3.3. Selection of Solid-State Mode for Raw Fermentation
The sensory evaluation results for the three fermentation modes are shown in
Table 6. It can be seen in the table that the four sensory evaluation indexes all showed significant differences (
p < 0.05), indicating that the different types of fermentation modes had strong influences on the fermentation results. Among them, the differences in the product odor, pH value, and appearance were the most obvious. The pH value of model C was the lowest and the smell was the most acidic; the appearance of the fermented product was bright, uniform, and without mold pollution. To understand the situation of bacterium infection in the fermentation more clearly, after the observation of the appearance, the sample was viewed under an optical microscope to determine whether there were non-mycotic bacteria in the fermentation products. It was found that, in both mode B and mode C, there were bacteria other than the three kinds of artificial inoculating strains, which might have been other bacteria growing in the fermentation system. The main problem facing uncooked raw fermentation is how to prevent the endogenous contamination of the fermentation system, especially by
E. coli,
Salmonella, and other pathogenic bacteria.
Pediococcus acidilactici can produce bacteriocin to inhibit
Listeria monocytogenes,
Shigella, and
Clostridium perfringens [
23,
46,
47]. The results showed that the lactic acid, acetic acid, and total acid contents in the fermentation system of
Pediococcus acidilactici were positively correlated with the antibacterial activity [
24]. Zhang Yiran et al. reported that
Bacillus subtilis MA139 could secrete antimicrobial peptides that kill
Escherichia coli,
Salmonella, and
Staphylococcus aureus, but they have no effect on the growth and metabolism of yeast and lactic acid bacteria [
48]. In this experiment, the bacteriostatic characteristics of
Pediococcus acidilactici and
Bacillus subtilis were used to antagonize pathogenic bacteria in the fermentation system. As
E. coli,
Salmonella, and other intestinal pathogenic bacteria cannot tolerate pH levels below 4.5, to effectively inhibit pathogenic bacteria in the fermented feed, rapid initial lactic acid production was required [
49,
50,
51]. Since
Pediococcus acidilactici can simultaneously produce bacteriostatic substances and lower pH, it was more conducive to inhibiting the growth of pathogenic bacteria such as
E. coli and
Salmonella. Therefore, model C was determined to be the best model for complex probiotic fermentation; that is,
Pediococcus acidilactici was inoculated first for closed fermentation and then
Saccharomyces cerevisiae and
Bacillus subtilis were inoculated simultaneously for static fermentation.
3.4. Pediococcus Acidilactici Fermentation Test
Although it has been reported in the literature that pediocin can be produced by
Pediococcus acidilactici [
52,
53], it is more commonly used for food preservative or fermented silage [
54,
55,
56] and rarely for solid-state fermentation of raw corn–soybean meal. As a facultative anaerobic fermentation bacterium,
Pediococcus acidilactici is a micro-aerobic microorganism with a strong bacteriocin production ability under micro-oxygen conditions [
52], and its SSF should be carried out in a closed environment. Therefore, in this experiment, the bag was filled with the raw materials and then closed static SSF was adopted. Low moisture content in a solid-state fermentation medium results in high substrate porosity, which is conducive to air penetration, leading to relatively abundant oxygen in the substrate. High moisture content in a solid-state fermentation medium results in low substrate porosity, which, in turn, prevents air penetration and an anaerobic environment is created. At the same time, the water content in the substrate, the water activity, and the mass transfer rate between microbial cells increase, which is conducive to the generation of microbial metabolites [
3,
10]. As shown in
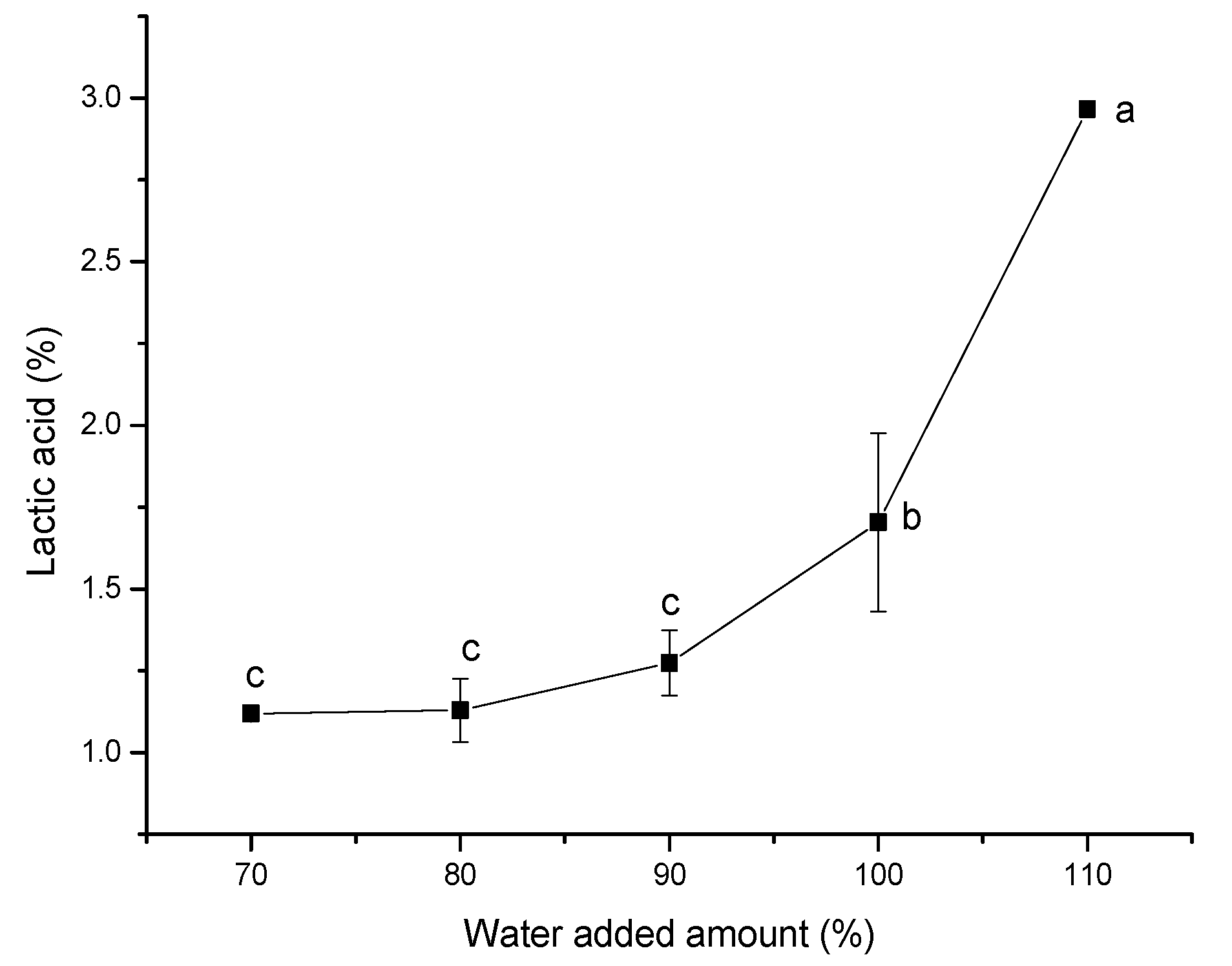
Figure 2, in the single-factor experiments for the water addition amount, the lactic acid content in the system gradually increased with the increase in the amount of water added in the solid medium, and the increase in lactic acid content was not significant when the water addition amount was less than 90%. When the water addition amount exceeded 90%, the lactic acid content in the system increased significantly. When the water addition amount reached 110%, although the lactic acid content was relatively the highest, the actual state of the medium was semi-solid, which was not conducive to the subsequent fermentation. Taking into account the actual situation, it was judged appropriate to choose the 100% water addition amount for the experiment.
