
Comparison of Aqueous and Lactobacterial-Fermented Mercurialis perennis L. (Dog’s Mercury) Extracts with Respect to Their Immunostimulating Activity
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General
2.2. Plant Material
2.3. Preparation of Aqueous Extracts (AE) and Model Fermentation after Inoculation with Lactobacteria Starter Cultures (LBF)
2.4. Whey-Inoculated Fermentation (WF) According to GHP Procedure No. 34c
2.5. D-/L-Lactic Acid (LA) Quantitation
 Pyruvate + NADH + H+. The recovery rate for D-/L-LA amounted to 101.13% (SD 0.03) utilizing the EnzytecTM Liquid multi-acid standard high (R-Biopharm AG). Prior to analysis the aqueous samples or reference standard were diluted with water (1:50 or 1:100 (v/v)).
Pyruvate + NADH + H+. The recovery rate for D-/L-LA amounted to 101.13% (SD 0.03) utilizing the EnzytecTM Liquid multi-acid standard high (R-Biopharm AG). Prior to analysis the aqueous samples or reference standard were diluted with water (1:50 or 1:100 (v/v)).2.6. HPLC(DAD)-ESI-MSn Analysis of Secondary Metabolites
2.7. GC-MS Analysis of Low Molecular Metabolites (<350 Dalton)
2.8. Cell Culture of THP-1 NFκB-eGFP Reporter Cells
2.9. NFκB Activity Assay
2.10. Analysis of Cytokines
2.11. Viability Assay
2.12. Micronucleus Test
2.13. Luminescent Bacteria Toxicity Test
2.14. Statistical Analysis
3. Results and Discussion
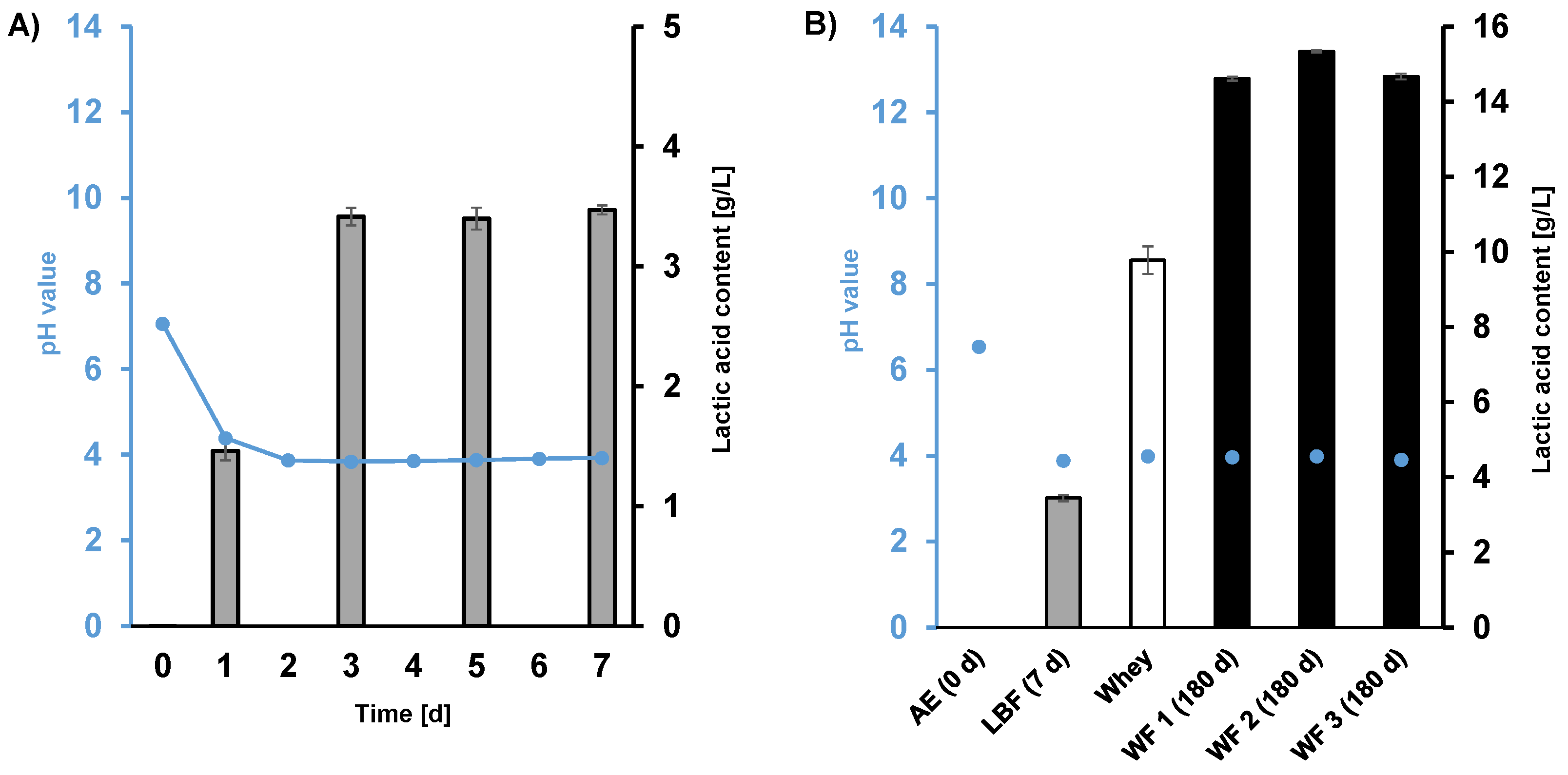
3.1. pH Change and Lactic Acid Formation in Fermentation Samples Inoculated with Lactobacteria (LBF) and Whey (WF)
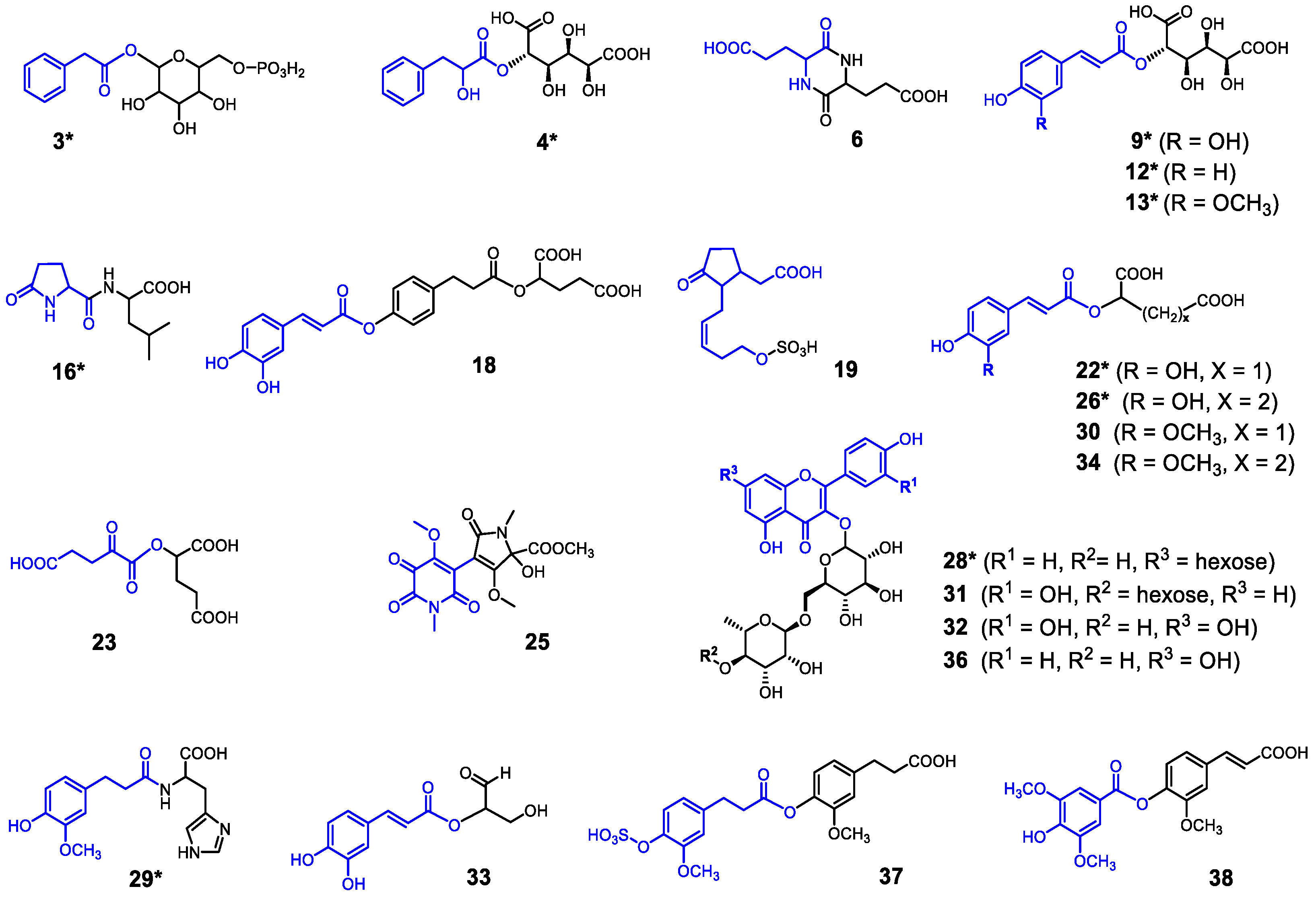
3.2. Characterization of Depsides, Flavonoid Glycosides and Fermentation Metabolites in AE, LBF and WF Samples by LC-MSn
