Abstract
Anaerobic digestion technology is regarded as the most ideal technology for the treatment of a distiller in terms of environmental protection, resource utilization, and cost. However, there are some limitations to this process, the most prominent of which is microbial activity. The purpose of this paper is to provide a critical review of the microorganisms involved in the anaerobic digestion process of a distiller, with emphasis on the archaea community. The effects of operating parameters on microbial activity and process, such as pH, temperature, TAN, etc., are discussed. By understanding the activity of microorganisms, the anaerobic treatment technology of a distiller can be more mature. Aiming at the problem that anaerobic treatment of a distiller alone is not effective, the synergistic effect of different substrates is briefly discussed. In addition, the recent literature on the use of microorganisms to purify a distiller was collected in order to better purify the distiller and reduce harm. In the future, more studies are needed to elucidate the interactions between microorganisms and establish the mechanisms of microbial interactions in different environments.
1. Introduction
Over-reliance on conventional fossil fuels, a kind of non-renewable energy, would not only result in a monolithic energy system but also negatively affect the environment. So, increasing the use of renewable energy is central to the development strategies of many countries [1,2,3]. In recent years, various technologies are widely studied to make full use of renewable energy. Solid, liquid, or gaseous energy products or modified bio-based materials with a higher energy density can be produced by combining renewable energy with one or more physical, chemical, and biological transformation technologies [4,5,6,7].
In the techniques mentioned above, anaerobic digestion (AD) is one of the most mature biotechnologies at present [8]. Organic waste is degraded by microorganisms to produce biogas and stable residue. Biogas (mainly carbon dioxide and methane) can be converted into electrical or thermal energy, and can also be upgraded to biomethane gas. It can be used as a chemical raw material or fuel to reduce dependence on fossil fuels. It has an important impact on mitigating global warming and can provide up to a 27% greenhouse gas mitigation rate [9,10]. It is reported that methane produced by anaerobic metabolism accounts for more than 90% of the earth [11,12,13,14]. Generally speaking, AD can largely convert organic waste into energy without significant costs, can avoid the waste of resources, and be environmentally friendly.
In the process of ethanol production, a large amount of distiller will be produced. Its composition is complex and has a large number of chemical pollutants, which not only pollute the environment but also have a carcinogenic effect on the human body [15,16,17,18]. However, as a typical biomass resource, distillers have good potential for resource utilization. Distiller is a decent sort of unrefined substance for natural manure, which is wealthy in natural matter, nitrogen, phosphorus, and potassium supplements for plant development, and the amount of destructive substances, for example, anti-microbials and weighty metals is low. Therefore, properly treated distillers can not only be converted into high-quality feed but also generate energy. The application into the soil can boost the number of nutrients in the soil, improve the structure of the microbial community, and help plants grow and develop. Moreover, a distiller is a type of biomass energy carrier that primarily consists of crude fiber, crude fat, crude protein, and starch, among other components. There is evidence that the energy utilization and ecological cycle of distillers are conducive to decarbonization, and it is crucial to make good use of these potential resources [9,14,19,20]. In most cases, the treatment of distillers is inefficient and extensive, which is no longer suitable for today’s green development environment. Therefore, an efficient and environmentally friendly treatment process is urgently required.
AD is currently the most developed and economically feasible process for treating distillers [21,22,23]. Distiller can be used to produce specific resources or all resources at the same time through AD. Studies have shown that the quality of the resources produced depends on the cooperation between microorganisms [11]. Among all microorganisms, although archaea account for 1–2% of the total prokaryotic organisms, it has an important impact on the activity of the microbial community [24]. Because of the macromolecular synthesis pathways and cellular energy consumption structure, its growth speed is limited [25,26]. Therefore, the growth of methane archaea is considered to be the speed-limit stage in the AD process and is vulnerable to environmental factors. Jimenez et al. [27] show bacterial richness in relation to SO42−/COD (chemical oxygen demand), but it is mainly affected by the substrate. Cremonez et al. [28] argued that methane production was limited in a simple single-stage AD system due to the limited activity of methanogenic microorganisms. Peng et al. [29] found that the combined inhibition caused by high ammonia concentration led to changes in the structure and activity of the archaea community, which aggravated the deterioration of the process. In anaerobic digestion, microorganisms can not only basically break down the substrate, but also effectively convert the substances produced by the decomposition into energy. Therefore, microbial activity is important for the anaerobic digestion process. In order for this technology to mature, it is necessary to have a comprehensive understanding of microorganisms, in terms of microbial activity, environmental factors, and certain key microorganisms. This review addresses these issues as a whole.
In recent years, researchers have produced more and more research on the dynamics of microbial communities. To our knowledge, this is the first time in recent years that information on methanogenic archaea has been compiled. Silva et al. [8] mainly discuss the resources produced by distillers, emphasizing the need for microbial research. Costa et al. [9] mainly discussed the metabolic diversity of methanogens but did not consider the influence of operating parameters. The purpose of this study is to review the basic information of methanogenic archaea based on previous studies, especially in terms of methanogenesis. Firstly, the characteristics and energy potential of distillers from different fermentation materials were introduced. Through the discussion of the results of previous studies, the important role and parameter influence of the archaea community were discussed. Then the AD is analyzed from the microbial point of view and the functions of some archaea are discussed. Moreover, considering the pollution of distillers, recent microbial purification techniques are also briefly presented. In view of the diversity of operating parameters and the complex linkage effect between different factors, microbial research on the anaerobic digestion of organic waste is still in the development stage, and molecular detection tools and data analysis methods need to be further optimized [30,31]. A worldwide data-sharing database should be established to consolidate data and promote the further development of microbial research.
2. Present Situation of Distiller
2.1. Distiller Production and Characterization
Distiller comes from the ethanol distillation stage after the fermentation of raw material, and its composition is affected by the raw material type, fermentation process, and the distillation stage [32]. The characteristics of the distiller corresponding to different raw materials are shown in Table 1. Sugar beet distiller usually requires additional nutrients for co-digestion, because it is so high in nitrogen, resulting in a very low C/N [33]. The utilization rate of sugarcane distiller is also the focus of discussion because of its huge yield [9,11,34,35]. Regardless of the raw material, the distiller is acidic and highly contaminated due to its complex composition [36,37]. According to Fuess et al. [23], a large distillery can produce 4.7 billion liters of distiller per year. Distiller contains many toxic substances, such as melanoidins, phenolic compounds, and heavy metals. Chowdhary et al. [17] discuss in detail the harmful substances in distillery wastewater and highlight the harm. The effective treatment of distillers is still an important challenge for the future.

Table 1.
Characteristics of distiller produced by different fermentation materials.
2.2. Different Treatment Methods of Distiller
If the distiller is discharged directly into the water, it will increase water pollution, lead to water eutrophication, and even affect biodiversity. Although some biological solutions have been developed to reduce water pollution, clearance rate and genotoxicity are urgent problems still to be solved [41]. Many treatments have been used to treat distillers [8,35]. Up to 76% of nitrogen loss is a major problem in compost, and although this can be reduced by adding biological carbon, the cost is another factor to consider [42]. Fertilization is by far the most common practice because it contains most of the nutrients needed by crops, but it is not the best option for recycling [7,11,34]. The risks include soil salinization, pore plugging, reduced microbial activity and reduced dissolved oxygen concentration, and the energy recovery is not high. Toxic metals such as cadmium, lead, copper, chromium, and nickel, increase human carcinogenic risk [22,23], but sodium accumulation may be the main reason for limiting fertilization [43]. Therefore, the distiller cannot be treated naturally in the soil. Other applications of distillers, such as for fermentation materials and animal feed, still have problems to be solved in terms of safety [36]. AD has been recommended by many researchers as the core processing technology for a distiller, which has the advantage of a high energy recovery rate and can maximize the energy potential of the distiller and be environmentally friendly.
2.3. Potential Energy and the Synergy of Co-Digestion
Biogas is usually converted into electricity and heat, and if this energy is used to produce ethanol, it can not only save 17 percent of the electricity used to process the raw material but also effectively reduce the carbon footprint [32]. The maximum annual benefits would be USD 886,096.50 and USD 450,256.01, respectively [44]. Biogas upgraded to biomethane gas can be used not only as an alternative to electricity generation but also as a substitute for gasoline. It can avoid greenhouse gas emissions and maintain energy balance. Longati et al. [45] evaluated the economic viability and environmental impact of AD of distillers and also reached positive conclusions. On a laboratory scale, different studies use different metrics to evaluate the energy potential of a sample. The most commonly used is the BMP test. We can judge the conversion of organic matter by the actual methane production rate obtained by the Biochemical methane potential (BMP) test, in order to determine its suitability as a feedstock for AD [46,47,48,49,50]. The high BMP of the distiller indicates that it contains abundant energy. Silva et al. [11] first discussed the volatile fatty acids (VFAs), bioH2, and bioCH4 obtained from the AD treatment and suggested that the energy obtained was about 3.5 GW if all sugarcane vinasse was treated in Brazil. Despite the huge energy potential of distillers, the anaerobic treatment of a single distiller is not ideal [51,52,53,54,55].
The combination of different substrates has long been an efficient way to treat waste with the potential to recover more resources [56,57,58,59,60,61]. Various studies on the co-digestion of distiller have shown synergistic effects [39,50,51]. This synergy is the result of the combination of two or more raw materials that is greater than the sum of the process results of a single raw material, the synergy performance is shown in Table A1. This synergistic effect can increase biogas production, and methane content, alleviate the accumulation and inhibition of TAN and VFAs, and so on. It is important to note that the synergy is not a superposition of the anaerobic fermentation characteristics of individual feedstocks. Urea and trace elements were added to the distiller, which addressed the problem of the time-consuming biological method proposed by Chowdhary et al. [17]. In recent years, Sillero’s team has also discovered the great potential for inactivating pathogens through AD [13,47,51]. AD of more raw materials is also used to treat sludge and food waste [55]. Overall, the synergistic effect makes it easier for the reaction vessel to reach a state of balanced nutrition, and the microorganisms can better evolve and grow, finally making the process achieve an ideal result. What calls for special attention is that the synergistic effect will be affected by the type of raw material, raw material mixing ratio, reaction vessel, reaction temperature, and other factors. For instance, Krzysztof et al. [37] studied the effects of anaerobic co-digestion in sugar beet pulp silage and vinasse at three different ratios, and it was found that the biogas yield at 3:1 was 598.1 mL/g VS and the process was more stable. Moreover, high-temperature treatment is more attractive than medium-temperature treatment under the same raw material and ratio [13,51,62].
However, the observation of this synergistic effect is mostly based on experimental results, and its detailed mechanism is not clear. Chuenchart et al. [53] studied the anaerobic co-digestion performance of food waste and chicken manure, emphasizing the synergistic effect among microorganisms, but did not conduct microbial detection. Peng et al. [29] identified microorganisms that frequently interact with the outside world during long-term AD, constructed interaction networks under specific circumstances, and determined the key roles of certain microorganisms. Jiang et al. [39] also identified the enhancement of the microbial community. Although different authors have optimized the AD process in different ways [11,23,63,64,65,66], low biogas production remains the biggest problem. Few studies have shown the continuity of microbial action. Elucidating the relationship between microbial interactions and the environment and understanding the AD process from the microbial perspective are of great significance for improving process quality [29,38].
3. Analysis Based on Microbial Perspective
3.1. Methanogenesis Pathway and Parameter Influence
In the process of AD of distiller grains, there are two main ways to generate methane, including the acetic acid nutrient pathway Equation (1) and the hydrogen nutrient pathway Equation (2). The impact of operation parameters on the path is shown in Table 2.
Table 2.
Impact of operation parameters.
3.1.1. Hydrogenotrophic Methanogenesis
Leading the hydrogenotrophic methanogenesis are hydrogenotrophic methanogens, such as Methanobacterium, Methanoculleus, and Methanothermobacter. Hydrogenotrophic methanogen is very important for the whole process because it not only consumes the H2 and CO2 to produce methane but also can let syntrophic acetogenic microorganisms remain active microorganisms [10,69,70]. Recently, it has been proven to play a key role in phenol degradation [67]. However, its activity and quantity are easily disturbed by many substances. With increasing ammonium concentration, the activity of hydrogenotrophic methanogens first increases and then decreases [7,29]. At different concentrations of hydrogen, the growth of archaea, H2 utilization, and other parameters are greatly affected [12,71]. Hydrogenotrophic methanogens are more tolerant to cations, high OLR, and ammonia concentrations than acetic acid-utilizing methanogens [16,68,72,73]. However, it is not as tolerant to acetic acid concentration, and Rachbauer et al. [6] recommend acetate concentration below 0.8 g/L. Microorganisms prefer hydrogenotrophic methanogenesis to other pathways because hydrogenotrophic methanogens are more widely distributed at both high and mesophilic temperatures [38], and hydrogenotrophic methanogens are more active at high temperatures, which can lead to shorter process times and thus more economic benefits [28,63,74].