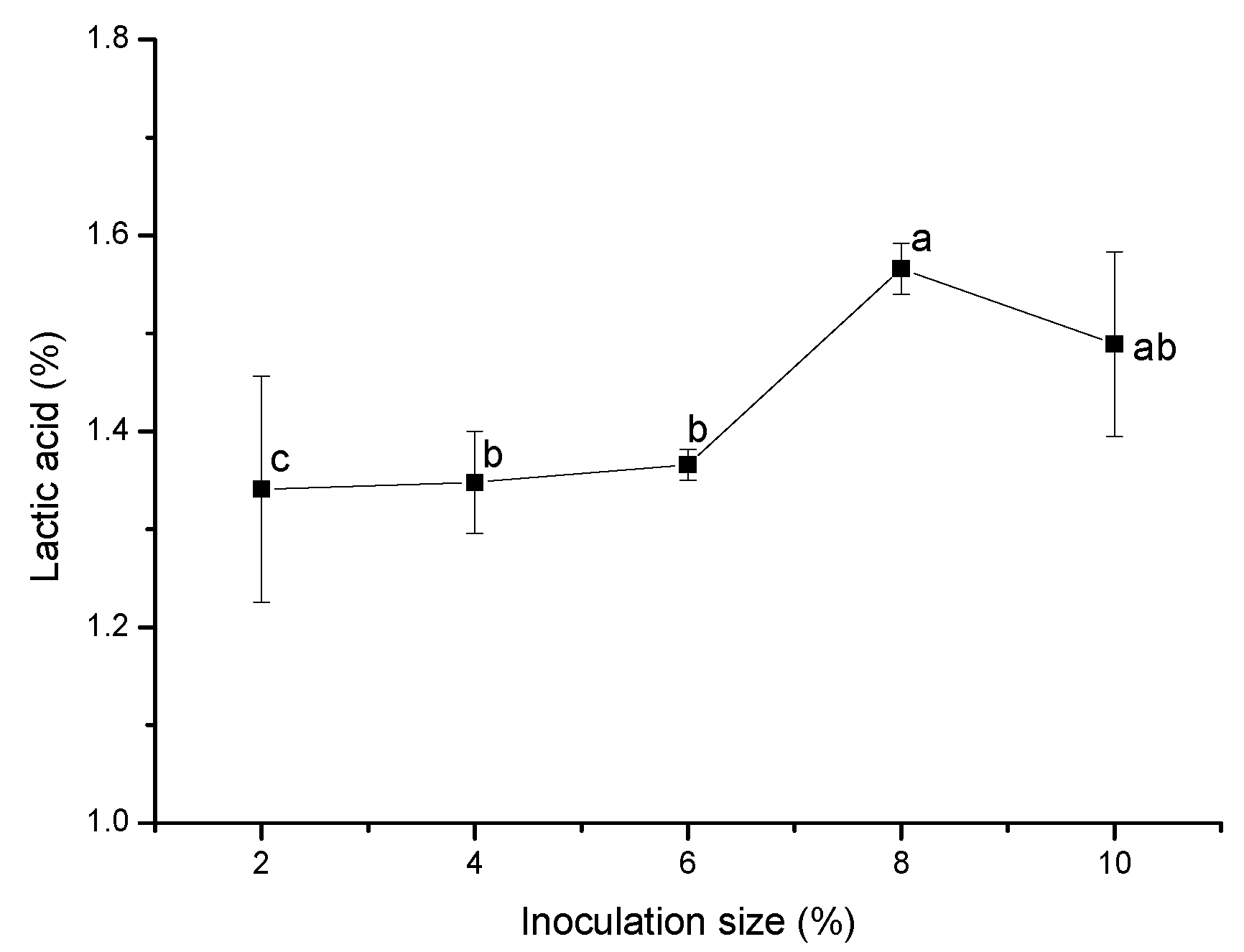
Excessive inoculation size leads to excessive growth of bacteria in the early stage of fermentation and consumes a lot of nutrients, which is not conducive to the accumulation of metabolic substances. If the inoculation size is too small, the microbial growth in the system will be slow, and the fermentation cycle will be prolonged [
57]. As shown in
Figure 3, with the increase in inoculation size, lactic acid production first increased and then decreased. This was because, with the increase in inoculation size in the fermentation system, the initial numbers of lactic acid bacteria cells and the cell proliferation of
Pediococcus acidilactici increased and, thus, the production of lactic acid and metabolites by the bacteria also increased. However, when the inoculation size exceeded 8%, the metabolic acid production in the system no longer increased due to the proliferation of cells and the consumption of nutrients in the system. The lactic acid yield reached its maximum when the inoculation size was 8%, so 8% was the optimal inoculation size.
