Abstract
The leachate from municipal solid waste transfer stations has a complex composition of pollutants, contains high concentrations of organic matter, and produces odors. The anaerobic digestion of leachate generates excessive volatile organic acids (VFAs), acidifying the digestate and destabilizing or even collapsing the anaerobic process. To solve these issues, we set up a facultative anaerobic system with air exposure and an obligate anaerobic system without air exposure to explore the impact of anaerobic degree on intermediate accumulation and anaerobic process stability. When the organic loading raised from 0.6 to 6.0 kg COD/(m3·d), VFA accumulation in the facultative system peaked at 3.30 g COD/L, and the chemical oxygen demand (COD) removal rate dropped from 95% to 60%. By contrast, the obligate system remained stable, with VFA concentrations below 0.50 g COD/L and COD removal rates between 85% and 99%. Microbial sequencing shows that VFA-oxidizing bacteria enhanced VFA degradation, such as Syntrophomonas and Smithella, which increased from 4.62% and 0.76% in the facultative system to 17.64% and 0.88% in the obligate system, respectively. Coenzyme F420 concentration as an indicator of methanogenic activity was 50% higher in the obligate system. Therefore, an obligate anaerobic system is promising to alleviate acidification for stable anaerobic digestion of leachate.
1. Introduction
Municipal waste transfer stations are vital in municipal solid waste (MSW) treatment, linking an MSW collection scheme and the final disposal facilities [1]. At transfer stations, fresh leachate is generated as a secondary pollutant, formed from the waste because of extrusion, waste decomposition, and infiltration of surface water during stacking and compression [2]. The leachate is typically 5% to 15% of the MSW mass [3]. According to the World Bank, the global production of MSW was 2.01 billion in 2016 and will reach 2.59 billion tons in 2030 [4]. Therefore, more than 129.5 million tons of fresh leachate will be generated from transfer stations in 2030. Fresh leachate has a potential hydrogen (pH) of 4.2 to 6.0, chemical oxygen demand (COD) of 40 to 70 g/L, biochemical oxygen demand (BOD5) of 15 to 25 g/L, ammonia nitrogen of 0.40 to 0.70 g/L, total phosphorus of 0.05 to 0.20 g/L, and suspended solids of 1.00 to 80.00 g/L [5]. Its direct discharges can pollute water and soil, significantly threatening the environment and public health [6]. Thus, the effective, economical, and environmentally friendly in situ treatment of fresh leachate is an urgent task.
Anaerobic digestion (AD) is the most effective technology for treating high-strength organic wastewater such as fresh leachate [7]. AD consists of four tandem biochemical reactions: hydrolysis, acidogenesis, acetogenesis, and methanogenesis [8]. Each reaction is carried out by certain microorganisms, and different functional species constitute a complex community. The AD of leachate can produce valuable methane with advantages of a high organic loading rate (OLR), low-solids biomass, and a low operating cost [9]. However, AD is limited by the difficulty in balancing hydrolytic, acidogenic, acetogenic, and methanogenic microbial activities. The major manifestation is the accumulation of volatile fatty acids (VFAs), destabilizing and lowering the efficiency of the anaerobic process [10].
VFAs, such as acetate, propionate, butyrate, and valerate, are essential intermediates of organic pollutants during acidogenesis. Among them, propionate, butyrate, and valerate are further converted to acetate and hydrogen via syntrophic VFA oxidizing bacteria. The accumulation of VFAs is due to an imbalance between the production of VFAs in the acidogenesis phase and the degradation of VFAs in the acetogenesis phase [10]. The balance is extremely sensitive since the concentrations of acetate and hydrogen must be low enough to allow thermodynamically favorable VFA degradation, but high enough for methanogens to grow [11]. Therefore, maintaining the metabolic balance between different functional species is critical. Current solutions to VFA accumulation aim to improve the reactor configuration, add reagents, regulate environmental factors, and implement bioaugmentation. Microbial diversity and methane production of food waste were improved in a two-phase anaerobic membrane bioreactor [12]. However, the two-phase system only avoids the inhibition of methanogens using acid production and does not solve the microbial metabolic imbalance. Increasing the abundance of propionate-oxidizing bacteria with bioaugmentation could alleviate VFA accumulation in anaerobic co-digestion of cattle manure and corn straw [13]. Nonetheless, the microbe introduced by a single bioaugmentation will not persist [14], and the bioaugmentation promotes only propionate oxidation without enhancing methanogenesis.
Most syntrophic VFA-oxidizing bacteria and methanogens are obligate anaerobes, and their metabolism is affected by anaerobic degree. Anaerobic degree indicates the extent of the anaerobic biochemical reaction as a progressive process, from facultative to obligate anaerobic [15]. Anaerobic degree is closely related to local environmental conditions (e.g., oxygen partial pressure, pH, oxidation-reduction potential, and hydraulic flushing intensity) and the succession of anaerobic microbial communities. Obligate anaerobes are sensitive to oxygen, slowing or stopping growth at oxygen volume concentrations above 0.5%, and facultative anaerobes grow slower at oxygen volume concentrations above 3% [16,17]. AD processes exposed to air reduce hydrolytic activity, methane yield, and methanogen abundance [18]. Furthermore, a lower oxidation-reduction potential (ORP) will reduce the accumulation of propionate in an anaerobic process [19]. These studies indicate that the growth of anaerobes and performance of anaerobic systems are influenced by oxygen partial pressure and ORP, factors related to anaerobic degree. However, the effects of anaerobic degree on VFA accumulation and the stability of the anaerobic process of waste leachate have not been reported. Therefore, this study aimed to address the low efficiency and stability of the anaerobic digestion of fresh leachate caused by VFA accumulation. We hypothesized that an obligate anaerobic system would enhance the diversity and activity of obligate anaerobes at high organic loading and rapidly consume accumulated VFA. To test the hypothesis, we assessed the effect of anaerobic degree on AD performance, intermediate accumulation, and the stability of the anaerobic processes in a facultative and an obligate anaerobic system.
2. Materials and Methods
2.1. Reactor Design and Operation
Two anaerobic baffled reactors (ABRs; ABR1; and ABR2) were operated continuously with a hydraulic retention time (HRT) of 5 d. ABR1 is a facultative anaerobic system with air exposure. ABR2 is an obligate anaerobic system without air exposure. Each ABR is made of Plexiglas panels 360 mm high, 90 mm wide, 270 mm long, and which have an effective working volume of 8.5 L. The ABR is divided into three compartments at a volume ratio of 1:2:3. The three compartments of ABR1 are ABR1-1, ABR1-2, and ABR1-3, whereas the three compartments of ABR2 are ABR2-1, ABR2-2, and ABR2-3. Each compartment includes an up-flow zone and a down-flow zone with a volume ratio of 5:1 (Figure S1). The ABRs were operated at room temperature (20 to 30 °C), and a heating rod kept the wastewater temperature at 25 ± 1 °C in winter. The mean ORP of ABR1-1, ABR1-2, ABR1-3, ABR2-1, ABR2-2, and ABR2-3 was −212 ± 4, −278 ± 13, −290 ± 16, −308 ± 13, −374 ± 27, and −386 ± 21 mV, respectively. Other conditions of the two systems were identical.