3.1.2. Acetotrophic Pathway
The methanogenesis of acetate follows two different pathways: acetoclastic methanogenesis and SAO [24]. Acetoclastic methanogens and syntrophic acetate-oxidizing bacteria (SAOB) compete for acetate. Acetate is an important intermediate and inhibitor of methanogenesis. High concentrations of acetate are toxic to microorganisms [25,29]. At the same time, high concentrations of acetic acid limited propionic acid degradation and inhibited acetate methanogenic bacteria [7,16].
Acetoclastic methanogens include Methanosarcina and Methanosaeta. 13C isotope analysis of Xiao et al. [75] indicates that methane comes from direct cleavage of acetic acid when abundant, but this has not been proven in AD. There are few studies on SAOB. SAOB is a difficult and slow-growing bacterium to culture. SAOB has been found in: Ca.Syntrophonatronum acetioxidants, Ca.Contubernalis alkalaceticum, Syntrophaceticus, and Coprothermobacter [24,25,38,68,76]. As the total ammonium concentration and temperature increase, SAO will gradually replace acetoclastic methanogenesis.
3.2. Stability of System
The stability of the AD process has been the focus of discussion [63]. Acidogenic bacteria and methanogenic microorganisms differ greatly in their sensitivity to environmental conditions. Failure to maintain a balance between these two groups of microbes is the main cause of reactor instability. Poor operational stability has prevented the widespread use of AD [77,78,79].
Numerous experiments have shown that the two-stage AD strategy plays a positive role in improving stability [32,53,80,81]. It can effectively alleviate the accumulation of inhibitors in single-stage AD and is more efficient in terms of biogas production and methane production [26,63,82]. Notably, in two-stage AD, the acidogenic and methanogenic processes run separately, producing methane via the acetotrophic route only.
It is extremely important to use an appropriate combination of operational and control parameters in a bioreactor to ensure maximum efficiency of the process [77]. Short HRT and high OLR have been recognized as causing system instability [51,52,53,54]. Therefore, low OLR and high HRT (hydraulic retention time) are generally used to start the experimental initiation phase in order to minimize the pressure on the microbial population during the adaptation period [40]. Reactor optimization can also deal with such problems [16,63]. Effective monitoring of AD is essential for optimal process results. Different stability indicators have been applied in different studies to describe system stability [40,77]. Because of the complexity of the AD process, it is recommended to use different combinations of stability indicators to monitor the AD process.
Acidic conditions are known to negatively affect methanogenesis. pH is often used as a key indicator to monitor anaerobic degradation processes [77]. The pH value remains neutral, which meets the growth requirements of most methanogens and is suitable for methane production. Although slight fluctuations in pH may also have beneficial effects, the pH should be kept within the appropriate range for methane generation [29,68]. As described in Part 2.3, the added substrate has a positive effect on pH stabilization, especially anaerobic triple digestion. More importantly, this synergy automatically balances out small pH fluctuations caused by changes in operating parameters [50,51]. In general, the pH of two-stage AD is generally more stable than that of one-stage digestion and therefore has better biogas yields [80,82,83]. Despite the benefits of using pH as a stability indicator, it may be less sensitive to monitoring substrates with higher organic loading rates and may not be appropriate as an early indicator [77]. For the anaerobic treatment of the distiller, we can still ensure pH stability and system stability by adding appropriate substrates and additives (NH4Cl, NaHCO3, NaOH) [46].
TAN has been a hot topic of research. Optimal TAN ensures the adequate buffering capacity of the medium, whereas high concentrations lead to inhibition and collapse of anaerobic processes [84,85,86]. The statistics are shown in Table 3 below.
Table 3.
Ammonia concentration thresholds recommended in different studies.
For optimal methane production at higher temperatures, ammonium concentration should not exceed 5000 mg/L [87,88]. It is important to note that the concentration limit for TAN is not fixed and must be studied for each case, as it depends on factors such as the type of substrate used, gas emissions, operating conditions, and the degree of microbial acclimation [29,78,85]. Because TAN concentration is the result of microbial degradation of nitrogenous substances, the tolerance of TAN can be improved by pretreatment of the substrate to improve the process stability [89,90,91]. In the experiment of Gao et al. [73], when the TAN concentration of the unacclimated reactor is about 4000 mg/L, the methanogenesis process stops completely, while the acclimated reactor can operate stably at an average TAN concentration of 4293 mg/L. It should be emphasized that high TAN values were observed at the end of anaerobic triple digestion, but no inhibition was observed [46,50]. The specific mechanism of this synergy has yet to be studied. Some studies believe that FAN (free ammonia) has a stronger correlation with process stability than TAN [92]. FAN neutralizes the acid to reduce the risk of acidification in the reactor, but because of its membrane permeability, it interferes with cell activity [73,78]. It was reported that FAN content above 200 mg L−1 would hinder stability, and that above 300 mg L−1 would lead to a sharp reduction in biogas production [29].
A very important parameter that must be considered in AD is the C/N ratio [92]. The C/N recommended by many studies is different, which is due to the different optimal C/N ratios for biogas production from different raw materials [33,38]. Too low C/N distiller cannot be digested anaerobically due to lack of nutrients, and too high C/N will limit the formation of new cells. Co-substrates such as straw and cow manure can be commonly added to regulate C/N and stimulate the growth of methanogens to maintain the stability of AD [7,33]. Some substrates such as cassava alcohol wastewater (CAW) can not only improve the C/N ratio of raw materials but also have a stronger synergistic effect [39].
Sulfur content in various forms is also discussed [59,93,94,95,96]. Sulfur is a nutrient required by methanogens, the optimum sulfur content is 1 to 25 mg/L, but the sulfur content in distillery grains is approximately 4 to 10 times the optimum concentration for AD [4,19,93]. Sulfur-rich distillers have a powerful inhibiting effect on methane production. There are two possible pathways of inhibition: (1) Sulfation occurs simultaneously with methanogenesis due to the presence of sulfate-reducing bacteria (SRB), and soluble sulfide inhibits methanogenic flora [11,27]. (2) The competition between SRB and methanogens reduces the conversion of organic matter to biogas. The effect of SO42−/COD on biogas production has been the focus of recent research and has been shown to have a huge correlation with stability [27,59,93]. Generally, a ratio of less than 0.1 does not pose a threat to stability [97]. With the increase of SO42−/COD, the process continues to deteriorate until no biogas is produced. It should be emphasized that at low SO42−/COD, the total sulfide will accumulate in the form of gas sulfide to ensure the stable production of biogas. The concentration of SO42− shall not exceed 3000 mg/L [93]. Competition between methanogenic archaea and SRB is an important topic in anaerobic treatment. The change in the COD/SO42− ratio does not change the result of competition between methanogens and SRB. However, SRB has an advantage in competing for electrons [97]. When the reaction lasts long enough, it produces a lot of sulfides. Controlling reaction time is also one of the measures to optimize system stability.
Total VFA concentrations are considered ideal indicators as they are the main methanogenic intermediates and catalyze methanogenesis, reflecting the balance between the four steps of AD [77,80]. It is well known that methane production usually goes up as VFAs break down. The process can be inhibited by decreasing the pH when the VFA concentration is too high [93]. The digestive process is less sensitive to VFA inhibition, but maintaining low VFA levels is important for the optimal performance of the AD process [25,46]. The concentration of a single VFA and the conversion between different VFAs have complex effects on the AD system. Propionic acid concentration of less than 1000 mg/L is more conducive to biogas production [7,25]. However, single VFA monitoring is expensive and requires some labor costs, which limits the detection of single VFA [77].
Alkalinity has also been proposed as a parameter for AD process monitoring, which can represent the buffer capacity of AD systems [40,77]. Alkalinity may come from the substrate itself or from compounds such as sodium bicarbonate being added. Although some experiments confirmed the stability of the system, a negative effect was observed due to the increase in alkalinity [16]. Alkalinity can be defined in many different ways. However, some studies have shown that IA/PA (intermediate alkalinity/partial alkalinity) is recommended as an indicator of the AD process due to its high sensitivity [38,77]. The two-stage anaerobic strategy can mitigate the effects of alkalinity fluctuations and has a more stable process [32]. Although the method for determining alkalinity is simple, the cost of balancing alkalinity is still an important factor to consider.
Phenol concentration has also been recently proposed as a stability index. Poirier et al. [79] studied the inhibitory effect of different phenol concentrations on AD and proposed phenol 0.50 g/L to 1.00 g/L as an early warning indicator since microbial recombination is initiated at this range but the final AD performance is not affected.
3.3. Methanogenic Archaea
Methane production is directly related to the activity of methanogenic archaea [98,99,100]. In this section, methanogenic archaea will be the focus of the discussion and the influence of operating parameters on them will be highlighted.
3.3.1. Diversity of Methanogenic Archaea
Regardless of the raw material used in the fermentation process, the low number of microorganisms detected is a common feature of all distillers due to the acidic pH and high temperature of the production process [99,100]. Anaerobic microbial communities can be categorized into two domains: bacteria and archaea. The archaeal domain is primarily responsible for methane production. The statistical methanogenic archaea communities are shown in Figure 1. They are strictly anaerobic microbes that can form methane from substrates such as carbon dioxide, formate, and acetate [101,102,103]. Methane-producing communities are stable over time [69].
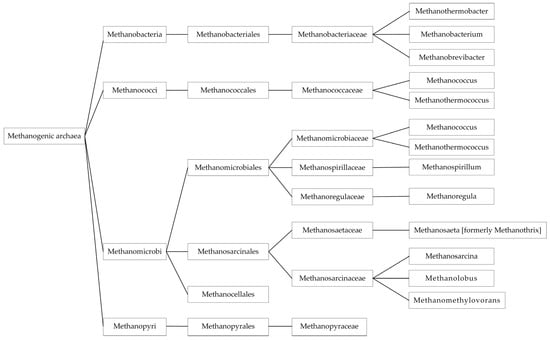
Figure 1.
Methanogenic archaea.
3.3.2. Effects of Operating Parameters on Archaeal Community
In this section, the effects of various process parameters on the AD of distillers were discussed and the reasons for the process changes were analyzed from the perspective of archaea. The relationship is shown in Figure 2. More data are in Table A2.
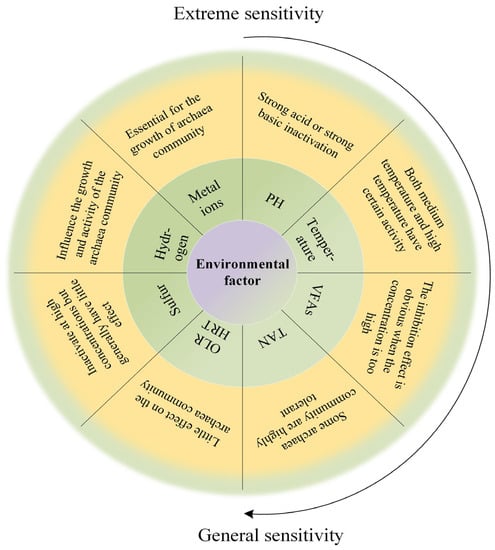
Figure 2.
Relationship between archaea activity and environmental factors.
pH
In all experiments aimed at producing biogas, almost all authors have controlled pH. This is because methanogenic archaea are extremely sensitive to pH and can be inhibited or even inactivated in highly acidic or alkaline environments. Acidic conditions will promote the hydrolysis of a distiller, but after acidic degradation, different archaea communities will be negatively affected to different degrees (such as Methanosaeta sp., Methanobacteriales, Methanomicrobiales, Methanococcales) [104,105]. Notably, the co-inhibition caused by pH may be the deep reason for the unsatisfactory process [29,32].
Ammonia/Ammonium
Microbes use ammonia/ammonium as a nitrogen source. However, high concentrations can be inhibitory or toxic. and are the two main forms in aqueous solutions and can be converted to each other depending on the pH, as shown in Equation (3) [68]:
The higher ammonium concentration inhibits the metabolism of hydrophilic methanogens through lead to VFA accumulation and a pH decrease, such as Methanobacterium and Methanospirillum [68], biogas production decreases or even stops production directly, which may also be related to the synergistic inhibition of high ammonium concentration and high VFAs concentration [85]. Methanogenic archaea have the lowest tolerance to ammonium inhibition compared with hydrolytic and acidogenic bacteria [78,87].
