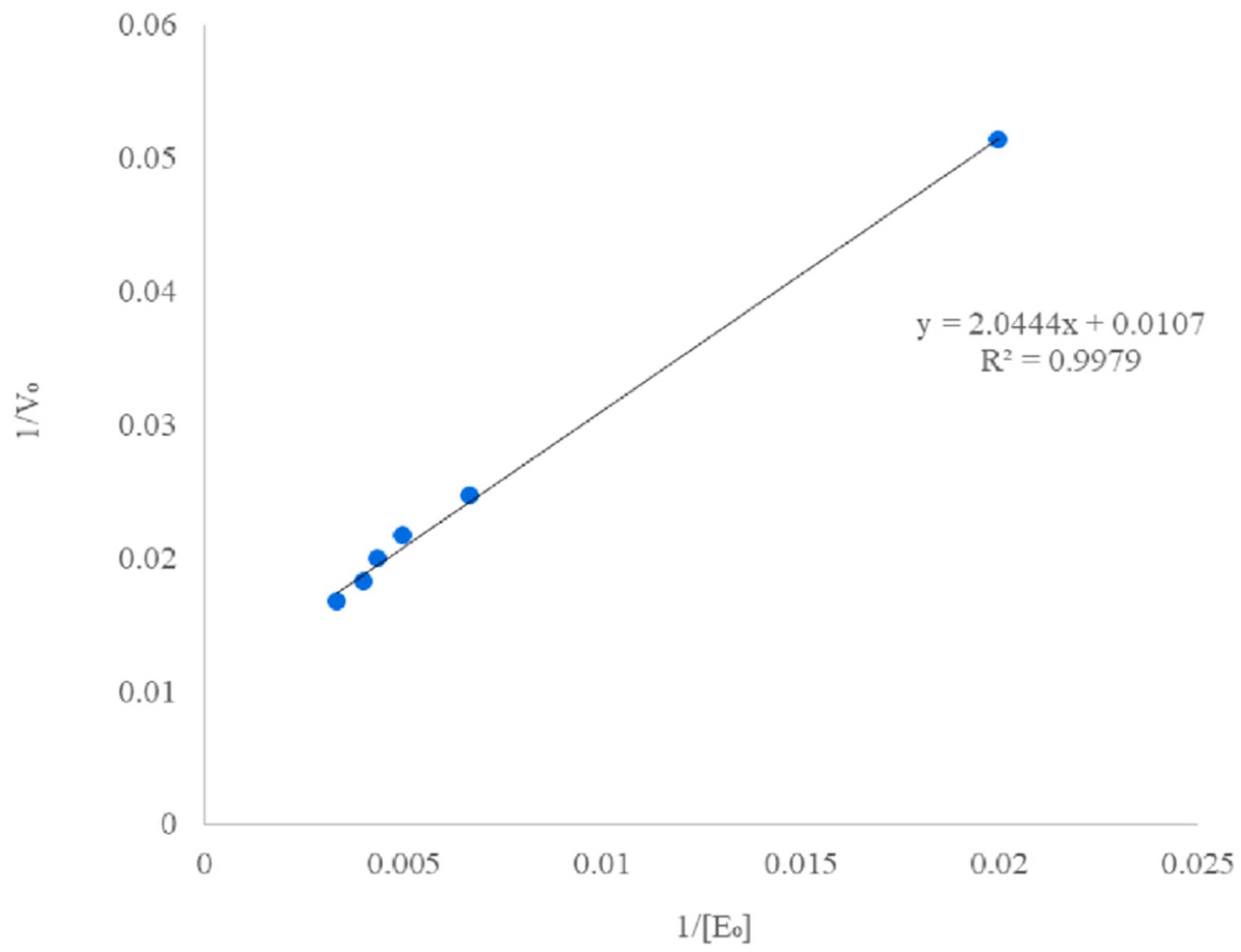
2.6. Kinetics Modelling: Estimation of the Parameters
3. Results
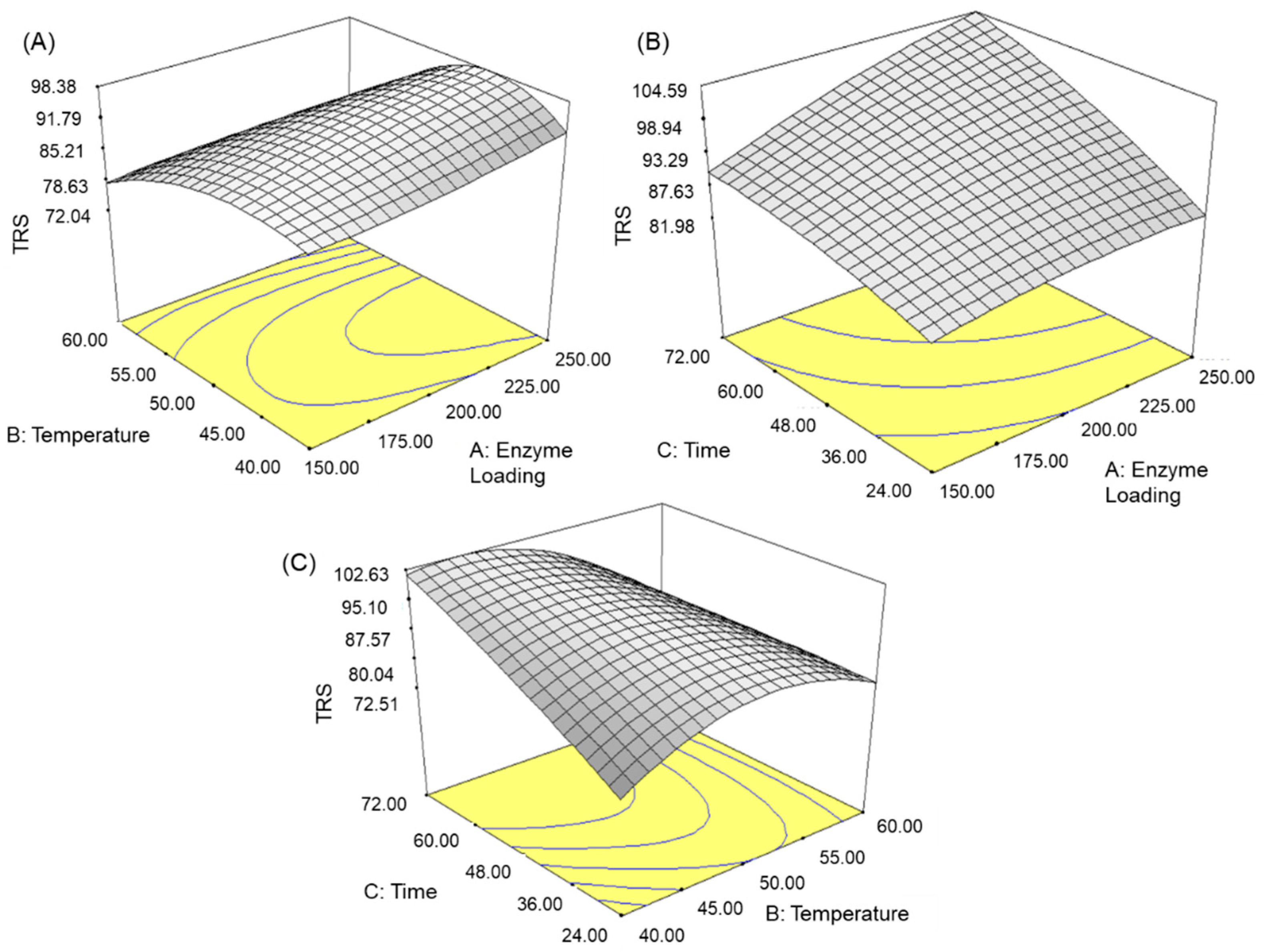
3.1. Optimisation of the Enzymatic Hydrolysis Process
3.2. ANOVA and Statistical Data Analysis
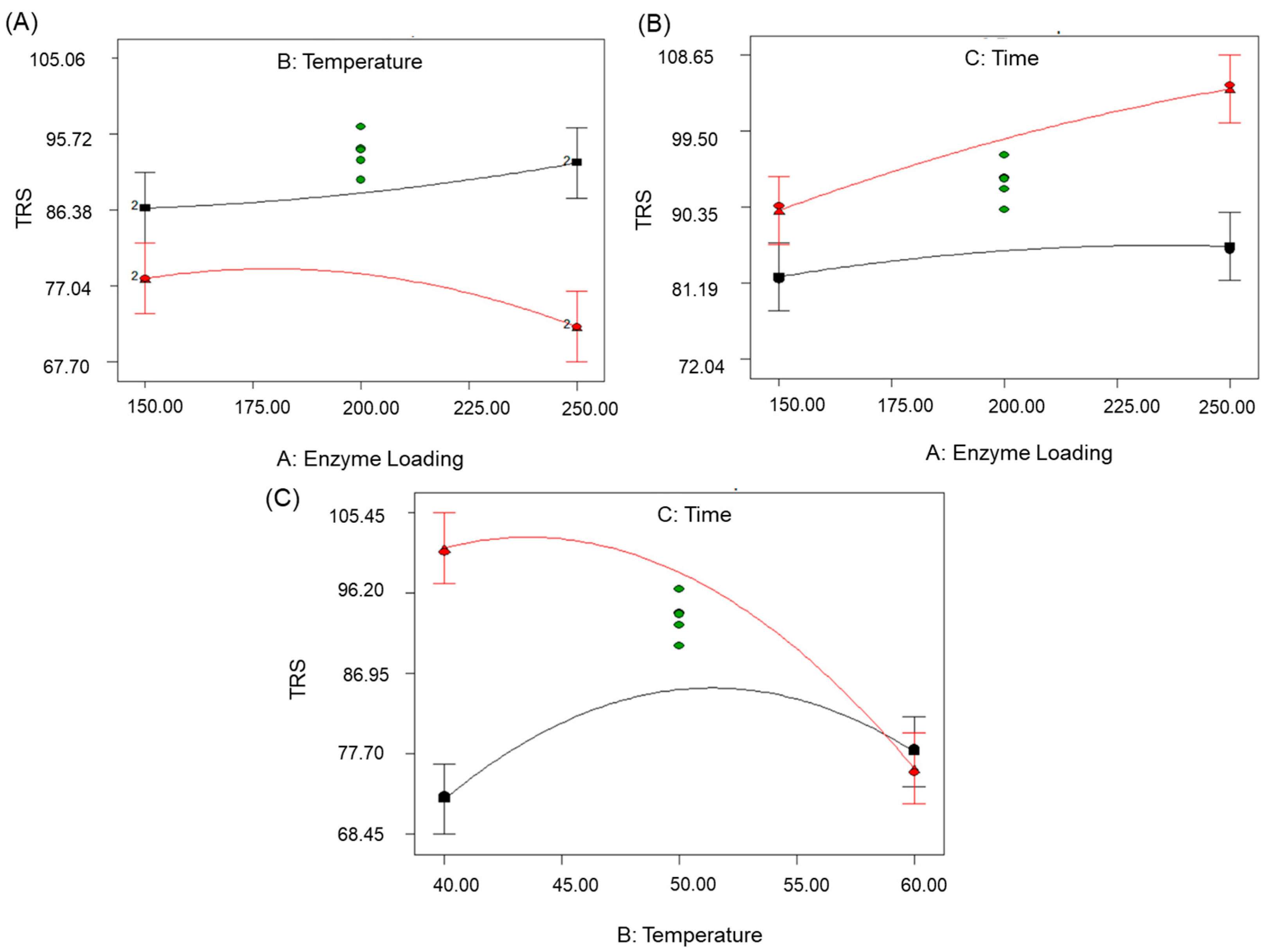
3.3. Effect of Variables on TRS Yield
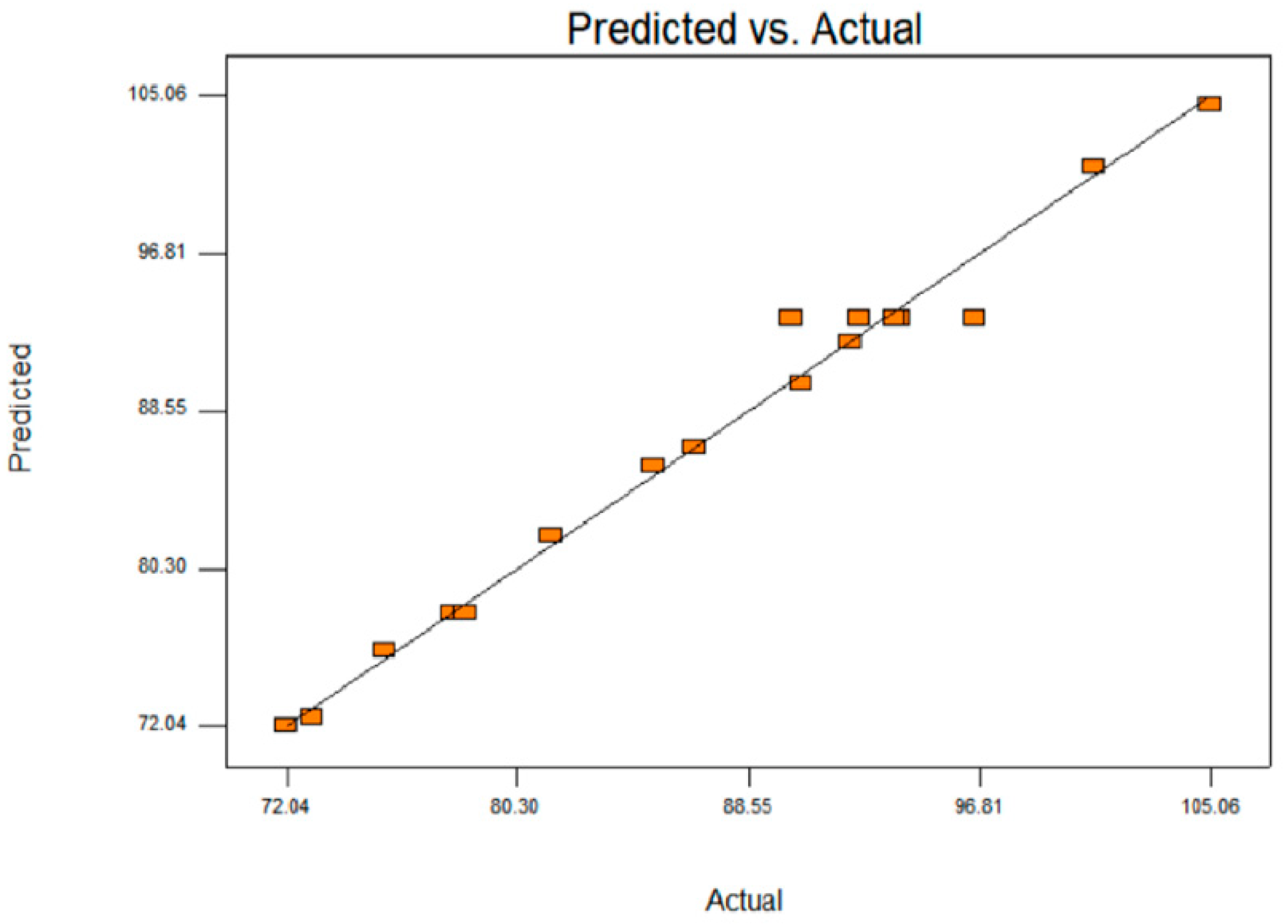
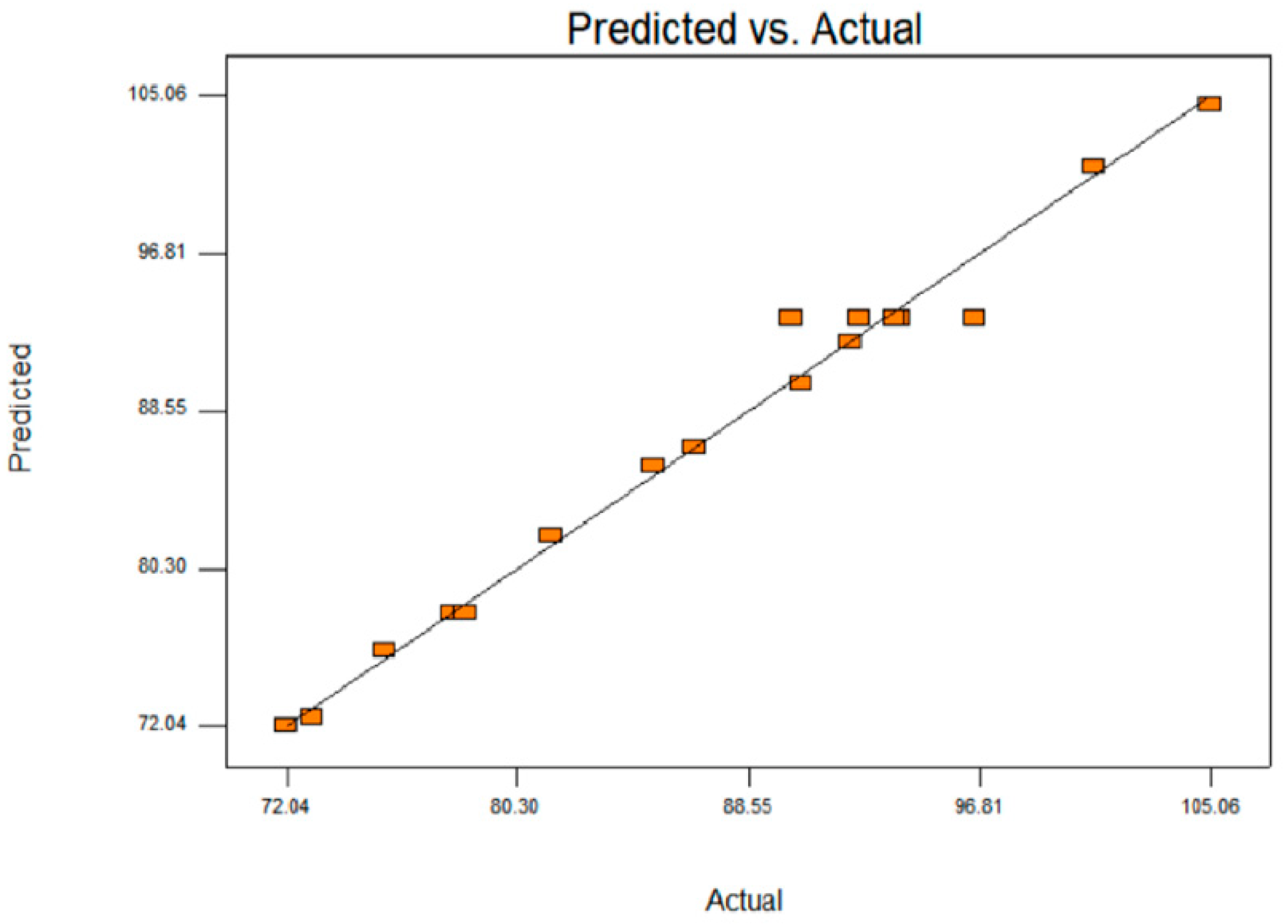
3.4. Validation of Optimisation Conditions
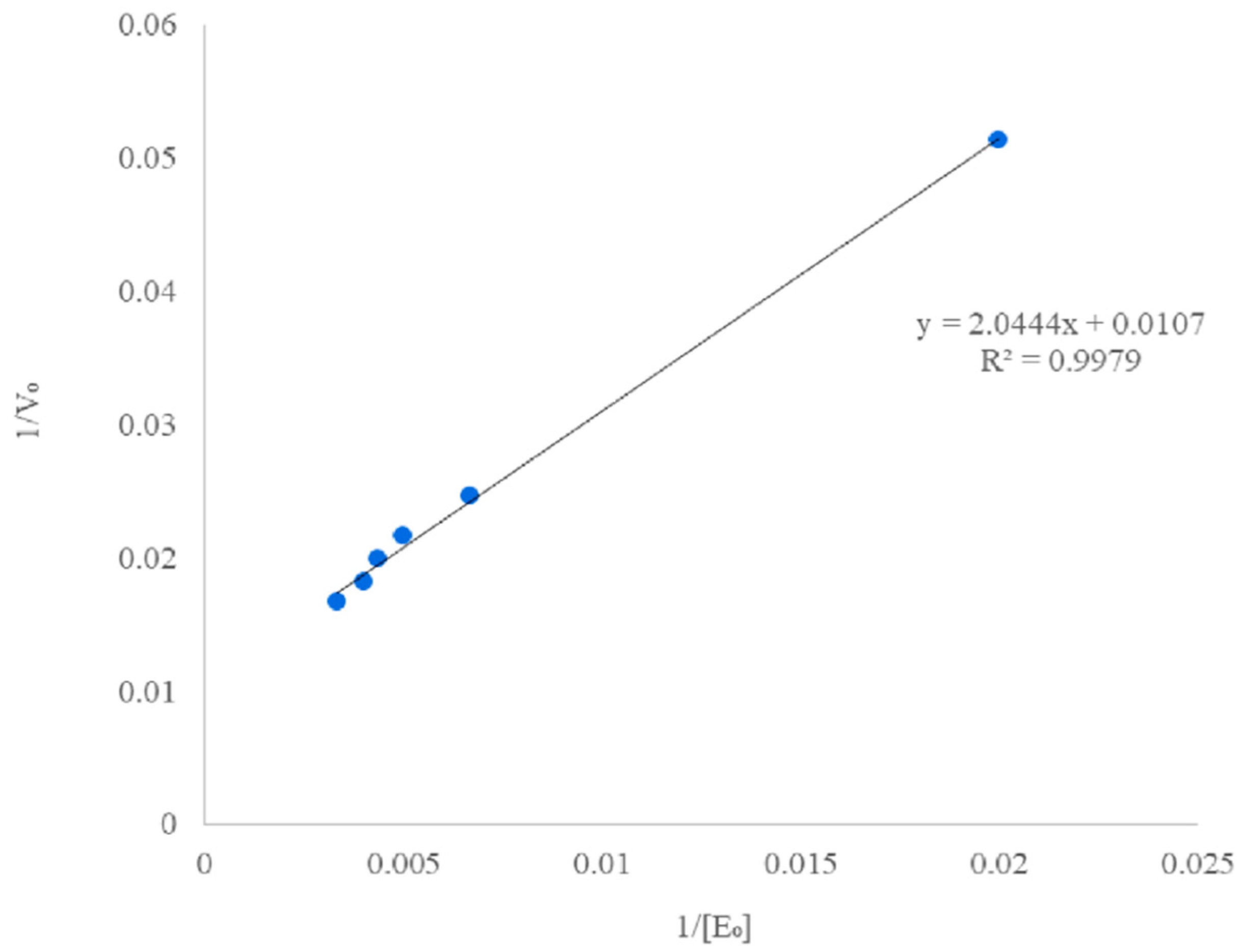
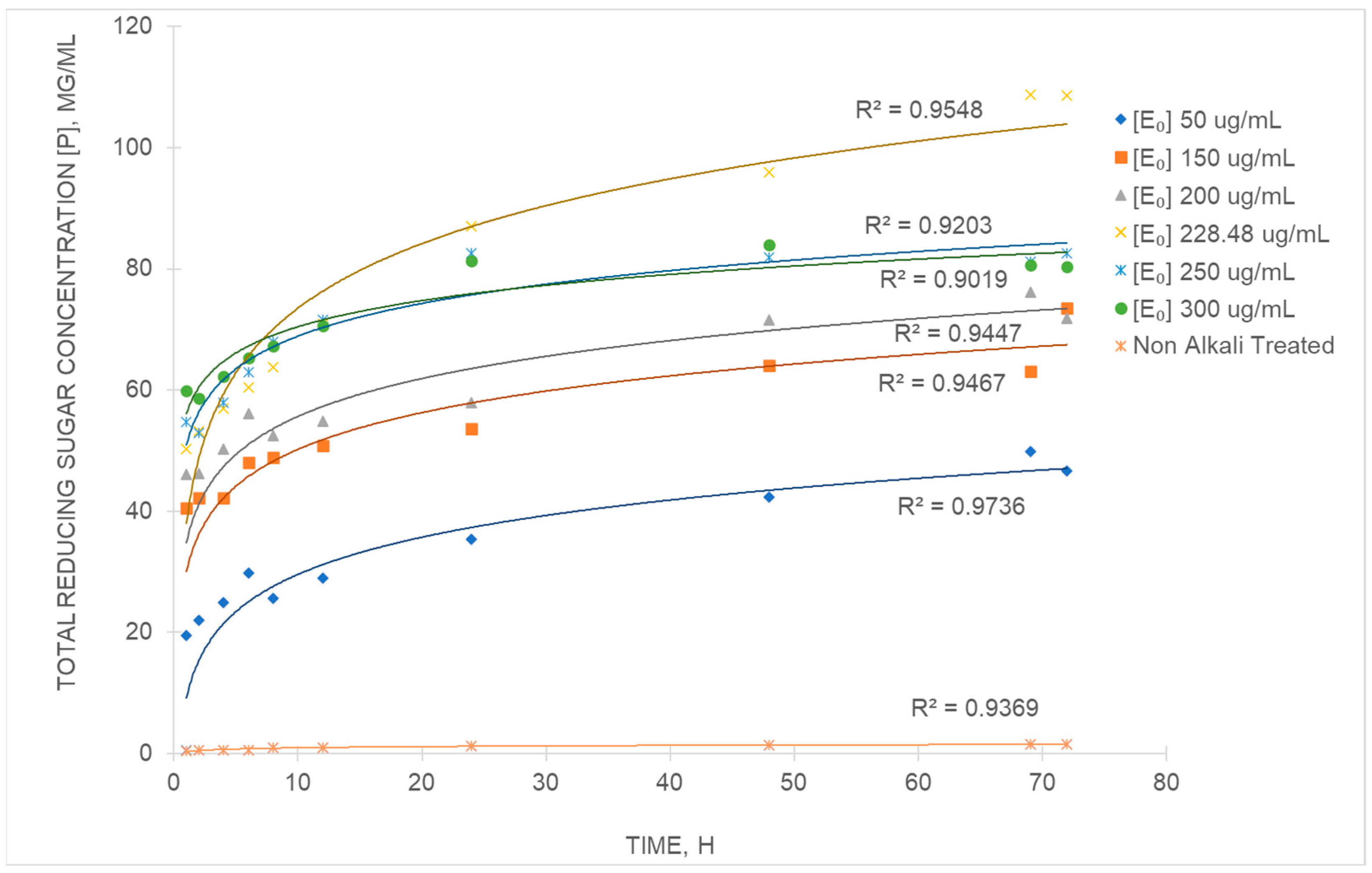
3.5. Kinetic Study of the Enzymatic Hydrolysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. Crop and Livestocks Product; FAO: Rome, Italy, 2021. [Google Scholar]
- El-Zawawy, W.K.; Ibrahim, M.M.; Abdel-Fattah, Y.R.; Soliman, N.A.; Mahmoud, M.M. Acid and Enzyme Hydrolysis to Convert Pretreated Lignocellulosic Materials into Glucose for Ethanol Production. Carbohydr. Polym. 2011, 84, 865–871. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, J. Hydrolysis of Lignocellulosic Materials for Ethanol Production: A Review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Taher, I.B.; Fickers, P.; Chniti, S.; Hassouna, M. Optimization of Enzymatic Hydrolysis and Fermentation Conditions for Improved Bioethanol Production from Potato Peel