
The Effect of Bacillus velezensis LJ02 Compounded with Different Fungi on the Growth of Watermelon Seedlings and Microbial Community Structure
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Design
2.3. Disease Survey
2.4. Growth Index
2.5. Rhizosphere Soil Metagenome Analysis
2.5.1. Sample Collection
2.5.2. Extraction of DNA from Rhizosphere Soil
2.5.3. Metagenomic Data Analysis and Processing
2.6. Statistical Analysis
3. Results
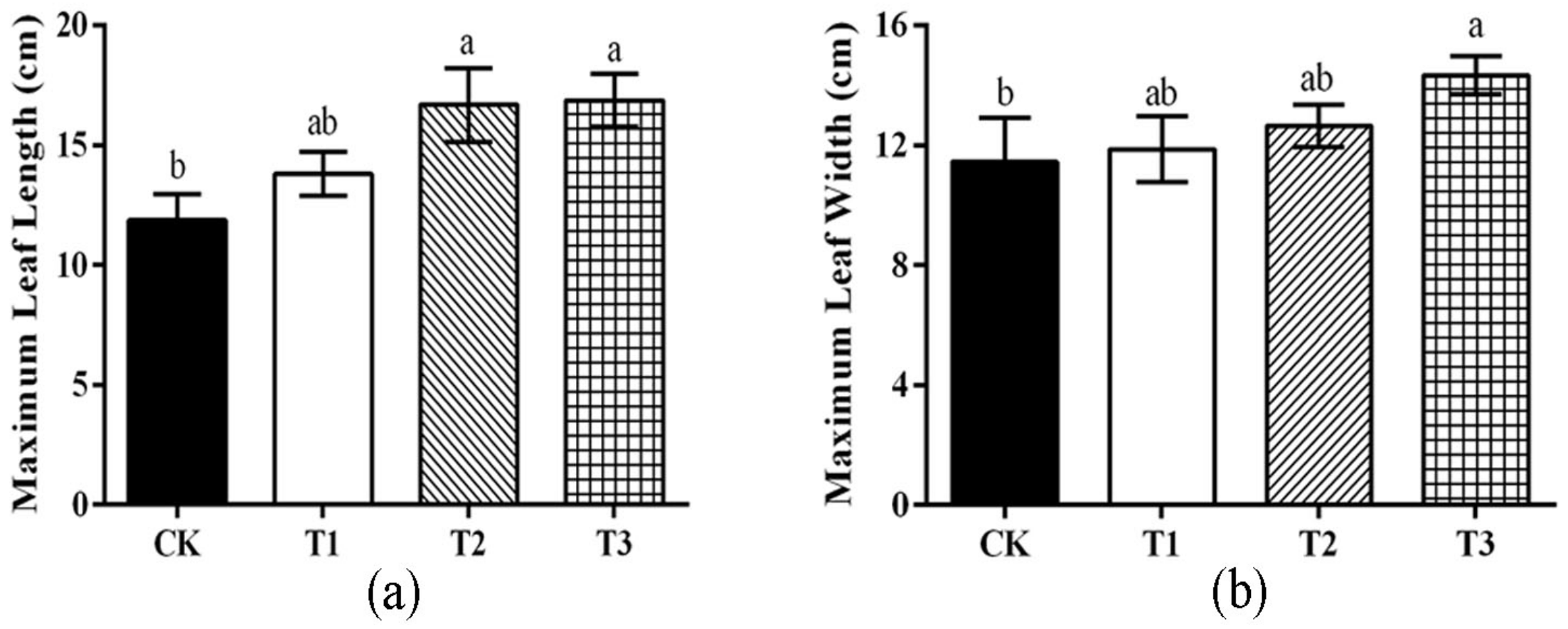
3.1. Effect of Different Treatments on the Growth of Watermelon Seedlings
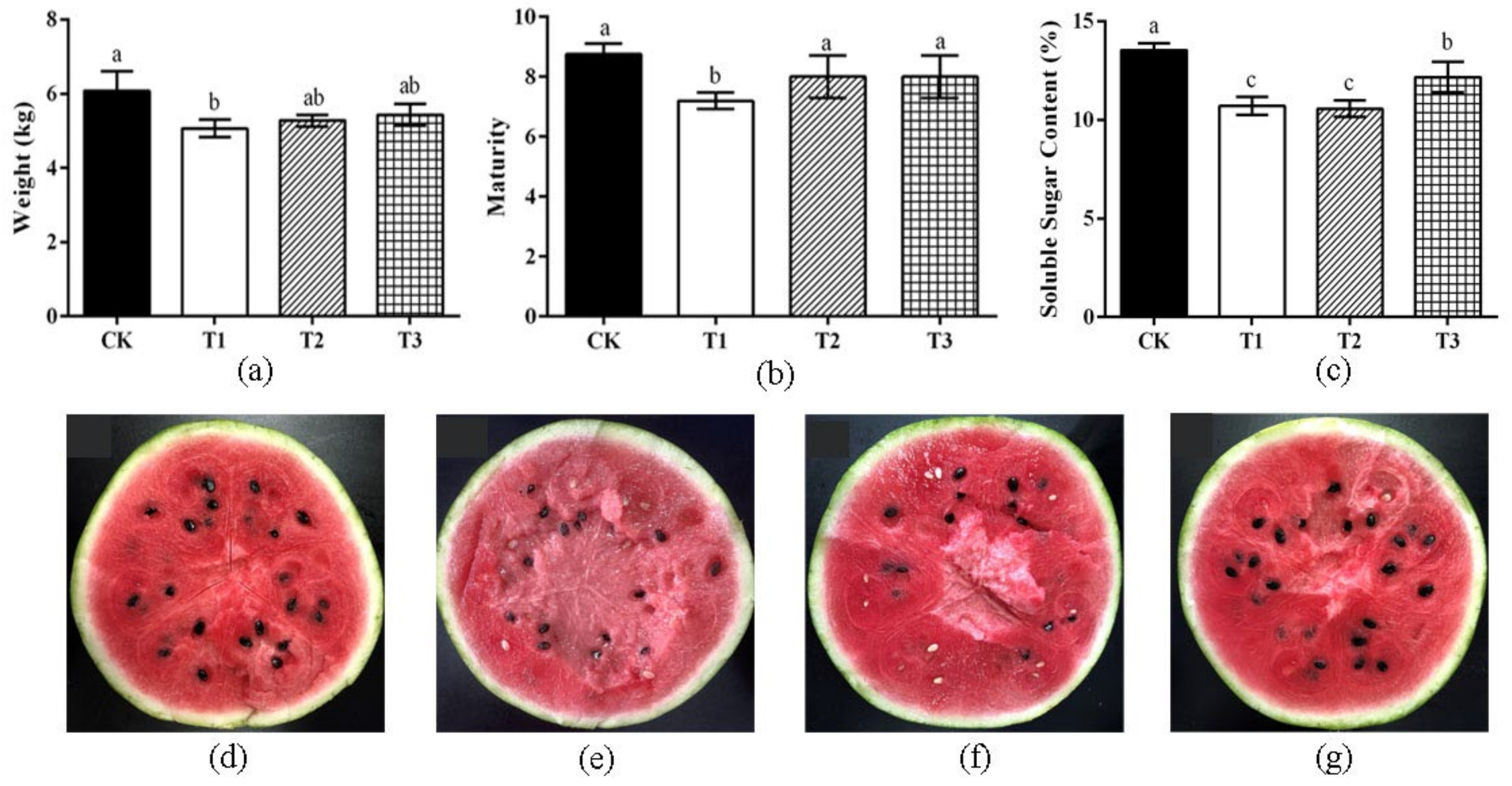
3.2. Effect of Different Treatments on the Quality of Watermelon Fruits
3.3. Effect of Different Treatments on Watermelon Gummy Stem Blight and Powdery Mildew
3.4. Effect of Different Treatments on Rhizosphere Soil Microorganisms in the Watermelon
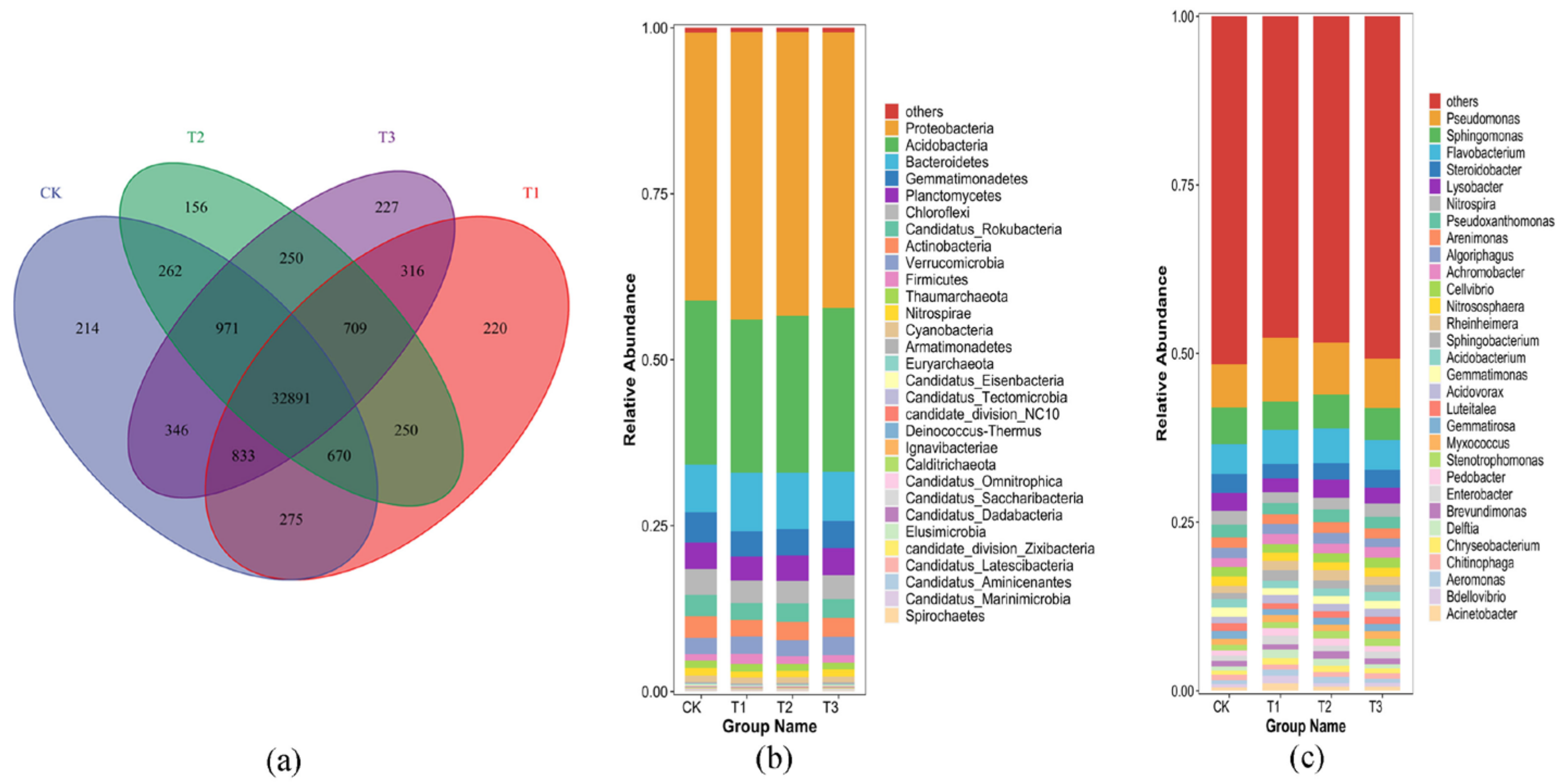
3.4.1. Microbial Community Structure
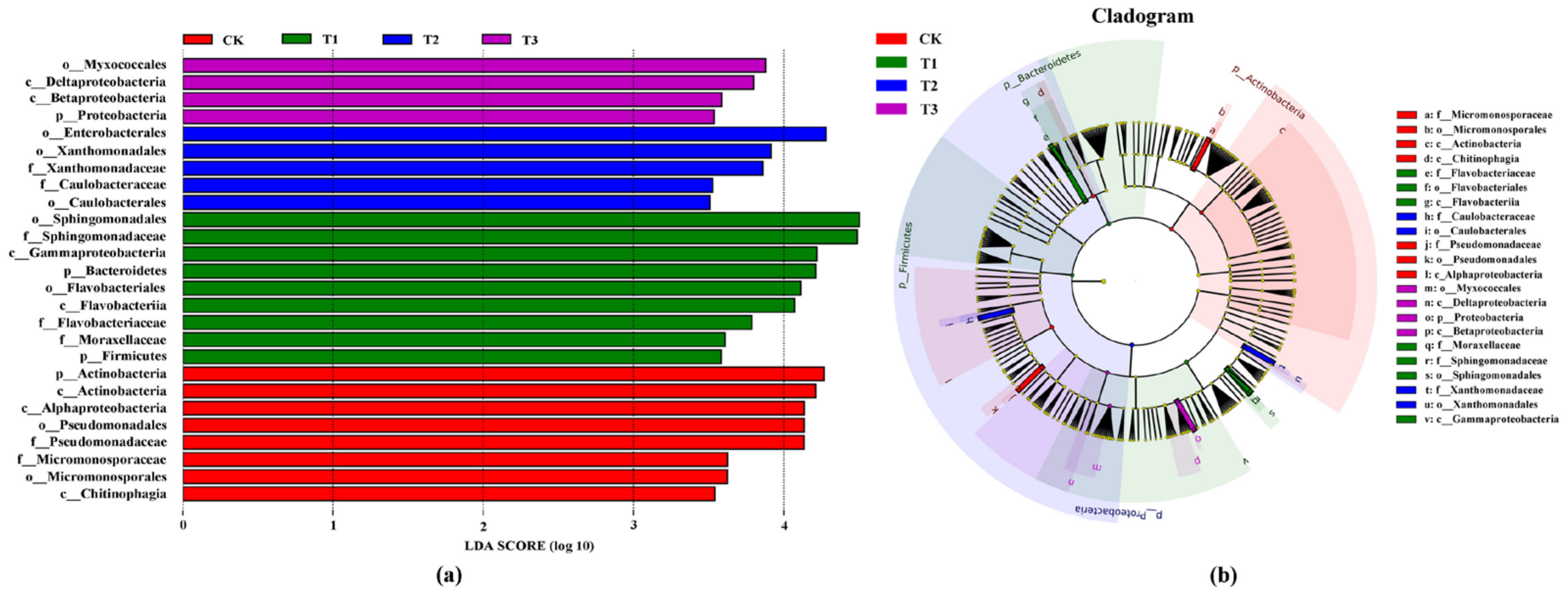
3.4.2. LEfSe Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- FAO. FAO Statistical Databases FAOSTAT. Available online: http://www.fao.org/ (accessed on 27 December 2023).