Free ammonia is much more toxic than ammonium. The influence mechanism of free ammonia on archaea is complex and varies with different substrates and operating conditions. In the AD of food waste, free ammonia leads to the accumulation of acetic acid and propionic acid, resulting in pH reduction and pH-related synergistic inhibition. It should be noted that a moderate increase in ammonia content will stimulate the development of vegetative hydrogen methanogens, resulting in desirable process results [68]. Peng et al. [29] continued to study the inhibition mechanism of free ammonia and believed that the influence of ammonia on the process was caused by the accumulation of VFAs. The instability of the process was rooted in the accumulation of acetate [6,24]. The relative abundances of archaea were generally inversely correlated with FAN, such as Methanosaetaceae and Methanosaeta, while Methanosarcina was positively correlated with FAN [32,67]. This indicated that the free ammonia concentration caused the transformation of the dominant archaea community, but the limited ability of the dominant archaea community to decompose acetic acid led to the process failure.
At short HRT and high OLR, the ammonia production rate is not sufficient for TAN accumulation, so it is difficult to observe TAN inhibition [51]. Although TAN tolerance of different substrates varies, co-inhibition due to high TAN can lead to process failure [85,87]. With the increase in TAN concentration, although the important role of the methanogenic meat family in acetic acid decomposition was observed, the archaea community remained at an overall inhibitory level [68].
Organic Load Rate and Hydraulic Retention Time
Although the archaea community in the distiller can work within a wide range of OLR and HRT, the sensitivity of different archaea communities is different. Overall, microbial activity was directly proportional to OLR and inversely proportional to HRT, and the hydrogenotrophic methanogens were the dominant methanogens at low OLR levels [51,67,106]. With the increase of OLR, acetoclastic methanogens were severely affected, such as Methanosarcina and Methanosaeta. However, there has been a significant increase in the relative abundance and activity of some archaea communities such as Methanobacterium [18,51]. Gamboa et al. [93] first identified the best Methanobacteriales and Methanosarcinales at the order level under optimal OLR, Methanobacterium, and Methanosaeta were found to produce methane. Cervantes et al. [16] confirmed the important role of Methanobacterium in high OLR. It is worth mentioning that Methanogenic archaea were severely inhibited under high OLR, which may be related to the accumulation of acetic acid and propionic acid in the reactor.
Hydrogen
Hydrogen partial pressure and hydrogen concentration are related to methane production. There are two metabolic pathways for the conversion of CO2 and H2, Equation (4):
Methanogens that consume H2 in low hydrogen pressure systems maintain low concentrations and facilitate propionate degradation [39]. With the increase of hydrogen partial pressure, although the acetoclastic methanogens and acetotrophic pathway are inhibited, it enriched certain archaea communities, such as Methanobacterium, Methanothrix, and Acetobacterium woodii to maintain the normal operation of the process [6,67,95].
Hydrogen concentration has a significant effect on the growth of archaea. Karadagli et al. [71] assessed the effect of hydrogen concentration on the growth of Methanobacterium bryantii M.o.H and found that when hydrogen concentration was greater than 2.4 μM, the net growth rate was positive. There is evidence that hydrogen is produced during the hydrolysis of glucose [107,108]. Increasing hydrogen concentration stimulated the acetoanaerobium genus and Methanobacterium, which enhanced the degradation of phenol, but Methanosaeta will decrease [67].
VFAs
Different volatile fatty acids have different toxicity to methanogens. There are few reports on formic acid in the AD process. Methanogens need trace elements to oxidize formic acid. The accumulation of formic acid in the absence of micronutrients will impede the degradation of propionate (feedback inhibition) and is related to the formation of hydrogen sulfide in biogas [33,38]. Figure 3 establishes the anaerobic mechanism under high acetic acid concentration. Studies have shown that methane production is highly correlated with acetic acid concentration [75]. As a major precursor to methane production, with the increase in total acetate concentration, total carbon conversion and methane production decreased [6,25]. Acetate concentration clearly dominates its metabolic pathway. High concentrations of acetic acid could inhibit acetoclastic methanogenesis (Methanosarcina and Methanosaeta) and affect the acetotrophic pathway [16,17]. However, acetic acid coexisted with acetone bacteria, SAOB, detritus acetone, and hydrogenotrophic methanogens even at inhibitory levels. Hydrogenotrophic methanogens are strengthened to varying degrees, which strengthens the SAO pathway to maintain methanogenesis [6,10,24]. Zhao et al. [25] found that the degradation of acetate is more difficult under high temperatures and high acetic acid concentrations, but this seems to be only applicable to simple substrates. Many experiments have observed that Methanothermobacter plays a key role in methane production at high temperatures [38,66].
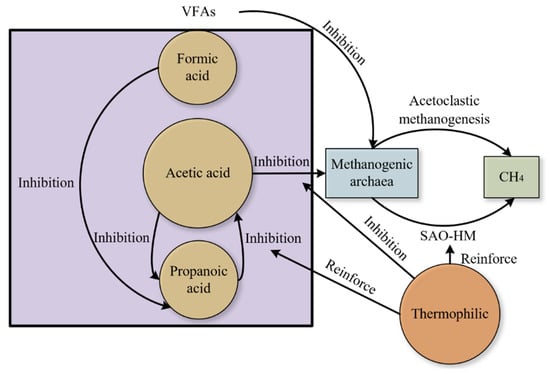
Figure 3.
Inhibition of methane production by high acetic acid concentration.
Propionic acid can be converted to acetic acid and hydrogen, and Equation (5) is:
Propionic acid is much more toxic than acetic acid, and its strong inhibitory effect at low concentrations has been observed [73]. Although the degradation of propionic acid is thermodynamically advantageous, it is easier to accumulate propionic acid under high-temperature AD of a distiller [66]. The degradation of propionic acid to methane is a slow process, and the appropriate extension of HRT has a greater influence on the degradation of propionic acid [25]. Butyrate was usually higher in distiller AD but was not significantly toxic to methanogenic bacteria [80].
Appropriate VFA concentrations can catalyze methane production, but high concentrations can also inhibit methane production. Inhibition concentration correlates with the synergistic effect of the substrate [7,46]. In the AD of the distiller, almost all archaea are inhibited under high VFAs, resulting in restricted methanogenesis, and the ability to break down substrates is reduced [38,107].
Temperature
Temperature is one of the most important parameters, as it can affect the activity of almost any microorganism. Processes at high temperatures seem more attractive than those at moderate temperatures, with the advantages of more complete destruction of viral and bacterial pathogens, and higher microbial activity [66,74,109]. Hydrogenotrophic methanogens (Methanoculleus and Methothermobacter) have higher activity and play a key role in the production of methane. However, the process using acetic acid as substrate was affected, mainly because the activity of Methanosaeate at high temperatures was inhibited [25,38]. However, there are different views: the increasing temperature will decrease microbial diversity, especially bacterial diversity, and mesophilic temperature is the optimal methanogenic temperature [74]. Methanobacterium is the main archaea at moderate temperatures and plays an important role in methane production [33,38]. The reactor at high temperatures is more sensitive to VFA, NH3, and alkalinity fluctuations. Anaerobic co-digestion in the medium temperature range is more stable, and microbial metabolism is more likely to reach equilibrium, which seems to be related to the activity of thermophilic bacteria [13,82]. Therefore, more authors use a two-stage reactor with high-temperature hydrolysis for acid production and medium-temperature methane production in order to obtain good process results [8,51].
Sulfur
COD/SO42− is significantly correlated with the inhibitory effect [95,97,110]. According to the literature, the best archaea orders are Methanobacteriales and Methanosarcinales at low COD/SO42− [27,93]. Subsequently, many authors examined the deeper changes of the archaeal community at different ratios and found that the content and activity of Methanosaeta were greatly affected by the increase of sulfur. Among them, Methanosaeta concilii’s ability to break down acetic acid was barely affected, Methanosaeta harundinacea rose slightly, while Methanosaeta thermophile disappeared [27,97]. This indicates that the concentration of SO42− has a certain selectivity for archaea. Although hydrogenotrophic methanogens are more active at low ratios, methanogens are generally at an inhibitory level [15,93]. When we looked at the competition between SRB and methanogens, the largest population of archaea community was Methanosaeta and its relative abundance increased during the long-term competition. Methanoregula disappeared and the relative abundance of total hydrogenotrophic methanogens decreased.
Metal Ions
The inhibitory effect on microorganisms in wastewater is mainly related to cations. Moderate concentrations of metal ions such as potassium and sodium will stimulate the growth of microorganisms, but excessive light metal ions will slow down the reproduction of microorganisms, inhibit their activity, and eventually destroy the stability of cell membranes [86,111]. Onodera et al. [72] assessed the effect of different concentrations of potassium on methane-producing activity (MPA) and found that the reduction of MPA was proportional to the concentration of potassium. The inhibition effect of a rapid increase (<1 min) was also greater than that of a gradual increase (>60 min). However, in the AD of the distiller, there was no significant change in potassium concentration before and after the experiment, which may be good for fertilizing [93]. The concentration of potassium had little effect on the structure of the archaea community. The relative abundance of Methanobacterium and Methanosaeta was almost unrelated to the concentration of potassium, but the overall reduction of archaea inhibited the anaerobic process. Lefebvre et al. [58] studied the effect of NaCl concentration on microbial communities. Only when NaCl concentration was higher than 20 g/L, were microorganisms seriously affected. Methanosaeteta sp. and M. beijingense were first recorded as salt-tolerant bacteria.
4. Archaea and Process Performance
4.1. Methanobacterium
Methanobacterium, a hydrogenotrophic methanogen, often appears in the AD of various substances. The ability of Methanobacterium to metabolize hydrogen is affected by a number of factors. Methanobacterium interact frequently with the outside world and can effectively decompose hydrogen produced by microorganisms such as Syntrophomonadaceae, Sedimentibacter, and Proteiniphilum at medium and low ammonia concentrations. It is important to stress that its activity is completely unaffected by low ammonia concentrations. However, high ammonium concentrations also inhibited its activity, and hydrogen accumulation was observed in the reactor [29,68]. However, these results were all observed at moderate temperatures. Methanobacterium often functions at moderate temperatures and produces desirable results through synergistic interaction with mesothermophilic bacteria. The proportion of the archaea community can be as high as 95%. However, it is almost undetectable at high temperatures and has nothing to do with the degradation of the distiller [25,38]. It should be stressed that Methanobacterium plays no significant role in the synergistic effect of co-digestion of the two distilleries [39]. It appears to be less sensitive to changes in environmental factors than other archaea [72,112], therefore, Methanobacterium can metabolize hydrogen efficiently under high OLR and low HRT [27,93]. Methanobacterium was also found to play an important role in phenol degradation recently, as shown in Figure 4.
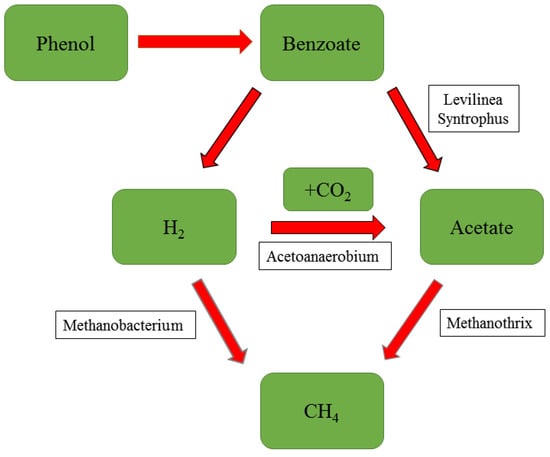
Figure 4.
Hydrogen enrichment enhances the anaerobic degradation of phenol [67] (The red line is the reinforced part).
4.2. Methanosaeta
Methanosaeta is a broken shell methanogen that strictly uses acetic acid as substrate and grows slowly. Acetotrophic pathways can be observed under various conditions [67,93,97]. In general, Methanosaeta’s ability to decompose acetic acid was negatively correlated with operating parameters [28]. What needs to be emphasized is that when TAN concentration is very low, Methanosaeta has higher activity and is not inhibited by ammonia. At high ammonium concentrations, the ability to decompose acetic acid is limited due to the synergistic inhibition of VFAs. However, with the enrichment of hydrogen, the inhibition effect under high ammonia can be effectively alleviated [67,79].
The concentration of acetate as the sole substrate had a significant effect on the activity of Methanosaeta. Numerous experiments have shown that Methanosaeta grows very slowly on acetic acid and is unable to decompose high concentrations of acetic acid [16,25,68]. This inhibitory effect can be mitigated by a synergistic effect of heating and substrate [39,113,114]. It should be noted that Methanosaeta has a lower affinity for acetate than Methanosarcina, although acetate is the only substrate for Methanosaeta [72].