Residues. Biotechnol. Prog. 2017, 33, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Umagiliyage, A.L.; Choudhary, R.; Liang, Y.; Haddock, J.; Watson, D.G. Laboratory Scale Optimization of Alkali Pretreatment for Improving Enzymatic Hydrolysis of Sweet Sorghum Bagasse. Ind. Crops Prod. 2015, 74, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.; Bishnoi, N.R. Enzymatic Hydrolysis Optimization of Microwave Alkali Pretreated Wheat Straw and Ethanol Production by Yeast. Bioresour. Technol. 2012, 108, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Chen, X.; Shen, F.; Su, Y.; Wan, Y. Optimization of Enzymatic Hydrolysis of Wheat Straw Pretreated by Alkaline Peroxide Using Response Surface Methodology. Ind. Eng. Chem. Res. 2009, 48, 7346–7353. [Google Scholar] [CrossRef]
- Maitan-Alfenas, G.P.; Visser, E.M.; Guimarães, V.M. Enzymatic Hydrolysis of Lignocellulosic Biomass: Converting Food Waste in Valuable Products. Curr. Opin. Food Sci. 2015, 1, 44–49. [Google Scholar] [CrossRef]
- Ayyachamy, M.; Gupta, V.K.; Cliffe, F.E.; Tuohy, M.G. Enzymatic Saccharification of Lignocellulosic Biomass BT-Laboratory Protocols in Fungal Biology: Current Methods in Fungal Biology; Gupta, V.K., Tuohy, M.G., Ayyachamy, M., Turner, K.M., O’Donovan, A., Eds.; Springer: New York, NY, USA, 2013; pp. 475–481. [Google Scholar]