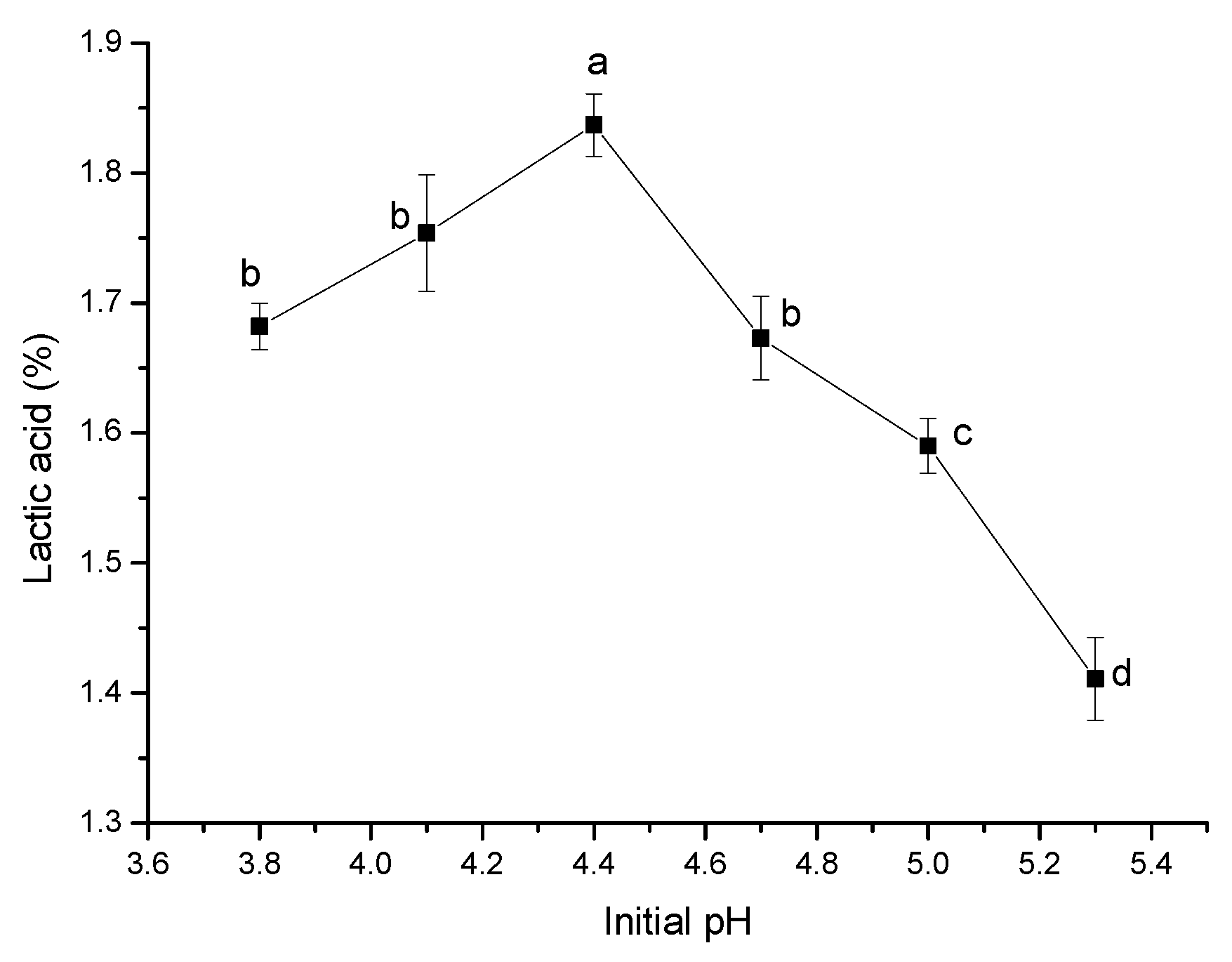
pH affects the function of membrane proteins by affecting the charge distribution on the cells, changes the absorption of nutrients, and ultimately affects the growth of the cell itself and the accumulation of metabolites [
57]. As shown in
Figure 4, with the increase in initial pH, lactic acid production in the fermentation system increased first and then decreased. When the initial pH was 4.4, lactic acid production was relatively the highest. As intestinal pathogenic bacteria, such as
Escherichia coli and
Salmonella, cannot tolerate pH below 4.5 [
51], the growth of intestinal pathogenic bacteria would be inhibited in systems with pH values lower than 4.4, and such systems would be more conducive to the growth of
Pediococcus acidilactici. When the pH value of the system was above 4.6, the growth of various bacteria, including intestinal pathogenic bacteria, affected the proliferation of
Pediococcus acidilactici cells, leading to a greater decline in lactic acid production. However, too low an initial pH value could also affect the membrane protein function, the intake of nutrients, the cell proliferation, and the acid production of
Pediococcus acidilactici. Therefore, the optimal initial pH was 4.4.
The orthogonal test of the fermentation conditions of
Pediococcus acidilactici is described in
Table 7. The factors influencing lactic acid yield in the fermentation system were as follows: initial pH value > water addition amount > empty column > inoculation amount. The extremum difference analysis showed that the optimal process conditions were A
2B
3D
1; that is, initial pH 4.4, 100% water addition amount, and 6% inoculation size.
Table 8 shows the variance analysis of the orthogonal experiment. The table shows that the major factors affecting lactic acid yield were as follows: initial pH value > water addition amount > empty column > inoculation size. The initial pH value and water addition amount had significant effects on lactic acid production, while the empty column and inoculation size had no significant effects on lactic acid production. Furthermore, MS
inoculation size< 2 MS
empty column. Therefore, the sum of the squares of deviation and freedom for the inoculation size were merged with the sum of the squares of deviation and freedom for the blank column, respectively, and then transformed into the sum of the squares of deviation and freedom for the error term, respectively, to improve the sensitivity of the
F test. Considering the economic cost and other factors, the optimal process conditions were pH 4.4, 100% water addition amount, and 6% inoculation size.