The COD/SO42− ratio has no significant effect on Methanosaeta’s ability to decompose acetic acid in a short time, and the effect of Methanosaeta concilii on the decomposition of acetic acid cannot be ignored [93,97]. However, SRB was more active than Methanosaeta over time, resulting in a process dominated by sulfide generation [112]. This may have something to do with the dramatic reduction in Methanosaeta concilii, although some new colonies have emerged (Methanosaeta harundinacea, Methanobacterium beijingense). However, it is not enough to offset the negative effects caused by the disappearance of highly active colonies, leading to an unsatisfactory process.
4.3. Methanosarcina
Methanosarcina is a kind of mixotrophic methanogen with more substrates available and plays an important role in high-speed AD processes [79]. Owing to the advantages of cell globule, high volume ratio, and high surface ratio, ammonia could tolerate concentrations as high as 7000 mg/L, but it could not efficiently metabolize acetic acid under higher concentrations of ammonia [29,68]. Its methanogenesis has been observed in many experiments even in uncomfortable environments because it has a strong tolerance to environmental changes compared with other archaea communities [16,25,93]. However, Methanosarcina disappears at a phenol concentration of 5 g/L [79]. The activity of Methanosarcina can be enhanced by feeding low concentrations of acetic acid [24,90].
4.4. Methanoculleus
As a hydrogenotrophic methanogen, Methanoculleus mainly uses H2 and CO2 as substrates. Although its role in hydrogen metabolism can be observed at moderate temperatures [25], Methanoculleus plays a key role in the high-temperature AD of the distiller. The high relative abundance (up to 90%) is a guarantee of process quality at high temperatures and also has syntrophic acetic acid oxidation (SAO) and hydrogenotrophic methanogens [38,113,114,115].
5. Removal of Harmful Ingredients
5.1. Melanoidins
One of the problems of AD of the distiller is that melanoids cannot be degraded. It is generally believed that carbonyl compounds (reducing sugars) and amino compounds are formed by the Maillard reaction under certain conditions. The chromophores and the specific formation mechanism of melanoids have not yet been determined and need to be studied with more advanced techniques [86,116,117,118].
Melanoids pose a serious threat to soil and aquatic ecosystems by inhibiting or reducing microbial activity. Although the specific structure is not clear, the overall color is dark, with a pungent smell. The toxicity to humans is also significant [17].
The reaction substrates are important for the formation of melanoids [119,120]. Bork et al. [19] studied the reactivity of norfuraneol and glyoxal, glycolaldehyde, or acetalaldehyde to characterize the precursors of melanoids. It is found that all typical short-chain degradation products of norfuraneol play a key role in the formation of coloring agents, and the formation of polar pre-melanoidins was also found to be the most efficient for carbonyls, but the definite structure of the coloring products could not be determined due to technical reasons. The Maillard reaction occurs all the time in the winemaking process. Due to the uncertainty of melanoids structure and the complexity of the Maillard reaction, it is not feasible to find a reliable method from the winemaking process. Melanoids can be reduced by aerobic methods with physicochemical treatments (ozonation, flocculation, coagulation, active carbon adsorption), but the biggest problem with flocculation and coagulation is specificity. It is difficult to select the most suitable flocculant/coagulant due to different substrates and operating conditions. Adsorption is a surface-based physical phenomenon, and studies on adsorbents (such as powdered activated carbon (PAC), activated carbon, CTS, etc.) have also attracted attention, but these methods are costly and produce a lot of secondary pollutants [17,89]. Bioremediation seems to be a better option. It can effectively alleviate industrial pollutants. Microbial decolorization is a hot topic in recent years and the statistics are shown in Table 4 below. Although there are a lot of advantages of microbial decolorization, for example, the toxicity of distiller wastewater can be greatly reduced and laccase can be produced at the same time (phenolic compounds catalyzed oxidation) and other resources, the microbial decolorization efficiency is related to many factors, such as added nutrients (sodium chloride and potassium chloride), distiller concentration, culture conditions, etc. [8,100]. As a result, the decolorization efficiency varies greatly under different conditions, and the selection of the best microorganism becomes a problem. The influence of the interaction between different microorganisms on the degradation of melanoids is rarely reported. A coupling of various methods should be developed to deal with melanoids.
Table 4.
Study on decolorization of distillers by microorganisms.
5.2. Phenolic Compounds
Polyphenols are known to be toxic compounds. They are byproducts of the ethanol production process. Their concentrations are significantly affected by fermentation materials and the fermentation process [79,114,116].
Although these phenolic compounds have antioxidant and antimicrobial benefits [17], they are more notable for their harm in interfering with the activity of methanogenic archaea and even killing bacteria by (1) reaction with membrane cells, (2) inactivation of basic enzymes, and (3) disruption, and inactivation of cytogenetic functions. The concentration of polyphenols that produce inhibition/toxicity to methanogenic microorganisms during AD is not clearly defined. This can be attributed to environmental conditions, substrate properties, etc. Phenolic compounds are difficult to degrade by bioactivity and are phytotoxic, thus interfering with the decomposition of organic matter in the digester [58,84]. Because of its presence, it can lead to the inhibition or even inactivation of some communities, such as Syntrophomonadaceae, Methanosarcina, and Methanoculleus. which in turn affects the methanogenesis process and leads to a longer delay period [50,67,79].
Pretreatment of a distiller has long been shown to be effective in reducing phenolic substances [116]. After the pretreatment of the distiller with ozone (15 min), phenolic substances were reduced by more than 50%. After anaerobic treatment, the COD removal rate was almost not affected, and more methane was generated [89]. This demonstrates the potential for the biochemical treatment of wastewater. Some biotechnologies have certain advantages. The role of Clostridium, Proteobacteria and Chloroflexi, Trametes sp., Syntrophus, and Syntroporhabdus in polyphenol degradation has been demonstrated [17,63,100]. Wu et al. [67] used hydrogen-rich technology to alleviate the inhibitory effect of ammonia on AD of phenol-containing wastewater. Phenol can only be solved at very low hydrogen partial pressure under methanogenic conditions. The decomposition path is shown in Figure 4. This indicates a great potential for phenol removal in conjunction with methane production.
5.3. Sulfur
The transformation of sulfur is shown in Figure 5.
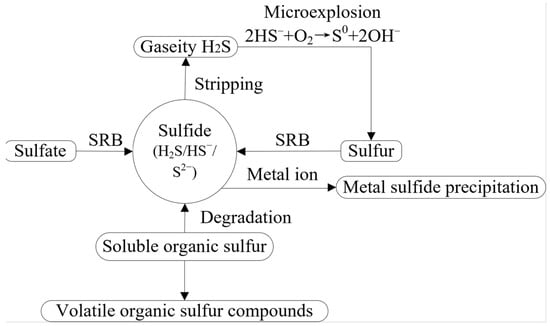
Figure 5.
Conversion of sulfur in AD [32,119].
5.3.1. Removal of H2S from Biogas
Among the compounds present in biogas, H2S is the most hazardous because even at low concentrations it can cause serious damage to equipment and harm the environment [11,15]. It has been reported that H2S concentrations of 150 mg/L inhibit methanogens [27].
Krayzelova et al. [123] applied micro-aeration to remove hydrogen sulfide in the UASB reactor. In the long-term operation, the H2S removal efficiency was 73%. What is more beneficial is that the micro-aeration did not lead to the reduction of COD removal rate or methanogenic activity. There are many other benefits of micro-aeration, such as sulfur recovery and enhanced hydrolysis process. Microaeration has the potential to remove large amounts of hydrogen sulfide from biogas [119]. Two-stage AD can also be used to reduce H2S content in biogas [26,32].
5.3.2. Removal of Sulfate and Sulfide
There are many reasons for the formation of sulfides, such as the degradation of organic sulfur and the reduction of sulfate. Sulfides have been reported to range from 30 to 250 mg/L [59,93].
It should be noted that an increase in the sulfate concentration of the influents does not necessarily increase the total sulfide produced, but rather the formation of sulfate and sulfur-containing precipitates in the effluent, which is inconsistent with Jiménez et al. [27], probably due to the use of a two-stage anaerobic membrane bioreactor in Silva et al. [15], which limits microbial activity. Gamboa et al. [93] achieved a 100% sulfate removal rate through the improved UASB reactor, but the COD removal rate was not high. As a co-matrix, waste oils form unsaturated LCFA (long-chain fatty acids) and mitigate the negative effects of increased sulfide levels on methane formation; however, this may lead to reduced methane production from waste oils [121,124]. For the distiller, the only source of sulfate is sulfuric acid added in the fermentation stage, so the ethanol fermentation stage can also be improved, and the sulfate concentration in the distiller can be reduced by finding suitable antibiotics and other products to replace sulfuric acid. Sulfide generation can also be reduced by using different operating parameters, such as COD/SO42−, OLR, etc. [15,97,111].
5.3.3. Inhibition of Sulfate-Reducing Bacteria Activity
SRB removes a high proportion of organic matter in the first few steps of the AD process to produce hydrogen sulfide, carbon dioxide, and acetic acid, and numerous studies have focused on factors influencing SRB activity [15,27,125].
Reasonable control of SO42−/COD can inhibit the activity of SRB. Jiménez et al. [27] treated a distiller rich in sulfate with AD and found that SRB was inhibited more and more strongly with the increase of SO42−/COD, but the proportion of organic matter degraded by sulfate-reducing bacteria increased to 27.1 ± 0.6%. However, the experimental results of Hu et al. [97] show that the change of SO42−/COD does not change the result of competition between MPA and SRB, which may be the result of using different substrates. This indicates that the synergistic effect of different substrates will affect the competition between MPA and SRB. The processing time is also important for SRB because of its competitive advantage over methanogens in terms of electron donors and its substrate adaptability [95]. Although some methanogenic archaea are affected by elevated temperatures, methanogenic archaea are more active than SRB at elevated temperatures [37].
6. Future Prospects for Distiller Treatment
6.1. Integration with Membrane Separation Technology
Many authors emphasize the necessity of combining with the membrane system [8,15,126]. Combining AD with the membrane system to optimize AD: the advantage of this technology is that the microorganisms in the bioreactor can be completely retained, which is conducive to the growth of microorganisms. It is more conducive to methane production [11,64]. While maintaining a high clearance rate, other resources can also be recovered [15,127].
6.2. Inactivated Pathogens
When distillers were used as one of the substrates to participate in the anaerobic process, Escherichia coli and salmonella were removed well below the limits set by European law [12,51]. In the future, some key microorganisms should be studied to better understand the clearance mechanism.
6.3. Coupling with Built-In Microbial Batteries
Bioelectrochemical systems (BES) can affect the growth and activity of microorganisms. Some studies have emphasized that combining anaerobic digesters with BES can increase biogas production [128]. In addition, BES can not only improve the system stability but also restore the anaerobic system [94]. BES treatment of a distiller seems to be a good choice.
6.4. Pretreatment
Different pretreatments of a distiller have great development prospects. Using different pretreatments, such as ozone oxidation, enzyme treatment, and Advanced oxidation process (AOP), the microbial activity was enhanced to varying degrees and the association between bacteria and archaea became closer, ultimately achieving desirable results [40,89,129,130]. The presence of certain chemicals can hinder methane production, and studies at the microbial level help the prospects for pretreatment applications [131,132]. Different raw materials have different pretreatment methods [133].
7. Conclusions
The AD of a distiller has a lot of potential for energy, but more research on microorganisms is needed to increase the rate at which energy is converted. This paper sets the foundation for microbial management by summarizing the fundamental knowledge about the archaea community. Although more and more researchers have observed and discussed microbial colonies, there is a lack of studies on the role of microbial continuity. Therefore, more researchers are required to pay attention to the role of microbial continuity. More attention should also be paid to the relationship between microbial structure and activity and the environment. Additionally, to better understand the changes in microbial community structure and metabolic activity, conventional biomolecular approaches must be enhanced. More research at the microbial level is the future direction of development.
Author Contributions
Conceptualization, G.L. and F.X.; methodology, F.X. and T.Y.; validation, G.L., F.X. and T.Y.; formal analysis, F.X. and T.Y.; resources, G.L.; data curation, F.X. and T.Y.; writing—original draft preparation, F.X., T.Y. and X.W.; writing—review and editing, F.X., T.Y., X.W., G.L. and T.L.; supervision, G.L. and Z.H.; funding acquisition, Z.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (Grant NO. 32172277).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We wish to thank the National Natural Science Foundation of China (32172277) and Beijing Technology and Business University for their support.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1.
Anaerobic co-digestion.
Table A1.