- Ferreira, S.; Duarte, A.P.; Ribeiro, M.H.L.; Queiroz, J.A.; Domingues, F.C. Response Surface Optimization of Enzymatic Hydrolysis of Cistus Ladanifer and Cytisus Striatus for Bioethanol Production. Biochem. Eng. J. 2009, 45, 192–200. [Google Scholar] [CrossRef]
- Kaur, S.; Sarkar, B.C.; Sharma, H.K.; Singh, C. Optimization of Enzymatic Hydrolysis Pretreatment Conditions for Enhanced Juice Recovery from Guava Fruit Using Response Surface Methodology. Food Bioproc. Tech. 2009, 2, 96–100. [Google Scholar] [CrossRef]
- Pandiyan, K.; Tiwari, R.; Singh, S.; Nain, P.K.S.; Rana, S.; Arora, A.; Singh, S.B.; Nain, L. Optimization of Enzymatic Saccharification of Alkali Pretreated Parthenium Sp. Using Response Surface Methodology. Enzym. Res. 2014, 2014, 764898. [Google Scholar] [CrossRef]
- Ruangmee, A.; Sangwichien, C. Response Surface Optimization of Enzymatic Hydrolysis of Narrow-Leaf Cattail for Bioethanol Production. Energy Convers. Manag 2013, 73, 381–388. [Google Scholar] [CrossRef]
- Saini, J.K.; Anurag, R.K.; Arya, A.; Kumbhar, B.K.; Tewari, L. Optimization of Saccharification of Sweet Sorghum Bagasse Using Response Surface Methodology. Ind. Crops Prod. 2013, 44, 211–219. [Google Scholar] [CrossRef]