3.5. Optimization of Solid-State Fermentation Conditions in the Second Stage
With different strains, inoculation proportions, and culture temperatures, the bacteria body protein content of the fermented feed will differ, which will affect the feeding effect in animals [
22]. Therefore, it is necessary to optimize the fermentation feed preparation process to obtain higher bacteria body protein content at lower cost and thus achieve higher cost benefits. The purpose of the second stage of the SSF was to use
Bacillus subtilis to degrade macromolecular substances in raw material, which is conducive to the proliferation of
Saccharomyces cerevisiae, and thus increase the protein content in fermented feed. Therefore, the inoculation ratio of
Bacillus subtilis to
Saccharomyces cerevisiae, inoculation size, and SSF temperature were the main factors affecting the protein content. Firstly, the single-factor experiments for the inoculation ratio, inoculation size, and SSF temperature were respectively conducted, and the results are shown in
Figure 5,
Figure 6 and
Figure 7. In previous research,
Bacillus subtilis could grow on raw materials consisting of grains and beans and secrete protease, cellulase, amylase, lipase, and other hydrolytic enzymes, especially those that show strong neutral protease and alkaline protease activity. Furthermore, the raw organic matter was decomposed, and the digestion and absorption rate of animals were improved [
18,
42,
58,
59]. It has been shown that inoculated probiotics, such as
Saccharomyces cerevisiae,
Bacillus subtilis, and lactic acid bacteria, can convert non-protein nitrogen in soybean meal into bacteria body protein with higher nutritional value during the cell proliferation process, improve the nutritional structure of raw materials, and increase the protein content of raw materials [
59,
60,
61,
62]. As shown in
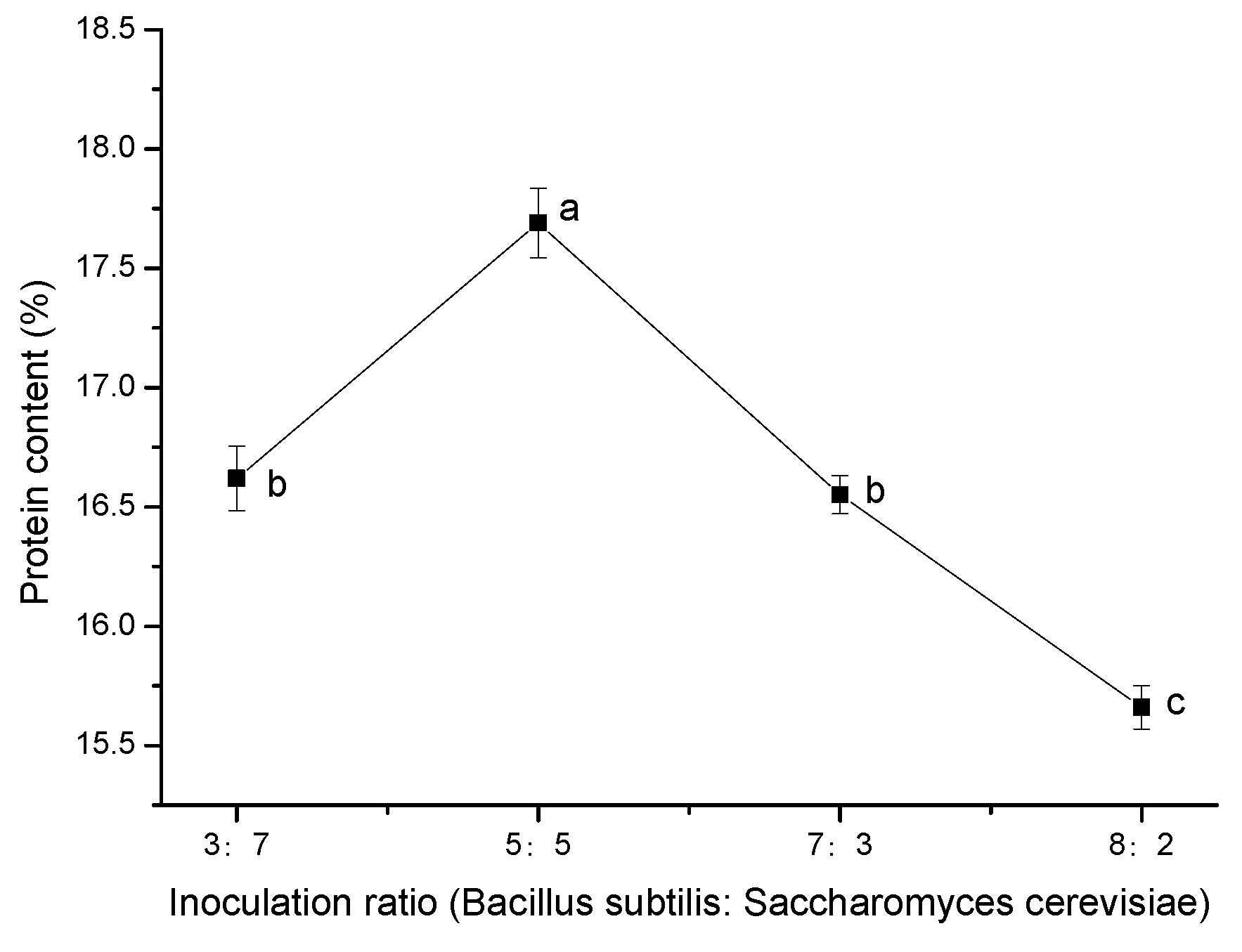
Figure 6, protein content in the fermentation system initially increased with the increase in the
Bacillus subtilis inoculation ratio. When the ratio of the two strains was 5:5, the protein content was the highest. When the proportion of
Bacillus subtilis inoculated exceeded that of
Saccharomyces cerevisiae, the protein content in the system began to decrease. With certain nutrients in the system, the proportion of
Bacillus subtilis increased appropriately, which was conducive to its metabolism and production of enzymes and decomposition of starch, protein, and other macromolecular substances in the fermentation system; such changes are useful for yeast proliferation and to increase yeast protein. However, when the proportion of
Bacillus subtilis was too high, the cell proliferation led to consumption of excessive nutrients, which is not conducive to the production of enzymes and the degradation of macromolecular substances, so the protein content in the fermentation system began to decline again. Therefore, the optimal inoculation ratio for the single-factor experiment was
Bacillus subtilis:
Saccharomyces cerevisiae = 5:5.
