
The Effects of L-Lysine-α-oxidase Enzyme and Trichoderma harzianum Rifai Culture Liquid on the Formation of Biofilms by Uropathogenic Multiresistant E. coli

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Enzyme
2.2. Antibiotics Resistance Tests
- Imipenem, 10 mg/disk (Imipenem, IPM10–SD073, HiMedia, Mumbai, India);
- Ceftazidime, 30 µg/disk (Ceftazidime, CAZ30–SD062, HiMedia, Mumbai, India);
- Ceftriaxone, 30 µg/disk (Ceftriaxone, CTR30–SD065, HiMedia, Mumbai, India);
- Ceftazidime with clavulanic acid, 30/10 µg/disc (Ceftazidime/Clavulanic acid, CAC30/10–SD207, HiMedia, Mumbai, India);
- Amoxiclav, 30 µg/disk (Amoxyclav, AMC30–SD063, HiMedia, Mumbai, India);
- Nitrofurantoin, 200 µg/disk (Nitrofurantoin, NIT200–SD090, HiMedia, Mumbai, India);
- Ampicillin, 25 µg/disk (Ampicillin, AMP25–SD077, HiMedia, Mumbai, India);
- Trimethoprim, 30 µg/disk (Trimethoprim, TR30–SD149, HiMedia, Mumbai, India);
- Ciprofloxacin, 30 µg/disk (Ciprofloxacin, CIP30–SD142, HiMedia, Mumbai, India);
- Tetracycline, 30 µg/disk (Tetracycline, TE30–SD037, HiMedia, Mumbai, India);
- Ceftriaxone, 30 µg/disk (Ceftriaxone, CTR30–SD065, HiMedia, Mumbai, India).
2.3. Biofilm Formation Ability
2.4. Statistical Analysis
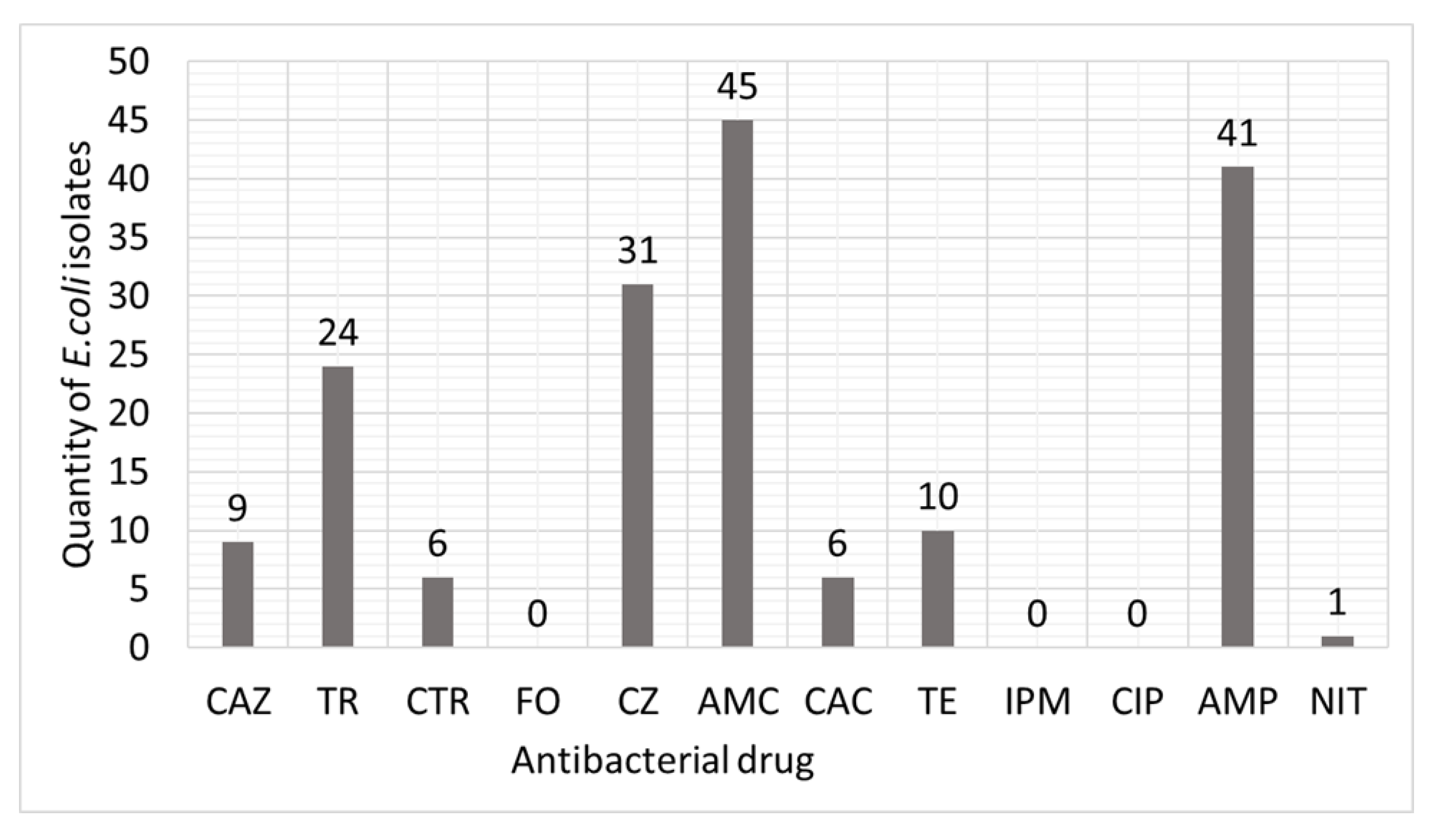
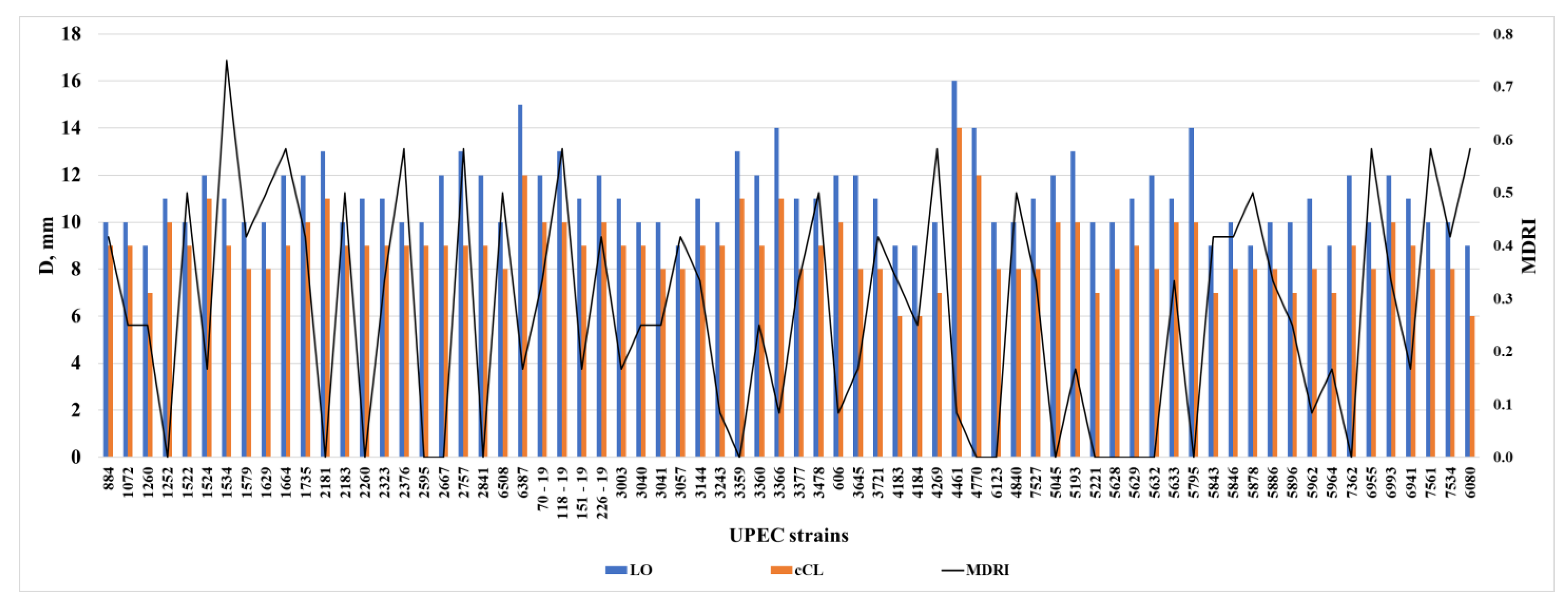
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Huemer, M.; Mairpady Shambat, S.; Brugger, S.D.; Zinkernagel, A.S. Antibiotic resistance and persistence-Implications for human health and treatment perspectives. EMBO Rep. 2020, 21, e51034. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.D.; Paladino, M.M.; Nagashima, K.; Brezel, E.R.; Holtzman, J.S.; Urso, S.J.; Ryno, L.M. Temperature-Dependent Influence of FliA Overexpression on PHL628 E. coli Biofilm Growth and Composition. Front. Cell. Infect. Microbiol. 2021, 11, 775270. [Google Scholar] [CrossRef] [PubMed]
- Rather, M.A.; Gupta, K.; Mandal, M. Microbial biofilm: Formation, architecture, antibiotic resistance, and control strategies. Braz. J. Microbiol. 2021, 52, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Baquero, F. Threats of antibiotic resistance: An obliged reappraisal. Int. Microbiol. Off. J. Span. Soc. Microbiol. 2021, 24, 499–506. [Google Scholar] [CrossRef]
