

Study on the Antibacterial Rule in Fermented Feed with Different Amounts of CaCO3 by Quantitative Real-Time Polymerase Chain Reaction


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Microorganisms
2.3. Expanding the Culture of Strains and Configuration of Culture Media
2.4. Primer Specificity Verification
2.5. Establishment of Strain Standard Curves for qPCR Determination
2.6. Solid-State Fermentation Process
2.7. Changes of Fermented Feed pH and Organic Acid Content with Different Amounts of CaCO3
2.8. Microorganism Detection with qPCR
2.9. Determination of Fermented Feed pH and Organic Acids
2.10. Statistical Analysis
3. Results
3.1. Strain Primer Specificity Verification
3.2. Establishment of qPCR Strain Standard Curves
3.3. Changes of Fermented Feed pH with Different Amounts of CaCO3
3.4. Growth Changes of Escherichia coli, Staphylococcus aureus, and Shigella flexneri with Different Amounts of CaCO3
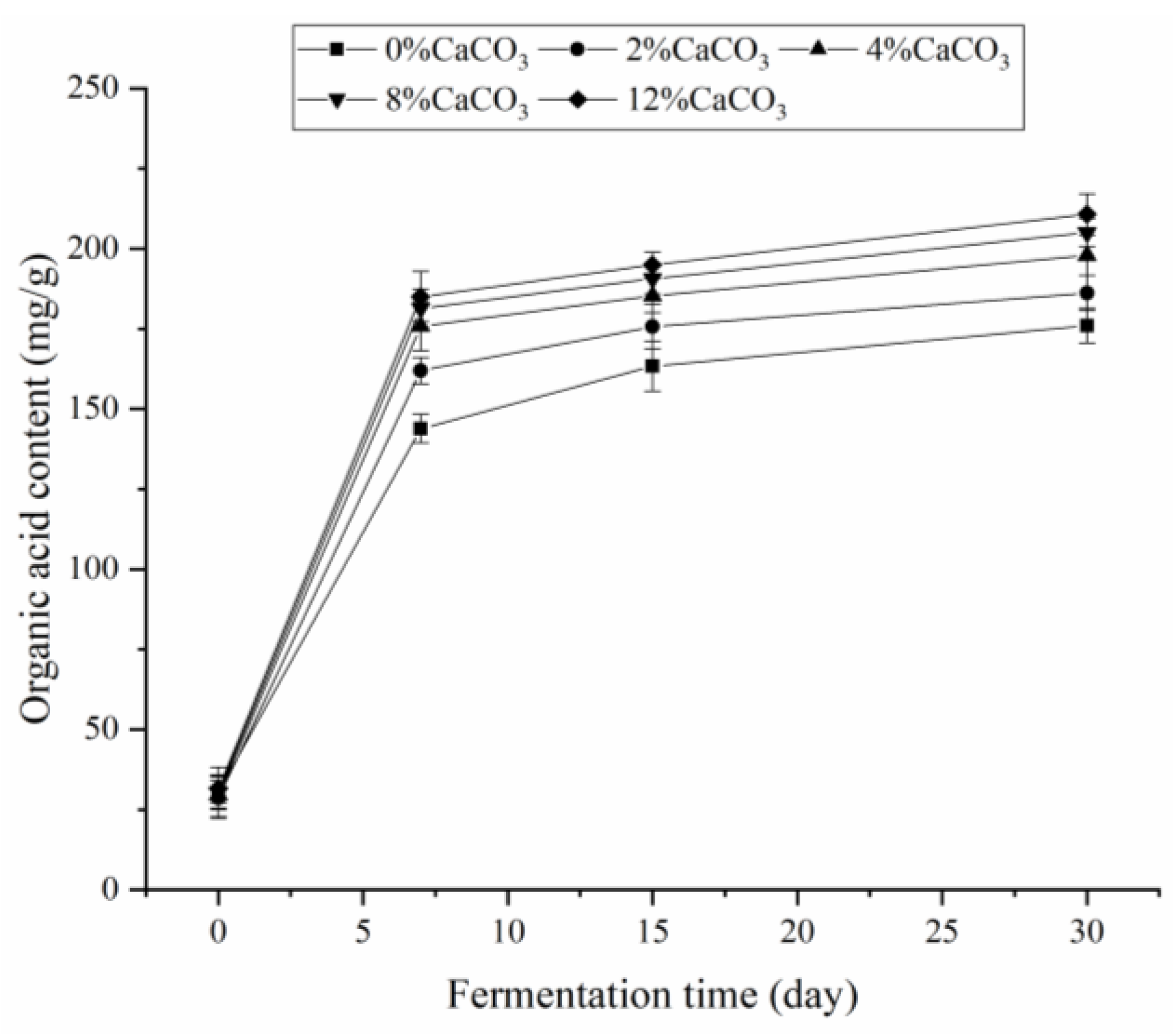
3.5. Changes of Organic Acid Content and Growth Changes of Fermenting Strains with Different Amounts of CaCO3
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Griffin, P.M.; Angulo, F.J.; Tauxe, R.V.; Hoekstra, R.M. Foodborne illness acquired in the United States—Unspecified agents. Emerg. Infect. Dis. 2011, 17, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Carter, B.; Mafura, M.; Liebana, E.; Woodward, M.J.; Anjum, M.F. Genetic diversity among Escherichia coli O157:H7 isolates and identification of genes linked to human infections. Infect. Immun. 2008, 76, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Bania, J.; Dabrowska, A.; Bystron, J.; Korzekwa, K.; Chrzanowska, J.; Molenda, J. Distribution of newly described enterotoxin-like genes in staphylococcus aureus from food. Int. J. Food Microbiol. 2006, 108, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.H.; Cho, S.J. Improvement of bioactivity of soybean meal by solid-state fermentation with Bacillus amyloliquefaciens versus Lactobacillus spp. and Saccharomyces cerevisiae. LWT-Food Sci. Technol. 2016, 68, 619–625. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Zong, X.; Wang, C.; Shi, C.; Wang, F.; Wang, Y.; Lu, Z. Peptides derived from fermented soybean meal suppresses intestinal inflammation and enhances epithelial barrier function in piglets. Food Agric. Immunol. 2020, 31, 120–135. [Google Scholar] [CrossRef]
- Vong, W.C.; Liu, S.Q. Biovalorisation of okara (soybean residue) for food and nutrition. Trends Food Sci. Technol. 2016, 52, 139–147. [Google Scholar] [CrossRef]
- Hawashi, M.; Altway, A.; Widjaja, T.; Gunawan, S. Optimization of process conditions for tannin content reduction in cassava leaves during solid state fermentation using saccharomyces cerevisiae. Heliyon 2019, 5, e02298. [Google Scholar] [CrossRef]