Anaerobic co-digestion.
| Substrate | Optimal Proportion | Synergy | Energy Potential of Distillers | Ref. |
|---|---|---|---|---|
| sewage sludge wine vinasse poultry manure | 49.5:49.5:1 | 1. pH is stable at neutral 2. The process effect is best at short HRT (13 days) | With the addition of wine vinasse, the process performance improved and methane production increased | [13] |
| 1. sugar beet vinasse 2. sugar beet vinasse, cow manure 3. sugar beet vinasse, straw. | 1. AD of sugar beet vinasse alone was not possible 2. The average values were 323.1 ± 48.6 mL CH4 g VS−1 3. The average values were 287.7 ± 22.2 mL CH4 g VS−1 | 1. BMP of sugar beet vinasse is 267.4 ± 4.5 L CH4 kg VS−1 2. Average VS/DM ratio is 71.5 ± 2.5% | [33] | |
| sugar beet pulp silage molasses vinasse | 3:1 | 1. The highest biogas productivity (598.1 mL/g VS) was achieved at the SBPS–vinasse ratio of 3:1 2. Biogas yield of SBPS and vinasse fermented alone decreased by 13% and 28.6%, respectively | SBPS contained 18.71% TS and 93.8% VS vinasse (6.75% TS and 75.11% VS) | [37] |
| sugarcane vinasse filter cake deacetylation liquor | Organic matter removal rate and methane production were the highest | High volatile fatty acid content | [38] | |
| sewage sludge wine vinasse poultry manure | 49.5:49.5:1 | 1. The co-digestion of three substrates was more advantageous 2. Relieves the inhibition of the high TAN | BMP increases with the addition of wine vinasse | [47] |
| sludge wine vinasse poultry manure | 49.5:49.5:1. | 1. In the experiment of 10 g/L poultry manure, H2 production, and biogas production were 18.20% and 27.57% higher, respectively, compared with 20 g/L. 2. The accumulation of TAN and FAN was alleviated | BHP tests: 22.34 mL H2/g SCOD and 27.1 mL H2/g VS | [48] |
| sludge wine vinasse | 25:75 | Under the optimum ratio, the highest hydrogen yield was 43.25 ± 1.52 mL H2/g VS, which was 14 times higher than that of sludge single fermentation | With the increase of wine vinasse in the mixture, hydrogen production increased significantly | [49] |
| sugarcane vinasse filter cake deacetylation liquor | 1. Compared with traditional AD, energy efficiency is improved by at least 16% 2. pH is stable at neutral | Single vinasse has high BMP, the mixture of BMP increased by about 38% | [50] | |
| sludge wine vinasse poultry manure | 49.5:49.5:1 | The VS and total volatile fatty acids reached, respectively, 93.13% and 97.43% of removal efficiency | With the addition of wine vinasse, the process performance improved and methane production increased | [51] |
| FOG slaughterhouse wastewater | FOG concentrations are 5–10% | 1. Methane production increased by a factor of two to five 2. The biodegradability of almost all substrates was improved | [52] | |
| food waste chicken manure | 1. Co-digestion improves system productivity for almost all parameters 2. The synergistic effect of microorganisms was obvious | [53] | ||
| sugarcane vinasse urea trace elements | 1. Stable operation under high OLR and low HRT 2. 79% higher methane production rate with a stable specific methane production of 239 mL g COD−1 | [54] | ||
| cow dung anaerobic granular sludge activated sludge food waste | 1:1:1:1 | 1. Optimal co-digested inoculum for biological methanation 2. The kinetics of organic degradation produces a scaling effect | [55] | |
| sugarcane vinasse chicken manure | 3:1 | 1. Co-digestion relieved the inhibition of TVFA and TAN 2. Co-digestion had the highest hydrolytic activity | BMP increased with the increase of vinasse | [56] |
| sewage sludge Sherry-wine distillery wastewater | 1:1 | 1. High methane production:154 L CH4/kg COD 2. The archaea were enriched in co-digestion | [57] |
Table A2.
Effect of operating conditions on methanogenic archaea.
Table A2.
Effect of operating conditions on methanogenic archaea.
| Order | Type | Available Substrate | Other Features | |
|---|---|---|---|---|
| Methanomicrobiales | hydrogenotrophic methanogens | H2 | Little affected by temperature and acetic acid concentration It is negatively correlated with SO42−/COD Concentrations of high total ammonia and salt showed a clearer effect than Methanobacteriales Higher acid resistance than Methanosaeta sp., Methanobacteriales, Methanococcales | [25] [27] [69] [104] |
| Methanobacteriales | hydrogenotrophic methanogens | H2, CO2, formic acid | Strictly hydrogenotrophic methanogens’; syntrophic acetate oxidation and hydrogenotrophic methanogenesis Greatly affected by temperature and acetic acid concentration When SO42−/COD is 0.05, the dominant bacterial order; it is greatly influenced by SO42−/COD and negatively correlated It is widely distributed and positively correlated with VFA, OLR, and temperature Optimal methanogenesis at optimal OLR (17.05 kg COD/m3-day) | [6] [25] [27] [69] [93] |
| Methanococcales | H2 | Cover most methanogens encountered in anaerobic digesters | [104] | |
| Methanosarcinales | Greatly affected by acetic acid concentration and not sensitive to temperature Optimal methanogenesis at optimal OLR (17.05 kg COD/m3-day) | [25] [93] | ||
| Family | ||||
| Methanosaetaceae | acetate | High acetic acid concentration is suitable for growth and inversely proportional to temperature In contrast to Methanomicrobiales and Methanobacteriales, the ratio of SO42−/COD has little influence Methanosaetaceae was negatively correlated with FAN, total ammonia, volatile fatty acids, and conductivity; however, it was positively related to Methanosarcinaceae and played a crucial role in low total ammonia, salt, and volatile fatty acid In general, a filamentous shape | [25] [27] [29] [69]. | |
| Methanosarcinaceae | acetic acid, methanol, other methylated C1 compounds, H2, CO2, CO | When growing on acetic acid, it is greatly affected by temperature Methanosarcinaceae was positively correlated with Methanosaetaceae and negatively correlated with total ammonia, volatile fatty acids, and conductivity; Methanosarcinaceae have a spherical form Multicellular clusters Relative abundance increased with increasing TAN Plays a key role in promoting methane production from biological carbon The most robust methanogen from metabolic and physiologic points May grow on the C1 compound in the absence of hydrogen | [25] [29] [69] [68] [75] [119] [120] | |
| Genus | ||||
| Methanobacterium | hydrogenotrophic methanogens | CO2, H2, formic acid | The relative abundance increases with the increase of OLR Frequent interaction with the outside world and high acid and ammonia resistance Mainly in mesophilic temperatures(30–35 °C) Methanobacterium had no obvious influence on mono-digestion and co-digestion. Relative abundance increased with increasing hydrogen concentration Little affected by potassium concentration Relative abundance was positively correlated with low ammonia concentration and negatively correlated with high ammonia concentration Little change in long-term competition with SRB It decreases with the addition of CTS | [16] [29] [38] [39] [67] [72] [68] [112] [134] |
| Methanosaeta | acetoclastic methanogens | acetate | Decreased relative abundance under high OLR and high acetic acid In 20 g-COD/L acetate reactors, the relative abundance at high temperatures decreased, accounting for only 0.07% of the archaea community FAN, pH, and TA were all negatively correlated with Methanosaeta Methanosaeta was more sensitive to high concentrations of acetate than Methanosarcina Halt-tolerant Archaea Under the condition of rich hydrogen, decrease; Active acetyl nutrient methanogenesis at high ammonia concentrations Lower affinity for acetate than Methanosarcina When the TAN level was lower than 560 mg/L, Methanosaeta was more active and not inhibited by ammonia; the activity is inversely proportional to the concentration of ammonium and acetic acid At high COD/SO42−ratios, playing a major role With the decrease of COD/SO42−, the quantity increases and the growth rate is higher than Methanosarcina The relative abundance of Methanosaeta increases dramatically during long-term competition with SRB At high OLR, the activity is severely inhibited, which can be alleviated by adding CAW; A broken shell methanogen; Slow-growing; Addition of CTS increases relative abundance | [16] [25] [29] [39] [58] [67] [72] [68] [93,97] [95] [112] [134] |
| Methanosarcina | mixotrophic methanogen | methanol, methylamine acetate H2, CO2 | Decreased relative abundance at high OLR and high acetic acid Feeding acetate at a concentration of 10 mM resulted in enrichment At lower acetate concentrations (20 g-COD/L), dominant methanogen, 84%(mesophilic) and 88% (thermophilic) At high ammonia concentrations, acetate cannot be efficiently metabolized It is negatively correlated with ammonia concentration, but the tolerance concentration of ammonia is as high as 7000 mg/L; survives in a weakly acidic environment Methanosarcina disappears at phenol concentration of 5 g/L Grows slowly on acetate With the decrease of COD/SO42−, the quantity increases, and the growth rate is lower than Methanosaeta Involved in multiple methanogenic pathways Drop after adding CTS It was involved in the recovery after inhibitory events with high levels of acetate The effect of Methanosarcina was more obvious under thermo-alkali pretreatment | [16] [24] [25] [29] [68] [79] [92] [95] [103] [134] [113] [115] |
| Methanoculleus | hydrogenotrophic methanogens | mainly H2, CO2 | At medium temperature, plays an important role; The relative abundance increased with the concentration of acetate, and the increase in temperature Major genus of thermophilic processes, capable of co-trophic acetic acid oxidation (SAO) and hydrotrophic methanogenesis Methanoculleus is a mesophilicarchaea; may survive in a weakly acidic environment Disappear when phenol concentration is 5 g/L With the increase in phenol concentration (0.50 g/L to 2.00 g/L), it became the dominant community | [25] [38] [68] [79] [113] |
| Methanothermobacter | hydrogenotrophic methanogens | H2, CO2 | In high acetic acid concentrations (60 g-COD/L) and high temperatures become the dominant community (94% of the archaeal community); Participate in the SAO-HM pathway Major Genera in thermophilic processes Emerged in the long-running competition with SRB | [25] [38] [119] |
| Methanoregula | hydrogenotrophic methanogens | H2 | Disappeared from the long competition with SRB | [119] |
| Methanofollis | H2 | Disappeared from the long competition with SRB | [119] | |
| Methanospirillum | hydrogenotrophic methanogens | Disappear as the temperature rises When the concentration of ammonium accumulates to 6 g/L, the metabolism is inhibited Addition of CTS increases relative abundance | [25] [68] [134] | |
| Methanobrevibacter | hydrogenotrophic methanogens | H2, CO2, formate | Increases with the increase of hydrogen Relative abundance decreased with addition of CTS | [36] [134] |
| Species | ||||
| Methanosaeta concilii | acetoclastic methanogen | acetate | The optimal sodium concentration is <60 mM It does not change with COD/SO42 and plays an important role in the consumption of acetate as an energy source for methane production Reduced in long-term competition with SRB (40.0%-15.2%) | [72] [97] [112] |
| Methanosaeta thermophile | acetoclastic methanogen | acetate | The optimal sodium concentration is <130 mM Disappeared with the decrease of COD/SO42− | [72] [97] |