- Han, J.; Dong, S.; Shi, Y.; Miao, H.; Liu, X.; Beckles, D.M.; Gu, X.; Zhang, S. Fine mapping and candidate gene analysis of gummy stem blight resistance in cucumber stem. Theor. Appl. Genet. 2022, 135, 3117–3125. [Google Scholar] [CrossRef]
- Kong, Z.; Zhang, Y.; Zhuang, C.; Mao, C.; Zhang, C. Detection and characterization of difenoconazole resistance in Stagonosporopsis citrulli from watermelon and muskmelon in Zhejiang Province of China. Phytopathol. Res. 2023, 5, 1. [Google Scholar] [CrossRef]
- Seblani, R.; Keinath, A.P.; Munkvold, G. Gummy stem blight: One disease, three pathogens. Mol. Plant Pathol. 2023, 24, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Mandal, M.K.; Suren, H.; Kousik, C. Elucidation of resistance signaling and identification of powdery mildew resistant mapping loci (ClaPMR2) during watermelon-Podosphaera xanthii interaction using RNA-Seq and whole-genome resequencing approach. Sci. Rep. 2020, 10, 14038. [Google Scholar] [CrossRef] [PubMed]
- McRae, A.G.; Taneja, J.; Yee, K.; Shi, X.; Haridas, S.; LaButti, K.; Singan, V.; Grigoriev, I.V.; Wildermuth, M.C. Spray-induced gene silencing to identify powdery mildew gene targets and processes for powdery mildew control. Mol. Plant Pathol. 2023, 24, 1168–1183. [Google Scholar] [CrossRef] [PubMed]