- Iraqi, K.G.E.; Fayed, R.H. Effect of yeast as feed supplement on behavioural and productive performance of broiler chickens. Life Sci. J. 2012, 9, 4026–4031. [Google Scholar]
- Paryad, A.; Mahmoudi, M. Effect of different levels of supplemental yeast (Saccharomyces cerevisiae) on performance, blood constituents and carcass characteristics of broiler chicks. Afr. J. Agric. Res. 2008, 3, 835–842. [Google Scholar]
- Yan, Y.; Li, Y.; Zhang, Z.; Wang, X.; Niu, Y.; Zhang, S.; Xu, W.; Ren, C. Advances of peptides for antibacterial applications. Colloid Surf. B 2021, 202, 111682. [Google Scholar] [CrossRef]
- Amrutha, B.; Sundar, K.; Shetty, P.H. Effect of organic acids on biofilm formation and quorum signaling of pathogens from fresh fruits and vegetables. Microb. Pathog. 2017, 111, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Pézsa, N.P.; Kovács, D.; Somogyi, F.; Karancsi, Z.; Móritz, A.V.; Jerzsele, A.; Rácz, B.; Farkas, O. Effects of Lactobacillus rhamnosus DSM7133 on Intestinal Porcine Epithelial Cells. Animals 2023, 13, 3007. [Google Scholar] [CrossRef] [PubMed]
- Leal, K.; Truong, L.; Maga, E.; King, A. Lactobacillus (L. plantarum & L. rhamnosus) and Saccharomyces (S. cerevisiae): Effects on performance, biochemical parameters, ammonium ion in manure, and digestibility of broiler chickens. Poult. Sci. 2023, 102, 102525. [Google Scholar] [CrossRef] [PubMed]
- Lobo, R.R.; Roesch, L.F.W.; Grilli, E.; Faciola, A.P. Effects of Microencapsulated Blend of Organic Acids and Pure Botanicals on the Ruminal Microbiota in an In Vitro Dual-Flow Continuous Culture System. Fermentation 2023, 9, 730. [Google Scholar] [CrossRef]
- Nithya, V.; Halami, P.M. Antibacterial peptides, probiotic properties and biopreservative efficacy of native Bacillus species isolated from different food sources. Probiotics Antimicrob. Proteins 2012, 4, 279–290. [Google Scholar] [CrossRef]
- Valle, J.C.; Bonadero, M.C.; Fernández-Gimenez, A.V. Saccharomyces cerevisiae as probiotic, prebiotic, synbiotic, postbiotics and parabiotics in aquaculture: An overview. Aquaculture 2023, 569, 739342. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; El-Maaty, H.M.A. Calcium and cholecalciferol levels on late phase-laying hens’ diets: Effects on productive and egg quality traits, blood biochemistry and immune responses. Front. Vet. Sci. 2020, 7, 389. [Google Scholar] [CrossRef]