3.3. GC-MS Assignment of Low Molecular Weight Compounds in AE and WF Samples
3.4. Stimulation of NFκB Activity of THP-1 Cells by Treatment with AE, LBF and WF Extracts
3.5. Effect of Extracts on Cytokine Expression
3.6. Bacterial Toxicity: Inhibition of Bioluminescence Induced by WF Extracts in V. fischeri
3.7. Genotoxic Potential: In Vitro Micronucleus Test
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Embacher, K.; Zilkowski, I.; Turek, C.; Stintzing, F.C. Topische Anwendung von Heilpflanzen zur Unterstützung der Wundheilung. Z. Für Phytother. 2021, 42, 249–262. [Google Scholar] [CrossRef]
- Talhouk, R.S.; Karam, C.; Fostok, S.; El-Jouni, W.; Barbour, E.K. Anti-inflammatory bioactivities in plant extracts. J. Med. Food 2007, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.; Kostyuk, V.; de Luca, C.; Pastore, S. Plant phenylpropanoids as emerging anti-inflammatory agents. Mini Rev. Med. Chem. 2011, 11, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Recio, M.C.; Andujar, I.; Rios, J.L. Anti-inflammatory agents from plants: Progress and potential. Curr. Med. Chem. 2012, 19, 2088–2103. [Google Scholar] [CrossRef]
- Miranda, R.d.S.; de Jesus, B.d.S.M.; da Silva Luiz, S.R.; Viana, C.B.; Adão Malafaia, C.R.; Figueiredo, F.d.S.; Carvalho, T.D.S.C.; Silva, M.L.; Londero, V.S.; da Costa-Silva, T.A.; et al. Antiinflammatory activity of natural triterpenes-An overview from 2006 to 2021. Phytother. Res. 2022, 36, 1459–1506. [Google Scholar] [CrossRef]
- Li, X.; Li, X.; Huang, N.; Liu, R.; Sun, R. A comprehensive review and perspectives on pharmacology and toxicology of saikosaponins. Phytomedicine 2018, 50, 73–87. [Google Scholar] [CrossRef]