- Eisenreich, W.; Rudel, T.; Heesemann, J.; Goebel, W. Link Between Antibiotic Persistence and Antibiotic Resistance in Bacterial Pathogens. Front. Cell. Infect. Microbiol. 2022, 12, 900848. [Google Scholar] [CrossRef]
- Gabibov, A.G.; Dontsova, O.A.; Egorov, A.M. Overcoming Antibiotic Resistance in Microorganisms: Molecular Mechanisms. Biochem. Biokhimiia 2020, 85, 1289–1291. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ikeda, M.; Okada, Y.; Higurashi, Y.; Okugawa, S.; Moriya, K. Clinical and Microbiological Characteristics of Recurrent Escherichia coli Bacteremia. Microbiol. Spectr. 2021, 9, e0139921. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, H.; Bi, D.; Khaledi, A.; Qiao, M. A systematic review and meta-analysis of antibiotic resistance patterns, and the correlation between biofilm formation with virulence factors in uropathogenic E. coli isolated from urinary tract infections. Microb. Pathog. 2020, 144, 104196. [Google Scholar] [CrossRef]
- Senyagin, A.N.; Larichev, A.F.; Smirnova, I.P.; Podoprigora, I.V. A Novel Express Method to Determine Activity of Antitumor Enzyme L-Lysine-α-Oxidase of Trichoderma harzianum Rifai F-180. Bull. Exp. Biol. Med. 2020, 169, 119–121. [Google Scholar] [CrossRef]
- Kot, B. Antibiotic Resistance among Uropathogenic Escherichia coli. Pol. J. Microbiol. 2019, 68, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Asadi Karam, M.R.; Habibi, M.; Bouzari, S. Urinary tract infection: Pathogenicity, antibiotic resistance and development of effective vaccines against Uropathogenic Escherichia coli. Mol. Immunol. 2019, 108, 56–67. [Google Scholar] [CrossRef]
- Lemke, P.; Zoheir, A.E.; Rabe, K.S.; Niemeyer, C.M. Microfluidic cultivation and analysis of productive biofilms. Biotechnol. Bioeng. 2021, 118, 3860–3870. [Google Scholar] [CrossRef]
- Raeispour, M.; Ranjbar, R. Antibiotic resistance, virulence factors and genotyping of Uropathogenic Escherichia coli strains. Antimicrob. Resist. Infect. Control. 2018, 7, 118. [Google Scholar] [CrossRef] [Green Version]
- York, A. Uropathogenic E. coli creates a memory. Nat. Rev. Microbiol. 2023, 21, 345. [Google Scholar] [CrossRef]
- Rathi, B.; Gupta, S.; Kumar, P.; Kesarwani, V.; Dhanda, R.S.; Kushwaha, S.K.; Yadav, M. Anti-biofilm activity of caffeine against uropathogenic E. coli is mediated by curli biogenesis. Sci. Rep. 2022, 12, 18903. [Google Scholar] [CrossRef]
