Abstract
Comprehensive yeast strain characterization is an important issue for the wine industry as market demands require controlled production of distinctive high-quality wines. Glycosides form an important reservoir of varietal grape wine aroma, and their hydrolysis into olfactory-active compounds essentially depends on the fermenting yeast genera and strains. Among the 14 Metschnikowia, Pichia, Torulaspora and 18 Saccharomyces spp., rapid screenings by agar plate and activity assay, including the substrates arbutin, cellobiose and p-nitrophenol-β-D-glucopyranoside, revealed the most glycosidase-active strains. In the novel co-fermentation setups, five selected non-Saccharomyces and a Saccharomyces strain were separated by a 14 kDa cut-off membrane, allowing respective viable cell counts but facilitating metabolite transfer. Chemical analysis focused on aroma glycosides, with extensive quantification by GC-MS with SIDA on the extracted and hydrolyzed compounds. Olfactory profiles obtained for the non-Saccharomyces wines demonstrated a significant impact of these yeasts, albeit mainly correlated with increased hydrolysis of monoterpene glycosides, and surpassed by a technical Aspergillus niger enzyme. While screenings of non-Saccharomyces strains indicated enhanced glucosidase activity under winemaking conditions, their effect was lower than expected and dominated by ester formation. Interestingly, Saccharomyces yeast cell vitality was increased via in co-fermentation, and non-Saccharomyces strains displayed extended viabilities with high ethanol tolerances.
1. Introduction
Currently, there is a trend in winemaking to return to more natural ways of fermentation, with the objective to produce high-quality distinctive wines and to pronounce the origin of the product. From mainly using fermentation-efficient single-strain Saccharomyces yeasts, the wine industry and related research have started to focus on the cultivation of vineyard- and winery-owned native yeast strains of various genera, and the term “microbiological terroir” has become established [1]. Non-Saccharomyces strains have been commercially available as starter cultures since the early 2000s, with the main purpose of creating wines with more complex aroma, taste and texture profiles. Nevertheless, they are still an underexplored microbiological resource and are limited to several strains of a few genera [2].
Saccharomyces and non-Saccharomyces yeasts have also been subject to research in terms of glycosidase activity. Glycosidic aroma precursors derived from Vitis vinifera L. grapes represent an odorless reservoir of important varietal compounds including monoterpenes, C13-norisoprenoids, C6-compounds and benzene derivatives [3,4]. For these glycosides, the sugar moiety may be present in the form of monosaccharide β-D-glucose, but more commonly as a disaccharide, where β-D-glucose is linked to an additional α-L-arabinofuranosyl-, α-L-rhamnopyranosyl- or β-apiofuranosyl residue [5]. The β-1,4-glycosidic bonds can be hydrolyzed either via acidic hydrolysis induced by low pH or via enzymatic activities expressed by microorganisms or added engineering enzymes. The glycosidase activity of Saccharomyces spp. is generally limited to the exo-1,3-β-glucanase EXG1p [6,7], and with a few exceptions, Saccharomyces yeasts exhibit only very low or no β-glucosidase activity in model systems [8,9,10,11,12]. Thus, their release of glycosidic aroma precursors during fermentation is reported to be rather low [13,14]. In contrast, several non-Saccharomyces yeasts have been found to hydrolyze aroma compounds from glycosylated precursors [15,16] leading to significant positive sensory effects [17,18], and β-glucosidase activity has been specifically reported for strains of Brettanoymces, Candida, Dekkera, Debaromyces, Hanseniaspora, Kloeckera, Metschnikowia, Torulaspora and Pichia [8,9,10,11,15,18]. However, non-Saccharomyces yeasts show much weaker fermentation performance than Saccharomyces yeasts [19] and furthermore tend to produce off-flavors such as ethyl acetate or volatile phenols [20]. Mixed or co-fermentations of non-Saccharomyces and Saccharomyces yeasts are therefore common in winemaking.
Nevertheless, Saccharomyces yeasts have been shown to inhibit the growth and metabolism of non-Saccharomyces yeasts [21,22]. Other studies in contrast demonstrated increased cell density and higher viability of mixed non-Saccharomyces and Saccharomyces fermentations compared to their pure cultures [23,24], and even a synergistic effect on aroma formation by both genera has been hypothesized [25,26]. Overall, interactions between yeasts are subject to complex mechanisms and include nutrition factors, secretion of potentially affecting compounds, and effects due to cell–cell-contacts, however, they strongly depend on the specific conditions and yeasts present [22].
With this previous knowledge, the presented work aims to reveal the respective contributions of Saccharomyces and non-Saccharomyces yeast strains on the hydrolysis of glycosidic aroma precursors, and the resulting impact on the sensory profiles of grape wines. The general hypothesis is that non-Saccharomyces strains exhibit significantly higher glycosidase activities, increase precursor degradation rates, and raise the expression of varietal sensory profiles. It is furthermore assumed that glycosidase activity can be pre-determined using rapid screening methods, and translates to increased precursor hydrolysis rates in grape wine fermentation. In using a fermentation model system with a 14 cut-off membrane spatially separating the co-fermenting yeasts, the analyses of viable cell counts can provide insights into their respective contributions over the course of fermentation. The membrane also minimizes non-Saccharomyces-to-Saccharomyces yeast cell contacts, aiming to increase non-Saccharomyces cell viability and maximum expression of glycosidase activities.
2. Materials and Methods
2.1. Screening for Glycosidase and ß-Glucosidase Activity of Yeasts
To select the most glycosidase-active Saccharomyces and non-Saccharomyces strains relevant to winemaking, rapid screenings were carried out based on long-established methods. They are used complementarily; arbutin has been shown to be a specific substrate for the β-glucosidase enzyme [27], whereas cellobiose and pNPG can be degraded by non-specific glycosidase enzymes [28].
2.1.1. Qualitative Agar Plate Assay
Qualitative agar plate screening according to Rosi et al. [8] and Daenen et al. [9] was carried out on glucose-free Yeast Nitrogen Base (YNB) medium, enriched either by hydrochinon-β-D-glucopyranosid (arbutin) or 4-O-β-D-glucopyranosyl-D-glucose (cellobiose). In order to utilize glucose and grow, yeasts need to express the relevant enzymes to hydrolyze the ß-1,4-linked glucose moiety. Media (20 gL−1 agar), adjusted to pH 5, consisted of the following: (i) arbutin-plates: 6.7 g L−1 YNB, 5 g L−1 arbutin, with or without 0.2 g L−1 ferric ammonium citrate; and (ii) cellobiose-plates: 6.7 g L−1 YNB, 5 g L−1 cellobiose. The media were autoclaved for 15 min at 121 °C and poured into Petri dishes (92 × 16 mm, Sarstedt AG & Co, Nümbrecht, Germany). Dry plates were inoculated with four yeast strains each in horizontal streaks and incubated at 30 °C for seven days. Not inoculated plates served as negative controls. In order to validate the general ability of the yeast strains to grow on agar plates, all strains were plated on YPD (yeast extract peptone dextrose) full medium, including glucose as the major carbon source.
2.1.2. Quantitative Activity Assay
Quantification of glycosidase activity was conducted according to the method of Spagna et al. [29], expressed by the amount of p-nitrophenol (pNP) released from its precursor p-nitrophenol-β-D-glucopyranoside (pNPG). Yeast cultures following the cultivation as described below were centrifuged (4500 rpm, 5 min) and washed twice with deionized water. Pellets were weighed out and resuspended in 10 mL of reaction buffer (sodium acetate 0.1 mol L−1, pH 5). A total of 2 mL of yeast suspension was incubated with 1 mL reaction solution (5 mmolL−1 pNPG in reaction buffer) for 2 h at 37 °C. The reaction was stopped with a stop buffer (sodium carbonate 0.2 mol L−1, pH 9). The blank sample for the photometric assay consisted of 1 mL of yeast suspension in 0.5 mL reaction buffer and 1.5 mL of stop buffer. Samples were centrifuged and pNP released into the supernatant was determined at 400 nm (photometer Nanocolor UV/Vis, Macherey Nagel GmbH & Co. KG, Düren, Germany). Quantification was based on a calibration series of pNP ranging from 0 µmol L−1 to 100 µmol L−1. Enzymatic activities of the yeasts and a technical enzyme (0.01 mg mL−1) were expressed arbitrarily as µU per mg of yeast or enzyme dry weight (µU mg−1). The technical enzyme (its manufacturer specification is detailed below) is obtained from Aspergillus niger and naturally contains β-glucosidase, arabinosidase, apiosidase and rhamnosidase [30]. Its glycosidase activities have been shown to range between 36 mU mg−1 and 380 mU mg−1 [29,31,32].
2.2. Yeasts and Enzymes
A total of 14 commercial and laboratory non-Saccharomyces yeast strains belonging to the species of Metschnikowia fructicola (1), Metschnikowia pulcherrima (3), Pichia anomala (2), Pichia fermentans (2), Pichia kluyveri (3) and Torulaspora delbrueckii (3) and 18 commercially available Saccharomyces spp. strains were tested. Yeast strains were obtained from the yeast collection of the Institute for Viticulture and Oenology at the DLR Rheinpfalz, Neustadt an der Weinstraße, Germany, the German Collection of Microorganisms and Cell Cultures (DSMZ) of the Leibniz Institute, Braunschweig, Germany, and provided by the following yeast producers: Lallemand Inc., Montreal, QC, Canada, Erbslöh Geisenheim AG, Geisenheim, Germany, Eaton Begerow GmbH, Langenlonsheim, Germany and Chr. Hansen Holding A/S, Hørsholm, Denmark. The technical enzyme product was Rapidase Revelation Aroma (formerly AR2000, DSM, Heerlen, The Netherlands).
2.3. Yeast Cultivation
Yeasts were available as glycerol stocks, lyophilized sterile cultures or bacteria-free dry yeast cultures. Yeasts maintained in glycerol stocks were incubated in liquid YPD medium (100 µL stock per 100 µL YPD) at 30 °C for 12–24 h. Lyophilized yeast pellets were rehydrated using 0.5 mL of YPD medium in the provided vessels. Dry yeasts were rehydrated for 15 min at 37 °C in autoclaved tap water, followed by a 15 min period after the addition of 50% pasteurized grape juice. All yeasts were inoculated on YPD plates containing cycloheximide and kanamycin (50 mg mL−1) to prevent any bacterial growth, which may exhibit glycosidase activities. Plates were incubated at 30 °C for 24–48 h. Yeast colonies were picked and transferred to 20 mL of liquid YPD medium and incubated for 24–48 h at 30 °C and 100 rpm on an automatic shaker. All yeasts were maintained on the YPD agar plates at 4 °C to ensure yeast availability during the study. For long-term storage, glycerol stocks were made with 500 µL of autoclaved glycerol and 500 µL of yeast suspension in 2 mL of cryo-vials (neolab, Heidelberg, Germany) and stored at −80 °C.