- Zhang, A.; Yang, X.; Li, Q.; Yang, Y.; Zhao, G.; Wang, B.; Wu, D. Immunostimulatory activity of water-extractable polysaccharides from Cistanche deserticola as a plant adjuvant in vitro and in vivo. PLoS ONE 2018, 13, e0191356. [Google Scholar] [CrossRef] [Green Version]
- Gründemann, C.; Garcia-Käufer, M.; Sauer, B.; Scheer, R.; Merdivan, S.; Bettin, P.; Huber, R.; Lindequist, U. Comparative chemical and biological investigations of β-glucan-containing products from shiitake mushrooms. J. Funct. Foods 2015, 18, 692–702. [Google Scholar] [CrossRef]
- Huang, K.; Li, Y.; Tao, S.; Wei, G.; Huang, Y.; Chen, D.; Wu, C. Purification, characterization and biological activity of polysaccharides from Dendrobium officinale. Molecules 2016, 21, 701. [Google Scholar] [CrossRef] [Green Version]
- Kardošová, A.; Ebringerová, A.; Alföldi, J.; Nosál’ová, G.; Matáková, T.; Hříbalová, V. Structural features and biological activity of an acidic polysaccharide complex from Mahonia aquifolium (Pursh) Nutt. Carbohydr. Polym. 2004, 57, 165–176. [Google Scholar] [CrossRef]
- Lorenz, P.; Beckmann, C.; Felenda, J.; Meyer, U.; Stintzing, F. Das Waldbingelkraut (Mercurialis perennis L.)—Pharmakognosie einer alten Arzneipflanze. Z Phytother 2013, 34, 40–46. [Google Scholar] [CrossRef]
- Rugman, F.; Meecham, J.; Edmondson, J. Mercurialis perennis (dog’s mercury) poisoning: A case of mistaken identity. Br. Med. J. 1983, 287, 1924. [Google Scholar] [CrossRef] [Green Version]
- Watson, P.J. Suspected dog’s mercury (Mercurialis perennis) poisoning in cattle. Vet. Rec. 1998, 142, 116–117. [Google Scholar] [CrossRef] [PubMed]