- Ranjani, S.; Kathun, U.R.; Hemalatha, S. Silver Decorated Myconanoparticles Control Growth and Biofilm Formation in Uropathogenic E. coli. Appl. Biochem. Biotechnol. 2022, 194, 504–516. [Google Scholar] [CrossRef]
- Pousti, M.; Zarabadi, M.P.; Abbaszadeh Amirdehi, M.; Paquet-Mercier, F.; Greener, J. Microfluidic bioanalytical flow cells for biofilm studies: A review. Analyst 2018, 144, 68–86. [Google Scholar] [CrossRef]
- Henly, E.L.; Norris, K.; Rawson, K.; Zoulias, N.; Jaques, L.; Chirila, P.G.; Parkin, K.L.; Kadirvel, M.; Whiteoak, C.; Lacey, M.M.; et al. Impact of long-term quorum sensing inhibition on uropathogenic Escherichia coli. J. Antimicrob. Chemother. 2021, 76, 909–919. [Google Scholar] [CrossRef]
- Naziri, Z.; Kilegolan, J.A.; Moezzi, M.S.; Derakhshandeh, A. Biofilm formation by uropathogenic Escherichia coli: A complicating factor for treatment and recurrence of urinary tract infections. J. Hosp. Infect. 2021, 117, 9–16. [Google Scholar] [CrossRef]
- Katongole, P.; Nalubega, F.; Florence, N.C.; Asiimwe, B.; Andia, I. Biofilm formation, antimicrobial susceptibility and virulence genes of Uropathogenic Escherichia coli isolated from clinical isolates in Uganda. BMC Infect. Dis. 2020, 20, 453. [Google Scholar] [CrossRef]
- Maharjan, G.; Khadka, P.; Siddhi Shilpakar, G.; Chapagain, G.; Dhungana, G.R. Catheter-Associated Urinary Tract Infection and Obstinate Biofilm Producers. Can. J. Infect. Dis. Med. 2018, 2018, 7624857. [Google Scholar] [CrossRef] [Green Version]
- Raad, N.; Tandon, D.; Hapfelmeier, S.; Polacek, N. The stationary phase-specific sRNA FimR2 is a multifunctional regulator of bacterial motility, biofilm formation and virulence. Nucleic Acids Res. 2022, 50, 11858–11875. [Google Scholar] [CrossRef] [PubMed]
- Nwabuife, J.C.; Omolo, C.A.; Govender, T. Nano delivery systems to the rescue of ciprofloxacin against resistant bacteria “E. coli; P. aeruginosa; Saureus; and MRSA” and their infections. J. Control. Release Off. J. Control. Release Soc. 2022, 349, 338–353. [Google Scholar] [CrossRef]