2.4. Co-Fermentations in a Spatial Separation Model System
2.4.1. Fermentation Setup and Conditions
The model system consisted of Schott flasks (2 L) equipped with a dialysis tube (cut-off 14 kDa, Carl Roth GmbH & Co KG, Karlsruhe, Germany), which spatially divided the fermentation flask into two compartments: non-Saccharomyces strains remained within the dialysis tube (inner compartment, 350 mL) and the Saccharomyces strain was inoculated into the flask itself (outer compartment, 1500 mL). The dialysis membrane functioned as a barrier for yeast cells but allowed the migration of substrates and fermentation metabolites. The successful separation of yeast genera was verified via by microscopic examination (Zeiss AX10, Carl Zeiss AG, Jena, Germany). FT-IR analysis (WineScan FT120, FOSS A/S, Hilerød, Denmark) of both compartments verified the transfer of sugars and ethanol through the membrane. Sterile inoculation and sampling were performed via silicon tubes submerged into the ferment of both compartments, which were permanently kept in place by a perforated polystyrene plug. The sampling ports were equipped with cannulas and Luer locks (neoLab Migge GmbH, Heidelberg, Germany) allowing sampling with disposable syringes (1 mL, neoLab). The ports were sterilized with 70% ethanol prior to and after sampling. Before sampling, ferments were homogenized by stirring the outer compartment with a magnetic rod at 60 rpm for a minute.
All fermentations were conducted in sterilized juice (2 min at 83 °C) of cultivar Muscat Blanc, which was obtained from the research winery of the Institute for Viticulture and Oenology, DLR Rheinpfalz. The grape variety was chosen for its specifically high concentrations of glycosidic aroma precursors. The grape juice with a pH of 3.3 consisted of 79 g of L−1 glucose, 88 g of L−1 fructose, 7.1 g of L−1 titratable acidity (calculated as tartaric acid), and 140 mg of L−1 yeast assimilate nitrogen (YAN, α-amino-nitrogen 84 mg L−1 and 56 mg L−1 ammonium), without juice parameter correction before fermentation.
Co-fermentations were carried out by combining five non-Saccharomyces yeast strains Metschnikowia fructicola MF1, Metschnikowia pulcherrima MP1, Pichia kluyveri PK1, Torulaspora delbrueckii strain TD1 and Torulaspora delbrueckii strain TD2, in sequential inoculation with the Saccharomyces yeast strain SY1. A fermentation of a pure culture of Saccharomyces strain SY1 served as a control. Additionally, another pure SY1 culture was inoculated with a commercial glycosidase enzyme. Fermentations were performed in triplicates at 20 °C in a temperature-controlled room.
2.4.2. Analysis of Yeast Cell Numbers and Fermentation Kinetics
All yeast strains were prepared as described above and inoculated with a targeted cell density of 5 × 106 cells mL−1. The Saccharomyces yeast strain was inoculated 48 h subsequent to the non-Saccharomyces strain in the co-fermentations. After 30 min of inoculation, the actual viable cells were determined to be on average 1.3 × 106 cells mL−1 for strain MF1, 4.7 × 106 cells mL−1 for strain MP1, 3.5 × 106 cells mL−1 for strain PK1, 5.7 × 106 cells mL−1 for strain TD1, 3.9 × 106 cells mL−1 for strain TD2 and 4.2 × 106 cells mL−1 for SY1.
The first sample was taken 30 min after inoculation, and all subsequent samples were taken in 48 h or 72 h intervals. Glucose, fructose, and ethanol were monitored by HPLC (MetaCarb 67C column, 300 × 6.5 mm, Agilent Technologies, Santa Clara, CA, USA), at a column temperature of 85 °C, and the eluent used was demineralized water at 0.6 mL min−1. Detection was conducted using the refractive index (all instruments were from Jasco). The density of viable yeast cells was measured using flow cytometry (Cube 6 with software CyFlowCube 13, Sysmex Partec GmbH, Görlitz, Germany).
After fermentation to dryness, the inner and outer compartments of each fermentation vessel were pooled and clarified via by centrifugation (4 °C, 4500 rpm, 5 min, Rotina 420 R, Andreas Hettich GmbH & Co KG, Tuttlingen, Germany), but by keeping the triplicates separate. The wines were decanted into 750 mL bottles and stabilized by adding 60 mgL−1 of SO2 (Solution sulfureuse, La Littorale S.A.S., Servian, France). Wines were stored at 12 °C for 8–10 weeks prior to chemical and sensory analysis. Additionally, a blend of aliquots from all finished pure SY1 fermentation triplicates was treated using the technical enzyme preparation (50 mg L−1) and stored for 8 weeks at 20 °C. The enzymatic treatment was applied to the dry wine due to the advertised glucose-mediated inhibition of the enzyme during fermentation.
2.5. Chemical Analysis of Glycosylated Aroma Precursors
2.5.1. Extraction and Enzymatic Hydrolysis
Glycosidic precursors were isolated via solid phase extraction (SPE, cartridges Lichrolut EN, 500 mg 6 mL, Merck, Darmstadt, Germany) according to previous publications [33], automated with a robotic system (SmartPrepExtractor Horizon Technology, Salem, MA, USA). Subsequent to enzymatic hydrolysis with a commercial glycosidase enzyme (Rapidase Revelation Aroma, formerly known as AR2000, DSM, Heerlen, The Netherlands) of the extracts as described by Schneider et al. [34], the obtained aglycons (i.e., released aroma compounds) were determined using an optimized setup of gas chromatography–mass spectrometry (GC-MS). A detailed description of the process can be found in the supplementary information (Section S1).
2.5.2. GC-MS with SIDA Analysis of the Hydrolyzed Aroma Compounds
GC-MS analysis was performed using a Trace GC coupled to a Trace MS quadrupole mass spectrometer. The system was equipped with an AS 3000 autosampler (all ThermoFisher Scientific, Waltham, MA, USA). A method known as “concurrent solvent re-condensation large volume splitless injection” according to Magni and Porzano (2003) was applied, as it combines the advantages of splitless injection (non-vaporizing material is kept within the injector liner) and the option to inject larger volumes of sample solutions (up to 50 µL) with a standard split-splitless injector. The separation column consisted of a phenyl–methyl-deactivated uncoated pre-column (4 m × 0.53 mm i.d.; BGB Analytik AG, Rheinfelden, Switzerland) coupled via a press-fit (BGB) to a fused silica capillary (30 m × 0.25 mm i.d.), coated with 0.5 µm of a polyethylene glycol type stationary phase (Phenomenex Inc., Torrance, CA, USA). The carrier gas used was helium at a constant inlet pressure of 75 kPa. The oven temperature was programmed from 47 °C (8 min hold), increased by 5 °C/min to 240 °C (15 min hold). The quadrupole MS was operated in full scan mode (EI+, 70 eV, source temperature 230 °C, scan range 29–300 m/z). Instrument control and data acquisition were performed using Xcalibur software vers. 1.2, and further data processing was performed using software vers. 2.1 (all ThermoFisher Scientific). In total, 23 volatile aroma compounds were quantified via a stable isotope dilution assay (SIDA), whenever corresponding deuterated internal standards were available. Structural identification of substances analyzed was based on reference compounds (if available), linear retention indices and a positive match of the obtained MS spectra with internal databases and the NIST’08 library (Gaithersburg, MD, USA). All measurements were performed in duplicates. Quantification was performed on extracted ion traces. Further details on the analytical method and the aroma chemistry can be found in the supplementary material (Section S1, and Tables S1–S3). The aroma compounds were assigned to five categories: monoterpenes, C6-alcohols, benzene derivatives, C13-norisoprenoids and volatile phenols (Table 1). Calibration (5-point) was performed for concentrations ranging from 5 µg L−1 to 100 µg L−1 for n-hexanol, linalool, β-damascenone and 4-ethylguaiacol, 10 µg L−1 to 300 µg L−1 for (E)-linalooloxide furanoid and 2-phenylethyl ethanol. Non-deuterated compounds and isotopic standards were added to the organic phase and analyzed via GC-MS. With samples, the addition of the isotopic standards was performed after the enzymatic hydrolysis because glycosylated standards are not commercially available. Calibration curves resulted in linear fits with R2 ≥ 0.97. To monitor the performance of sample preparation and enzymatic hydrolysis, n-octyl-β-D-glycopyranoside was added to the wine before SPE, and its hydrolysis product n-octanol was quantified via GC-MS.

Table 1.
Aroma compounds from glycosidic precursors after enzymatic hydrolysis.
2.6. Chemical Analysis of Esters
A headspace solid-phase microextraction (HS-SPME)–GC-MS method with SIDA quantification was used for the quantification of eight esters: ethyl acetate, ethyl butanoate, ethyl hexanoate, ethyl octanoate, ethyl decanoate, 2-methylbutyl acetate, 3-methylbutyl acetate and hexyl acetate. Calibration curves obtained by using the corresponding non-deuterated substance revealed linear fits with R2 ≥ 0.97. Structural identification of all substances was based on linear retention indices and on a positive match of the obtained spectra. Measurements were performed in duplicates; detailed information can be found in the supplementary material (Section S1, Tables S1, S3 and S4). A Trace GC Ultra gas chromatograph equipped with a split–splitless injector was coupled to a DSQ quadrupole mass spectrometer (all ThermoFisher Scientific, Waltham, MA, USA). For sample preparation, 1 mL of wine and 9 mL of brine were pipetted into 20 mL headspace vials, and 10 µL of standard solution was added (supplementary Table S4). A CombiPAL autosampler was used (CTC Analytics, Zwingen, Switzerland). Agitation and incubation were performed using a single magnet mixer (Chromtech, Bad Camberg, Germany) at 40 °C for 10 min. Extraction of analytes was performed using a polydimethylsiloxane (PDMS) fiber (100 µm thickness, 10 mm length, BGB Analytik AG) at 40 °C for 20 min. Desorption of the analytes was performed for 2 min in the injector, held at 240 °C using a split-less time of 2 min. Split flow was then set to 15 mL min−1. GC analysis was performed using a fused silica capillary column (30 m × 0.25 mm i.d.), coated with a phenyl-methyl polydimethysiloxane stationary phase with 0.5 µm film thickness (ZB-5, Phenomenex). The carrier gas used was helium with a constant flow of 1 mL min−1. The oven temperature program started at 40 °C (2 min hold), raised to 200 °C at 3.5 °C/min and finally to 300 °C (5 min hold) at 25 °C/min. Analytes were detected in full scan mode (EI+, 70 eV, ion source temperature 240 °C, scan range 29–300 m/z). Instrument control and data acquisition were performed using Xcalibur software, vers. 2.2 (ThermoFisher Scientific).
2.7. Sensory Analysis
2.7.1. Projective Mapping
All six wines, encoded by three-digit numbers, were simultaneously presented to 12 panelists (trained and experienced staff members of the Institute for Viticulture and Oenology of the DLR Rheinpfalz, 9 male and 5 female; 29–54 years old). Two wines were served in duplicates. Panelists were requested to position the wines on a DIN A3 sheet of paper (297 × 420 mm) according to their own criteria based on similarities and differences among all samples. For flash profiling, all assessors were asked to write free choice descriptors for each wine onto the sheet of paper. The resulting coordinates were digitalized using the FIZZ software (version 2.45 A, Biosystémes, Couternon, France). For statistical evaluation, the x- and y-coordinates and frequencies of grouped free choice descriptors were included in a multiple factor analysis (MFA) in R studio, and the method used was as described by Pagès [35]. In a subsequent discussion with the panelists, seven aromas and five taste attributes were selected to discriminate the aroma of the wines best, and were used as descriptors in the subsequent descriptive analysis (DA).