| Methanosaeta harundinacea | acetoclastic methanogen | acetate | The optimal sodium concentration is <20 mM It increases slightly with the decrease of COD/SO42− Significant increase in long-term competition with SRB (2.5–35.2%) | [72] [97] [112] |
| Methanoregula formicicum | hydrogenotrophic methanogen | H2 | Disappeared from the long competition with SRB | [112] |
| Methanoregula boonei | hydrogenotrophic methanogen | H2 | Disappeared from the long competition with SRB | [112] |
| Methanobacterium petrolearium | H2 | Emerging in long-term competition with SRB (0–6.9%) | [112] | |
| Methanobacterium beijingense | H2 | Salt tolerance Emerging in long-term competition with SRB (0–4.8%) | [58] [112] | |
| Methanothermobacter tenebrarum | H2 | Emerging from a long rivalry with the SRB | [112] | |
| Methanosarcina mazei | acetate | Occurs in long-term competition with SRB, but in low relative abundance | [112] | |
| Methanosarcina acetivorans | M. acetivorans may grow nonmethanogenically, using CO as a substrate | [103] | ||
| Methanomethylovoran hollandica | methanol, methylamines acetate | Occurs in long-term competition with SRB, but in low relative abundance | [112] | |
| Methanofollis liminatans | hydrogenotrophic methanogen | H2 | Occurs in long-term competition with SRB, but in low relative abundance | [112] |
References
- Li, G.; Hao, Y.; Yang, T.; Xiao, W.; Pan, M.; Huo, S.; Lyu, T. Enhancing bioenergy production from the raw and defatted microalgal biomass using wastewater as the cultivation medium. Bioengineering 2022, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hossen, E.H.; Aziz, T.N.; Ducoste, J.J.; de Los Reyes, F.L. Increased loading stress leads to convergence of microbial communities and high methane yields in adapted anaerobic co-digesters. Water Res. 2020, 169, 115155. [Google Scholar] [CrossRef] [PubMed]
- Toshinari, M.; Sarah, S.; Viviana, S.; Yuki, H.; Shotaro, T. Engineering anaerobic digestion via optimizing microbial community: Efects of bactericidal agents, quorum sensing inhibitors, and inorganic materials. Appl. Microbiol. Biotechnol. 2021, 105, 7607–7618. [Google Scholar]
- Janke, L.; Leite, A.; Nikolausz, M.; Schmidt, T.; Liebetrau, J.; Nelles, M.; Stinner, W. Biogas Production from Sugarcane Waste: Assessment on Kinetic Challenges for Process Designing. Int. J. Mol. Sci. 2015, 16, 20685–20703. [Google Scholar] [CrossRef] [PubMed]
- Gao, N.; Milandile, M.H.; Quan, C.; Rundong, L. Critical assessment of plasma tar reforming during biomass gasification: A review on advancement in plasma technology. J. Hazard. Mater. 2022, 421, 126764. [Google Scholar] [CrossRef] [PubMed]
- Rachbauer, L.; Beyer, R.; Bochmann, G.; Fuchs, W. Characteristics of adapted hydrogenotrophic community during biomethanation. Sci. Total Environ. 2017, 595, 912–919. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Jin, K.; Yi, R.; Chen, M.; Peng, J.; Pan, Y. Enhancement of bioenergy recovery from agricultural wastes through recycling of cellulosic alcoholic fermentation vinasse for anaerobic co-digestion. Bioresour. Technol. 2020, 311, 123511. [Google Scholar] [CrossRef]
- Hoarau, J.; Caro, Y.; Grondin, I.; Petit, T. Sugarcane vinasse processing: Toward a status shift from waste to valuable resource. A review. J. Water Process Eng. 2018, 24, 11–25. [Google Scholar] [CrossRef]
- Buller, L.S.; Romero, C.; Lamparelli, R.A.C.; Ferreira, S.F.; Bortoleto, A.P.; Mussatto, S.I.; Forster-Carneiro, T. A spatially explicit assessment of sugarcane vinasse as a sustainable by-product. Sci. Total Environ. 2021, 765, 142717. [Google Scholar] [CrossRef]
- Lopes, S.I.; Capela, M.I.; Lens, P.N. Sulfate reduction during the acidification of sucrose at pH 5 under thermophilic (55 degrees C) conditions. II: Effect of sulfide and COD/ SO42-ratio. Bioresour. Technol. 2010, 101, 4278–4284. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Brasil, Y.L.; Koch, K.; Amaral, M.C.S. Resource recovery from sugarcane vinasse by anaerobic digestion—A review. J. Environ. Manag. 2021, 295, 113137. [Google Scholar] [CrossRef] [PubMed]
- Costa, K.C.; Leigh, J.A. Metabolic versatility in methanogens. Curr. Opin. Biotechnol. 2014, 29, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Sillero, L.; Solera, R.; Perez, M. Improvement of the anaerobic digestion of sewage sludge by co-digestion with wine vinasse and poultry manure: Effect of different hydraulic retention times. Fuel 2022, 321, 124104. [Google Scholar] [CrossRef]
- Rodrigues Reis, C.E.; Hu, B. Vinasse from Sugarcane Ethanol Production: Better Treatment or Better Utilization? Front. Energy Res. 2017, 5, 7. [Google Scholar] [CrossRef]
- Silva, A.F.R.; Magalhaes, N.C.; Cunha, P.V.M.; Amaral, M.C.S.; Koch, K. Influence of COD/SO42-ratio on vinasse treatment performance by two-stage anaerobic membrane bioreactor. J. Environ. Manag. 2020, 259, 110034. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Cervantes, A.; Guevara-Santos, N.; Arreola-Vargas, J.; Snell-Castro, R.; Méndez-Acosta, H.O. Performance and microbial dynamics in packed-bed reactors during the long-term two-stage anaerobic treatment of tequila vinasses. Biochem. Eng. J. 2018, 138, 12–20. [Google Scholar] [CrossRef]
- Chowdhary, P.; Raj, A.; Bharagava, R.N. Environmental pollution and health hazards from distillery wastewater and treatment approaches to combat the environmental threats: A review. Chemosphere 2018, 194, 229–246. [Google Scholar] [CrossRef]
- Kainthola, J.; Kalamdhad, A.S.; Goud, V.V. A review on enhanced biogas production from anaerobic digestion of lignocellulosic biomass by different enhancement techniques. Process Biochem. 2019, 84, 81–90. [Google Scholar] [CrossRef]
- Bork, L.V.; Haase, P.T.; Rohn, S.; Kanzler, C. Structural characterization of polar melanoidins deriving from Maillard reaction intermediates—A model approach. Food Chem. 2022, 395, 133592. [Google Scholar] [CrossRef]
- da Silva Neto, J.V.; Gallo, W.L.R.; Nour, E.A.A. Production and use of biogas from vinasse: Implications for the energy balance and GHG emissions of sugar cane ethanol in the brazilian context. Environ. Prog. Sustain. Energy 2019, 39, 13226. [Google Scholar] [CrossRef]
- Peiter, F.S.; Hankins, N.P.; Pires, E.C. Evaluation of concentration technologies in the design of biorefineries for the recovery of resources from vinasse. Water Res. 2019, 157, 483–497. [Google Scholar] [CrossRef]
- Fuess, L.T.; Garcia, M.L. Bioenergy from stillage anaerobic digestion to enhance the energy balance ratio of ethanol production. J. Environ. Manag. 2015, 162, 102–114. [Google Scholar] [CrossRef]
- Fuess, L.T.; Garcia, M.L. Implications of stillage land disposal: A critical review on the impacts of fertigation. J. Environ. Manag. 2014, 145, 210–229. [Google Scholar] [CrossRef]
- Dyksma, S.; Jansen, L.; Gallert, C. Syntrophic acetate oxidation replaces acetoclastic methanogenesis during thermophilic digestion of biowaste. Microbiome 2020, 8, 105. [Google Scholar] [CrossRef]
- Zhao, J.; Westerholm, M.; Qiao, W.; Yin, D.; Bi, S.; Jiang, M.; Dong, R. Impact of temperature and substrate concentration on degradation rates of acetate, propionate and hydrogen and their links to microbial community structure. Bioresour. Technol. 2018, 256, 44–52. [Google Scholar] [CrossRef]
- Ramos, L.R.; Lovato, G.; Rodrigues, J.A.D.; Silva, E.L. Anaerobic digestion of vinasse in fluidized bed reactors: Process robustness between two-stage thermophilic-thermophilic and thermophilic-mesophilic systems. J. Clean. Prod. 2021, 314, 128066. [Google Scholar] [CrossRef]
- Jiménez, J.; Barrera, E.L.; De Vrieze, J.; Boon, N.; DeMeester, S.; Spanjers, H.; Romero, O.R.; Dewulf, J. Microbial community dynamics reflect reactor stability during the anaerobic digestion of a very high strength and sulfate-rich vinasse. J. Chem. Technol. Biotechnol. 2018, 93, 975–984. [Google Scholar] [CrossRef]
- Cremonez, P.A.; Teleken, J.G.; Weiser Meier, T.R.; Alves, H.J. Two-Stage anaerobic digestion in agroindustrial waste treatment: A review. J. Environ. Manag. 2021, 281, 111854. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, S.; Li, L.; Zhao, X.; Ma, Y.; Shi, D. Long-term high-solids anaerobic digestion of food waste: Effects of ammonia on process performance and microbial community. Bioresour. Technol. 2018, 262, 148–158. [Google Scholar] [CrossRef]
- Prangya, R.; Daya, S.; Macsen, H.; Caitlin, B.; Helmer, L.; Reddicherla, U.; Sanjay, M.; Sagarika, P.; Mukesh, G. Sustainable Valorisation of Animal Manures via Thermochemical Conversion Technologies: An Inclusive Review on Recent Trends. Waste Biomass Valorization 2022, 14, 553–582. [Google Scholar]
- Umapathi, R.; Park, B.; Sonwal, S.; Rani, G.M.; Cho, Y.J.; Huh, Y. Advances in optical-sensing strategies for the on-site detection of pesticides in agricultural foods. Trends Food Sci. Technol. 2022, 119, 69–89. [Google Scholar] [CrossRef]
- Santana Junior, A.E.; Duda, R.M.; Oliveira, R.A.D. Improving the energy balance of ethanol industry with methane production from vinasse and molasses in two-stage anaerobic reactors. J. Clean. Prod. 2019, 238, 117577. [Google Scholar] [CrossRef]
- Moraes, B.S.; Triolo, J.M.; Lecona, V.P.; Zaiat, M.; Sommer, S.G. Biogas production within the bioethanol production chain: Use of co-substrates for anaerobic digestion of sugar beet vinasse. Bioresour. Technol. 2015, 190, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Carpanez, T.G.; Moreira, V.R.; Assis, I.R.; Amaral, M.C.S. Sugarcane vinasse as organo-mineral fertilizers feedstock: Opportunities and environmental risks. Sci. Total Environ. 2022, 832, 154998. [Google Scholar] [CrossRef]
- Sydney, E.B.; Carvalho, J.C.; Letti, L.A.J.; Magalhaes, A.I., Jr.; Karp, S.G.; Martinez-Burgos, W.J.; Candeo, E.S.; Rodrigues, C.; Vandenberghe, L.P.S.; Neto, C.J.D.; et al. Current developments and challenges of green technologies for the valorization of liquid, solid, and gaseous wastes from sugarcane ethanol production. J. Hazard. Mater 2021, 404, 124059. [Google Scholar] [CrossRef]
- Buenavista, R.M.E.; Siliveru, K.; Zheng, Y. Utilization of Distiller’s dried grains with solubles: A review. J. Agric. Food Res. 2021, 5, 100195. [Google Scholar] [CrossRef]
- Zieminski, K.; Kowalska-Wentel, M. Effect of enzymatic pretreatment on anaerobic co-digestion of sugar beet pulp silage and vinasse. Bioresour. Technol. 2015, 180, 274–280. [Google Scholar] [CrossRef]
- Volpi, M.P.C.; Junior, A.; Franco, T.T.; Moraes, B.S. Operational and biochemical aspects of co-digestion (co-AD) from sugarcane vinasse, filter cake, and deacetylation liquor. Appl. Microbiol. Biotechnol. 2021, 105, 8969–8987. [Google Scholar] [CrossRef]
- Jiang, Q.; Xin, Y.; Jiang, Y.; Huang, L.; Shen, P. Improving the efficiency of anaerobic digestion of Molasses alcohol wastewater using Cassava alcohol wastewater as a mixed feedstock. Bioresour. Technol. 2022, 344, 126179. [Google Scholar] [CrossRef]
- Mendez-Acosta, H.O.; Snell-Castro, R.; Alcaraz-Gonzalez, V.; Gonzalez-Alvarez, V.; Pelayo-Ortiz, C. Anaerobic treatment of Tequila vinasses in a CSTR-type digester. Biodegradation 2010, 21, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Oñate, J.; Arenas, A.; Ruiz, A.; Rivera, K.; Peláez, C. Evaluation of Mutagenic and Genotoxic Activity in Vinasses Subjected to Different Treatments. Water Air Soil. Pollut. 2015, 226, 144. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, Y.; Wang, H.; Zhao, X.; Cui, H.; Wei, Z. Reducing nitrogen loss and phytotoxicity during beer vinasse composting with biochar addition. Waste Manag. 2017, 61, 150–156. [Google Scholar] [CrossRef]