Wastewater quality and quantity fluctuate significantly in fresh leachate from transfer stations. To compare the resistance of the two systems to loading fluctuations, the two systems were operated in seven stages with the OLR progressively increasing from 0.6 to 6.0 kg COD/(m3·d) (Table 1). During the operation, AD performance, VFA accumulation, and anaerobic stability were monitored to investigate the differences in the capability of the two systems to cope with OLRs.

Table 1.
The operation stages for ABR1 and ABR2.
2.2. Seed Sludge and Feeding
Seed sludge was from the anaerobic tank for a leachate treatment facility at an MSW transfer station in Tianhe District (Guangzhou, China). The sludge was screened to remove impurities, washed three times, and loaded equally into each compartment of two ABRs. The inoculum volume for each compartment was 1.4 L, approximately half of the compartment volume; the initial sludge concentrations are shown in Table S1.
The two ABRs were fed with fresh leachate from an MSW transfer station in Tianhe District, Guangzhou, China. The main wastewater quality parameters of the fresh leachate were COD of 20 to 35 g/L, NH4+-N of 0.40 to 0.80 g/L, total phosphorus (TP) of 0.06 to 0.10 g/L, and pH of 3.5 to 5.5.
2.3. Analytical Methods
COD, total suspended solids (TSS), and volatile suspended solids (VSS) were analyzed according to the Standard Methods [20]. VFAs were analyzed using gas chromatography (GC) with a flame ionization detector (FID) (Tianmei, GC-7890, Shanghai, China). Samples for VFA analysis were centrifuged at 1095× g for 5 min, and the supernatant was filtered with 0.22 μm polyethersulfone membrane filters (Jingteng, Tianjin, China). The filtrate was acidified with 5% volume of formic acid (Aladdin, Shanghai, China). The pH was measured with a pH meter (PHS-3C, Sanxin, Shanghai, China). The ORP was measured using a composite electrode (501, Leici, Shanghai, China).
Because of the limitation of ABR1 to collect biogas in contact with air, the F420 index was used to characterize the methanogenic activity. The coenzyme F420 is a critical electron carrier in methanogens and a widely used indicator of methanogenesis [21,22]. The activity of coenzyme F420 was measured spectrophotometrically [23].
2.4. DNA Extraction, PCR Amplification, and High-Throughput 16S rRNA Gene Sequencing
Six suspended sludge samples were obtained from the six compartments of the two ABRs at the end of Stage IV and analyzed for microbial communities via high-throughput 16S rRNA gene sequencing. Genomic DNA was extracted from the sludge using an E.Z.N.A™ Mag-Bind Soil DNA Kit (Omega Bio-Tek, Guangzhou, China). The extracted DNA was quantified using a Qubit 3.0 DNA Assay (Thermo Fisher Scientific, Shanghai, China). Amplification of the V3-V4 region of the 16S rRNA gene of archaea was conducted with three rounds of nested polymerase chain reaction (PCR). The first round was conducted using primers GU1ST-340F and GU1ST-1000R (Shanghai Sangon Biotech Co., Shanghai, China). The second round was conducted using the first round PCR product with primers 349F and 806R [24]. The third round was conducted with Illumina bridge PCR-compatible primers. Amplification of the V3-V4 region of the 16S rRNA gene of bacteria was conducted with two rounds of nested PCR. The first round was conducted using primers 341F and 805R, and the second round was conducted with Illumina bridge PCR-compatible primers. DNA libraries were prepared by converging purified PCR products, and the library size was determined using 2% agarose gel electrophoresis. To obtain a uniform long cluster effect and high-quality sequencing data, we measured library concentration using a Qubit 3.0 fluorescence quantifier (Shanghai Sangon Biotech Co., Shanghai, China).
2.5. Date Analysis
The indicators monitored daily (pH and COD) were measured once for each sample, and the other indicators (ORP, F420, VFA, and sludge concentration) were measured three times for an arithmetic mean and a standard deviation. The arithmetic means and standard deviations were calculated using Microsoft Excel 2019 (Microsoft Office 2019, Microsoft Corporation, Redmond, WA, USA). The data were plotted with Origin 2019b (OriginLab Corporation, Northampton, MA, USA).
3. Results and Discussion
3.1. The Performance of the ABRs
To evaluate the performance of the two ABRs at different anaerobic degrees, the pH, COD removal rate, and F420 concentration in ABR1 and ABR2 were monitored (Figure 1). Stages I to III were the start-up and domestication period, where the pH between corresponding compartments of ABR1 and ABR2 differed by less than 0.5. Starting from Stage IV, the pH of ABR1 decreased sharply, where the minimum pH value of ABR1-1 was 2.2 lower than that of ABR2-1. Likewise, the COD removal rates of the two ABRs in Stages I to III were comparable and above 95%. The COD removal rate of ABR1 and ABR2 dropped sharply in Stage IV to a minimum of 60.19% and 85.58%, respectively, and gradually recovered in Stage V. In Stage IV, the COD removal rate of ABR1 significantly differed from that of ABR2, with the mean COD removal rates of 81.36% ± 10.22% and 91.39% ± 4.16%, respectively. Therefore, ABR2 was more stable and resistant to pulse OLRs. Meanwhile, the F420 level in ABR2 was 50% higher than that in ABR1, indicating ABR2 had enhanced methanogen activity.
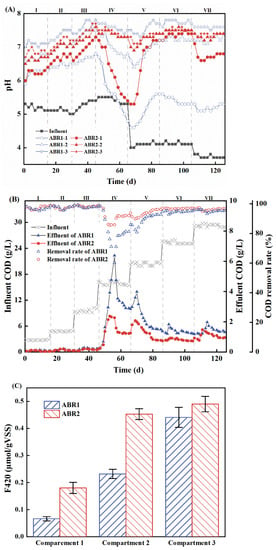
Figure 1.
pH (A), effluent COD and COD removal rates (B), and coenzyme F420 (C) of ABR1 and ABR2.