2.7.2. Descriptive Analysis
The six wines were then evaluated by the same 12 panelists and two additional staff members of the institute (n = 14). Evaluation sessions were performed in a temperature-controlled sensory room at 20 °C with single booths for each taster equipped with a computer for data acquisition, also using the software FIZZ. To obtain the required volumes for sensory analysis, and to unify within treatments, fermentation triplicates were pooled. Panelists were asked to rate on a 10 cm unstructured scale for each of the twelve descriptors defined by the previous mapping session. Intensities scales were anchored on the left by “weak” and on the right by “intense”. signals and on the right by “intense” signals. The panelists were trained in the recognition of the attributes and the use of the scale. Wines were tasted in duplicates on different days and the pure Saccharomyces fermentation was served twice as the control. Samples encoded with three-digit numbers were presented in randomized order. The ratings were sequential for odor descriptors and monadic for taste attributes. Samples were served at 12 °C in clear DIN 10960 glasses (Schott Zwiesel, Zwiesel, Germany) and covered with plastic lids. Sensory standards were prepared in a neutral white wine according to Table 2 and provided in each tasting booth during the sessions. Taste standard preparation and taste descriptor evaluation results are not shown because there were no significant differences found including for sweet, sour, bitter, mouthfeel and body in the wines (analysis of variance, ANOVA LSD, ≤0.05).
Table 2.
Sensory standards for descriptive analysis.
2.8. Data Analysis
Analysis of variance (ANOVA) with Fisher’s Least Significant Difference (LSD) post hoc testing was calculated using the Add-In XLSTAT version 2011. 4.04. (Addinsoft, Paris, France) in Excel 2007 software (Microsoft, Redmond, WA, USA). The software R (R, 2018) and the FactoMineR package were used for the multi-block PCA method multiple factor analysis (MFA) of the Projective Mapping.
2.9. Chemicals
YPD, YNB, cycloheximide, kanamycin, n-octanol, nerol, geraniol, linalool, α-terpineol, 2-phenyl ethanol, 4-ethylguaiacol, n-decane, as well as ethyl esters of carboxylic acids and acetate esters were purchased from Sigma Aldrich (St. Louis, MO, USA). Agar, glycerol, arbutin, cellobiose, pNPG, pNP, sodium acetate, sodium carbonate, n-hexane, dichloromethane and methanol were from Carl Roth. Purchase of isotopic standards: d13-n-hexanol (99.2% d), d5-phenethyl alcohol (99.6% d) and d3-4-ethylguaiacol (99.2% d) were from Dr. Ehrenstorfer GmbH (Augsburg, Germany). d5-Linalool, d5-linalool oxide (furanoid), d4-β-damascenone, deuterated ethyl esters of carboxylic acids and acetate esters were synthesized in-house as described earlier [36]. Octyl-ß-D-glucoside, n-hexanol and linalool oxide (furanoid) were obtained from Fluka (Buchs, Switzerland). β-damascenone was donated from Symrise AG (Holzminden, Germany).
3. Results and Discussion
3.1. Glycosidase and ß-Glucosidase Activity Determined by Screenings
Among the 14 non-Saccharomyces yeasts, the agar plate tests resulted positive for the Metschnikowia strains MF1, MP1, MP2 and MP3 and for the Pichia strains PA1, PA2, PF1 and PK1, indicated by growth on both the arbutin and cellobiose agar plates. These results furthermore lead to the conclusion that the detected glycosidase activity can be specified as β-glucosidase activity because arbutin has been shown to be a specific substrate to that enzyme [27]. In contrast, none of the 18 Saccharomyces strains was able to hydrolyze the substrates arbutin or cellobiose, as they showed no growth on the agar plates (Table 3).
Table 3.
Glycosidase activity of all yeasts and enzyme preparation.
This had been expected, and the results were in accordance with the previously cited studies showing glycosidase activity in non-Saccharomyces, but not in Saccharomyces yeasts [8,9,10,11,12]. However, the quantitative and less specific pNPG assay revealed certain glycosidase activities also among Saccharomyces strains between 24 and 74 µU mg−1, whereas the non-Saccharomyces strains varied in a range of 7 and 138 µU mg−1. It is known that Saccharomyces yeasts in conditions of sugar deficiency can express certain glycosidase activity, which can be attributed to an unspecific side activity of an exo-1,3-β-glucanase [6,7]. According to these findings, and shown by the results obtained in this work, Saccharomyces yeasts are probably contributing to the hydrolysis of glycosylated aroma precursors, at least to a certain quantity, albeit outnumbered by non-Saccharomyces yeasts. However, in the same assay, the technical enzyme preparation, its biological origin is Aspergillus niger, showed a more than 1000-fold higher activity of 89,744 µU mg−1 compared to all yeasts, which reveals the potentially possible degradation of aroma precursors by glycosidase activity from the Muscat Blanc grape juice. Although such a high value cannot be expected from yeasts, the enzyme treatment serves as a reference point for high glycosidase activities. Only two non-Saccharomyces strains, Torulaspora delbrueckii TD1 and TD2, yielded activity values above 100 µU mg−1, which was three times higher than the mean activity of the tested non-Saccharomyces strains at 41 ± 46 µU mg−1 or Saccharomyces strains at 47 ± 14 µU mg−1.
Qualitative screening by growth on arbutin and cellobiose agar plates was limited in accordance with the quantitative activities determined by the pNPG assay. Presumably, and consistent with the previously discussed substrate affinity of yeast glycosidase enzymes, the different ß-1,4-linked moieties including glucose (cellobiose), p-hydroxyphenol (arbutin) and p-nitrophenol (pNPG) had a strong impact on the activity readings. Overall, the detected glycosidase activities of all yeasts, including non-Saccharomyces and Saccharomyces, were rather low in comparison to the technical enzyme preparation. To obtain more authentic insights on the contribution of the glycosidase activities on the complex pool of aroma precursors, and other effects of non-Saccharomyces and Saccharomyces yeasts under fermentation conditions, experimental co-fermentations provided further insights.
Five non-Saccharomyces yeast strains were selected for the experimental winemaking: Metschnikowia strains MF1 and MP1 and Pichia strain PK1 due to their high ß-glucosidase activity determined in the agar plate tests, and Torulaspora delbrueckii strains TD1 and TD2 with the highest glycosidase activity in the pNPG assay. Saccharomyces cerevisiae SY1 was chosen for the pure Saccharomyces control fermentation and for sequential inoculation in all co-fermentations. This commercially available yeast strain shows robust fermentation behavior, also in mixed fermentations [25,37].
3.2. Monitoring of Yeasts and Metabolites in the Model System Co-Fermentations
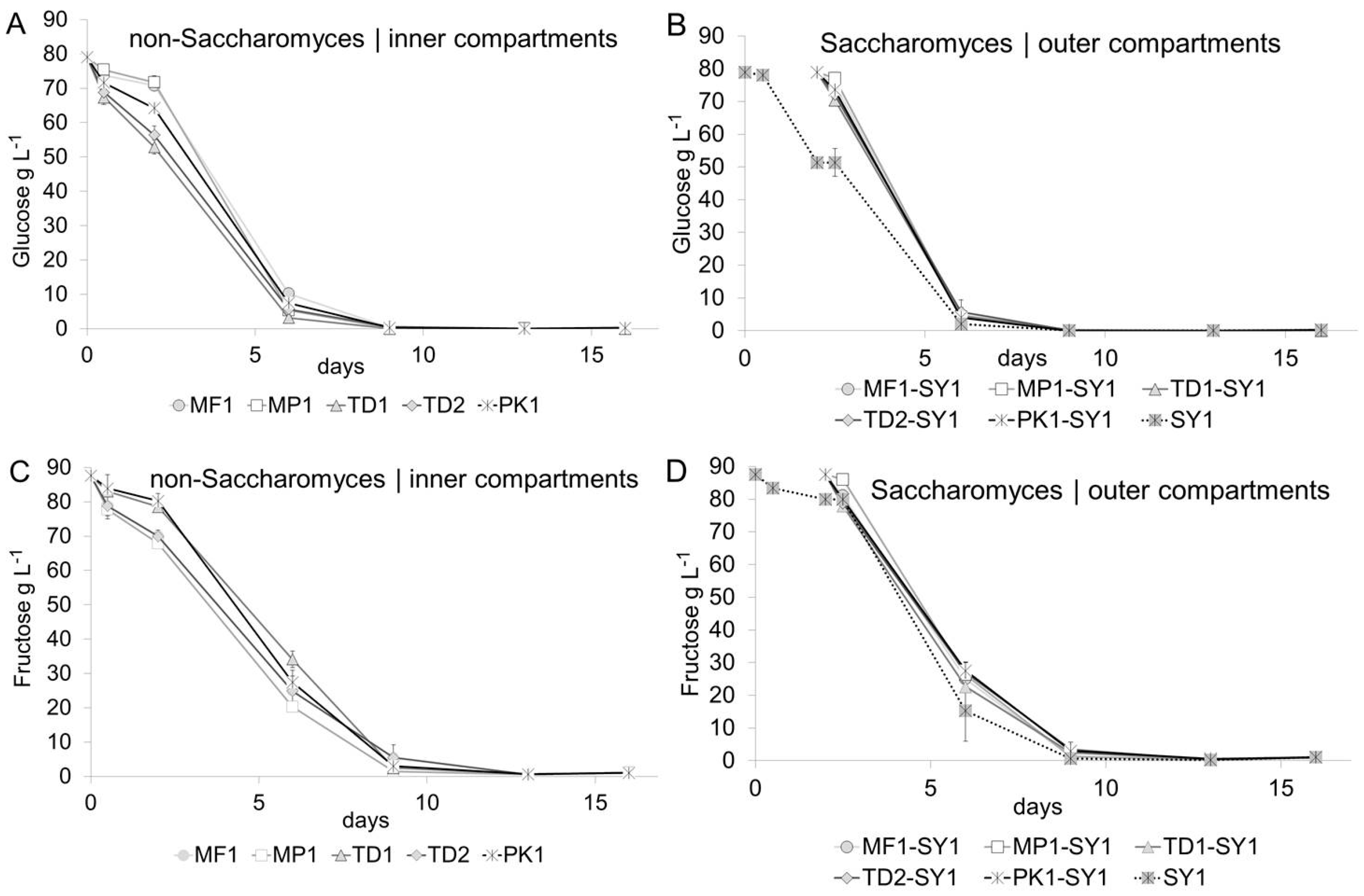
Fermentation progress and cell counts were closely monitored to link the biochemistry of the co-fermenting yeasts with their growth curves. Figure 1 summarizes the sugar kinetics of all fermentations. A generally slightly faster glucose (Figure 1A,B) than fructose (Figure 1C,D) uptake was found for all non-Saccharomyces and Saccharomyces yeasts, which indicated that all yeasts in this study were glycophilic [38]. Only slight variations and non-significant variations in the glucose and fructose degradation rates comparing the individual yeast treatments were found, and they were more pronounced in the inner (non-Saccharomyces) than in the outer (Saccharomyces) compartments. Prior to the inoculation with Saccharomyces on day two of fermentation, non-Saccharomyces yeasts had started to lower the glucose concentrations in the inner compartments by 5–15 g L−1, and the consumption kinetics increased with the inoculation of the strong-fermenting Saccharomyces yeast over the course of fermentation. Glucose was also generally metabolized faster in the outer compartments due to higher activity of the Saccharomyces yeast, which demonstrated a fast, but not instantaneous migration of sugars via the dialysis membrane. All treatments fermented to dryness within 13 days, which was defined by the summed concentration of glucose and fructose below 1 g L−1.