- Wang, J.; Wang, W.W.; Qi, G.H.; Cui, C.F.; Wu, S.G.; Zhang, H.J.; Wang, J. Effects of dietary Bacillus subtilis supplementation and calcium levels on performance and eggshell quality of laying hens in the late phase of production. Poult. Sci. 2020, 100, 100970. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, L.; Shen, Q.; Qi, L.; Jiang, S.; Guo, Y.; Zhang, C.; Richel, A. Preparation of cattle bone collagen peptides-calcium chelate and its structural characterization and stability. LWT-Food Sci. Technol. 2021, 144, 111264. [Google Scholar] [CrossRef]
- Wei, C.; Liu, G.; Zhang, J.; Bao, J. Elevating fermentation yield of cellulosic lactic acid in calcium lactate form from corn stover feedstock. Ind. Crops Prod. 2018, 126, 415–420. [Google Scholar] [CrossRef]
- Chon, J.W.; Kim, Y.J.; Kim, D.H.; Song, K.Y.; Kim, H.; Kun, H.S. Supplementation of modified mannitol-yolk-polymyxin b agar with cefuroxime for quantitative detection of bacillus cereus in food. J. Food Sci. 2019, 84, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Xiong, T.; Chen, J.; Huang, T.; Xie, M.; Xiao, Y.; Liu, C.; Peng, Z. Fast evaluation by quantitative PCR of microbial diversity and safety of Chinese Paocai inoculated with Lactobacillus plantarum NCU116 as the culture starter. LWT-Food Sci. Technol. 2019, 101, 201–206. [Google Scholar] [CrossRef]
- Xu, H.S.; Roberts, N.; Singleton, F.L.; Attwell, R.W.; Grimes, D.J.; Colwell, R.R. Survival and viability of nonculturable Escherichia coli and Vibrio cholerae in the estuarine and marine environment. Microb. Ecol. 1982, 8, 313–323. [Google Scholar] [CrossRef]
- Wang, X.; Tang, T.; Miao, Q.; Xie, S.; Chen, X.; Tang, J.; Peng, C.; Xu, X.; Wei, W.; You, Z.; et al. Detection of transgenic rice line TT51-1 in processed foods using conventional PCR, real-time PCR, and droplet digital PCR. Food Control 2019, 98, 380–388. [Google Scholar] [CrossRef]
- Costa, G.N.; Vilas-Boas, G.T.; Vilas-Boas, L.A.; Miglioranza, L.H.S. In Silico phylogenetic analysis of lactic acid bacteria and new primer set for identification of lactobacillus plantarum in food samples. Eur. Food Res. Technol. 2011, 233, 233–241. [Google Scholar] [CrossRef]
- Chhetri, V.S.; Han, Y.; Janes, M.E.; Adhikari, A. Evaluation of viability of E. coli o157: h7 on chlorine and lactic acid treated spinach leaves using combined propidium monoazide staining and real-time PCR. LWT-Food Sci. Technol. 2020, 125, 109259. [Google Scholar] [CrossRef]
- Aprodu, I.; Walcher, G.; Schelin, J.; Hein, I.; Norling, B.; Rådström, P.; Nicolau, A.; Wagner, M. Advanced sample preparation for the molecular quantification of Staphylococcus aureus in artificially and naturally contaminated milk. Int. J. Food Microbiol. 2011, 145, S61–S65. [Google Scholar] [CrossRef]