The high concentration of readily biodegradable organic matter in the fresh leachate causes rapid acidification, especially when a high OLR is applied [25]. After the acidic influent entered each compartment, the pH raised rapidly via biochemical processes and dilution. From Stages I to III, the pH of the two ABRs was stable at 6.5 ± 0.3 (ABR1-1 and ABR2-1), 7.0 ± 0.3 (ABR1-2 and ABR2-2), and 7.3 ± 0.3 (ABR1-3 and ABR2-3). However, the pH dropped dramatically when the OLR increased to 3.0 kg COD/(m3·d) in Stage IV, and the influent pH suddenly decreased to 4.0 because of the change in weather in Stage V. The drop in pH in each compartment varied, especially in ABR1-1 and ABR2-1, where the pH dropped from 6.9 to a minimum of 4.6 and from 7.3 to a minimum of 5.3, respectively. Therefore, ABR2 had significantly better buffering capacity against influent shock loading. From Stages VI to VII, the pH was stabilized at 5.4 ± 0.2 (ABR1-1), 7.2 ± 0.1 (ABR1-2), 7.7 ± 0.1 (ABR1-3), 7.0 ± 0.3 (ABR2-1), 7.4 ± 0.1(ABR2-2), and 7.4 ± 0.1 (ABR2-3). For comparison, the pH fluctuated in ABR2 within a narrow range, closer to the optimal pH range of 6.8 to 7.2 for methanogens [26]. These results demonstrate that the obligate anaerobic system has better pH buffering capacity and is more stable under high organic loading without alkalinity addition.
The effluent COD of ABR1 and ABR2 was approximately 0.2 g/L from Stages I to III. When the OLR rose to 3.0 kg COD/(m3·d) in Stage IV, the effluent of the two systems deteriorated at different degrees. The effluent COD concentrations of ABR1 and ABR2 peaked at 6.38 g/L (60.19% removal) and 2.37 g/L (85.58% removal), respectively. From Stages V to VII, the effluent COD of ABR1 and ABR2 decreased and stabilized at 1.45 ± 0.25 and 0.93 ± 0.17 g/L, respectively. The stable effluent COD levels were due to the succession of microbial populations when the organic loading increased from Stages IV to V, which enhanced the tolerance of the system to high organic loading [27]. The largest difference between the COD removal rates of the two ABRs existed in ABR1-1 and ABR2-1. From Stages IV to VII, ABR2-1 removed over half of the influent COD in ABR2, while ABR1-1 removed only 23.15% ± 12.31% (Table 2). The low COD removal rate of ABR1-1 might be associated with its low pH value (5.3 ± 0.4). Anaerobes are sensitive to pH fluctuations, and abrupt changes in pH would inhibit the microbial activity of methanogens [26]. Low pH would inhibit microbial activity and reduce the hydrolysis and methanogenic capacity, hindering the anaerobic degradation of VFA [28]. Even worse, VFA accumulation would further decrease the pH of the system [29]. Fortunately, the obligate anaerobic system alleviated pH fluctuation by maintaining excellent buffering performance and system stability.
Table 2.
The mean COD removal (%) of each compartment in ABR1 and ABR2.
3.2. The Accumulative Concentration and Type of VFA
During AD processes, VFAs are requisite intermediates utilized by syntrophic VFA-oxidizing bacteria and methanogens to produce methane [31]. Low COD removal rates and acidification are caused by VFA accumulation [29]. VFAs are accumulated in ABR1 and ABR2 (Figure 2). From Stages I to III, VFA concentrations in the two ABRs were low, where ABR1-3 and ABR2-3 had VFA concentrations below 0.03 g COD/L, mainly composed of acetate and propionate. Starting from Stage IV, the VFA concentration of the ABRs increased sharply, corresponding to the increase in COD concentration. The VFA concentrations in ABR1-1, ABR1-2, and ABR1-3 peaked at 12.54, 4.82, and 3.30 g COD/L, respectively. By contrast, the VFA concentrations in ABR2-1, ABR2-2, and ABR2-3 peaked at 5.23, 1.30, and 0.50 g COD/L, respectively, much lower than those in ABR1. Therefore, the obligate anaerobic system (ABR2) is more favorable for VFA degradation and alleviating VFA accumulation. The predominant VFA in the two systems was propionate, which is more difficult to degrade than other VFAs because of the thermodynamic constraint of positive Gibbs free energy change at pH 7 for propionate (ΔG0′) (Equation (1)). The propionate concentrations in the effluent of ABR1 and ABR2 were 1.54 ± 0.21 and 0.58 ± 0.24 g COD/L in Stage IV, respectively. From Stages V to VII, the propionate concentrations in the effluent of ABR1 and ABR2 decreased to 0.32 ± 0.19 and 0.09 ± 0.12 g COD/L, respectively. Compared with the facultative anaerobic system, the obligate anaerobic system enhanced propionate degradation.
CH3CH2COO− + 3H2O → CH3COO− + HCO3− + H+ + 3H2 ΔG0′ = +76.1 kJ/mol
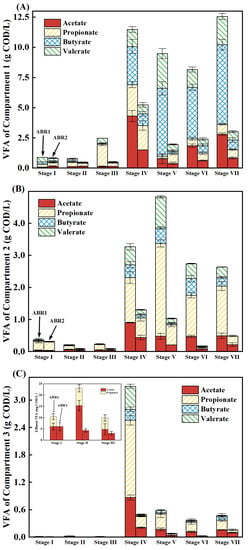
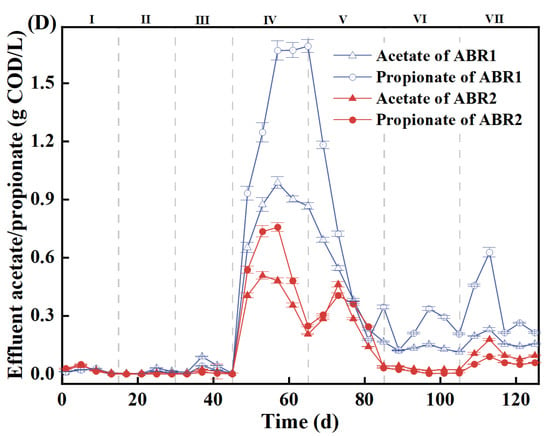
Figure 2.
The VFA of Compartment 1 (A), Compartment 2 (B), and Compartment 3 (C) and the concentration of acetate and propionate in the effluent of ABR1 and ABR2 (D).
As basic operating parameters, pH and anaerobic degree control the distribution and activity of functional microbes, resulting in various profiles of VFAs in different compartments. The VFA composition and concentration in ABR1-1 and ABR2-1 differed significantly (Figure 2A). ABR1-1 was dominated by butyrate and valerate, while ABR2-1 was dominated by acetate and propionate. This might be related to the different pH of the two compartments, which would cause different types of primary fermentation [32]. From Stages IV to VII, the VFA concentration in ABR1-1 was 2.2 to 4.8 times higher than that in ABR2-1. When the influent OLR was 3.0 kg COD/(m3·d), the VFA of ABR1-1 exceeded 11.00 g COD/L, deteriorating the effluent quality and destabilizing the system. The predominant VFA in ABR1-2 and ABR2-2 was propionate (Figure 2B). In ABR1-2, VFA peaked at 4.82 g COD/L with 2.89 g COD/L of propionate. The peak VFA concentration in ABR2-2 was 1.30 g COD/L with 0.61 g COD/L of propionate, a 78.8% decrease in propionate compared with that of ABR1-2. The propionate concentration in ABR2-2 was always lower than that in ABR2-1, while in ABR1, the propionate concentration increased from 0.54 ± 0.26 g COD/L in ABR1-1 to 1.91 ± 0.71 g COD/L in ABR1-2 from Stages IV to VII. The increase in propionate concentration indicates that the main biochemical reaction in ABR1-2 was the conversion of long-chain fatty acids to propionate and acetate via β-oxidation rather than the degradation of propionate [33]. The significant difference in the VFA concentration of ABR1-2 and ABR2-2 demonstrates that VFA degradation was much stronger in ABR2-2 than in ABR1-2.