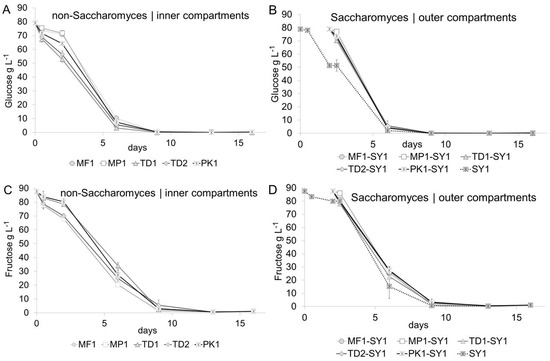
Figure 1.
Fermentation kinetics. A slightly faster decrease in glucose concentrations (A,B) compared to fructose (C,D) indicated that non-Saccharomyces, inner compartment (A,C) and Saccharomyces, outer compartment (B,D), were glycophilic.
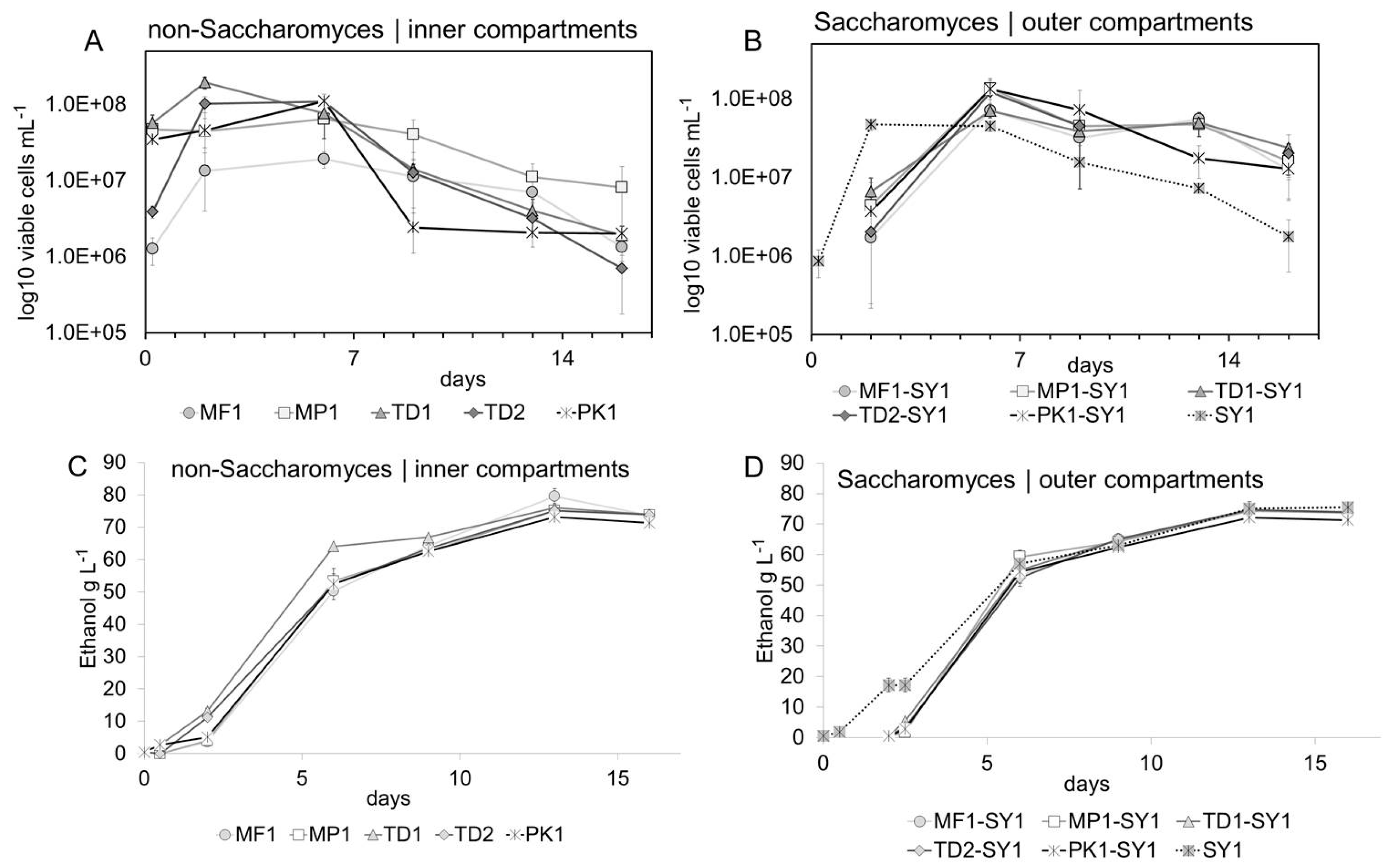
Figure 2A,B display the yeast growth in the inner and outer compartments during fermentation. The initial populations of viable non-Saccharomyces yeasts were determined 30 min after inoculation, varied in the range of 3.4 × 107 and 5.7 × 107 viable cells mL−1 for MP1, PK1 and TD1 and were lower at 1.3 × 106 viable cells mL−1 for MF1 and at 3.94 × 106 viable cells mL−1 for TD2. Variations in the growth curves during the first 48 h occurred presumably due to different amounts of viable cells at the beginning of fermentation which may have already occurred due to differences in their tolerance of ethanol which at this time differed between 5 and 15 g/L (0.6–1.9% v/v alc., Figure 2C,D). Within the first two days, all yeasts reached the maximum population size, followed by a stationary phase of at least four days. A decline in viable non-Saccharomyces cells started at different time points, and in these concentration ranges the effect surely was to be explained by the different tolerances towards ethanol of non-Saccharomyces yeasts [19,39]. The number of living cells of Pichia strain PK1 decreased most rapidly between day six and nine, at an ethanol concentration between 50 and 60 g L−1 (6.3–7.6% v/v), while strain MP1 from this point showed the highest tolerance by constantly maintaining the highest cell numbers. For the other non-Saccharomyces strains, the number of living cells dropped more continuously and slowly on day six and nine, respectively. The unusually long presence of viable non-Saccharomyces cells, particularly up to the end of fermentation, was presumably achieved by the prevention of cell-to-cell contact with the Saccharomyces cells via the dialysis membrane. Due to its cut-off of 14 kDa, the membrane might have served as a protective barrier for toxic compounds secreted by Saccharomyces yeasts [40].
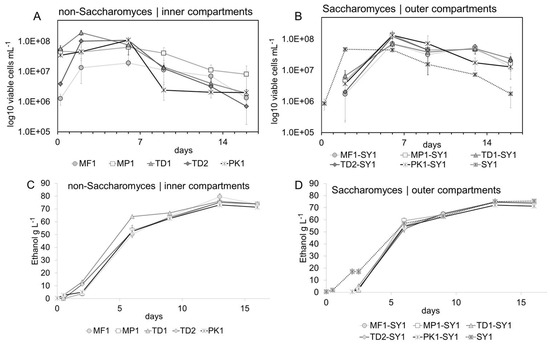
Figure 2.
Yeast growth during fermentation. Non-Saccharomyces yeasts displayed high viable cell numbers during an extended time during fermentation (A) with varying but generally high tolerance towards ethanol (C). Saccharomyces yeasts benefitted significantly from co-fermentation shown by significantly increased viable cell numbers compared to pure fermentation (B), visible over the whole course of fermentation (D), ANOVA, p ≤ 0.05.
Saccharomyces yeasts in the corresponding outer compartments of the co-fermentations and in the pure fermentation had an initial population of 3 × 106 viable cells mL−1 (Figure 2A,B), and growth curves in all treatments evolved in a similar pattern. Cell counts decreased slowly from day six on; however, by the end of fermentation on day 13, the number of living Saccharomyces cells was slightly but significantly higher in the co-fermentations than in the pure Saccharomyces fermentation (p ≤ 0.001, F value 11.6). Saccharomyces yeasts probably benefited from released compounds of autolysis of non-Saccharomyces yeasts such as polysaccharides, proteins, peptides, free amino acids and lipids [41]. Although the transfer of these compounds was probably limited by the used dialysis membrane in the model system, this was a valuable side observation on the benefits of (controlled) mixed fermentations.
3.3. Yeast Mediated Hydrolysis of Glycosylated Aroma Precursors from Grape Juice to Wine in Model Co-Fermentations
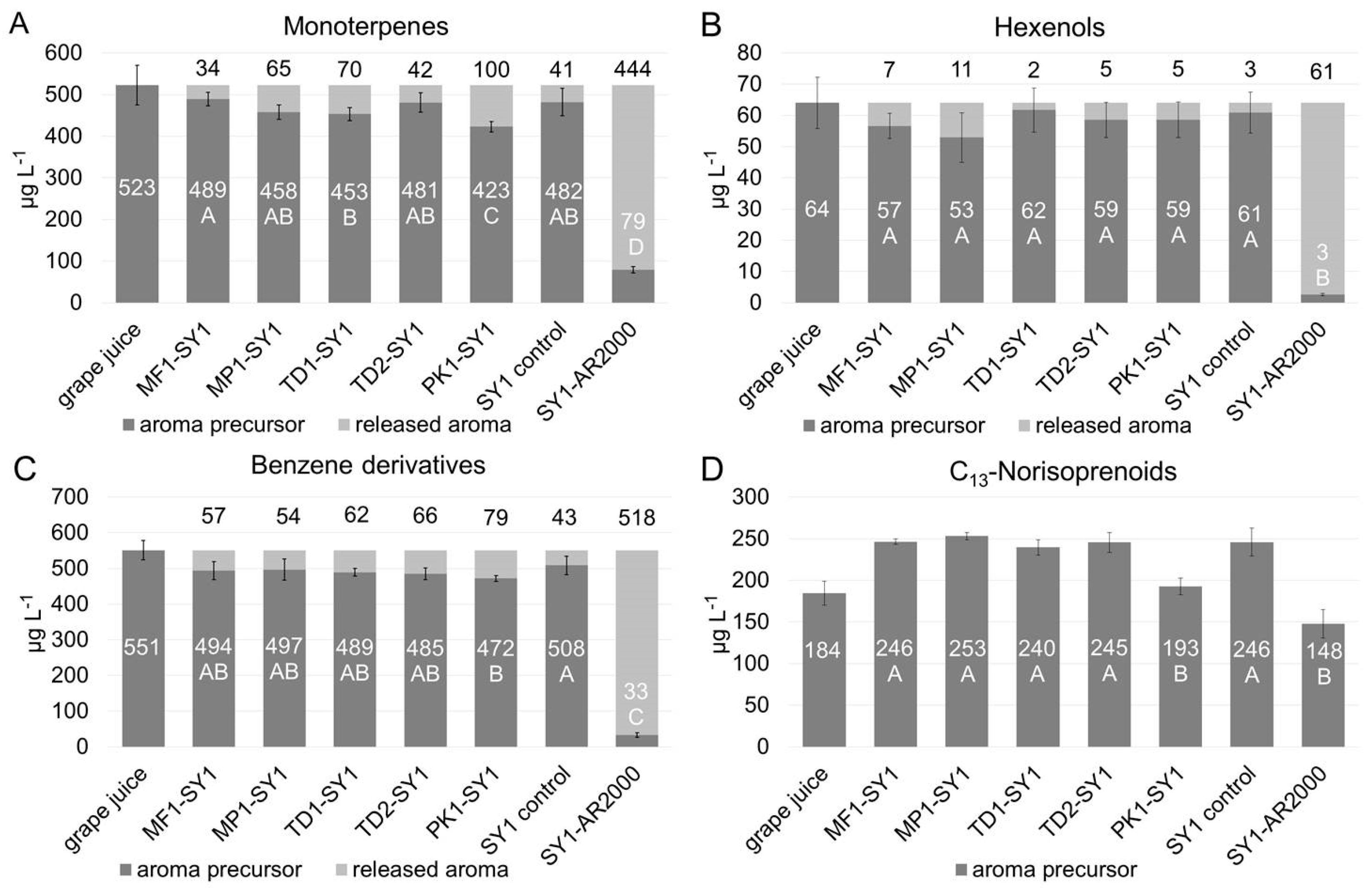
Glycosidase activity exhibited by the selected yeasts during fermentation was determined indirectly by the difference of aroma precursor concentrations being present in the grape juice, and their concentrations in the wines after completed fermentation. The results are displayed in Figure 3A–D, as sum parameters according to the previously defined classes.