- German Homoeopathic Pharmacopoeia: GHP; WVG medPharm Scientific Publishers: Stuttgart, Germany, 2005; ISBN 978-3-8047-5088-3.
- Lorenz, P.; Bunse, M.; Sauer, S.; Conrad, J.; Stintzing, F.C.; Kammerer, D.R. Conversion of plant secondary metabolites upon fermentation of Mercurialis perennis L. extracts with two Lactobacteria strains. Fermentation 2019, 5, 42. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, P.; Conrad, J.; Stintzing, F.C. Metabolic fate of depsides and alkaloid constituents in aqueous extracts from Mercurialis perennis L. during fermentation. Chem. Biodivers. 2013, 10, 1706–1723. [Google Scholar] [CrossRef]
- Sauer, S.; Dlugosch, L.; Milke, F.; Brinkhoff, T.; Kammerer, D.R.; Stintzing, F.C.; Simon, M. Succession of bacterial and fungal communities during fermentation of medicinal plants. Fermentation 2022, 8, 383. [Google Scholar] [CrossRef]
- Test No. 487: In Vitro Mammalian Cell Micronucleus Test; OECD Publishing: Paris, France, 2016; ISBN 9789264264861.
- Bryce, S.M.; Bemis, J.C.; Avlasevich, S.L.; Dertinger, S.D. In vitro micronucleus assay scored by flow cytometry provides a comprehensive evaluation of cytogenetic damage and cytotoxicity. Mutat. Res. 2007, 630, 78–91. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, K.; Nakajima, Y.; Matsumura, S.; Chatani, F. An in vitro micronucleus assay with size-classified micronucleus counting to discriminate aneugens from clastogens. Toxicol. Vitr. 2010, 24, 208–216. [Google Scholar] [CrossRef]
- ISO 11348-1:2007; Water Quality—Determination of the Inhibitory Effect of Water Samples on the Light Emission of Vibrio fischeri (Luminescent bacteria test)—Part 1: Method Using Freshly Prepared Bacteria. Beuth Verlag GmbH: Berlin, Germany, 2009.