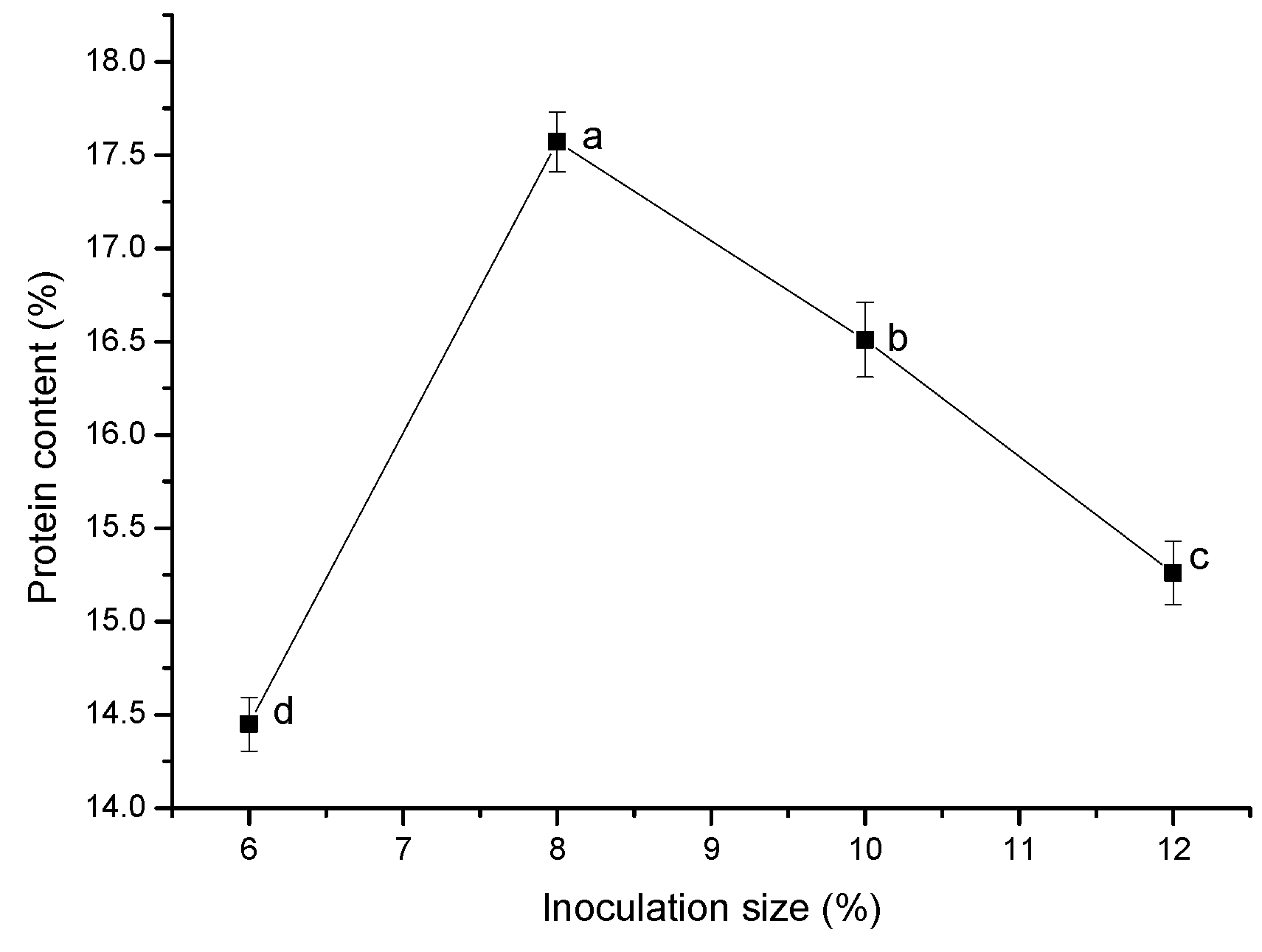
The inoculation size affected the proliferation rate and metabolite yield of bacteria [
57]. As shown in
Figure 6, protein content increased as the inoculation size increased from 6% to 8%. When the inoculation size exceeded 8%, the protein content tended to decline due to excessive proliferation of cells and excessive consumption of nutrients in the system. It can be seen that a larger inoculation size is not always better for fermented feed, which is consistent with the results published by Guo Ruyi et al. [
57]. Therefore, the optimal inoculation size for the single-factor experiment was 8%.