- Carvalho, M.L.; Sousa, R.; Rodríguez-Zúñiga, U.F.; Suarez, C.A.G.; Rodrigues, D.S.; Giordano, R.C.; Giordano, R.L.C. Kinetic Study of the Enzymatic Hydrolysis of Sugarcane Bagasse. Braz. J. Chem. Eng. 2013, 30, 437–447. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.J. Enzyme Kinetics of Cellulose Hydrolysis. Biochem. J. 1989, 262, 1001. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, F.; Lis, M.J.; Colom, X.; López-Mesas, M.; Valldeperas, J. Effect of Alkali Pretreatment on Cellulase Hydrolysis of Wheat Straw: Kinetic Study. Process. Biochem. 2005, 40, 3360–3364. [Google Scholar] [CrossRef]
- Pratto, B.; de Souza, R.B.A.; Sousa, R.; da Cruz, A.J.G. Enzymatic Hydrolysis of Pretreated Sugarcane Straw: Kinetic Study and Semi-Mechanistic Modeling. Appl. Biochem. Biotechnol. 2016, 178, 1430–1444. [Google Scholar] [CrossRef]
- Kadam, K.L.; Rydholm, E.C.; McMillan, J.D. Development and Validation of a Kinetic Model for Enzymatic Saccharification of Lignocellulosic Biomass. Biotechnol. Prog. 2004, 20, 698–705. [Google Scholar] [CrossRef]
- Lin, S.-H.; Chou, L.-M.; Chien, Y.-W.; Chang, J.-S.; Lin, C.-I. Prebiotic Effects of Xylooligosaccharides on the Improvement of Microbiota Balance in Human Subjects. Gastroenterol. Res. Pract 2016, 2016, 5789232. [Google Scholar] [CrossRef] [Green Version]