- Yu, Z.; Dong, B.; Lu, W. Dynamics of bacterial community in solid-state fermented feed revealed by 16S rRNA. Lett. Appl. Microbiol. 2009, 49, 166–172. [Google Scholar] [CrossRef]
- Heng, X.; Chen, H.; Lu, C.; Feng, T.; Li, K.; Gao, E. Study on synergistic fermentation of bean dregs and soybean meal by multiple strains and proteases. LWT-Food Sci. Technol. 2022, 154, 112626. [Google Scholar] [CrossRef]
- Elfalleh, W.; Sun, C.; He, S.; Kong, B.; Ma, Y. Changes in enzymatic activities during “koji” incubation and natural fermentation of soybean paste. J. Food Process. Preserv. 2017, 41, e13302. [Google Scholar] [CrossRef]
- Moghaddam, T.K.; Zhang, J.; Du, G. Uvra expression of lactococcus lactis nz9000 improve multiple stresses tolerance and fermentation of lactic acid against salt stress. J. Food Sci. Technol. 2017, 54, 639–649. [Google Scholar] [CrossRef]
- Postollec, F.; Falentin, H.; Pavan, S.; Combrisson, J.; Sohier, D. Recent advances in quantitative PCR (qPCR) applications in food microbiology. Food Microbiol. 2011, 28, 848–861. [Google Scholar] [CrossRef] [PubMed]
- Beal, J.D.; Niven, S.J.; Campbell, A.; Brooks, P.H. The effect of temperature on the growth and persistence of Salmonella in fermented liquid pig feed. Int. J. Food Microbiol. 2002, 79, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Wilks, J.C.; Slonczewski, J.L. pH of the cytoplasm and periplasm of Escherichia coli: Rapid measurement by green fluorescent protein fluorimetry. J. Bacteriol. 2007, 189, 5601–5607. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.O.; Lee, R.E.; Lee, W. A pursuit of Staphylococcus aureus continues: A role of persister cells. Arch. Pharm. Res. 2020, 43, 630–638. [Google Scholar] [CrossRef]
- Peng, S.; Song, J.; Zeng, W.; Wang, H.; Zang, Y.; Xin, J.; Suo, H. A broad-spectrum novel bacteriocin produced by Lactobacillus plantarum SHY 21–2 from yak yogurt: Purification, antimicrobial characteristics and antibacterial mechanism. LWT-Food Sci. Technol. 2021, 142, 110955. [Google Scholar] [CrossRef]
- Costa, W.K.A.D.; Souza, G.T.D.; Brandao, L.R.; Lima, R.C.D.; Garcia, E.F.; Lima, M.D.S.; Souza, E.L.D.; Saarela, M.; Magnani, M. Exploiting antagonistic activity of fruit-derived lactobacillus to control pathogenic bacteria in fresh cheese and chicken meat. Food Res. Int. 2018, 108, 172–182. [Google Scholar] [CrossRef]
- Wang, N.; Le, G.; Shi, Y.; Zeng, Y. Production of bioactive peptides from soybean meal by solid state fermentation with lactic acid bacteria and protease. Adv. J. Food Sci. Technol. 2014, 6, 1080–1085. [Google Scholar] [CrossRef]
- Vázquez, J.A.; González, M.P.; Murado, M.A. Effects of lactic acid bacteria cultures on pathogenic microbiota from fish. Aquaculture 2005, 245, 149–161. [Google Scholar] [CrossRef]