- Fuess, L.T.; Garcia, M.L.; Zaiat, M. Seasonal characterization of sugarcane vinasse: Assessing environmental impacts from fertirrigation and the bioenergy recovery potential through biodigestion. Sci. Total Environ. 2018, 634, 29–40. [Google Scholar] [CrossRef]
- Tena, M.; Buller, L.S.; Sganzerla, W.G.; Berni, M.; Forster-Carneiro, T.; Solera, R.; Pérez, M. Techno-economic evaluation of bioenergy production from anaerobic digestion of by-products from ethanol flex plants. Fuel 2022, 309, 122171. [Google Scholar] [CrossRef]
- Longati, A.A.; Lino, A.R.A.; Giordano, R.C.; Furlan, F.F.; Cruz, A.J.G. Biogas Production from Anaerobic Digestion of Vinasse in Sugarcane Biorefinery: A Techno-economic and Environmental Analysis. Waste Biomass Valorization 2019, 11, 4573–4591. [Google Scholar] [CrossRef]
- Kondusamy, D.; Kalamdhad, A.S. Pre-treatment and anaerobic digestion of food waste for high rate methane production—A review. J. Environ. Chem. Eng. 2014, 2, 1821–1830. [Google Scholar] [CrossRef]
- Sillero, L.; Solera, R.; Perez, M. Biochemical assays of potential methane to test biogas production from dark fermentation of sewage sludge and agricultural residues. Int. J. Hydrogen Energy 2022, 47, 13289–13299. [Google Scholar] [CrossRef]
- Sillero, L.; Solera, R.; Perez, M. Anaerobic co-digestion of sewage sludge, wine vinasse and poultry manure for bio-hydrogen production. Int. J. Hydrogen Energy 2022, 47, 3667–3678. [Google Scholar] [CrossRef]
- Tena, M.; Luque, B.; Perez, M.; Solera, R. Enhanced hydrogen production from sewage sludge by cofermentation with wine vinasse. Int. J. Hydrogen Energy 2020, 45, 15977–15984. [Google Scholar] [CrossRef]
- Volpi, M.P.C.; Brenelli, L.B.; Mockaitis, G.; Rabelo, S.C.; Franco, T.T.; Moraes, B.S. Use of Lignocellulosic Residue from Second-Generation Ethanol Production to Enhance Methane Production Through Co-digestion. BioEnergy Res. 2021, 15, 602–616. [Google Scholar] [CrossRef]
- Sillero, L.; Solera, R.; Pérez, M. Thermophilic-mesophilic temperature phase anaerobic co-digestion of sewage sludge, wine vinasse and poultry manure: Effect of hydraulic retention time on mesophilic-methanogenic stage. Chem. Eng. J. 2023, 451, 138478. [Google Scholar] [CrossRef]
- Agabo-Garcia, C.; Solera, R.; Perez, M. First approaches to valorizate fat, oil and grease (FOG) as anaerobic co-substrate with slaughterhouse wastewater: Biomethane potential, settling capacity and microbial dynamics. Chemosphere 2020, 259, 127474. [Google Scholar] [CrossRef] [PubMed]
- Chuenchart, W.; Logan, M.; Leelayouthayotin, C.; Visvanathan, C. Enhancement of food waste thermophilic anaerobic digestion through synergistic effect with chicken manure. Biomass Bioenergy 2020, 136, 105541. [Google Scholar] [CrossRef]
- Janke, L.; Leite, A.F.; Batista, K.; Silva, W.; Nikolausz, M.; Nelles, M.; Stinner, W. Enhancing biogas production from vinasse in sugarcane biorefineries: Effects of urea and trace elements supplementation on process performance and stability. Bioresour. Technol. 2016, 217, 10–20. [Google Scholar] [CrossRef]
- Gaur, R.Z.; Suthar, S. Anaerobic digestion of activated sludge, anaerobic granular sludge and cow dung with food waste for enhanced methane production. J. Clean. Prod. 2017, 164, 557–566. [Google Scholar] [CrossRef]
- Marin Batista, J.D.; Salazar, L.; Castro, L.; Escalante, H. Co-digestión anaerobia de vinaza y gallinaza de jaula: Alternativa para el manejo de residuos agrícolas colombianos. Rev. Colomb. Biotecnol. 2016, 18, 6–12. [Google Scholar] [CrossRef]
- Ripoll, V.; Agabo-García, C.; Perez, M.; Solera, R. Improvement of biomethane potential of sewage sludge anaerobic co-digestion by addition of “sherry-wine” distillery wastewater. J. Clean. Prod. 2020, 251, 119667. [Google Scholar] [CrossRef]
- Lefebvre, O.; Quentin, S.; Torrijos, M.; Godon, J.J.; Delgenes, J.P.; Moletta, R. Impact of increasing NaCl concentrations on the performance and community composition of two anaerobic reactors. Appl. Microbiol. Biotechnol. 2007, 75, 61–69. [Google Scholar] [CrossRef]
- Cabrera-Diaz, A.; Pereda-Reyes, I.; Oliva-Merencio, D.; Lebrero, R.; Zaiat, M. Anaerobic Digestion of Sugarcane Vinasse Through a Methanogenic UASB Reactor Followed by a Packed Bed Reactor. Appl. Biochem. Biotechnol. 2017, 183, 1127–1145. [Google Scholar] [CrossRef]
- De Vrieze, J.; De Lathouwer, L.; Verstraete, W.; Boon, N. High-rate iron-rich activated sludge as stabilizing agent for the anaerobic digestion of kitchen waste. Water Res. 2013, 47, 3732–3741. [Google Scholar] [CrossRef]
- Wang, B.; Stromberg, S.; Li, C.; Nges, I.A.; Nistor, M.; Deng, L.; Liu, J. Effects of substrate concentration on methane potential and degradation kinetics in batch anaerobic digestion. Bioresour. Technol. 2015, 194, 240–246. [Google Scholar] [CrossRef]
- Zhang, W.; Werner, J.J.; Agler, M.T.; Angenent, L.T. Substrate type drives variation in reactor microbiomes of anaerobic digesters. Bioresour. Technol. 2014, 151, 397–401. [Google Scholar] [CrossRef]
- Kiani Deh Kiani, M.; Parsaee, M.; Safieddin Ardebili, S.M.; Reyes, I.P.; Fuess, L.T.; Karimi, K. Different bioreactor configurations for biogas production from sugarcane vinasse: A comprehensive review. Biomass Bioenergy 2022, 161, 106446. [Google Scholar] [CrossRef]
- Maaz, M.; Yasin, M.; Aslam, M.; Kumar, G.; Atabani, A.E.; Idrees, M.; Anjum, F.; Jamil, F.; Ahmad, R.; Khan, A.L.; et al. Anaerobic membrane bioreactors for wastewater treatment: Novel configurations, fouling control and energy considerations. Bioresour. Technol. 2019, 283, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Harihastuti, N.; Yuliasni, R.; Djayanti, S.; Handayani, N.; Rame, R.; Prasetio, A.; Kadier, A. Full-Scale Application of Up-flow High Rate Anaerobic Reactor with Substrate Modification and Effluent Recirculation for Sugarcane Vinasse Degradation and Biogas Generation. J. Ecol. Eng. 2021, 22, 314–324. [Google Scholar] [CrossRef]
- Mostafa, P.; Mostafa, K.D.; Keikhosro, K. A review of biogas production from sugarcane vinasse. Biomass Bioenerg. 2019, 122, 117–125. [Google Scholar]
- Wu, B.; He, C.; Yuan, S.; Hu, Z.; Wang, W. Hydrogen enrichment as a bioaugmentation tool to alleviate ammonia inhibition on anaerobic digestion of phenol-containing wastewater. Bioresour. Technol. 2019, 276, 97–102. [Google Scholar] [CrossRef]
- Chen, H.; Wang, W.; Xue, L.; Chen, C.; Liu, G.; Zhang, R. Effects of Ammonia on Anaerobic Digestion of Food Waste: Process Performance and Microbial Community. Energy Fuels 2016, 30, 5749–5757. [Google Scholar] [CrossRef]
- De Vrieze, J.; Saunders, A.M.; He, Y.; Fang, J.; Nielsen, P.H.; Verstraete, W.; Boon, N. Ammonia and temperature determine potential clustering in the anaerobic digestion microbiome. Water Res. 2015, 75, 312–323. [Google Scholar] [CrossRef]
- Cheah, W.Y.; Ling, T.C.; Juan, J.C.; Lee, D.J.; Chang, J.S.; Show, P.L. Biorefineries of carbon dioxide: From carbon capture and storage (CCS) to bioenergies production. Bioresour. Technol. 2016, 215, 346–356. [Google Scholar] [CrossRef]
- Karadagli, F.; Rittmann, B.E. Kinetic characterization of Methanobacterium bryantii MoH. Environ. Sci. Technol. 2005, 39, 4900–4905. [Google Scholar] [CrossRef]
- Onodera, T.; Syutsubo, K.; Hatamoto, M.; Nakahara, N.; Yamaguchi, T. Evaluation of cation inhibition and adaptation based on microbial activity and community structure in anaerobic wastewater treatment under elevated saline concentration. Chem. Eng. J. 2017, 325, 442–448. [Google Scholar] [CrossRef]
- Gao, S.; Zhao, M.; Chen, Y.; Yu, M.; Ruan, W. Tolerance response to in situ ammonia stress in a pilot-scale anaerobic digestion reactor for alleviating ammonia inhibition. Bioresour. Technol. 2015, 198, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Espinoza-Escalante, F.M.; Pelayo-Ortíz, C.; Navarro-Corona, J.; González-García, Y.; Bories, A.; Gutiérrez-Pulido, H. Anaerobic digestion of the vinasses from the fermentation of Agave tequilana Weber to tequila: The effect of pH, temperature and hydraulic retention time on the production of hydrogen and methane. Biomass Bioenergy 2009, 33, 14–20. [Google Scholar] [CrossRef]
- Xiao, L.; Liu, F.; Xu, H.; Feng, D.; Liu, J.; Han, G. Biochar promotes methane production at high acetate concentrations in anaerobic soils. Environ. Chem. Lett. 2019, 17, 1347–1352. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Abbas, B.; Geleijnse, M.; Kolganova, T.V.; Kleerebezem, R.; van Loosdrecht, M.C. Syntrophic associations from hypersaline soda lakes converting organic acids and alcohols to methane at extremely haloalkaline conditions. Environ. Microbiol. 2016, 18, 3189–3202. [Google Scholar] [CrossRef]
- Sun, H.; Ni, P.; Angelidaki, I.; Dong, R.; Wu, S. Exploring stability indicators for efficient monitoring of anaerobic digestion of pig manure under perturbations. Waste Manag. 2019, 91, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cheng, J.J.; Creamer, K.S. Inhibition of anaerobic digestion process: A review. Bioresour. Technol. 2008, 99, 4044–4064. [Google Scholar] [CrossRef] [PubMed]
- Poirier, S.; Bize, A.; Bureau, C.; Bouchez, T.; Chapleur, O. Community shifts within anaerobic digestion microbiota facing phenol inhibition: Towards early warning microbial indicators? Water Res. 2016, 100, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.-F.; Xu, X.-H.; Dai, M.; Yuan, X.-Z.; Guo, R.-B. Hydrogen and methane production from vinasse using two-stage anaerobic digestion. Process Saf. Environ. Prot. 2017, 107, 81–86. [Google Scholar] [CrossRef]
- Lindner, J.; Zielonka, S.; Oechsner, H.; Lemmer, A. Is the continuous two-stage anaerobic digestion process well suited for all substrates? Bioresour. Technol. 2016, 200, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hu, R.; Hao, Y.; Yang, T.; Li, L.; Luo, Z.; Xie, L.; Zhao, N.; Liu, C.; Sun, C.; et al. CO2 and air pollutant emissions from bio-coal briquettes. Environ. Technol. Innov. 2023, 29, 102975. [Google Scholar] [CrossRef]
- Lloret, E.; Salar, M.J.; Blaya, J.; Pascual, J.A. Two-stage mesophilic anaerobic–thermophilic digestion for sludge sanitation to obtain advanced treated sludge. Chem. Eng. J. 2013, 230, 59–63. [Google Scholar] [CrossRef]
- Syaichurrozi, I.; Sumardiono, S. Predicting kinetic model of biogas production and biodegradability organic materials: Biogas production from vinasse at variation of COD/N ratio. Bioresour. Technol. 2013, 149, 390–397. [Google Scholar] [CrossRef]
- Paranhos, A.G.O.; Adarme, O.F.H.; Barreto, G.F.; Silva, S.Q.; Aquino, S.F. Methane production by co-digestion of poultry manure and lignocellulosic biomass: Kinetic and energy assessment. Bioresour. Technol. 2020, 300, 122588. [Google Scholar] [CrossRef]
- Czatzkowska, M.; Harnisz, M.; Korzeniewska, E.; Koniuszewska, I. Inhibitors of the methane fermentation process with particular emphasis on the microbiological aspect: A review. Energy. Sci. Eng. 2020, 8, 1880–1897. [Google Scholar] [CrossRef]
- Niu, Q.; Qiao, W.; Qiang, H.; Hojo, T.; Li, Y.Y. Mesophilic methane fermentation of chicken manure at a wide range of ammonia concentration: Stability, inhibition and recovery. Bioresour. Technol. 2013, 137, 358–367. [Google Scholar] [CrossRef]