- Sajjanshetty, R.; Kulkarni, N.S.; Shankar, K.; Jayalakshmi, S.K.; Sreeramulu, K. Enhanced Production and In-Situ Removal of Butanol during the Fermentation of Lignocellulosic Hydrolysate of Pineapple Leaves. Ind. Crops Prod. 2021, 173, 114147. [Google Scholar] [CrossRef]
- Nashiruddin, N.I.; Mansor, A.F.; Rahman, R.A.; Ilias, R.M.; Yussof, H.W. Process Parameter Optimization of Pretreated Pineapple Leaves Fiber for Enhancement of Sugar Recovery. Ind. Crops Prod. 2020, 152, 112514. [Google Scholar] [CrossRef]
- Ghose, T.K. Measurement of Cellulase Activities. Methods Mol. Biol. 1987, 59, 257–268. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J. Statistical Optimization of Sodium Hydroxide Pretreatment and Enzymatic Hydrolysis of Corn Stover Powder for Enhancing Sugar Production Using Response Surface Methodology. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Q. Response Surface Optimization of Enzymatic Hydrolysis Process of Wet Oxidation Pretreated Wood Pulp Waste. Cellul. Chem. Technol. 2016, 50, 803–809. [Google Scholar]
- Lee, Y.H.; Fan, L.T. Kinetic Studies of Enzymatic Hydrolysis of Insoluble Cellulose: (II). Analysis of Extended Hydrolysis Times. Biotechnol. Bioeng. 1983, 25, 939–966. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Fan, L.T. Kinetic Studies of Enzymatic Hydrolysis of Insoluble Cellulose: Analysis of the Initial Rates. Biotechnol. Bioeng. 1982, 24, 2383–2406. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Coded Name | −1 | 0 | 1 |
|---|---|---|---|---|
| Enzyme loading (µg/g) | X1 | 150 | 200 | 250 |
| Temperature (°C) | X2 | 40 | 50 | 60 |
| Time (h) | X3 | 24 | 48 | 72 |
| Source | Sum of Squares | DF | Mean Square | F Value | Prob > F | |
|---|---|---|---|---|---|---|
| Model | 1498.26 | 11 | 136.21 | 27.82 | 0.0009 | significant |
| X1-Enzyme loading | 83.45 | 1 | 83.45 | 17.04 | 0.0091 | |
| X2-Temperature | 99.00 | 1 | 99.00 | 20.22 | 0.0064 | |
| X3-Time | 363.02 | 1 | 363.02 | 74.15 | 0.0003 | |
| X12 | 7.14 | 1 | 7.14 | 1.46 | 0.2811 | |
| X22 | 411.01 | 1 | 411.01 | 83.95 | 0.0003 | |
| X32 | 10.12 | 1 | 10.12 | 2.07 | 0.2101 | |
| X1X2 | 33.12 | 1 | 33.12 | 6.76 | 0.0482 | |
| X1X3 | 29.98 | 1 | 29.98 | 6.12 | 0.0562 | |
| X2X3 | 237.16 | 1 | 237.16 | 48.44 | 0.0009 | |
| X12X2 | 9.92 | 1 | 9.92 | 2.03 | 0.2138 | |
| X1X22 | 43.52 | 1 | 43.52 | 8.89 | 0.0307 | |
| Residual | 24.48 | 5 | 4.90 | |||
| Lack of fit | 1.78 | 1 | 1.78 | 0.31 | 0.6057 | Not significant |
| Pure error | 22.70 | 4 | 5.68 |
| Cycle | Variables | Response | ||
|---|---|---|---|---|