- Alav, I.; Sutton, J.M.; Rahman, K.M. Role of bacterial efflux pumps in biofilm formation. J. Antimicrob. Chemother. 2018, 73, 2003–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gajdács, M.; Kárpáti, K.; Nagy, Á.L.; Gugolya, M.; Stájer, A.; Burián, K. Association between biofilm-production and antibiotic resistance in Escherichia coli isolates: A laboratory-based case study and a literature review. Acta Microbiol. Immunol. Hung. 2021, 68, 217–226. [Google Scholar] [CrossRef]
- Carvalho, F.M.; Mergulhão FJ, M.; Gomes, L.C. Using Lactobacilli to Fight Escherichia coli and Staphylococcus aureus Biofilms on Urinary Tract Devices. Antibiotics 2021, 10, 1525. [Google Scholar] [CrossRef]
- Kusakabe, H.; Kodama, K.; Kuninaka, A.; Yoshino, H.; Misono, H.; Soda, K. A new antitumor enzyme, L-lysine alpha-oxidase from Trichoderma viride. Purification and enzymological properties. J. Biol. Chem. 1980, 255, 976–981. [Google Scholar] [CrossRef]
- Anand, U.; Nandy, S.; Mundhra, A.; Das, N.; Pandey, D.K.; Dey, A. A review on antimicrobial botanicals, phytochemicals and natural resistance modifying agents from Apocynaceae family: Possible therapeutic approaches against multidrug resistance in pathogenic microorganisms. Drug Resist. Updates 2020, 51, 100695. [Google Scholar] [CrossRef]
- Lukasheva, E.V.; Babayeva, G.; Karshieva, S.S.; Zhdanov, D.D.; Pokrovsky, V.S. L-Lysine α-Oxidase: Enzyme with Anticancer Properties. Pharmaceuticals 2021, 14, 1070. [Google Scholar] [CrossRef]
- Pokrovsky, V.S.; Chepikova, O.E.; Davydov, D.Z.; Zamyatnin, A.A.; Lukashev, A.N., Jr.; Lukasheva, E.V. Amino Acid Degrading Enzymes and their Application in Cancer Therapy. Curr. Med. Chem. 2019, 26, 446–464. [Google Scholar] [CrossRef]
- Sobowale, A.; Uzoma, L.; Aduramigba-Modupe, A.; Bamkefa, B. Fungitoxicity of Trichoderma longibrachiatum (Rifai) Metabolites against Fusarium oxysporum, Aspergillus niger and Aspergillus tamarii. Am. J. Plant Sci. 2022, 13, 984–993. [Google Scholar] [CrossRef]
- Brunelle, J.L.; Green, R. One-dimensional SDS-polyacrylamide gel electrophoresis (1D SDS-PAGE). Methods Enzymol. 2014, 541, 151–159. [Google Scholar] [CrossRef]
- Olabode, I.R.; Sachivkina, N.; Karamyan, A.; Mannapova, R.; Kuznetsova, O.; Bobunova, A.; Zhabo, N.; Avdonina, M.; Gurina, R. In Vitro Activity of Farnesol against Malassezia pachydermatis Isolates from Otitis Externa Cases in Dogs. Animals 2023, 13, 1259. [Google Scholar] [CrossRef]