- Rizzotti, L.; Levav, N.; Fracchetti, F.; Felis, G.E.; Torriani, S. Effect of UV-C treatment on the microbial population of white and red wines, as revealed by conventional plating and PMA-qPCR methods. Food Control 2015, 47, 407–412. [Google Scholar] [CrossRef]
- Huang, T.; Shi, Y.; Zhang, J.; Han, Q.; Xia, X.S.; Zhang, A.M.; Song, Y. Rapid and Simultaneous Detection of Five, Viable, Foodborne Pathogenic Bacteria by Photoinduced PMAxx-Coupled Multiplex PCR in Fresh Juice. Foodborne Pathog. Dis. 2021, 18, 640–646. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Primers (5′-3′) | Amplified Fragment |
|---|---|---|
| Bacillus subtilis | F: CGTAGAGCCACTTGAGCG R: CTGCCGTTACAGTTCCTT | 257 |
| Saccharomyces cerevisiae | F: GCGATAACGAACGAGACCCTAA R: CCAGCACGACGGAGTTTCACAAGAT | 225 |
| Lactiplantibacillus plantarum subsp. | F: GTGGTGCGGTCGATATTTTAGTT R: TCAGCCGCGCTTGTAACC | 108 |
| Lacticaseibacillus rhamnosus | F: GACGCAGCCGGTTGACCCAA R: GGCGGCAGTTGCCCCAGAAT | 376 |
| Escherichia coli | F: GCACTAAAAGCTTGGAGCAGTTC R: AACAATGGGTCAGCGGTAAGGCTA | 178 |
| Staphylococcus aureus | F: GCGATTGATGGTGATACGGTT R: AGCCAAGCCTTGACGAACTAAAGC | 279 |
| Shigella flexneri | F: GAGATTCCTGCTCCGCTAA R: TGCGAGGTAGTTGACATTGG | 323 |
| Strains | Standard Curves | Related Coefficient | Efficiency Coefficient |
|---|---|---|---|
| Bacillus subtilis | Y = −3.3493X + 35.973 | R2 = 0.9996 | 98.87% |
| Saccharomyces cerevisiae | Y = −3.4505X + 38.2299 | R2 = 0.9977 | 94.90% |
| Lactiplantibacillus plantarum subsp. plantarum | Y = −3.4474X + 35.226 | R2 = 0.9999 | 95.01% |
| Lacticaseibacillus rhamnosus | Y = −3.4732X + 36.781 | R2 = 0.9951 | 94.05% |
| Escherichia coli | Y = −3.3464X + 39.203 | R2 = 0.9990 | 98.98% |
| Staphylococcus aureus | Y = −3.4632X + 38.2299 | R2 = 0.9991 | 94.42% |
| Shigella flexneri | Y = −3.2115X + 35.226 | R2 = 0.9985 | 104.48% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Heng, X.; Ni, Z.; Chen, H. Study on the Antibacterial Rule in Fermented Feed with Different Amounts of CaCO3 by Quantitative Real-Time Polymerase Chain Reaction. Fermentation 2023, 9, 940. https://doi.org/10.3390/fermentation9110940
Li J, Heng X, Ni Z, Chen H. Study on the Antibacterial Rule in Fermented Feed with Different Amounts of CaCO3 by Quantitative Real-Time Polymerase Chain Reaction. Fermentation. 2023; 9(11):940. https://doi.org/10.3390/fermentation9110940
Chicago/Turabian StyleLi, Jibin, Xinyu Heng, Zhong Ni, and Huayou Chen. 2023. "Study on the Antibacterial Rule in Fermented Feed with Different Amounts of CaCO3 by Quantitative Real-Time Polymerase Chain Reaction" Fermentation 9, no. 11: 940. https://doi.org/10.3390/fermentation9110940