The VFA accumulation in ABR1-3 and ABR2-3 represents the effluent quality and treatment efficiency. From Stages I to III, the effluent of ABR1 comprised mainly acetate and propionate with a total concentration of 0.02 to 0.03 g COD/L, whereas the effluent of ABR2 contained only acetate with a concentration of approximately 0.01 g COD/L. When the OLR increased to 3.0 kg COD/(m3·d) in Stage IV, VFA accumulated in ABR1, with the total concentration increased from 3.27 g COD/L in ABR1-2 to 3.30 g COD/L in ABR1-3. The peak concentrations of acetate and propionate in ABR1-2 and ABR 1-3 reached 0.99 and 1.69 g COD/L, respectively (Figure 2D). By contrast, the VFA in ABR2 decreased from 1.30 g COD/L in ABR2-2 to 0.49 g COD/L in ABR2-3 at the same time. This is exactly why the effluent COD concentrations between ABR1 and ABR2 were significantly different. ABR1-3, which was the main methanogenic phase like ABR2-3, showed no significant degradation of VFAs, indicating the absence of propionate-oxidizing bacteria and acetoclastic methanogens [34]. Stimulated by the high substrate concentration in Stage IV, the corresponding functional microorganisms grew rapidly [35]. The increase in the relative abundance of functional microbes enhanced the resistance to high OLR of anaerobic activated sludge [27]. Therefore, after the restoration of stability, the buffering capacity and load resistance of ABR1 were enhanced and the VFA degradation rate increased. From Stages V to VII, although the gap between the two systems narrowed, the effluent VFA of ABR1 (0.48 ± 0.09 g COD/L) was higher than that of ABR2 (0.09 ± 0.05 g COD/L).
The VFA distribution in the two ABRs indicates that the kinetics of biochemical reactions in ABR2 were faster than those in ABR1. When the VFA concentration in ABR2 decreased substantially with the increment of the compartment, VFA degraded little in ABR1, resulting in VFA accumulation. Furthermore, the VFA concentration in ABR1 was always higher than that in ABR2, indicating that ABR1 was limited in acetogenesis and methanogenesis. As a result, the high pH and low ORP in the obligate anaerobic system are more favorable for the degradation of VFA.
3.3. Microbial Diversity and Community Analysis
Stage IV was the key period at which ABR1 and ABR2 differed significantly. To explore the underlying mechanisms for the difference, the microbial community composition and diversity of the two ABRs were analyzed at the end of Stage IV.
Alpha diversity indices indicate the diversity of microbial communities within samples. Among them, Shannon and Shannoneven indices characterize the diversity and evenness of species distribution in the communities. Chao and Ace indices estimate the total number of species in the sample [36]. The values of the four alpha diversity indices of the compartments of ABR2 are higher than the corresponding compartments of ABR1, with significant differences in Compartment one (Table 3). The Shannon index of ABR1-1 is 2.303, while the other compartments of two ABRs exceed 4. The low microbial diversity in ABR1-1 resulted in its low resistance to environmental stress [37], corresponding to the low pH and low COD removal rate of ABR1-1 (Figure 1). The more abundant species in ABR2 promoted the metabolism (e.g., organic matter utilization and degradation) of the microbial community and maintained the stability of the anaerobic system.
Table 3.
Alpha diversity indices of ABR1 and ABR2.
In addition to microbial diversity, principal component analysis (PCA) on six samples showed the differences and similarities among samples by analyzing the composition of communities. The more similar the composition of samples, the closer they were in the PCA plot. PC1 and PC2 contributed 59.18% and 23.66%, respectively, to the community variations (Figure 3). The corresponding compartments of ABR1 and ABR2 are far apart in the PCA plot, and their microbial communities significantly differed, especially between ARB1-1 and ARB2-1, consistent with previous analysis (Table 3). The sample similarity between ABR1-3 and ABR2-2 was presumably due to the similar environment of these two compartments, such as similar OLR and VFA concentrations. The mean VFA concentrations of ABR1-3 and ABR2-2 were 0.68 and 0.49 g COD/L, respectively, and acetate and propionate were the main components with an average total proportion of 89.94% and 86.79%, respectively. In summary, although the initial state of sludge in each compartment was the same, varying environmental conditions affected the structure of microbial communities [38]. This discrepancy in microbial community structure is the source of the difference in the performance of the two ABRs.
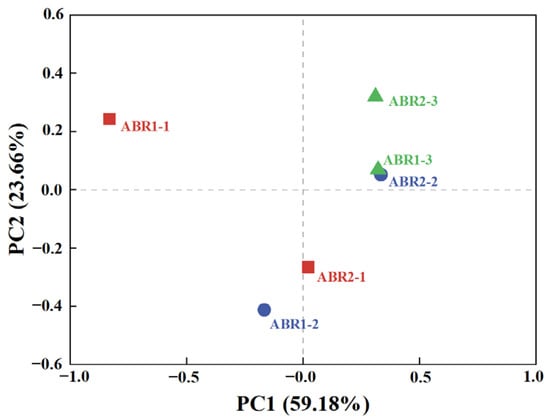
Figure 3.
Principal component analysis (PCA) of microbial communities in the two ABRs.
Bacterial communities of the two ABRs mainly contained nine phyla, among which Firmicutes (13.26% to 38.3%), Bacteroidetes (8.04% to 62.29%), Chloroflexi (0.19% to 20.52%), Proteobacteria (2.55% to 15.02%), Synergistetes (0.21% to 2.07%), and Actinobacteria (0.2% to 2.07%) were common fermentative groups in mesophilic anaerobic reactors (Figure 4A) [39]. Firmicutes and Bacteroidetes hydrolyze polysaccharides such as cellulose and starch [40]. The relative abundance of Synergistetes was 1.31% ± 0.78% in ABR1 and 1.85% ± 0.18% in ABR2. Synergistetes can participate in syntrophic methanogenesis processes [41]. Planctomycetes were uncommon in other anaerobic systems; however, the relative abundance of Planctomycetes was 13.75% to 44.06% in all compartments except for ABR1-1. In addition to Anammox and the obligate aerobes Planctomyces and Pirellula, Planctomycetes include some genera isolated from lakes and wetlands for fermentation [42]. Unclassified Planctomycetaceae, presumably fermenting bacteria, accounted for a significant proportion (26.71% ± 10.24%) in all compartments except for ABR1-1 (Figure 4B).