- Sachivkina, N.; Vasilieva, E.; Lenchenko, E.; Kuznetsova, O.; Karamyan, A.; Ibragimova, A.; Zhabo, N.; Molchanova, M. Reduction in Pathogenicity in Yeast-like Fungi by Farnesol in Quail Model. Animals 2022, 12, 489. [Google Scholar] [CrossRef]
- Zalewska-Piątek, B.M.; Piątek, R.J. Alternative treatment approaches of urinary tract infections caused by uropathogenic Escherichia coli strains. Acta Biochim. Pol. 2019, 66, 129–138. [Google Scholar] [CrossRef]
- Koroleva, E.A.; Soloveva, A.V.; Morgunova, E.Y.; Kapotina, L.N.; Luyksaar, S.I.; Luyksaar, S.V.; Bondareva, N.E.; Nelubina, S.A.; Lubenec, N.L.; Zigangirova, N.A.; et al. Fluorothiazinon inhibits the virulence factors of uropathogenic Escherichia coli involved in the development of urinary tract infection. J. Antibiot. 2023, 76, 279–290. [Google Scholar] [CrossRef]
- Takooree, H.; Aumeeruddy, M.Z.; Rengasamy KR, R.; Venugopala, K.N.; Jeewon, R.; Zengin, G.; Mahomoodally, M.F. A systematic review on black pepper (Piper nigrum L.): From folk uses to pharmacological applications. Crit. Rev. Food Sci. Nutr. 2019, 59, S210–S243. [Google Scholar] [CrossRef]
- Singh, K.; Gujju, R.; Bandaru, S.; Misra, S.; Babu, K.S.; Puvvada, N. Facet-Dependent Bactericidal Activity of Ag3PO4Nanostructures against Gram-Positive/Negative Bacteria. ACS Omega 2022, 7, 16616–16628. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Senyagin, A.; Sachivkina, N.; Das, M.; Semenova, V.; Kuznetsova, O.; Ibragimova, A. The Effects of L-Lysine-α-oxidase Enzyme and Trichoderma harzianum Rifai Culture Liquid on the Formation of Biofilms by Uropathogenic Multiresistant E. coli. Fermentation 2023, 9, 710. https://doi.org/10.3390/fermentation9080710
Senyagin A, Sachivkina N, Das M, Semenova V, Kuznetsova O, Ibragimova A. The Effects of L-Lysine-α-oxidase Enzyme and Trichoderma harzianum Rifai Culture Liquid on the Formation of Biofilms by Uropathogenic Multiresistant E. coli. Fermentation. 2023; 9(8):710. https://doi.org/10.3390/fermentation9080710
Chicago/Turabian StyleSenyagin, Alexander, Nadezhda Sachivkina, Milana Das, Valentina Semenova, Olga Kuznetsova, and Alfia Ibragimova. 2023. "The Effects of L-Lysine-α-oxidase Enzyme and Trichoderma harzianum Rifai Culture Liquid on the Formation of Biofilms by Uropathogenic Multiresistant E. coli" Fermentation 9, no. 8: 710. https://doi.org/10.3390/fermentation9080710