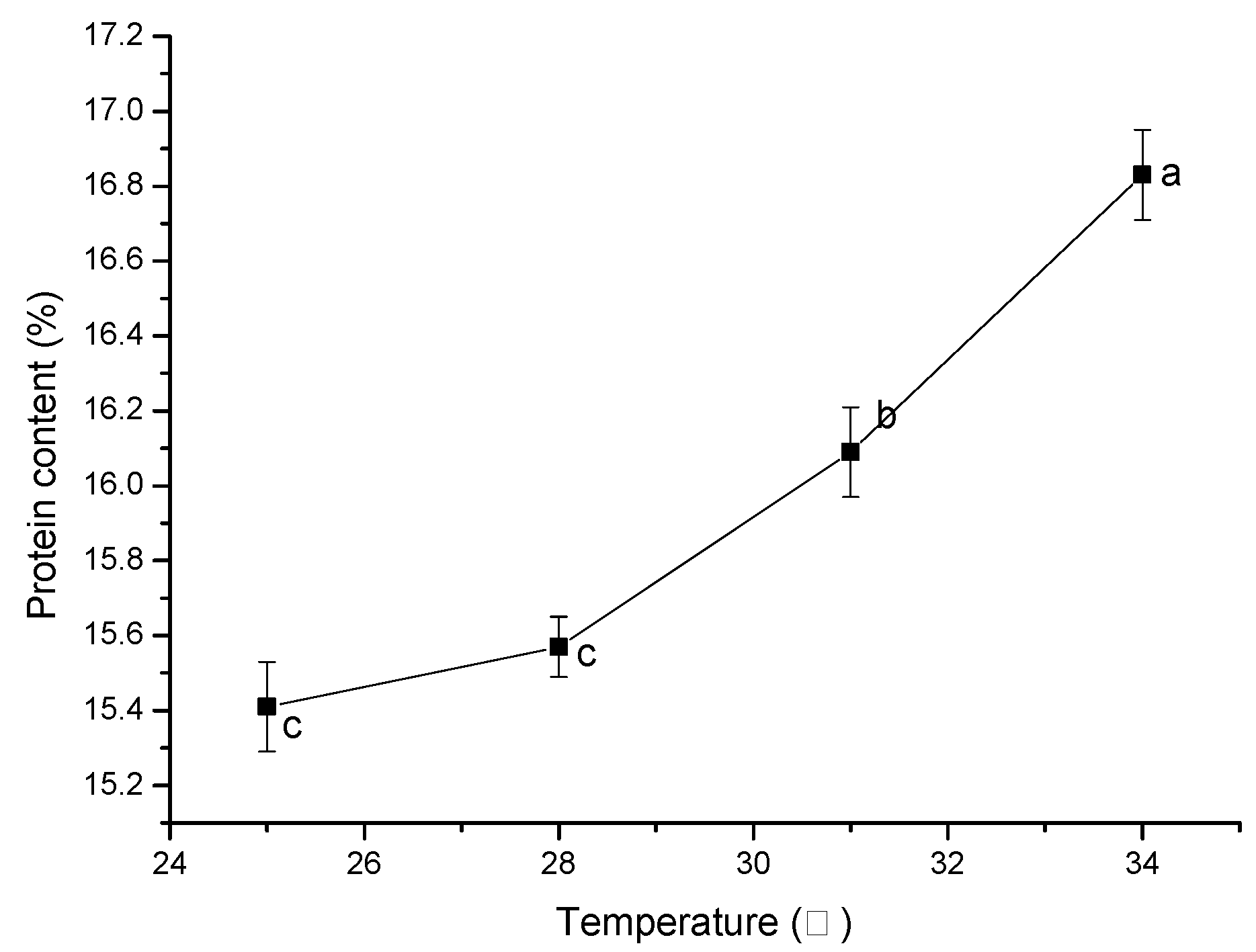
The optimum growth temperatures for the different microorganisms differ. The optimum growth temperature for
Bacillus subtilis is 37 °C, and that of
Saccharomyces cerevisiae is between 28 and 31 °C. High temperature is not conducive to the growth of
Saccharomyces cerevisiae, but
Bacillus subtilis can grow well at 40 °C since it has strong heat resistance [
58,
59]. Considering the temperature growth characteristics of
Bacillus subtilis and
Saccharomyces cerevisiae, the temperature range of 25 °C–34 °C was selected in the study. As shown in
Figure 7, the protein content in the system increased with the increase in culture temperature. This was because, with the increase in temperature, the exudation of soluble substances and the efficiency of enzymatic hydrolysis in the system increased, so
Bacillus subtilis and Saccharomyces cerevisiae could obtain more available substances and the protein content of the bacteria and yeast in the system increased. In the range of temperatures used, the optimum culture temperature was 34 °C. Based on the single-factor experiments, a response surface methodology (RSM) was used to further optimize the SSF conditions in the second stage. The experiment results are shown in
Table 9 and the variance analysis results are shown in
Table 10. The
p value of 0.0082 of the regression model in
Table 10 was less than 0.01, indicating that the regression relationship between the independent variable and the response value was extremely significant (
p < 0.01). The
p value for the lack of fit was 0.0689, indicating that the difference between the lack of fit and the pure error was not significant. According to the
F values in
Table 10, the contribution rates of the independent variable factors were as follows: A > B > C; that is to say, inoculation ratio > fermentation temperature > inoculation size. The inoculation ratio had the most significant effect on the protein content of the fermented feed, followed by the fermentation temperature, and the inoculation size had the least significant relative effect. In the model, the first-order term of the independent variable and the interaction between the independent variables had no significant influence on the response value. In the model, the quadratic term of the independent variable fermentation temperature had a very significant influence on the response value (
p < 0.01), and the quadratic term of the independent variable inoculation size had a significant influence on the response value (
p < 0.05).
Design-Expert was used to analyze the variance in the experimental data, and the quadratic regression model was established:
The correlation coefficient of the regression model R2 = 0.9023, indicating that 90.23% of the response value changes were related to the inoculation ratio, fermentation temperature, and inoculation amount, and the prediction was good.
The optimal process parameters for the model obtained by software optimization are shown in
Table 11: at an inoculation ratio of
Bacillus subtilis to
Saccharomyces cerevisiae (
v/
v) of 3.8:6.2, fermentation temperature of 33.52 °C, and inoculation size of 6%, the protein content of the fermented feed was the highest, and the predicted value was 19.66%. As the fermentation temperature of 33.52 °C was not easy to operate, the fermentation temperature was adjusted to 33.5 °C. The verification tests were carried out with the optimized process parameters and the protein content of the fermented feed was measured as 19.87%, which was basically consistent with the predicted value. Therefore, this proved that the model was reliable. Compared with unfermented raw material, the protein content of the fermented feed was increased by 5.21 percentage points and 35.74% (
p < 0.05). Hou Nannan et al. used soybean meal, corn meal, and bran as raw materials; solid-state fermentation was conducted with lactic acid bacteria and yeast added for 72 h, and the crude protein content of the fermented feed was 6.07% higher than that of the control [
61]. Shi Changyou et al. fermented a mixture of soybean meal, corn meal, and bran with
Bacillus subtilis and
Enterococcus faecium in two-stage solid-state fermentation. In the first stage,
Bacillus subtilis underwent aerobic fermentation for 24 h, and then anaerobic fermentation was conducted with
Enterococcus faecalis added again for 48 h. Compared with the control, the crude protein in the fermented feed increased by 3.58 percentage points (
p < 0.05) [
29]. In this study, the feed fermented by the compound probiotics had an obvious fruity taste, sweetness, and a bright appearance, and the sensory properties of the fermented feed were consistent with descriptions of high-quality fermented feed [
61].