| Enzyme Loading (µg/g) | Temperature (°C) | Time (h) | TRS (mg/mL) | |
| 1 | 200 (0) | 50 (0) | 48 (0) | 93.78 |
| 2 | 200 (0) | 60 (1) | 72 (1) | 75.56 |
| 3 | 250 (1) | 60 (1) | 48 (0) | 72.04 |
| 4 | 150 (−1) | 50 (0) | 72 (1) | 90.45 |
| 5 | 150 (−1) | 40 (−1) | 48 (0) | 86.64 |
| 6 | 150 (−1) | 50 (0) | 24 (−1) | 81.51 |
| 7 | 200 (0) | 60 (1) | 24 (−1) | 78.43 |
| 8 | 200 (0) | 50 (0) | 48 (0) | 90.10 |
| 9 | 200 (0) | 40 (−1) | 24 (−1) | 72.98 |
| 10 | 250 (1) | 50 (0) | 24 (−1) | 85.17 |
| 11 | 150 (−1) | 60 (1) | 48 (0) | 77.99 |
| 12 | 250 (1) | 40 (−1) | 48 (0) | 92.20 |
| 13 | 200 (0) | 40 (−1) | 72 (1) | 100.91 |
| 14 | 200 (0) | 50 (0) | 48 (0) | 93.96 |
| 15 | 250 (1) | 50 (0) | 72 (1) | 105.06 |
| 16 | 200 (0) | 50 (0) | 48 (0) | 92.51 |
| 17 | 200 (0) | 50 (0) | 48 (0) | 96.65 |
| Initial Enzyme Concentration, [Eo] (µg/g) | k (mg/mL.h) | n | R2 |
|---|---|---|---|
| PALFs pre-treated with NaOH 0.5 | 1.2 × 10−2 | 0.79 | 0.9736 |
| 15.0 | 8.0 × 10−3 | 0.62 | 0.9467 |
| 20.0 | 6.13 × 10−3 | 0.51 | 0.9447 |
| 22.9 | 5.77 × 10−3 | 0.41 | 0.9548 |
| 25.0 | 5.34 × 10−3 | 0.39 | 0.9203 |
| 30.0 | 4.98 × 10−4 | 0.31 | 0.9019 |