- Anthony, P.K.; Virginia, B.D. Controlling powdery mildew on cucurbit rootstock seedlings in the greenhouse with fungicides and biofungicides. Crop Prot. 2012, 42, 338–344. [Google Scholar] [CrossRef]
- Price, C.L.; Parker, J.E.; Warrilow, A.G.S.; Kelly, D.E.; Kelly, S.L. Azole fungicides—Understanding resistance mechanisms in agricultural fungal pathogens. Pest Manag. Sci. 2015, 71, 1054–1058. [Google Scholar] [CrossRef]
- Guo, S.; Tao, C.; Jousset, A.; Xiong, W.; Wang, Z.; Shen, Z.; Wang, B.; Xu, Z.; Gao, Z.; Liu, S.; et al. Trophic interactions between predatory protists and pathogen-suppressive bacteria impact plant health. ISME J. 2022, 16, 1932–1943. [Google Scholar] [CrossRef]
- Patrick, B.; Paul, E.; Gerardina, U. Defense-related gene expression in response to the application of biological control agents in banana. Biol. Control 2023, 186, 105317. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; Hamss, H.E.; Belabess, Z.; Barka, E.A. Biological control of plant pathogens: A global perspective. Microorganisms 2022, 10, 596. [Google Scholar] [CrossRef]