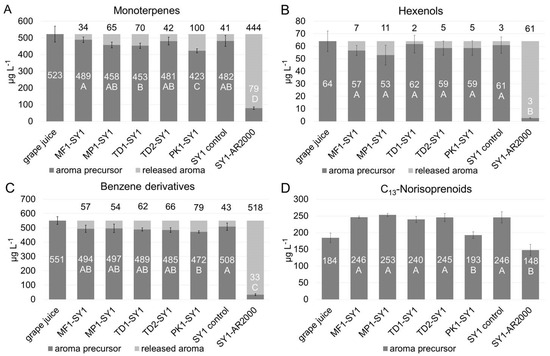
Figure 3.
Concentrations of measured glycosylated precursors (white numbers) and of theoretically released aroma compounds, calculated from the difference in precursors in the juice, and the wines (black numbers). Precursors classified as (glycosides of) monoterpenes (A), C6-alcohols (B), benzene derivatives (C), and C13-norisoprenoids (D) were significantly affected mainly by Pichia strain PK1 and the commercial enzyme (AR2000). ANOVA LSD, p ≤ 0.05, n = 3 fermentation replicates and n = 2 analytical replications.
Non-Saccharomyces yeast enzymatic activity led to overall only moderate degradation of aroma precursors over the course of fermentation. Significant differences among the yeast strains were detected for the remaining glycosylated monoterpenes (p ≤ 0.01, F value 5.56) and C13-norisoprenoids (p ≤ 0.001, F value 11.09). Compared to all other treatments, Pichia kluyveri strain PK1 was found to yield the highest rates, and significantly lowered monoterpene and C13-norisoprenoid precursors to 423 ± 12 µg L−1 (Figure 3A) and 193 ± 10 µg L−1 (Figure 3D), respectively. Thereof, PK1 released 100 µg L−1 of monoterpenes, compared to much lower amounts found for other co-fermentations, including a decrease of 34 µg L−1 for Metschnikowia MP1, or of 41 µg L−1 in the Saccharomyces SY1 control fermentation. Examining linalool, a monoterpene with high olfactory potency, its release was limited to 4–12 µg L−1 in co-fermented wines. Thus, the linalool concentration liberated by the yeasts remained below the sensory threshold determined at 25.2 µg L−1 in model wine [42]. Geraniol, another potent monoterpene, was decreased by concentrations between 23–39 µg L−1, not significantly different among the yeasts analyzed, but overall exceeding its sensory threshold of 30 µg L−1 (see supplementary Figures in Section S2). Non-Saccharomyces yeast strains might be sensitive to stereo chemical differences in the aglycons, as shown for Saccharomyces yeasts that preferentially hydrolyze primary alcohols such as geraniol compared to tertiary alcohols such as linalool [43], albeit with low absolute concentrations; however, overall summing up to a significant release of varietal Muscat Blanc monoterpenes with expected effects on sensory profiles of the wines.
Glycosylated C13-norisoprenoids, albeit significantly different between the yeast treatments compared in contrast to the other measurements, were found with higher concentrations in the wines after fermentation than in the grape juice. This unexpected increase instead of a decline might involve further breakdown or structural rearrangements of C13-norisprenoid precursors caused during sample preparation. More probably, the ethanol present in wine but not in grape juice enhanced the solubility of glycosylated C13-norisoprenoid precursors in the wine samples, and these rather nonpolar compounds adsorbed to a lower degree on the SPE columns from the grape juice than from the wine samples. This may have facilitated their extraction by the SPE material and the subsequent elution with methanol, generally resulting in higher measures of C13-norisoprenoids in wines after fermentation than in the grape juice. A similar effect of increasing free C13-norisoprenoids concentrations during fermentation was observed by Ganß et al. [36] but not by Schneider et al. [34]. The latter, however, analyzed grape juice samples only, so any effects due to sample matrices cannot be excluded by this publication. Despite this uncertainty, a comparison of the co-fermentation revealed that strain PK1 yielded the significantly lowest amount of remaining C13-norisoprenoids precursors compared to all non-Saccharomyces yeast fermentations, including Metschnikowia yeasts MF1 and MP1 as well as Torulaspora yeasts TD1 and TD2 and the Saccharomyces SY1 control.
Nevertheless, the significantly increased release rates of benzene derivatives confirmed that Pichia strain PK1 obtained the highest glycosidase activity among all yeasts tested, and Saccharomyces yeast SY1 the lowest. For C6-alcohol precursors, neither non-Saccharomyces nor Saccharomyces strains yielded differences, presumably due to their low concentrations in general, and precursors of volatile phenols could not be determined, because their concentration was below the calibration range of 5 µg L−1 (see Table 1).
In addition to Besides enzymatic activity, pH-governed chemical hydrolysis supposedly took place as well during fermentation, although it was aimed to limit this effect by keeping the fermentation short, storing the finished wines at a low temperature, and carrying out chemical and sensory analysis eight to ten weeks after the fermentation. Due to equal pH and storage temperature, it could be assumed that the chemical hydrolysis of glycolytic precursors was similar in all samples and was omitted from the discussion as a systematic error source.
However, in contrast to the findings for the yeast treatments, the technical enzyme from Aspergillus niger significantly lowered the concentration of all aroma precursors to a much larger extent than the yeasts alone. The analysis of monoterpene glycosides showed that SY1 pure fermentation released 41 µg L−1 of these odor-active compounds, whereas the additional application of the enzyme increased this number by ten times to 444 µg L−1. The applied enzyme is known to express glycosidase activity at high levels [29,30,31,32] and is very efficient in hydrolyzing both mono- and disaccharides [44]. A more detailed characterization of the enzyme was not the focus of this work, thus it served as a benchmark for high glycosidase activity as demonstrated in the enzyme activity screenings above and furthermore to show the potential of hydrolyzable precursors from the grape juice. Similar results were obtained for benzene derivative and C6-alcohol precursors, and to a lower extent on the C13-norisoprenoid precursors, and possible explanations have been discussed above.
Overall, under fermentation conditions, Saccharomyces and non-Saccharomyces yeast strains yielded a much lower release of glycosylated precursors than had been expected from the agar plate or enzyme activity tests. Only for Pichia kluyveri strain PK1, positive results from the screening tests corresponded to the significantly increased release of monoterpenes (see Table 3). Very possibly, the observed low real expression of glycosidase activity resulted from the winemaking settings, which were high glucose levels in the grape juice with increasing ethanol concentrations over the course of fermentation, anaerobic conditions and low pH levels, presumably strongly diminishing enzymatic activity in the fermentation trials [15]. Among those factors, particularly the free glucose present has been reported to drastically reduce glucosidase activity of both Saccharomyces and non-Saccharomyces yeasts [6,13,45]. Furthermore, hydrolysis of varietal wine aroma precursors requires glycosidase activity suitable for the more complex pool of glycosides including mono- and di-glucosides [44].
Therefore, the prediction validity of the presented rapid screening tests alone seems to be rather limited. Although they are suitable to test high numbers of yeasts at low cost in a short time to select candidates with increased glycosidase activity, they need to be complemented by further screenings for enzymatic abilities and tolerances [46,47] to achieve practical applicability in winemaking.
3.4. Ester Formation
Yeast-derived esters may modify the sensory properties of wines beyond the effects of released glycosylated aroma compounds, so their analysis was included in the study. In general, pure Saccharomyces SY1 fermentation formed the most ethyl esters, and the presence of co-fermenting non-Saccharomyces in contrast yielded fewer ethyl esters (Table 4). Pichia strain PK1 overall produced the most acetate esters, except for hexyl acetate, which was highest in the pure Saccharomyces fermentation. PK1 co-fermented wines were characterized by higher concentrations of 3-methylbutyl acetate, reminiscent of banana, and hexyl acetate, reminiscent of pear, which had no sensory impact presumably due to the masking effect of the high ethyl acetate concentration, which we will be discussing in the next section (On the other hand, Saccharomyces pure fermentation SY1 contained the highest concentrations of ethyl hexanoate, reminiscent of pineapple, ethyl butanoate evoking an apple note as well as ethyl octanoate and ethyl decanoate, contributing to fruitiness and presumably leading to the strongest intensities of exotic citrus fruit. In comparison, 26 mg L−1 of ethyl acetate in the SY1 wine did not mask the fruity perception as was the case for the PK1 co-fermented wine yielding 51 mg L−1 of this compound. All other wines, however, co-fermented with Metschnikowia and Torulaspora strains, did not yield any particularly outstanding concentrations of any ester, and were characterized by generally low-medium levels.
Table 4.
Concentrations of selected esters in co-fermented and control wines. Ethyl- and acetate ester formation was shown to be significantly impacted by the yeast treatment. (ANOVA LSD, p ≤ 0.05).