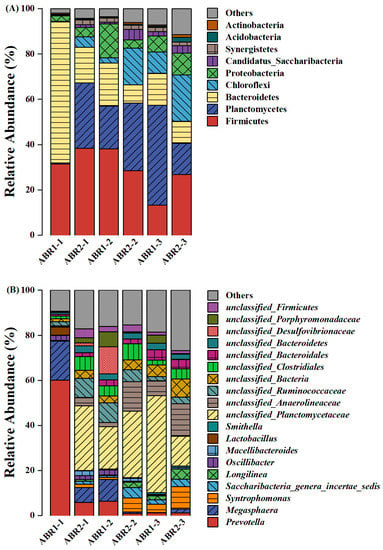
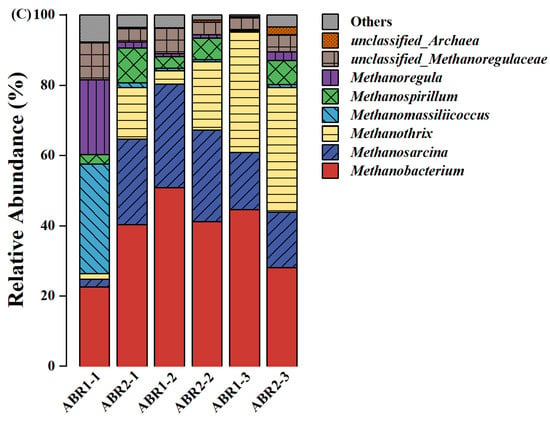
Figure 4.
The relative abundance of dominant bacteria at the phylum (A) and the genus (B) level and dominant archaea at the genus level (C).
ABR1-1 and ABR2-1 differed in bacterial composition at the genus level. The bacterial diversity of ABR1-1 was much lower than that of ABR2-1, with two genera, Prevotella (60.0%) and Megasphaera (17.4%), dominating and accounting for 77.4% of the total bacteria in ABR1-1 (Figure 4B). The abundance of Prevotella and Megasphaera in the other compartments was 3.78% ± 3.76% and 3.06% ± 2.44%, respectively. The relative abundance of syntrophic VFA-oxidizing bacteria in ABR1 (2.77% ± 2.67%) and ABR2 (9.24% ± 4.63%) also differed. VFA-oxidizing bacteria are necessary for the conversion of intermediate VFA to methane. Prevotella mainly breaks down proteins and carbohydrates [43], and Megasphaera utilizes fructose and lactic acid to produce hydrogen and organic acids [44]. The biochemical reactions in ABR1-1 were limited to hydrolysis and acidogenesis, where VFA accumulated to 11.00 g COD/L. The relative abundances of Syntrophomonas, Saccharibacteria-genera-incerae-sedis, and Smithella in ABR1 were 1.54% ± 1.55%, 0.98% ± 0.78%, and 0.25% ± 0.35%, respectively, and were 5.88% ± 3.41%, 3.07% ± 1.35%, and 0.29% ± 0.23%, respectively, in ABR2. Syntrophomonas is a syntrophic butyrate-oxidizing bacterium that oxidizes saturated fatty acids to produce acetate and hydrogen [45]. Most Saccharibacteria-genera-incerae-sedis can utilize glucose, amino acids, and butyrate [46]. Furthermore, Smithella utilizes propionate in the context of co-nutrition with H2/formate utilization partners [47]. Smithella includes S. propionica, which is the most discussed among syntrophic propionate-oxidizing bacteria that metabolize propionate through their unique pathway [48,49]. These functional bacteria, whose relative abundance was much higher in ABR2 than in ABR1, are syntrophic VFA-oxidizing bacteria that can oxidize saturated fatty acids to produce acetate/hydrogen and provide electron donors for methanogenesis. Therefore, the VFA of ABR1-3 accumulated to 3.30 g COD/L, whereas the VFA concentration in ABR2-3 peaked at a much lower concentration, 0.49 g COD/L, resulting in the pH of ABR1-3 dropping to 4.1. In summary, the syntrophic VFA-oxidizing bacteria in the obligate anaerobic systems promoted VFA degradation and maintained system stability.
The dominant archaea at the gene level were Methanobacterium (22.5% to 50.8%), Methanosarcina (2.21% to 29.48%), Methanothrix (1.66% to 35.52%), Methanomassiliicoccus (0.09% to 31.12%), Methanospirillum (0.27% to 9.98%), and Methanoregula (0.36% to 21.26%) (Figure 4C). Methanobacterium was the dominant archaeal genus in all compartments (44.20% ± 3.70%) except for ABR1-1 (22.5%) and ABR2-3 (28.14%), consistent with previous studies where Methanobacterium dominated methanogenic biocathodes [50,51]. The dominant genus of ABR1-1 was Methanomassiliicoccus (31.12%), which emerged during the recovery process after an organic overload and increased in abundance at high OLR [52]. The relative abundance of Methanomassiliicoccus in other compartments of two ABRs was 0.72% ± 0.38%, significantly lower than that of ABR1-1. The dominant genus of ABR2-3 was Methanothrix (Methanosaeta) (35.52%), which uses acetate to produce methane [53]. Likewise, Methanosarcina performs methanogenesis mainly via acetylation in combination with other methanogenesis pathways [54]. The relative abundance of Methanothrix and Methanosarcina was 13.25% ± 12.92% and 15.99% ± 9.64%, respectively, in ABR1 and 24.53% ± 12.72% and 22.03% ± 3.96%, respectively, in ABR2. Methanogens that can produce methane via direct acetate cleavage include only two genera (Methanosarcina and Methanothrix) [55]. The relative abundances of Methanospirillum in ABR1 and ABR2 were 2.13% ± 1.15% and 7.62% ± 1.47%, respectively. Methanospirillum degrades propionate in syntrophic cocultures with S. propionica through the C6-dismutation pathway [56]. Therefore, these genera are the key organisms in high-rate anaerobic reactors, as acetate and propionate are the dominant intermediates in methane production [57]. The high abundance of acetoclastic methanogens in ABR2 enhanced the utilization of acetate, and the rapid degradation of acetate could effectively mitigate the poor degradation of propionate, butyrate, and valerate. Overall, the obligate anaerobic conditions of ABR2 facilitated the growth of syntrophic VFA-oxidizing bacteria and acetoclastic methanogens, alleviating VFA accumulation and maintaining the stability of the system.