| Untreated PALF 22.9 | 1.30 × 10−3 | 0.35 | 0.9369 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nashiruddin, N.I.; Abd Rahman, N.H.; A. Rahman, R.; Md. Illias, R.; Ghazali, N.F.; Abomoelak, B.; El Enshasy, H.A. Improved Sugar Recovery of Alkaline Pre-Treated Pineapple Leaf Fibres via Enzymatic Hydrolysis and Its Enzymatic Kinetics. Fermentation 2022, 8, 640. https://doi.org/10.3390/fermentation8110640
Nashiruddin NI, Abd Rahman NH, A. Rahman R, Md. Illias R, Ghazali NF, Abomoelak B, El Enshasy HA. Improved Sugar Recovery of Alkaline Pre-Treated Pineapple Leaf Fibres via Enzymatic Hydrolysis and Its Enzymatic Kinetics. Fermentation. 2022; 8(11):640. https://doi.org/10.3390/fermentation8110640
Chicago/Turabian StyleNashiruddin, Noor Idayu, Noor Hidayah Abd Rahman, Roshanida A. Rahman, Rosli Md. Illias, Nazlee Faisal Ghazali, Bassam Abomoelak, and Hesham Ali El Enshasy. 2022. "Improved Sugar Recovery of Alkaline Pre-Treated Pineapple Leaf Fibres via Enzymatic Hydrolysis and Its Enzymatic Kinetics" Fermentation 8, no. 11: 640. https://doi.org/10.3390/fermentation8110640
APA StyleNashiruddin, N. I., Abd Rahman, N. H., A. Rahman, R., Md. Illias, R., Ghazali, N. F., Abomoelak, B., & El Enshasy, H. A. (2022). Improved Sugar Recovery of Alkaline Pre-Treated Pineapple Leaf Fibres via Enzymatic Hydrolysis and Its Enzymatic Kinetics. Fermentation, 8(11), 640. https://doi.org/10.3390/fermentation8110640