In general, ester formation found for the analyzed non-Saccharomyces yeasts was lower than shown in other publications [25,37,48]. Ester formation is strongly governed by genetically determined esterase activities expressed by the yeasts, but also strongly influenced by nitrogen supply. It was beyond the scope of this study to characterize the genetically determined expression of alcohol acyltransferase enzymes promoting acetates, biosynthesis and esterase enzymes promoting their hydrolysis [49]. However, to a certain point, the observed differences were probably explainable by the comparatively low yeast assimilable nitrogen (YAN) concentration present in the grape juice used in this study, albeit with 140 mg L−1 YAN (84 mg L−1 alpha-amino-N, 56 mg L−1 ammonium) it was still within the expected range for a grape juice [50]. YAN was therefore not augmented, considering the controlled experimental conditions with a juice at moderate sugar concentrations, lowering the risk of fermentation problems, and in order to avoid the entry of another artificial factor into the experiment. However, the above-cited study of Benito et al. [25] was based on a juice containing 300 mg L−1 YAN (147 mg L−1 alpha-amino-N, with supplemented 127 mg L−1 ammonium) and the results of González–Royo et al. [48] on a model juice with 600 mg L−1 YAN (180 mg L−1 alpha-amino-N, 420 mg L−1 ammonium). YAN variations between 117 mg L−1 and 500 mg L−1 have been shown to significantly modulate seven ethyl esters and four acetate esters including all esters monitored in our study. A lack of YAN, as well as a surplus of it, lowered ester formation, except for ethyl acetate which increased along with nitrogen supply [49].
The nutrient deficiency probably even had a greater effect on the high biomass of the non-Saccharomyces yeasts. Fermentation kinetics (Figure 2) revealed surprisingly high counts of viable cells in the non-Saccharomyces inner compartments over an extended time, from the beginning of fermentation up to rising ethanol levels. Their autolysis, therefore, began at a later stage of fermentation, when increased ethanol levels already started to hinder the nitrogen uptake by the Saccharomyces yeasts. The nutrition of the Saccharomyces co-ferments in this study was sufficient to complete fermentation to dryness; however, it may have been too low to produce as many esters as in the pure Saccharomyces fermentation and in other studies.
3.5. Sensory Profiles Obtained for the Wines of Different Yeast Treatments
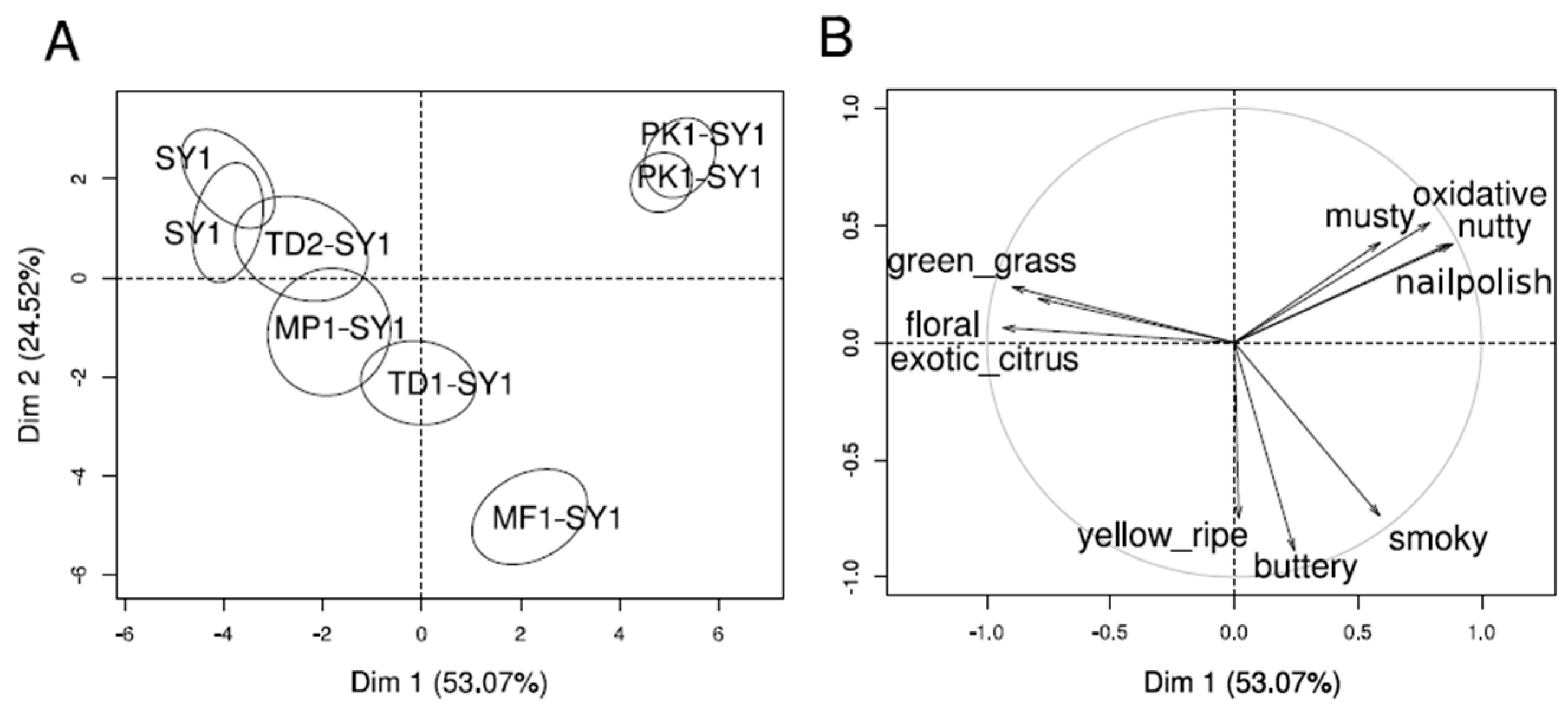
Projective mapping provided a holistic overview of the sensory diversity of the wines. The sample map and the correlations of free-choice descriptor groups are shown in Figure 4. Co-fermentations with strains PK1 and MF1 showed the largest distances and were clearly separated from the other wines. The two wines were mainly described as ‘buttery’ and ‘smoky’, and ‘musty’, ‘oxidized’ and ‘nail polish’ for MF1-SY1 and PK1-SY1, respectively. Wines from pure Saccharomyces fermentation (SY1) and co-fermentation TD2-SY1 were characterized by ‘exotic-citrus’, ‘floral’ and ‘green grass’ notes. Wines from the co-fermentations MP1-SY1 and TD1-SY1 were placed close to wines SY1 and co-fermented with wine TD2-SY1. The validity of the obtained data was confirmed visibly by the proximity of the replicates of SY1 in the upper right corner and PK1-SY1 in the upper left part (Figure 4A). The results of the projective mapping and a discussion with all panelists after the evaluation of all wines led to a list of sensory attributes for subsequent DA, which differentiated the wines best (see Table 2).
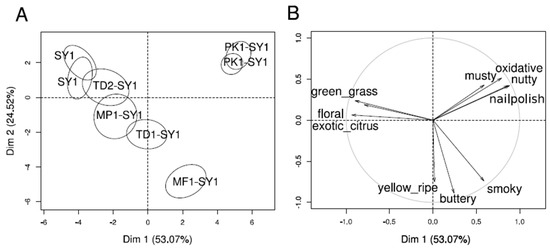
Figure 4.
Projective mapping of the wines separated the wines by aromas mediated by released glycosidic precursors and yeast-derived esters. Representation of wines with 90% confidence ellipses (A) and descriptors (B), scored by 12 judges (MFA, p ≤ 0.05).
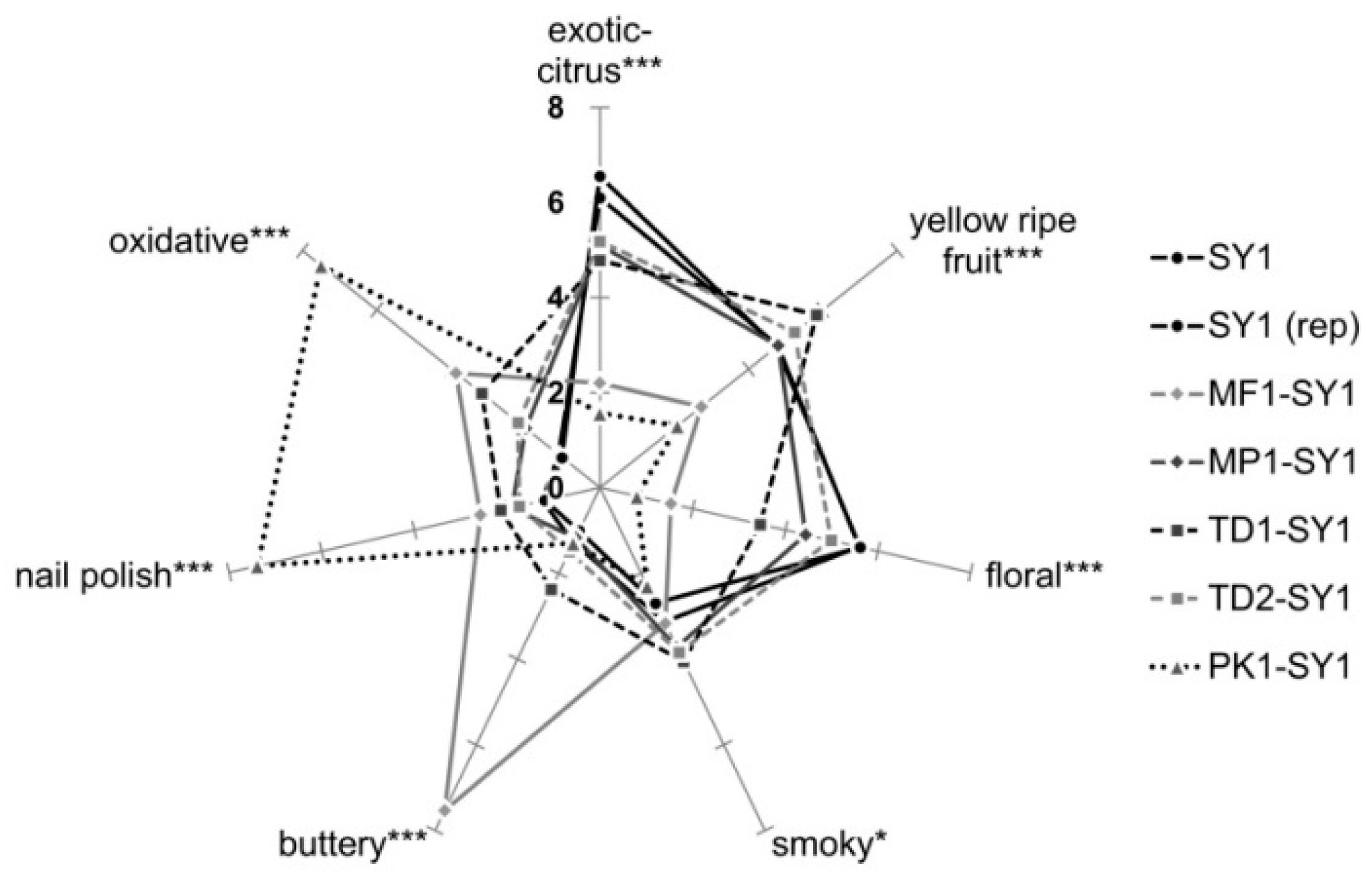
The sensory profiles of the wines obtained by DA are displayed in Figure 5. They confirm the results of the projective mapping and additionally provide more detailed information. Again, the validity of the data was confirmed by similar profiles of blind double-tested control replicate, sample SY1. From the aroma profiles, it was evident that the wine produced solely with Saccharomyces SY1 scored highest in the attributes ‘exotic-fruit’ and ‘floral’. Non-Saccharomyces yeasts significantly modified all seven evaluated sensory attributes, and the according co-fermentations were above all rated higher in the attribute ‘yellow-ripe fruit’. The attributes ‘smoky’, ‘oxidized’, ‘nail polish’ and ‘buttery’ were also scored higher in the presence of non-Saccharomyces strains, which might have masked more varietal aromas like ‘floral’, ‘exotic-fruit’ and ‘yellow-ripe. These findings seemed to show that any possible effects due to hydrolysis of glycosylated precursors were masked by the tremendous presence of dominant esters far above their odor thresholds. In particular, the attribute ‘floral’, mediated by free monoterpenes, is negatively correlated with undesired attributes such as ‘oxidized’. The assumption that off-flavors suppress the perception of desired varietal aromas such as ‘exotic-fruit’ and ‘floral’ became very evident for wine PK1-SY, which was dominated by high ‘nail polish’ and ‘oxidized’ intensities due to concentrations of ethyl acetate of 51 mg L−1 well above its odor threshold reported for 12.3–160 mg L−1 of wine [51,52], and close to the rejection threshold reported for 60 mg L−1 of grape juice [42] (see Table 4). Pichia yeasts have been reported to provide a strong glycosidase activity [53], which was confirmed by this study as strain PK1 significantly increased the hydrolysis on monoterpene precursors, but these yeasts produce high amounts of acetate esters as well [54]. Interestingly, co-fermentations with the Torulaspora delbrueckii strains TD1 and TD2 were rated higher in the ‘yellow-ripe’ fruit impression.