3.6. The Results of High-Throughput Sequencing for the Detection of the Relative Abundance of Bacterial Microflora in the Fermentation System
The fermentation samples were collected at 0 h (before inoculation), 48 h (after the second stage of inoculation), and 96 h and sent to Novogene Bioinformation Technology Co., Ltd., respectively, for third-generation high-throughput sequencing using the PacBio platform. The results of the 16S rDNA bacterial microflora abundance detection are shown in
Figure 8 and
Table S2, and the results of the ITS fungal microflora abundance detection are shown in
Figure 9 and
Table S3.
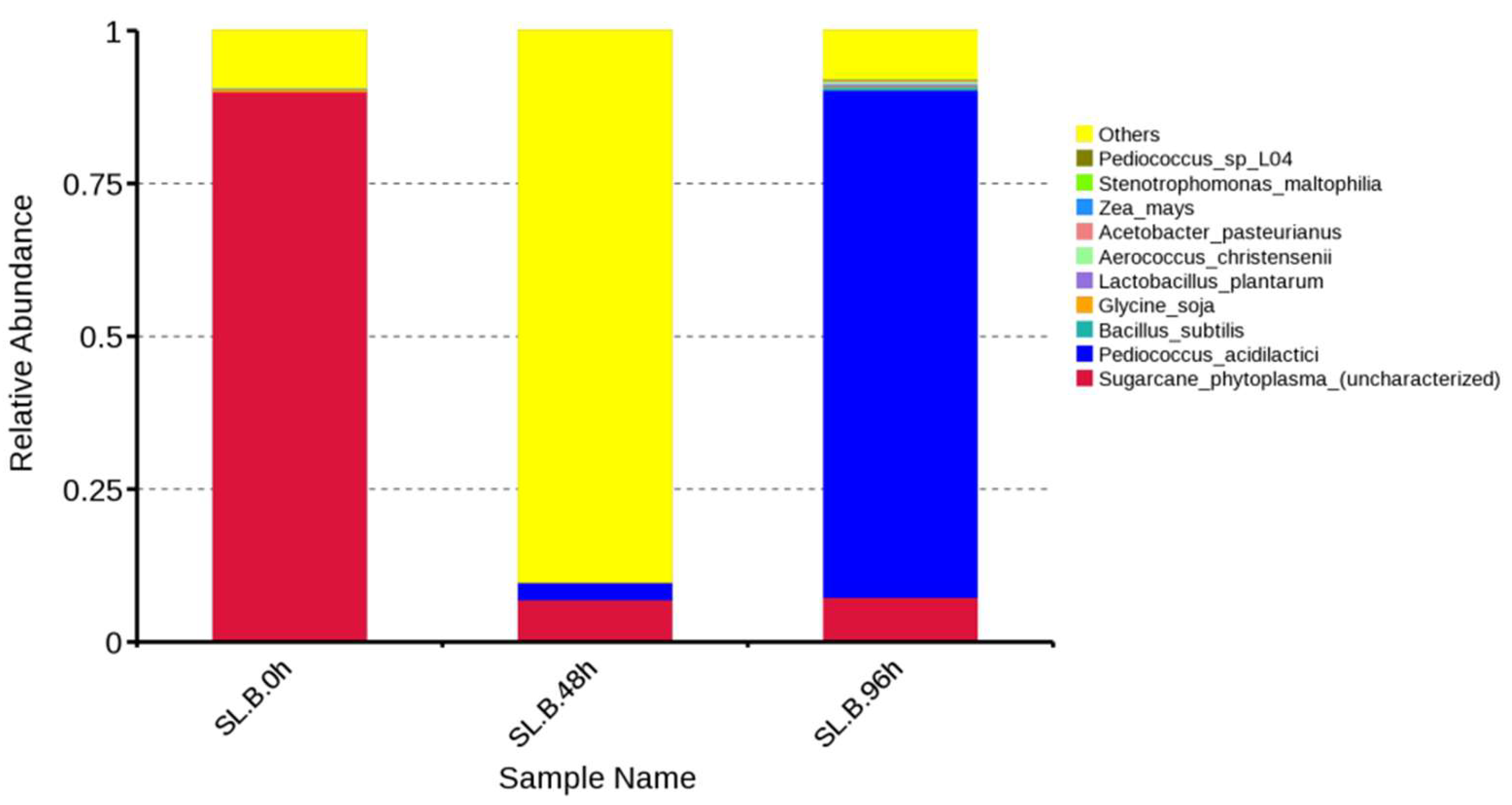
As shown in
Figure 8, the original bacterial communities in the uninoculated raw feed system were, in order,
Sugarcane phytoplasma (uncharacterized), “Others”,
Glycine soja, and
Zea mays, but the relative abundances of the latter two were low. After being fermented with
Pediococcus acidilactici for 48 h, the bacterial communities in the fermented system with relative abundances that increased were “Others” and
Pediococcus acidilactici. The relative abundances of the communities
Sugarcane_phytoplasma (uncharacterized) and
Glycine soja decreased, while
Zea mays disappeared. It could be seen that the initial distribution of the bacterial community in the system was changed by the fermentation of
Pediococcus acidilactici in corn–soybean meal for 48 h, and the relative abundance of
Sugarcane phytoplasma (uncharacterized) was reduced and that of “Others” increased. After the cooperative fermentation of
Bacillus subtilis and
Saccharomyces cerevisiae for 96 h,
Pediococcus acidilactici was the dominant bacterial community with the highest relative abundance. Among the top ten, the communities with relative abundances that increased were, in turn,
Bacillus subtilis,
Lactobacillus plantarum,
Aerococcus christensenii,
Acetobacter pasteurianus,
Stenotrophomonas maltophilia, and
Pediococcus sp_L04 (from high to low). The community with relative abundances that decreased was “Others”, while the relative abundances of
Sugarcane phytoplasma (uncharacterized) and
Glycine soja remained basically stable. Lian Mengran compared the community structure of soybean meal with that of fermented soybean meal and found that the original community in soybean meal, unidentified
Cyanobacteria, decreased significantly after fermentation. The relative abundances of
Lactobacillus,
Pediococcus,
Enterococcus, and
Staphylococcus and the diversity of bacterial communities increased after fermentation, which was similar to the change in the trend for bacterial communities in this study [
63].