- Laitila, A.; Sweins, H.; Vilpola, A.; Kotaviita, E.; Olkku, J.; Home, S.; Haikara, A. Lactobacillus plantarum and Pediococcus pentosaceus starter cultures as a tool for microflora management in malting and for enhancement of malt processability. J. Agric. Food Chem. 2006, 54, 3840–3851. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; He, Z.; Wang, X.; Zhao, M.; Cao, X.; Lin, X.; Ji, C.; Zhang, S.; Liang, H. Improving the quality of Suancai by inoculating with Lactobacillus plantarum and Pediococcus pentosaceus. Food Res. Int. 2021, 148, 110581. [Google Scholar] [CrossRef] [PubMed]
- Swan, G.A. Isolation, structure, and synthesis of hermidin, a chromogen from Mercurialis perennis L. J. Chem. Soc. Perkin Trans. 1 1985, 16, 1757–1766. [Google Scholar] [CrossRef]
- Lorenz, P.; Hradecky, M.; Berger, M.; Bertrams, J.; Meyer, U.; Stintzing, F.C. Lipophilic constituents from aerial and root parts of Mercurialis perennis L. Phytochem. Anal. 2010, 21, 234–245. [Google Scholar] [CrossRef]
- Lorenz, P.; Conrad, J.; Duckstein, S.; Kammerer, D.R.; Stintzing, F.C. Chemistry of hermidin: Insights from extraction experiments with the main alkaloid of Mercurialis perennis L. tracked by GC/MS and LC/MSn. HCA 2014, 97, 1606–1623. [Google Scholar] [CrossRef]
- Lorenz, P.; Knödler, M.; Bertrams, J.; Berger, M.; Meyer, U.; Stintzing, F.C. n-Alkylresorcinol occurrence in Mercurialis perennis L. (Euphorbiaceae). Z. Naturforsch. C J. Biosci. 2010, 65, 174–179. [Google Scholar] [CrossRef]
- Lorenz, P.; Conrad, J.; Bertrams, J.; Berger, M.; Duckstein, S.; Meyer, U.; Stintzing, F.C. Investigations into the phenolic constituents of dog’s mercury (Mercurialis perennis L.) by LC-MS/MS and GC-MS analyses. Phytochem. Anal. 2012, 23, 60–71. [Google Scholar] [CrossRef]
- Lorenz, P.; Duckstein, S.; Conrad, J.; Knödler, M.; Meyer, U.; Stintzing, F.C. An approach to the chemotaxonomic differentiation of two European dog’s mercury species: Mercurialis annua L. and M. perennis L. Chem. Biodivers. 2012, 9, 282–297. [Google Scholar] [CrossRef]
- Chamandy, A.; Zhao, M.; Rammal, H.; Ennahar, S. Hyphenated LC-ABTS·+ and LC-DAD-HRMS for simultaneous analysis and identification of antioxidant compounds in Astragalus emarginatus Labill. extracts. J. Pharm. Anal. 2022, 12, 253–262. [Google Scholar] [CrossRef]
- Kapp, K.; Hakala, E.; Orav, A.; Pohjala, L.; Vuorela, P.; Püssa, T.; Vuorela, H.; Raal, A. Commercial peppermint (Mentha × piperita L.) teas: Antichlamydial effect and polyphenolic composition. Food Res. Int. 2013, 53, 758–766. [Google Scholar] [CrossRef]
- Frerot, E.; Chen, T. Identification and quantitation of new glutamic acid derivatives in soy sauce by UPLC/MS/MS. Chem. Biodivers. 2013, 10, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Kiyono, T.; Hirooka, K.; Yamamoto, Y.; Kuniishi, S.; Ohtsuka, M.; Kimura, S.; Park, E.Y.; Nakamura, Y.; Sato, K. Identification of pyroglutamyl peptides in Japanese rice wine (sake): Presence of hepatoprotective pyroGlu-Leu. J. Agric. Food Chem. 2013, 61, 11660–11667. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, S.; Jaiswal, R.; Matei, M.F.; Kuhnert, N. Investigation of acyl migration in mono- and dicaffeoylquinic acids under aqueous basic, aqueous acidic, and dry roasting conditions. J. Agric. Food Chem. 2014, 62, 9160–9170. [Google Scholar] [CrossRef]