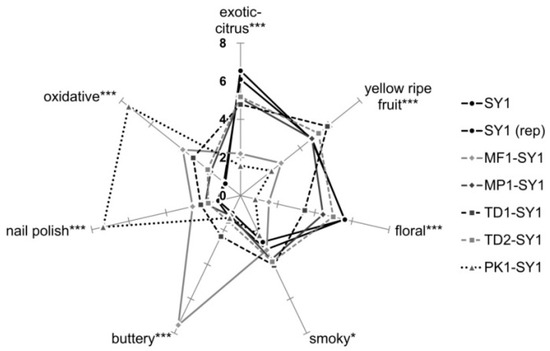
Figure 5.
Descriptive analysis of the wines revealed possible aroma modulation by non-Saccharomyces yeasts but also showed the masking and dominating effects of esters over the potential influence of released glycosydic aroma precursors (14 judges × 2 replications, ANOVA LSD, *, *** p ≤ 0.05).
Overall, the different aroma profiles clearly showed the possibility of modulating wine aroma by co-fermentations with non-Saccharomyces yeasts, but also stressed the importance of further tests, fermentation trials and sensorial evaluation of the produced wines to profoundly understand the complex activity of these yeasts under fermentation conditions. Non-targeted sensory evaluation could rapidly detect flavor scalping of off-flavors, whereas chemical analysis of aroma compounds alone fails to do so. With this need for screening large numbers of yeasts in terms of their sensory impact, extensive DA with long-term training of panel might not be necessary or suitable for practical application aiming to identify potent new yeast strains for practical industry application. Rapid methods including the presented descriptive mapping are able to replace extensive DA and have been validated in various publications [55].
4. Conclusions
The presented study demonstrated the modulation of wine aroma profiles by co-fermenting non-Saccharomyces compared to pure Saccharomyces yeasts. However, it was shown that increased glycosidase activity observed for several yeast strains in enzyme activity assays does not necessarily translate into actual hydrolysis of grape-derived aroma precursors under winemaking conditions, and the contribution of non-Saccharomyces yeasts to wine aroma was rather explained by ester formation than by release of varietal grape aroma precursors. Nevertheless, since the existence of effective glycosidase-active yeasts could be shown as for a selected Pichia strain, rapid activity screenings should be suitable to select promising candidates among high numbers of yeasts, possibly combined with further activity and tolerance screenings to improve their predictability. The co-fermentation model setup achieved the possibility to monitor non-Saccharomyces yeasts over extended life spans during fermentation, as their separation from Saccharomyces yeasts by the 14 kDa membranes maintained viable cells much longer than in usual mixed fermentations. These observed prolonged life spans demonstrated greater tolerance towards high ethanol and low glucose concentrations by non-Saccharomyces yeasts than expected. Nevertheless, significant contribution to aroma glycoside degradation among non-Saccharomyces strains seems to be rather rare. Although a maximized presence of these yeasts was provided, with the exception of one strain non-Saccharomyces, yeasts could not lead to significantly increased glycosidase activity-mediated hydrolysis rates of aroma precursors.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation9110931/s1, Section S1 contains Table S1: Deuterated isotopic standards, Table S2: Identification of important compounds by LRI and MS [56,57,58,59,60,61,62,63,64], Table S3: Quantifiers for all compounds and Table S4: Ester compounds and standards. Section S2 includes Figure S1: Concentration and odor thresholds of the monoterpenes linalool (-glycoside, A) and geraniol (-glycoside, B).
Author Contributions
Conceptualization, Supervision, Funding acquisition, and Writing: U.F.; Methodology, Formal Analysis, Data Curation: H.-G.S., Methodology, and Investigation: M.W.; Conceptualization, Project administration, Methodology, Investigation, Formal Analysis, Data Curation, and Writing: D.S. All authors have read and agreed to the published version of the manuscript.
Funding
The project was partially funded by the International Organization of Vine and Wine (OIV) via a postgraduate research grant, and by the German Ministry of Economics and Technology (via AiF) and the FEI (Forschungskreis der Ernährungsindustrie e. V., Bonn), project AiF-FV 16627 N. Additional support was given by Erbslöh Geisenheim AG, Germany, Eaton Begerow GmbH, German, Chr. Hansen Holding A/S, Denmark and Lallemand Inc., Canada, in providing purified yeast strains.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study (sensory analysis of alcoholic beverages).
Data Availability Statement
Original data are available by directly contacting the corresponding authors.
Acknowledgments
This paper is dedicated to the late Hans-Georg Schmarr (20 August 2019), who will always be remembered and honored for his outstanding knowledge and passion for gas chromatography and related topics. Special thanks again to his great support and co-authorship of this work. Furthermore, many thanks to Philipp Franke and Kathrin Diesler for the flow cytometry analysis, to Jochen Vestner and Martha Wicks-Müller for supporting the sensory analysis and statistics in R studio, and to Jutta Keiser for laboratory assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Mas, A.; Portillo, M.C. Strategies for microbiological control of the alcoholic fermentation in wines by exploiting the microbial terroir complexity: A mini-review. Int. J. Food Microbiol. 2022, 367, 109592. [Google Scholar] [CrossRef] [PubMed]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, X.-L.; Ullah, N.; Tao, Y.-S. Aroma Glycosides in Grapes and Wine. J. Food Sci. 2017, 82, 248–259. [Google Scholar] [CrossRef]
- Noble, A.C.; Strauss, C.R.; Williams, P.J.; Wilson, B. Sensory evaluation of non-volatile flavour precursors in wine. In Flavour Science and Technology, Proceedings of the 5th Weurman Flavour Research Symposium, Voksenasen, Oslo, 23–25 March 1987; Martens, M., Ed.; Wiley & Sons: New York, NY, USA, 1987; pp. 383–391. [Google Scholar]
- Voirin, S.G.; Baumes, R.L.; Sapis, J.-C.; Bayonove, C.L. Analytical methods for monoterpene glycosides in grape and wine: II. Qualitative and quantitative determination of monoterpene glycosides in grape. J. Chromatogr. 1992, 595, 269–281. [Google Scholar] [CrossRef]
- Gil, J.V.; Manzanares, P.; Genovés, S.; Vallés, S.; González-Candelas, L. Over-production of the major exoglucanase of Saccharomyces cerevisiae leads to an increase in the aroma of wine. Int. J. Food Microbiol. 2005, 103, 57–68. [Google Scholar] [CrossRef]
- Schmidt, S.; Rainieri, S.; Witte, S.; Matern, U.; Martens, S. Identification of a Saccharomyces cerevisiae Glucosidase That Hydrolyzes Flavonoid Glucosides. Appl. Environ. Microbiol. 2011, 77, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Rosi, I.; Vinella, M.; Domizio, P. Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 1994, 77, 519–527. [Google Scholar] [CrossRef]
- Daenen, L.; Saison, D.; Sterckx, F.; Delvaux, F.R.; Verachtert, H.; Derdelinckx, G. Screening and evaluation of the glucoside hydrolase activity in Saccharomyces and Brettanomyces brewing yeasts. J. Appl. Microbiol. 2008, 104, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Fia, G.; Giovani, G.; Rosi, I. Study of β-glucosidase production by wine-related yeasts during alcoholic fermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 2005, 99, 509–517. [Google Scholar] [CrossRef]
- Spagna, G.; Barbagallo, R.N.; Palmeri, R.; Restuccia, C.; Giudici, P. Properties of endogenous β-glucosidase of a Saccharomyces cerevisiae strain isolated from Sicilian musts and wines. Enzym. Microb. Technol. 2002, 31, 1030–1035. [Google Scholar] [CrossRef]
- Hernández, L.F.; Espinosa, J.C.; Fernández-González, M.; Briones, A. β-Glucosidase activity in a Saccharomyces cerevisiae wine strain. Int. J. Food Microbiol. 2003, 80, 171–176. [Google Scholar] [CrossRef]
- Delcroix, A.; Günata, Z.; Sapis, J.-C.; Salmon, J.-M.; Bayonove, C. Glycosidase Activities of Three Enological Yeast Strains During Winemaking: Effect on the Terpenol Content of Muscat Wine. Am. J. Enol. Vitic. 1994, 45, 291–296. [Google Scholar] [CrossRef]
- Zoecklein, B.W.; Marcy, J.E.; Williams, J.M.; Jasinski, Y. Effect of Native Yeasts and Selected Strains of Saccharomyces cerevisiaeon Glycosyl Glucose, Potential Volatile Terpenes, and Selected Aglycones of White Riesling (Vitis vinifera L.) Wines. J. Food. Compost. Anal. 1997, 10, 55–65. [Google Scholar] [CrossRef]
- Fernández-González, M.; Di Stefano, R.; Briones, A. Hydrolysis and transformation of terpene glycosides from muscat must by different yeast species. Food Microbiol. 2003, 20, 35–41. [Google Scholar] [CrossRef]
- Mendes Ferreira, A.; Clímaco, M.C.; Mendes Faia, A. The role of non-Saccharomyces species in releasing glycosidic bound fraction of grape aroma components—A preliminary study. J. Appl. Microbiol. 2001, 91, 67–71. [Google Scholar] [CrossRef]
- Varela, C.; Siebert, T.; Cozzolino, D.; Rose, L.; McLean, H.; Henschke, P.A. Discovering a chemical basis for differentiating wines made by fermentation with ‘wild’ indigenous and inoculated yeasts: Role of yeast volatile compounds. Aust. J. Grape Wine Res. 2009, 15, 238–248. [Google Scholar] [CrossRef]
- Hernández-Orte, P.; Cersosimo, M.; Loscos, N.; Cacho, J.; Garcia-Moruno, E.; Ferreira, V. The development of varietal aroma from non-floral grapes by yeasts of different genera. Food Chem. 2008, 107, 1064–1077. [Google Scholar] [CrossRef]