3.4. The Correlation between Environmental Factors and Functional Microbes
A canonical correlation analysis (CCA) map shows the relationship between environmental variables (ORP, VFA, and pH) and the dominant functional microbial genera in the ABRs (Figure 5). The correlation coefficients’ R’s between ORP, pH, and VFA and dominant functional microbial genera are 0.684, 0.964, and 0.986, respectively. The angle between ORP and VFA is acute, indicating a positive correlation [58]. Therefore, low ORP promotes the degradation of VFAs, supporting the conclusion that an obligate anaerobic system mitigates VFA accumulation. The angle between VFA and pH is obtuse (a negative correlation), indicating that VFA accumulation would decrease pH. Low pH hinders the metabolism of anaerobes and destabilizes the anaerobic process. The genera that favorite the degradation of VFAs, such as Syntrophomonas, Smithella, Methanothrix, and Methanosarcina, were more related to the samples from ABR2. Furthermore, these bacteria were in the opposite direction of ORP, indicating that low ORP promotes their proliferation. The samples from ABR2 are in the opposite direction of VFA for both bacteria and archaea, representing a negative correlation. Thus, VFA consumption was enhanced in ABR2. As a result, ORP, representing anaerobic degree in this study, significantly affects the composition of the functional microbial community and the system performance during anaerobic digestion.
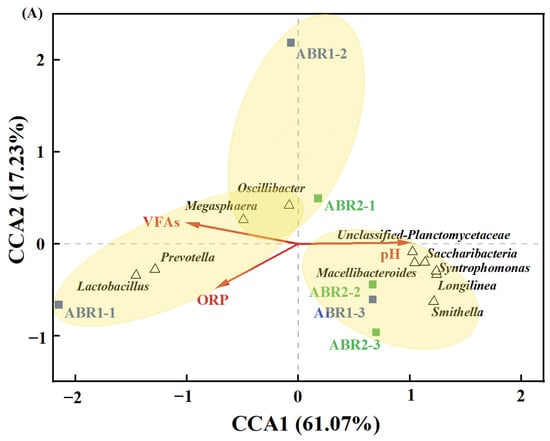
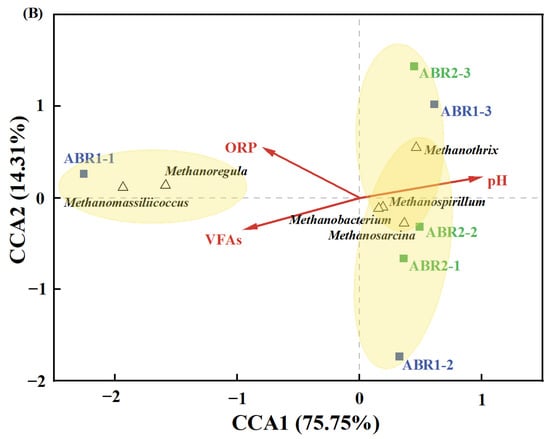
Figure 5.
Canonical correlation analysis (CCA) of environmental factors and major genera of bacteria (A) and archaea (B).
4. Conclusions
The effects of anaerobic degree on VFA accumulation and anaerobic process stability in facultative and obligate anaerobic systems treating fresh leachate were investigated. Compared with the facultative anaerobic system, the VFA accumulation in the obligate anaerobic system was more effectively alleviated from 3.30 to 0.49 g COD/L (84.98% reduction). The obligate anaerobic system achieved a dynamic balance by promoting the growth of syntrophic VFA-oxidizing bacteria and acetoclastic methanogens, maintaining the anaerobic stability (mean COD removal efficiency 95.84%). Therefore, obligate anaerobic digestion is a strategy to alleviate VFA accumulation and improve anaerobic process stability under high organic loading.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation9050465/s1, Figure S1: Schematic of the reactors; Table S1: Sludge concentration in each compartment of ABR1 and ABR2.
Author Contributions
Data curation, C.J.; Formal analysis, J.W.; Methodology, P.L.; Writing—original draft, B.L.; Writing—review and editing, C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Natural Science Foundation of Guangdong Province (China; 2020A1515011113), the Guangdong Science and Technology Program (China; 2020B121201003), the Applied Basic Research Project of Guangzhou (Guangdong province, China; 202002030455), and the Zhongshan Science and Technology Plan Project (Guangdong province, China; 2020AG021).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Thanks to the waste transfer station in Tianhe District, Guangzhou, China for the support and the provision of fresh leachate.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Arij, Y.; Fatihah, S.; Rakmi, A.R. Performance of pilot scale anaerobic biofilm digester (ABD) for the treatment of leachate from a municipal waste transfer station. Bioresour. Technol. 2018, 260, 213–220. [Google Scholar] [CrossRef]
- Chu, Y.; Fan, J.; Wang, R.; Liu, C.; Zheng, X. Preparation and immobilization of Bi2WO6/BiOI/g-C3N4 nanoparticles for the photocatalytic degradation of tetracycline and municipal waste transfer station leachate. Sep. Purif. Technol. 2022, 300, 121867. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, L.; Hu, S.; Diao, Y.; Jin, X.; Jin, P.; Chen, C.; Wu, X.; Wang, X.C. Electro-dissolved ozone flotation (E-DOF) integrated anoxic/oxic membrane reactor for leachate treatment from a waste transfer station. Environ. Sci. Pollut. Res. 2022, 29, 55803–55815. [Google Scholar] [CrossRef]
- Sondh, S.; Upadhyay, D.S.; Patel, S.; Patel, R.N. A strategic review on Municipal Solid Waste (living solid waste) management system focusing on policies, selection criteria and techniques for waste-to-value. J. Clean. Prod. 2022, 356, 131908. [Google Scholar] [CrossRef]
- Wang, X.; Xie, B.; Wu, D.; Hassan, M.; Huang, C. Characteristics and risks of secondary pollutants generation during compression and transfer of municipal solid waste in Shanghai. Waste Manag. 2015, 43, 1–8. [Google Scholar] [CrossRef]
- Thanh, B.X.; Dan, N.P.; Visvanathan, C. Low flux submerged membrane bioreactor treating high strength leachate from a solid waste transfer station. Bioresour. Technol. 2013, 141, 25–28. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.; Qian, G.; Liu, J. Effective anaerobic biodegradation of municipal solid waste fresh leachate using a novel pilot-scale reactor: Comparison under different seeding granular sludge. Bioresour. Technol. 2014, 165, 152–157. [Google Scholar] [CrossRef]
- Fernández-Rodríguez, J.; Pérez, M.; Romero, L.I. Semicontinuous Temperature-Phased Anaerobic Digestion (TPAD) of Organic Fraction of Municipal Solid Waste (OFMSW). Comparison with single-stage processes. Chem. Eng. J. 2016, 285, 409–416. [Google Scholar] [CrossRef]
- Nabi, M.; Liang, H.; Cheng, L.; Yang, W.; Gao, D. A comprehensive review on the use of conductive materials to improve anaerobic digestion: Focusing on landfill leachate treatment. J. Environ. Manag. 2022, 309, 114540. [Google Scholar] [CrossRef]