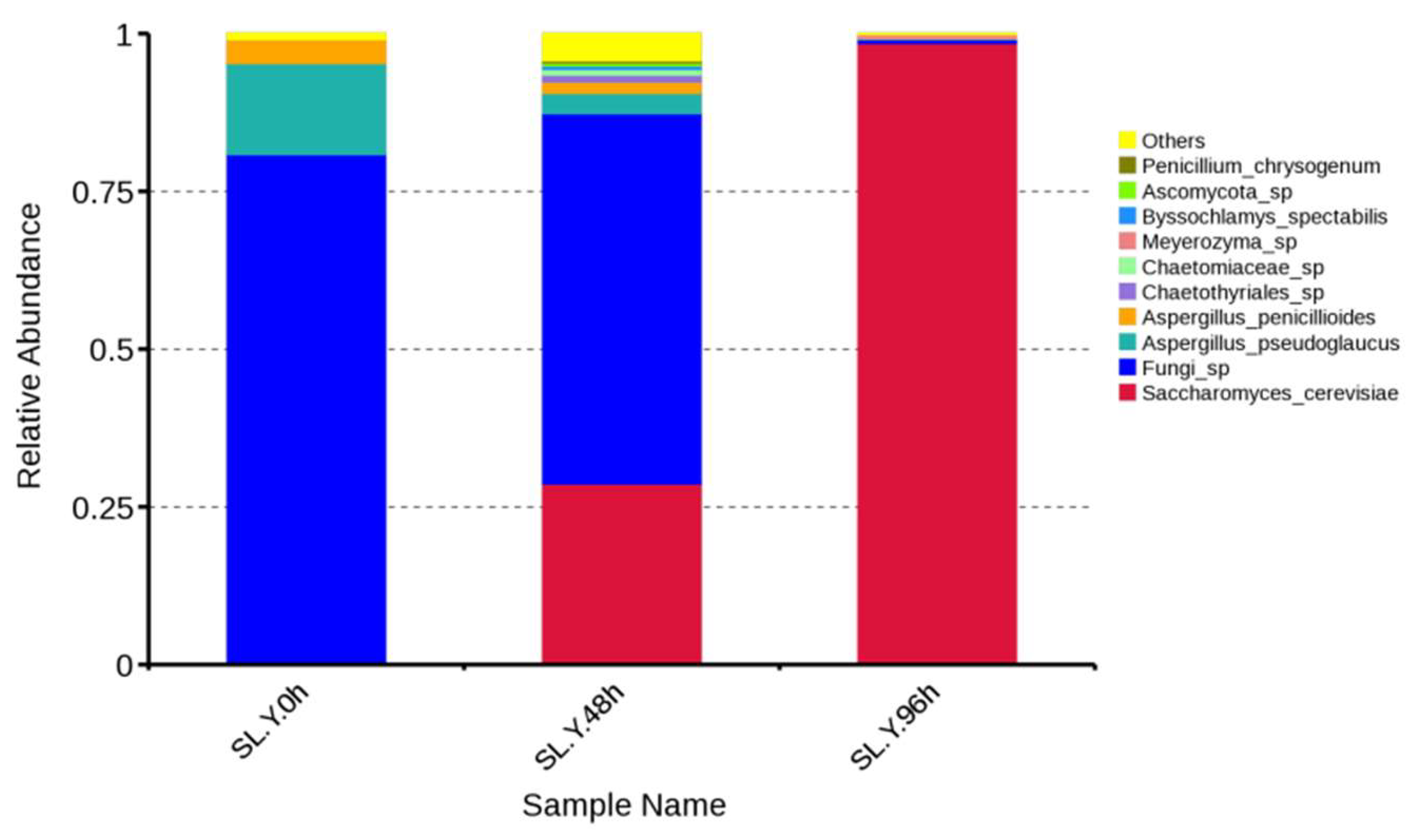
As shown in
Figure 9, the original fungi communities with high relative abundances in uncooked corn–soybean meal without inoculation were
Fungi sp.,
Aspergillus pseudoglaucus,
Aspergillus penicillioides, and “Others”. After 48 h fermentation with
Pediococcus acidilactici, the communities with decreasing relative abundances were
Fungi sp.,
Aspergillus pseudoglaucus, and
Aspergillus penicillioides. The fungi communities with increased relative abundance included
Saccharomyces cerevisiae, “Others”,
Chaetothyriales sp.,
Chaetomiaceae sp.,
Byssochlamys spectabilis,
Ascomycota sp., and
Penicillium chrysogenum. Therefore, the inoculation of
Pediococcus acidilactici had a great impact on the distribution of mycoflora in the system, and the relative abundance of
Fungi sp. decreased the most, while the diversity of mycoflora increased. After fermentation with
Bacillus subtilis and
Saccharomyces cerevisiae for 96 h, the absolute dominant community of mycoflora in the sample was
Saccharomyces cerevisiae, and the relative abundances of all the top ten fungi that appeared after 48 h, except
Saccharomyces cerevisiae, declined significantly or they disappeared. Meanwhile, a new microflora,
Meyerozyma sp., appeared in the fermentation system. Shi Changyou et al. studied the two-stage fermentation of corn–soybean meal with
Bacillus subsubis and
Enterococcus faecium, and the level of mold in the 96 h fermented feed dropped below the detection limit, which was similar to the results in this study [
29].
The fermented feed was sent to the Agricultural Products Testing Center at Heilongjiang Academy of Agricultural Sciences for
Salmonella detection, but no
Salmonella was detected (
Table S4).













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}