- Fleet, G.H.; Heard, G.M. Yeasts-Growth During Fermentation. In Wine Microbiology And Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers GmbH: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1997, 14, 199–203. [Google Scholar] [CrossRef]
- Liu, Y.; Rousseaux, S.; Tourdot-Maréchal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine microbiome: A dynamic world of microbial interactions. Crit. Rev. Food Sci. Nutr. 2017, 57, 856–873. [Google Scholar] [CrossRef] [PubMed]
- Bordet, F.; Joran, A.; Klein, G.; Roullier-Gall, C.; Alexandre, H. Yeast-Yeast Interactions: Mechanisms, Methodologies and Impact on Composition. Microorganisms 2020, 8, 600. [Google Scholar] [CrossRef]
- Cheraiti, N.; Guezenec, S.; Salmon, J.-M. Redox Interactions between Saccharomyces cerevisiae and Saccharomyces uvarum in Mixed Culture under Enological Conditions. Appl. Environ. Microbiol. 2005, 71, 255–260. [Google Scholar] [CrossRef]
- Mendoza, L.M.; de Nadra, M.C.M.; Farías, M.E. Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Sadoudi, M.; Tourdot-Maréchal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacón, J.-J.; Ballester, J.; Vichi, S.; Guérin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast–yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.A.; van Broock, M.; Valles, S.; Ramón, D.; Caballero, A.C. Screening and typing of Patagonian wine yeasts for glycosidase activities. J. Appl. Microbiol. 2004, 96, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Yabe, T.; Maruyama, Y.; Abe, K.; Nakajima, T. Characterization of Recombinant Yeast Exo-β-1,3-Glucanase (Exg 1p) Expressed in Escherichia coli Cells. Biosci. Biotechnol. Biochem. 2001, 65, 1310–1314. [Google Scholar] [PubMed]
- Spagna, G.; Romagnoli, D.; Angela, M.; Bianchi, G.; Pifferi, P.G. A simple Method for Purifying Glycosidases: α-l-arabinofuranosidase and β-d-glucopyranosidase from Aspergillus niger to Increase the Aroma of Wine. Part I. Enzym. Microb. Technol. 1998, 22, 298–304. [Google Scholar] [CrossRef]
- Gunata, Y.Z.; Dugelay, I.; Vallier, M.J.; Sapis, J.C.; Bayonove, C. Multiple forms of glycosidases in an enzyme preparation from Aspergillus niger: Partial characterization of a β-apiosidase. Enzym. Microb. Technol. 1997, 21, 39–44. [Google Scholar] [CrossRef]
- Baek, H.H.; Cadwallader, K.R. Contribution of Free and Glycosidically Bound Volatile Compounds to the Aroma of Muscadine Grape Juice. J. Food Sci. 1999, 64, 441–444. [Google Scholar] [CrossRef]
- Barbagallo, R.; Spagna, G.; Abbate, C.; Azzaro, G.; Palmeri, R. Inexpensive isolation of β-d-glucopyranosidase from α-l-arabinofuranosidase, α-l-rhamnopyranosidase, and o-acetylesterase. Appl. Biochem. Biotechnol. 2002, 101, 1–13. [Google Scholar] [CrossRef]
- Ibarz, M.; Ferreira, V.; Hernández-Orte, P.; Loscos, N.; Cacho, J. Optimization and evaluation of a procedure for the gas chromatographic–mass spectrometric analysis of the aromas generated by fast acid hydrolysis of flavor precursors extracted from grapes. J. Chromatogr. 2006, 1116, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Schneider, R.; Charrier, F.; Moutounet, M.; Baumes, R. Rapid analysis of grape aroma glycoconjugates using Fourier-transform infrared spectrometry and chemometric techniques. Anal. Chim. Acta 2004, 513, 91–96. [Google Scholar] [CrossRef]
- Pagès, J. Collection and analysis of perceived product inter-distances using multiple factor analysis: Application to the study of 10 white wines from the Loire Valley. Food Qual. Prefer. 2005, 16, 642–649. [Google Scholar] [CrossRef]
- Ganß, S.; Kirsch, F.; Winterhalter, P.; Fischer, U.; Schmarr, H.-G. Aroma Changes due to Second Fermentation and Glycosylated Precursors in Chardonnay and Riesling Sparkling Wines. J. Agric. Food Chem. 2011, 59, 2524–2533. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Bisson, L.F. Yeasts-Metabolism of sugars. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Chur, Switzerland, 1993; pp. 55–75. [Google Scholar]
- Fleet, G.H.; Lafon-Lafourcade, S.; Ribéreau-Gayon, P. Evolution of Yeasts and Lactic Acid Bacteria During Fermentation and Storage of Bordeaux Wines. Appl. Environ. Microbiol. 1984, 48, 1034–1038. [Google Scholar] [CrossRef]
- Albergaria, H.; Francisco, D.; Gori, K.; Arneborg, N.; Gírio, F. Saccharomyces cerevisiae CCMI 885 secretes peptides that inhibit the growth of some non-Saccharomyces wine-related strains. Appl. Microbiol. Biotechnol. 2010, 86, 965–972. [Google Scholar] [CrossRef]
- Hernawan, T.; Fleet, G. Chemical and cytological changes during the autolysis of yeasts. J. Int. Microbiol. 1995, 14, 440–450. [Google Scholar] [CrossRef]
- van Gemert, L.J. Compilations of Odour Threshold Values in Air, Water & Other Media and Compilations of Flavour Threshold Values in Water & Other Media; Oliemans Punter & Partners: Utrecht, The Netherlands, 2011. [Google Scholar]
- Günata, Y.Z.; Bayonove, C.L.; Tapiero, C.; Cordonnier, R.E. Hydrolysis of grape monoterpenyl beta-D-glucosides by various beta-glucosidases. J. Agric. Food Chem. 1990, 38, 1232–1236. [Google Scholar] [CrossRef]
- Cabaroglu, T.; Selli, S.; Canbas, A.; Lepoutre, J.-P.; Günata, Z. Wine flavor enhancement through the use of exogenous fungal glycosidases. Enzym. Microb. Technol. 2003, 33, 581–587. [Google Scholar] [CrossRef]
- Mateo, J.J.; Peris, L.; Ibañez, C.; Maicas, S. Characterization of glycolytic activities from non-Saccharomyces yeasts isolated from Bobal musts. J. Ind. Microbiol. Biotechnol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Madrigal, T.; Maicas, S.; Tolosa, J.J.M. Glucose and Ethanol Tolerant Enzymes Produced by Pichia (Wickerhamomyces) Isolates from Enological Ecosystems. Am. J. Enol. Vitic. 2013, 64, 126–133. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, R.; Sirisena, S.; Gan, R.; Fang, Z. Beta-glucosidase activity of wine yeasts and its impacts on wine volatiles and phenolics: A mini-review. Food Microbiol. 2021, 100, 103859. [Google Scholar] [CrossRef]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Vilanova, M.; Ugliano, M.; Varela, C.; Siebert, T.; Pretorius, I.S.; Henschke, P.A. Assimilable nitrogen utilisation and production of volatile and non-volatile compounds in chemically defined medium by Saccharomyces cerevisiae wine yeasts. Appl. Microbiol. Biotechnol. 2007, 77, 145–157. [Google Scholar] [CrossRef]
- Henschke, P.A.; Jiranek, V. Yeasts-metabolism of nitrogen compounds. In Wine Microbiology and Biotechnology, 1st ed.; Fleet, G.H., Ed.; Taylor and Francis: London, UK, 1993; pp. 77–164. [Google Scholar]
- Corison, C.A.; Ough, C.S.; Berg, H.W.; Nelson, K.E. Must Acetic Acid and Ethyl Acetate as Mold and Rot Indicators in Grapes. Am. J. Enol. Vitic. 1979, 30, 130–134. [Google Scholar] [CrossRef]
- Etiévant, P.X. Wine. In Volatile Compounds in Food. Food Science and Technology; Maarse, H., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1991; pp. 483–546. [Google Scholar]
- Swangkeaw, J.; Vichitphan, S.; Butzke, C.E.; Vichitphan, K. The characterisation of a novel Pichia anomala β-glucosidase with potentially aroma-enhancing capabilities in wine. Ann. Microbiol. 2009, 59, 335. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Varela, P.; Ares, G. Sensory profiling, the blurred line between sensory and consumer science. A review of novel methods for product characterization. Food Res. Int. 2012, 48, 893–908. [Google Scholar] [CrossRef]
- Boido, E.; Lloret, A.; Medina, K.; Fariña, L.; Carrau, F.; Versini, G.; Dellacassa, E. Aroma Composition of Vitis vinifera Cv. Tannat: the Typical Red Wine from Uruguay. J. Agric. Food Chem. 2003, 51, 5408–5413. [Google Scholar] [CrossRef]
- Osorio, C.; Alarcon, M.; Moreno, C.; Bonilla, A.; Barrios, J.; Garzon, C.; Duque, C. Characterization of Odor-Active Volatiles in Champa (Campomanesia lineatifolia R. & P.). J. Agric. Food Chem. 2006, 54, 509–516. [Google Scholar] [PubMed]
- Aasen, A.J.; Kimland, B.; Enzell, C.R. Tobacco chemistry. 18. Absolute configuration of (9R)-9-hydroxy-4,7E-megastigmadien-3-one (3-oxo-α-ionol). Acta Chem. Scand. (1947–1973) 1973, 27, 2107–2114. [Google Scholar] [CrossRef]
- Aubert, C.; Günata, Z.; Ambid, C.; Baumes, R. Changes in Physicochemical Characteristics and Volatile Constituents of Yellow- and White-Fleshed Nectarines during Maturation and Artificial Ripening. J. Agric. Food Chem. 2003, 51, 3083–3091. [Google Scholar] [CrossRef]
- Humpf, H.U.; Schreier, P. Bound aroma compounds from the fruit and the leaves of blackberry (Rubus laciniata L.). J. Agric. Food Chem. 1991, 39, 1830–1832. [Google Scholar] [CrossRef]
- Knapp, H.; Winterhalter, P. Mass spectra and retention indexes of monoterpene diols from rose petals. J. Essent. Oil Res. 2000, 12, 392–399. [Google Scholar] [CrossRef]
- Olle, D.; Baumes, R.L.; Bayonove, C.L.; Lozano, Y.F.; Sznaper, C.; Brillouet, J.-M. Comparison of Free and Glycosidically Linked Volatile Components from Polyembryonic and Monoembryonic Mango (Mangifera indica L.) Cultivars. J. Agric. Food Chem. 1998, 46, 1094–1100. [Google Scholar] [CrossRef]
- Lee, S.-J.; Noble, A.C. Characterization of Odor-Active Compounds in Californian Chardonnay Wines Using GC-Olfactometry and GC-Mass Spectrometry. J. Agric. Food Chem. 2003, 51, 8036–8044. [Google Scholar] [CrossRef] [PubMed]
- Genisheva, Z.; Oliveira, J.M. Monoterpenic characterization of white cultivars from Vinhos verdes appellation of origin (north Portugal). J. Inst. Brew. 2009, 115, 308–317. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).