- Nakano, H.; Tamai, N.; Tsukamoto, M.; Abe, N. Change in ultraviolet absorption spectra of histidine in aqueous solution as a result of femtosecond laser irradiation. JJSLSM 2010, 31, 390–393. [Google Scholar] [CrossRef] [Green Version]
- Turner, K.W.; Martley, F.G. Galactose fermentation and classification of thermophilic lactobacilli. Appl. Environ. Microbiol. 1983, 45, 1932–1934. [Google Scholar] [CrossRef] [Green Version]
- Iskandar, C.F.; Cailliez-Grimal, C.; Borges, F.; Revol-Junelles, A.-M. Review of lactose and galactose metabolism in lactic acid bacteria dedicated to expert genomic annotation. Trends Food Sci. Technol. 2019, 88, 121–132. [Google Scholar] [CrossRef]
- Loewus, F.A. Inositol and plant cell wall polysaccharide biogenesis. Subcell. Biochem. 2006, 39, 21–45. [Google Scholar] [CrossRef]
- Hadi Alkarawi, H.; Zotz, G. Phytic acid in green leaves. Plant Biol. 2014, 16, 697–701. [Google Scholar] [CrossRef]
- Arena, M.E.; Manca de Nadra, M.C. Biogenic amine production by Lactobacillus. J. Appl. Microbiol. 2001, 90, 158–162. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Capillas, C.; Jiménez-Colmenero, F. Biogenic amines in meat and meat products. Crit. Rev. Food Sci. Nutr. 2004, 44, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Capozzi, V.; Russo, P.; Ladero, V.; Fernández, M.; Fiocco, D.; Alvarez, M.A.; Grieco, F.; Spano, G. Biogenic amines degradation by Lactobacillus plantarum: Toward a potential application in wine. Front. Microbiol. 2012, 3, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucchetti, G.; Locci, F.; Massara, P.; Vitale, R.; Neviani, E. Production of pyroglutamic acid by thermophilic lactic acid bacteria in hard-cooked mini-cheeses. J. Dairy Sci. 2002, 85, 2489–2496. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Suomalainen, T.; Mäyrä-Mäkinen, A.; Huttunen, E. Antimicrobial activity of 2-pyrrolidone-5-carboxylic acid produced by lactic acid bacteria. J. Food Prot. 1997, 60, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Zvanych, R.; Lukenda, N.; Kim, J.J.; Li, X.; Petrof, E.O.; Khan, W.I.; Magarvey, N.A. Small molecule immunomodulins from cultures of the human microbiome member Lactobacillus plantarum. J. Antibiot. 2014, 67, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Bosshart, H.; Heinzelmann, M. THP-1 cells as a model for human monocytes. Ann. Transl. Med. 2016, 4, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartini; Piyaviriyakul, S.; Thongpraditchote, S.; Siripong, P.; Vallisuta, O. Effects of Plantago major extracts and its chemical compounds on proliferation of cancer cells and cytokines production of lipopolysaccharide-activated THP-1 macrophages. Pharmacogn. Mag. 2017, 13, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Eliseo, D.; Pannucci, E.; Bernini, R.; Campo, M.; Romani, A.; Santi, L.; Velotti, F. In vitro studies on anti-inflammatory activities of kiwifruit peel extract in human THP-1 monocytes. J. Ethnopharmacol. 2019, 233, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Albrahim, T.; Alnasser, M.M.; Al-Anazi, M.R.; ALKahtani, M.D.; Alkahtani, S.; Al-Qahtani, A.A. Potential anti-inflammatory and anti-apoptotic effect of Coccinia grandis plant extract in LPS stimulated-THP-1 cells. Environ. Sci. Pollut. Res. Int. 2020, 27, 21892–21904. [Google Scholar] [CrossRef]