- Lv, N.; Zhao, L.; Wang, R.; Ning, J.; Pan, X.; Li, C.; Cai, G.; Zhu, G. Novel strategy for relieving acid accumulation by enriching syntrophic associations of syntrophic fatty acid-oxidation bacteria and H2/formate-scavenging methanogens in anaerobic digestion. Bioresour. Technol. 2020, 313, 123702. [Google Scholar] [CrossRef]
- Dolfing, J. Thermodynamic Constraints on Syntrophic Acetate Oxidation. Appl. Environ. Microbiol. 2014, 80, 1539–1541. [Google Scholar] [CrossRef]
- Amha, Y.M.; Corbett, M.; Smith, A.L. Two-Phase Improves Performance of Anaerobic Membrane Bioreactor Treatment of Food Waste at High Organic Loading Rates. Environ. Sci. Technol. 2019, 53, 9572–9583. [Google Scholar] [CrossRef]
- Xu, X.; Sun, Y.; Sun, Y.; Li, Y. Bioaugmentation improves batch psychrophilic anaerobic co-digestion of cattle manure and corn straw. Bioresour. Technol. 2022, 343, 126118. [Google Scholar] [CrossRef]
- Rhee, C.; Park, S.G.; Kim, D.W.; Yu, S.I.; Shin, J.; Hwang, S.; Shin, S.G. Tracking microbial community shifts during recovery process in overloaded anaerobic digesters under biological and non-biological supplementation strategies. Bioresour. Technol. 2021, 340, 125614. [Google Scholar] [CrossRef]
- Rittmann, B.E.; Mccarty, P.L. Environmental Biotechnology Principles & Applications; McGraw-Hill Education: New York, NY, USA, 2014; Volume xiv. [Google Scholar]
- Loesche, W.J. Oxygen sensitivity of various anaerobic bacteria. Appl. Microbiol. 1969, 18, 723. [Google Scholar] [CrossRef]
- Li, J.; Ran, X.; Zhou, M.; Wang, K.; Wang, H.; Wang, Y. Oxidative stress and antioxidant mechanisms of obligate anaerobes involved in biological waste treatment processes: A review. Sci. Total Environ. 2022, 838, 156454. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J.; Gao, L.; Yu, J.; Yuan, X.; Zhu, W.; Wang, X.; Cui, Z. Aerobic deterioration of corn stalk silage and its effect on methane production and microbial community dynamics in anaerobic digestion. Bioresour. Technol. 2018, 250, 828–837. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, Q.; Li, F.T. Avoiding propionic acid accumulation in the anaerobic process for biohydrogen production. Biomass Bioenergy 2006, 30, 177–182. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water and Wastewater, 23rd ed.; APHA: Washington, DC, USA, 2017. [Google Scholar]
- Guo, Q.; Wang, Y.N.; Qian, J.; Zhang, B.; Hua, M.; Liu, C.; Pan, B. Enhanced production of methane in anaerobic water treatment as mediated by the immobilized fungi. Water Res. 2021, 190, 116761. [Google Scholar] [CrossRef]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260–296. [Google Scholar] [CrossRef]
- Heine-Dobbernack, E.; Schoberth, S.M.; Sahm, H. Relationship of Intracellular Coenzyme F420 Content to Growth and Metabolic Activity of Methanobacterium bryantii and Methanosarcina barkeri. Appl. Environ. Microbiol. 1988, 54, 454–459. [Google Scholar] [CrossRef]
- Yan, P.; Zhao, Y.; Zhang, H.; Chen, S.; Zhu, W.; Yuan, X.; Cui, Z. A comparison and evaluation of the effects of biochar on the anaerobic digestion of excess and anaerobic sludge. Sci. Total Environ. 2020, 736, 139159. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Wu, B.; Gu, L.; Deng, R. Upgrade the high-load anaerobic digestion and relieve acid stress through the strategy of side-stream micro-aeration: Biochemical performances, microbial response and intrinsic mechanisms. Water Res. 2022, 221, 118850. [Google Scholar] [CrossRef]
- Ali, S.; Hua, B.; Huang, J.J.; Droste, R.L.; Zhou, Q.; Zhao, W.; Chen, L. Effect of different initial low pH conditions on biogas production, composition, and shift in the aceticlastic methanogenic population. Bioresour. Technol. 2019, 289, 121579. [Google Scholar] [CrossRef]
- Lu, F.; Huang, L.; Qian, F.; Jiang, Q.; Khan, S.; Shen, P. Resistance of anaerobic activated sludge acclimated by different feeding patterns: Response to different stress shocks. Water Sci. Technol. 2022, 85, 3023–3035. [Google Scholar] [CrossRef]
- Latif, M.A.; Mehta, C.M.; Batstone, D.J. Influence of low pH on continuous anaerobic digestion of waste activated sludge. Water Res. 2017, 113, 42–49. [Google Scholar] [CrossRef]
- Wang, G.; Li, Q.; Gao, X.; Wang, X.C. Sawdust-Derived Biochar Much Mitigates VFAs Accumulation and Improves Microbial Activities To Enhance Methane Production in Thermophilic Anaerobic Digestion. ACS Sustain. Chem. Eng. 2019, 7, 2141–2150. [Google Scholar] [CrossRef]
- Yu, H.; Lai, B.; Yang, H.; Rong, H.; Liang, H.; Qu, F. In situ probing methanogenesis in anaerobic wastewater treatment using front-face excitation-emission matrix (FF-EEM) fluorescence. J. Clean. Prod. 2023, 387, 135734. [Google Scholar] [CrossRef]
- Zhang, C.; Su, H.; Baeyens, J.; Tan, T. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sustain. Energy Rev. 2014, 38, 383–392. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, Y.; Quan, X.; Zhang, Y. Towards engineering application: Potential mechanism for enhancing anaerobic digestion of complex organic waste with different types of conductive materials. Water Res. 2017, 115, 266–277. [Google Scholar] [CrossRef]
- Shin, H.S.; Kim, S.H.; Lee, C.Y.; Nam, S.Y. Inhibitory effects of long-chain fatty acids on VFA degradation and β-oxidation. Water Sci. Technol. 2003, 47, 139–146. [Google Scholar] [CrossRef]
- Mathai, P.P.; Nicholes, M.S.; Venkiteshwaran, K.; Brown, C.M.; Morris, R.L.; Zitomer, D.H.; Maki, J.S. Dynamic shifts within volatile fatty acid–degrading microbial communities indicate process imbalance in anaerobic digesters. Appl. Microbiol. Biotechnol. 2020, 104, 4563–4575. [Google Scholar] [CrossRef]
- Li, J.; Yan, H.; Chen, Q.; Meng, J.; Li, J.; Zhang, Y.; Jha, A.K. Performance of anaerobic sludge and the microbial social behaviors induced by quorum sensing in a UASB after a shock loading. Bioresour. Technol. 2021, 330, 124972. [Google Scholar] [CrossRef]