- Niu, Q.; Qiao, W.; Qiang, H.; Li, Y.Y. Microbial community shifts and biogas conversion computation during steady, inhibited and recovered stages of thermophilic methane fermentation on chicken manure with a wide variation of ammonia. Bioresour. Technol. 2013, 146, 223–233. [Google Scholar] [CrossRef]
- Siles, J.A.; Garcia-Garcia, I.; Martin, A.; Martin, M.A. Integrated ozonation and biomethanization treatments of vinasse derived from ethanol manufacturing. J. Hazard. Mater. 2011, 188, 247–253. [Google Scholar] [CrossRef]
- Nualsri, C.; Reungsang, A.; Plangklang, P. Biochemical hydrogen and methane potential of sugarcane syrup using a two-stage anaerobic fermentation process. Ind. Crop Prod. 2016, 82, 88–99. [Google Scholar] [CrossRef]
- Rajagopal, R.; Massé, D.; Singh, G. A critical review on inhibition of anaerobic digestion process by excess ammonia. Bioresour. Technol. 2013, 143, 632–641. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, Z.; Zhang, Y. Using straw as a bio-ethanol source to promote anaerobic digestion of waste activated sludge. Bioresour. Technol. 2019, 286, 121388. [Google Scholar] [CrossRef]
- España-Gamboa, E.I.; Mijangos-Cortés, J.O.; Hernández-Zárate, G.; Maldonado, J.A.D.; Alzate-Gaviria, L.M. Methane production by treating vinasses from hydrous ethanol using a modified UASB reactor. Biotechnol. Biofuels 2012, 5, 82. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Cheng, H.; Chen, F.; Zhang, Y.; Xu, X.; Huang, C.; Chen, C.; Liu, W.; Ding, C.; Li, Z.; et al. Enhanced methane production by alleviating sulfide inhibition with a microbial electrolysis coupled anaerobic digestion reactor. Environ. Int. 2020, 136, 105503. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhen, G.; Ni, J.; Hojo, T.; Kubota, K.; Li, Y.Y. Effect of influent COD/SO42− ratios on biodegradation behaviors of starch wastewater in an upflow anaerobic sludge blanket (UASB) reactor. Bioresour. Technol. 2016, 214, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Hao, Y.; Yang, T.; Wu, J.; Xu, F.; Li, L.; Wang, B.; Li, M.; Zhao, N.; Wang, N.; et al. Air pollutant emissions from sludge-bituminous briquettes as a potential household energy source. Case Stud. Therm. Eng. 2022, 37, 102251. [Google Scholar] [CrossRef]
- Hu, Y.; Jing, Z.; Sudo, Y.; Niu, Q.; Du, J.; Wu, J.; Li, Y.Y. Effect of influent COD/SO42− ratios on UASB treatment of a synthetic sulfate-containing wastewater. Chemosphere. 2015, 130, 24–33. [Google Scholar] [CrossRef]
- Eskicioglu, C.; Kennedy, K.J.; Marin, J.; Strehler, B. Anaerobic digestion of whole stillage from dry-grind corn ethanol plant under mesophilic and thermophilic conditions. Bioresour. Technol. 2011, 102, 1079–1086. [Google Scholar] [CrossRef]
- Degueurce, A.; Tomas, N.; Le Roux, S.; Martinez, J.; Peu, P. Biotic and abiotic roles of leachate recirculation in batch mode solid-state anaerobic digestion of cattle manure. Bioresour. Technol. 2016, 200, 388–395. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Wu, J. Enhancement of methane production in anaerobic digestion process: A review. Appl. Energy 2019, 240, 120–137. [Google Scholar] [CrossRef]
- Ahmed, P.M.; Nieto-Penalver, C.G.; de Figueroa, L.I.C.; Pajot, H.F. Vinasse odyssey: Sugarcane vinasse remediation and laccase production by Trametes sp. immobilized in polyurethane foam. Biodegradation 2022, 33, 333–348. [Google Scholar] [CrossRef]
- Raskin, L.; Stromley, J.M.; Rittmann, B.E.; Stahl, D.A. Group-Specific 16S rRNA Hybridization Probes To Describe Natural Communities of Methanogens. Appl. Environ. 1994, 60, 1232–1240. [Google Scholar] [CrossRef]
- Galagan, J.E.; Nusbaum, C.; Roy, A.; Endrizzi, M.G.; Macdonald, P.; FitzHugh, W.; Calvo, S.; Engels, R.; Smirnov, S.; Atnoor, D.; et al. The genome of M. acetivorans reveals extensive metabolic and physiological diversity. Genome Res. 2002, 12, 532–542. [Google Scholar] [CrossRef]
- Zahedi, S.; Solera, R.; Micolucci, F.; Cavinato, C.; Bolzonella, D. Changes in microbial community during hydrogen and methane production in two-stage thermophilic anaerobic co-digestion process from biowaste. Waste Manag. 2016, 49, 40–46. [Google Scholar] [CrossRef]
- Li, G.; Yang, T.; Xiao, W.; Wu, J.; Xu, F.; Li, L.; Gao, F.; Huang, Z. Sustainable environmental assessment of waste-to-energy practices: Co-pyrolysis of food waste and discarded meal boxes. Foods 2022, 11, 3840. [Google Scholar] [CrossRef]
- Zahedi, S.; Sales, D.; Romero, L.I.; Solera, R. Hydrogen production from the organic fraction of municipal solid waste in anaerobic thermophilic acidogenesis: Influence of organic loading rate and microbial content of the solid waste. Bioresour. Technol. 2013, 129, 85–91. [Google Scholar] [CrossRef]
- Ziganshin, A.M.; Schmidt, T.; Scholwin, F.; Il’inskaya, O.N.; Harms, H.; Kleinsteuber, S. Bacteria and archaea involved in anaerobic digestion of distillers grains with solubles. Appl. Microbiol. Biotechnol. 2011, 89, 2039–2052. [Google Scholar] [CrossRef]
- Li, J.; Zheng, G.; He, J.; Chang, S.; Qin, Z. Hydrogen-producing capability of anaerobic activated sludge in three types of fermentations in a continuous stirred-tank reactor. Biotechnol. Adv. 2009, 27, 573–577. [Google Scholar] [CrossRef]
- Ziganshin, A.M.; Liebetrau, J.; Proter, J.; Kleinsteuber, S. Microbial community structure and dynamics during anaerobic digestion of various agricultural waste materials. Appl. Microbiol. Biotechnol. 2013, 97, 5161–5174. [Google Scholar] [CrossRef]
- Sarti, A.; Pozzi, E.; Chinalia, F.A.; Ono, A.; Foresti, E. Microbial processes and bacterial populations associated to anaerobic treatment of sulfate-rich wastewater. Process. Biochem. 2010, 45, 164–170. [Google Scholar] [CrossRef]
- Yabuki, L.; Queluz, J.; Garcia, M. Assessment of phase distribution and removal of metals in anaerobic digesters. Int. J. Environ. Sci. Technol. 2022, 19, 463–474. [Google Scholar] [CrossRef]
- Wu, J.; Niu, Q.; Li, L.; Hu, Y.; Mribet, C.; Hojo, T.; Li, Y.Y. A gradual change between methanogenesis and sulfidogenesis during a long-term UASB treatment of sulfate-rich chemical wastewater. Sci. Total Environ. 2018, 636, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Mosbaek, F.; Kjeldal, H.; Mulat, D.G.; Albertsen, M.; Ward, A.J.; Feilberg, A.; Nielsen, J.L. Identification of syntrophic acetate-oxidizing bacteria in anaerobic digesters by combined protein-based stable isotope probing and metagenomics. ISME J. 2016, 10, 2405–2418. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhang, J.; Pan, M.; Hao, Y.; Hu, R.; Xiao, W.; Li, G.; Lv, T. Valorisation of microalgae residues after lipid extraction: Pyrolysis characteristics for biofuel production. Biochem. Eng. J. 2022, 179, 108330. [Google Scholar] [CrossRef]
- Fu, J.; Yan, B.; Gui, S.; Fu, Y.; Xia, S. Anaerobic co-digestion of thermo-alkaline pretreated microalgae and sewage sludge: Methane potential and microbial community. J. Environ. Sci. 2023, 127, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Parnaudeau, V.; Condom, N.; Oliver, R.; Cazevieille, P.; Recous, S. Vinasse organic matter quality and mineralization potential, as influenced by raw material, fermentation and concentration processes. Bioresour. Technol. 2008, 99, 1553–1562. [Google Scholar] [CrossRef]
- Wilk, M.; Krzywonos, M.; Seruga, P.; Walaszczyk, E. Effect of pH and temperature on vinasse decolorization by lactic acid bacteria in batch processes. Water Environ. Res. 2019, 91, 573–580. [Google Scholar] [CrossRef]
- Chandra, R.; Bharagava, R.N.; Rai, V. Melanoidins as major colourant in sugarcane molasses based distillery effluent and its degradation. Bioresour. Technol. 2008, 99, 4648–4660. [Google Scholar] [CrossRef]
- Forouzanmehr, F.; Solon, K.; Maisonnave, V.; Daniel, O.; Volcke, E.I.P.; Gillot, S.; Buffiere, P. Sulfur transformations during two-stage anaerobic digestion and intermediate thermal hydrolysis. Sci. Total Environ. 2022, 810, 151247. [Google Scholar] [CrossRef]
- Welte, C.; Deppenmeier, U. Bioenergetics and anaerobic respiratory chains of aceticlastic methanogens. Biochim. Biophys. Acta 2014, 1837, 1130–1147. [Google Scholar] [CrossRef]
- Krzywonos, M.; Chałupniak, A.; Zabochnicka-Świątek, M. Decolorization of beet molasses vinasse by Bacillus megaterium ATCC 14581. Bioremediation J. 2017, 21, 81–88. [Google Scholar] [CrossRef]
- Tiwari, S.; Rai, P.; Yadav, S.K.; Gaur, R. A novel thermotolerant Pediococcus acidilactici B-25 strain for color, COD, and BOD reduction of distillery effluent for end use applications. Environ. Sci. Pollut. Res. Int. 2013, 20, 4046–4058. [Google Scholar] [CrossRef]
- Krayzelova, L.; Bartacek, J.; Kolesarova, N.; Jenicek, P. Microaeration for hydrogen sulfide removal in UASB reactor. Bioresour. Technol. 2014, 172, 297–302. [Google Scholar] [CrossRef]
- Shakeri Yekta, S.; Elreedy, A.; Liu, T.; Hedenstrom, M.; Isaksson, S.; Fujii, M.; Schnurer, A. Influence of cysteine, serine, sulfate, and sulfide on anaerobic conversion of unsaturated long-chain fatty acid, oleate, to methane. Sci. Total Environ. 2022, 817, 152967. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhang, Y.; Li, Q.; Quan, X. Adding Fe0 powder to enhance the anaerobic conversion of propionate to acetate. Biochem. Eng. J. 2013, 73, 80–85. [Google Scholar] [CrossRef]
- Khan, M.A.; Ngo, H.H.; Guo, W.S.; Liu, Y.W.; Zhou, J.L.; Zhang, J.; Liang, S.; Ni, B.J.; Zhang, X.B.; Wang, J. Comparing the value of bioproducts from different stages of anaerobic membrane bioreactors. Bioresour. Technol. 2016, 214, 816–825. [Google Scholar] [CrossRef]
- De Sitter, K.; Garcia-Gonzalez, L.; Matassa, C.; Bertin, L.; De Wever, H. The use of membrane based reactive extraction for the recovery of carboxylic acids from thin stillage. Sep. Purif. Technol. 2018, 206, 177–185. [Google Scholar] [CrossRef]
- De Vrieze, J.; Gildemyn, S.; Arends, J.B.; Vanwonterghem, I.; Verbeken, K.; Boon, N.; Verstraete, W.; Tyson, G.W.; Hennebel, T.; Rabaey, K. Biomass retention on electrodes rather than electrical current enhances stability in anaerobic digestion. Water Res. 2014, 54, 211–221. [Google Scholar] [CrossRef]
- Sun, C.; Cao, W.; Banks, C.J.; Heaven, S.; Liu, R. Biogas production from undiluted chicken manure and maize silage: A study of ammonia inhibition in high solids anaerobic digestion. Bioresour. Technol. 2016, 218, 1215–1223. [Google Scholar] [CrossRef]
- Li, G.; Hu, R.; Wang, N.; Yang, T.; Xu, F.; Li, J.; Wu, J.; Huang, Z.; Pan, M.; Lv, T. Cultivation of microalgae in adjusted wastewater to enhance biofuel production and reduce environmental impact: Pyrolysis performances and life cycle assessment. J. Clean. Prod. 2022, 355, 131768. [Google Scholar] [CrossRef]
- Mao, C.; Feng, Y.; Wang, X.; Ren, G. Review on research achievements of biogas from anaerobic digestion. Renew. Sustain. Energy Rev. 2015, 45, 540–555. [Google Scholar] [CrossRef]
- Zhang, J.; Yip, C.; Xia, C.; Liang, Y. Evaluation of methane release from coals from the San Juan basin and Powder River basin. Fuel 2019, 244, 388–394. [Google Scholar] [CrossRef]
- Nallathambi, G. Anaerobic digestion of biomass for methane production: A review. Biomass Bioenergy 1997, 13, 83–114. [Google Scholar] [CrossRef]
- Yin, M.; Chen, H. Unveiling the dual faces of chitosan in anaerobic digestion of waste activated sludge. Bioresour. Technol. 2022, 344, 126182. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).