- Chun, S.-H.; Lee, H.A.; Lee, K.B.; Kim, S.H.; Park, K.-Y.; Lee, K.-W. Effects of glycated whey protein concentrate on pro-inflammatory cytokine expression and phagocytic activity in RAW264.7 macrophages. Biol. Pharm. Bull. 2016, 39, 199–206. [Google Scholar] [CrossRef] [Green Version]
- Rusu, D.; Drouin, R.; Pouliot, Y.; Gauthier, S.; Poubelle, P.E. A bovine whey protein extract stimulates human neutrophils to generate bioactive IL-1Ra through a NF-kappaB- and MAPK-dependent mechanism. J. Nutr. 2010, 140, 382–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiewiet, M.B.G.; Dekkers, R.; Gros, M.; van Neerven, R.J.J.; Groeneveld, A.; de Vos, P.; Faas, M.M. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS ONE 2017, 12, e0178191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimbrell, M.R.; Warshakoon, H.; Cromer, J.R.; Malladi, S.; Hood, J.D.; Balakrishna, R.; Scholdberg, T.A.; David, S.A. Comparison of the immunostimulatory and proinflammatory activities of candidate Gram-positive endotoxins, lipoteichoic acid, peptidoglycan, and lipopeptides, in murine and human cells. Immunol. Lett. 2008, 118, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahru, A. Ecotoxicological tests in non-ecotoxicological research: Contribution to the three Rs. Use of luminescent photobacteria for evaluating the toxicity of 47 MEIC reference chemicals. ALTEX 2006, 23, 302–308. [Google Scholar]
- In Vitro Evaluation of Genotoxicity and Irritation Potential of Eye Drops Containing Aqueous Plant Extracts, 9th ed.; Zilkowski, I.; Müller, A.; Mörbt, N.; Turek, C.; Stintzing, F.C. (Eds.) 11th World Congress (WC11), Maastricht; ALTEX Proceedings: Kreuzlingen, Switzerland, 2021. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lorenz, P.; Zilkowski, I.; Mailänder, L.K.; Klaiber, I.; Nicolay, S.; Garcia-Käufer, M.; Zimmermann-Klemd, A.M.; Turek, C.; Stintzing, F.C.; Kammerer, D.R.; et al. Comparison of Aqueous and Lactobacterial-Fermented Mercurialis perennis L. (Dog’s Mercury) Extracts with Respect to Their Immunostimulating Activity. Fermentation 2023, 9, 190. https://doi.org/10.3390/fermentation9020190
Lorenz P, Zilkowski I, Mailänder LK, Klaiber I, Nicolay S, Garcia-Käufer M, Zimmermann-Klemd AM, Turek C, Stintzing FC, Kammerer DR, et al. Comparison of Aqueous and Lactobacterial-Fermented Mercurialis perennis L. (Dog’s Mercury) Extracts with Respect to Their Immunostimulating Activity. Fermentation. 2023; 9(2):190. https://doi.org/10.3390/fermentation9020190
Chicago/Turabian StyleLorenz, Peter, Ilona Zilkowski, Lilo K. Mailänder, Iris Klaiber, Sven Nicolay, Manuel Garcia-Käufer, Amy M. Zimmermann-Klemd, Claudia Turek, Florian C. Stintzing, Dietmar R. Kammerer, and et al. 2023. "Comparison of Aqueous and Lactobacterial-Fermented Mercurialis perennis L. (Dog’s Mercury) Extracts with Respect to Their Immunostimulating Activity" Fermentation 9, no. 2: 190. https://doi.org/10.3390/fermentation9020190