- Nie, S.A.; Lei, X.; Zhao, L.; Wang, Y.; Wang, F.; Li, H.; Yang, W.; Xing, S. Response of activity, abundance, and composition of anammox bacterial community to different fertilization in a paddy soil. Biol. Fertil. Soils 2018, 54, 977–984. [Google Scholar] [CrossRef]
- Duong, C.M.; Lim, T.-T. Optimization and microbial diversity of anaerobic co-digestion of swine manure with waste kitchen oil at high organic loading rates. Waste Manag. 2022, 154, 199–208. [Google Scholar] [CrossRef]
- Yin, Q.; Wang, Z.; Wu, G. Impacts of environmental factors on microbial diversity, distribution patterns and syntrophic correlation in anaerobic processes. Arch. Microbiol. 2019, 201, 603–614. [Google Scholar] [CrossRef]
- Antwi, P.; Li, J.; Boadi, P.O.; Meng, J.; Shi, E.; Xue, C.; Zhang, Y.; Ayivi, F. Functional bacterial and archaeal diversity revealed by 16S rRNA gene pyrosequencing during potato starch processing wastewater treatment in an UASB. Bioresour. Technol. 2017, 235, 348–357. [Google Scholar] [CrossRef]
- Xu, S.; Lu, W.; Liu, Y.; Ming, Z.; Liu, Y.; Meng, R.; Wang, H. Structure and diversity of bacterial communities in two large sanitary landfills in China as revealed by high-throughput sequencing (MiSeq). Waste Manag. 2017, 63, 41–48. [Google Scholar] [CrossRef]
- Sieber, J.R.; McInerney, M.J.; Gunsalus, R.P. Genomic Insights into Syntrophy: The Paradigm for Anaerobic Metabolic Cooperation. Annu. Rev. Microbiol. 2012, 66, 429–452. [Google Scholar] [CrossRef]
- Kulichevskaya, I.S.; Ivanova, A.A.; Suzina, N.E.; Rijpstra, W.I.C.; Sinninghe Damsté, J.S.; Dedysh, S.N. Paludisphaera borealis gen. nov., sp. nov., a hydrolytic planctomycete from northern wetlands, and proposal of Isosphaeraceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 837–844. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Nguyen, A.Q.; Johir, M.A.H.; Guo, W.; Ngo, H.H.; Chaves, A.V.; Nghiem, L.D. Application of rumen and anaerobic sludge microbes for bio harvesting from lignocellulosic biomass. Chemosphere 2019, 228, 702–708. [Google Scholar] [CrossRef]
- Fuentes, L.; Braga, L.; Castelló, E.; Etchebehere, C. Work scheme to isolate the different micro-organisms found in hydrogen-producing reactors: A study of effectiveness by pyrosequencing analysis. J. Appl. Microbiol. 2018, 125, 96–110. [Google Scholar] [CrossRef]
- Amha, Y.M.; Sinha, P.; Lagman, J.; Gregori, M.; Smith, A.L. Elucidating microbial community adaptation to anaerobic co-digestion of fats, oils, and grease and food waste. Water Res. 2017, 123, 277–289. [Google Scholar] [CrossRef]
- Kindaichi, T.; Yamaoka, S.; Uehara, R.; Ozaki, N.; Ohashi, A.; Albertsen, M.; Nielsen, P.H.; Nielsen, J.L. Phylogenetic diversity and ecophysiology of Candidate phylum Saccharibacteria in activated sludge. FEMS Microbiol. Ecol. 2016, 92, fiw078. [Google Scholar] [CrossRef]
- Kuever, J. The Family Syntrophaceae. In The Prokaryotes: Deltaproteobacteria and Epsilonproteobacteria; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 281–288. [Google Scholar] [CrossRef]
- Kim, W.; Shin, S.G.; Han, G.; Cho, K.; Hwang, S. Structures of microbial communities found in anaerobic batch runs that produce methane from propionic acid—Seeded from full-scale anaerobic digesters above a certain threshold. J. Biotechnol. 2015, 214, 192–198. [Google Scholar] [CrossRef]
- Feng, D.; Xia, A.; Huang, Y.; Zhu, X.; Zhu, X.; Show, P.-L.; Liao, Q. Continuous electromethanogenesis of propionate wastewater: Effect of external voltage and hydraulic retention time. Chem. Eng. J. 2023, 454, 140267. [Google Scholar] [CrossRef]
- Zhen, G.; Kobayashi, T.; Lu, X.; Xu, K. Understanding methane bioelectrosynthesis from carbon dioxide in a two-chamber microbial electrolysis cells (MECs) containing a carbon biocathode. Bioresour. Technol. 2015, 186, 141–148. [Google Scholar] [CrossRef]
- Lee, B.; Park, J.-G.; Shin, W.-B.; Tian, D.-J.; Jun, H.-B. Microbial communities change in an anaerobic digestion after application of microbial electrolysis cells. Bioresour. Technol. 2017, 234, 273–280. [Google Scholar] [CrossRef]
- Moestedt, J.; Muller, B.; Westerholm, M.; Schnurer, A. Ammonia threshold for inhibition of anaerobic digestion of thin stillage and the importance of organic loading rate. Microb. Biotechnol. 2016, 9, 180–194. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, H.; Liu, J.; Hazen, T.C.; Huang, V.; He, Q. Unexpected competitiveness of Methanosaeta populations at elevated acetate concentrations in methanogenic treatment of animal wastewater. Appl. Microbiol. Biotechnol. 2017, 101, 1729–1738. [Google Scholar] [CrossRef]
- He, P.; Duan, H.; Han, W.; Liu, Y.; Shao, L.; Lü, F. Responses of Methanosarcina barkeri to acetate stress. Biotechnol. Biofuels 2019, 12, 289. [Google Scholar] [CrossRef]
- Evans, P.N.; Boyd, J.A.; Leu, A.O.; Woodcroft, B.J.; Parks, D.H.; Hugenholtz, P.; Tyson, G.W. An evolving view of methane metabolism in the Archaea. Nat. Rev. Microbiol. 2019, 17, 219–232. [Google Scholar] [CrossRef]
- De Bok, F.A.M.; Stams, A.J.M.; Dijkema, C.; Boone, D.R. Pathway of Propionate Oxidation by a Syntrophic Culture of Smithella propionica and Methanospirillum hungatei. Appl. Environ. Microbiol. 2001, 67, 1800–1804. [Google Scholar] [CrossRef]
- Thomas, M.; William, M. Energy Conservation and Hydrogenase Function in Methanogenic Archaea, in Particular the Genus Methanosarcina. Microbiol. Mol. Biol. Rev. 2019, 83, e00020-19. [Google Scholar] [CrossRef]
- Guo, X.; Yu, H.; Yan, Z.; Gao, H.; Zhang, Y. Tracking variations of fluorescent dissolved organic matter during wastewater treatment by accumulative fluorescence emission spectroscopy combined with principal component, second derivative and canonical correlation analyses. Chemosphere 2018, 194, 463–470. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).