- Li, Y.; Gu, Y.; Li, J.; Xu, M.; Wei, Q.; Wang, Y. Biocontrol agent Bacillus amyloliquefaciens LJ02 induces systemic resistance against cucurbits powdery mildew. Front. Microbiol. 2015, 6, 883. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Kim, S.J.; Kwon, S.W.; Rooney, A.P. Phylogenomic analysis shows that Bacillus amyloliquefaciens subsp. plantarum is a later heterotypic synonym of Bacillus methylotrophicus. Int. J. Syst. Evol. Microbiol. 2015, 65, 2104–2109. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, C.A.; Kim, S.J.; Kwon, S.W.; Rooney, A.P. Bacillus velezensis is not a later heterotypic synonym of Bacillus amyloliquefaciens; Bacillus methylotrophicus, Bacillus amyloliquefaciens subsp. plantarum and ‘Bacillus oryzicola’ are later heterotypic synonyms of Bacillus velezensis based on phylogenomics. Int. J. Syst. Evol. Microbiol. 2016, 66, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Nga, N.T.T.; Giau, N.T.; Long, N.T.; Lübeck, M.; Shetty, N.P.; Neergaard, E.D.; Thuy, T.T.T.; Kim, P.V.; Jørgensen, H.J.L. Rhizobacterially induced protection of watermelon against Didymella bryoniae. J. Appl. Microbiol. 2010, 109, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Meysam, S.; Abbas, J.; Jamalali, O. Chemical and biological products for control of powdery mildew on cucumber. Int. J. Veg. Sci. 2022, 28, 233–238. [Google Scholar] [CrossRef]
- González, V.; Armijos, E.; Garcés-Claver, A. Fungal endophytes as biocontrol agents against the main soil-borne diseases of melon and watermelon in Spain. Agronomy 2020, 10, 820. [Google Scholar] [CrossRef]
- Ruangwong, O.; Wonglom, P.; Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.; Sunpapao, A. Biological control activity of Trichoderma asperelloides PSU-P1 against gummy stem blight in muskmelon (Cucumis melo). Physiol. Mol. Plant Pathol. 2021, 115, 101663. [Google Scholar] [CrossRef]
- Zhang, J.; He, L.; Guo, C.; Liu, Z.; Kaliaperumal, K.; Zhong, B.; Jiang, Y. Evaluation of Aspergillus aculeatus GC-09 for the biological control of citrus blue mold caused by Penicillium italicum. Fungal Biol. 2022, 126, 201–212. [Google Scholar] [CrossRef]
- Boshra, A.H.; Rashid, A.A.; Abdullah, M.A. Talaromyces omanensis and Aspergillus fumigatus endophytic fungi suppress Pythium aphanidermatum and its induced damping-off diseases of cucumber and radish. Arch. Phytopathol. Plant Prot. 2023, 56, 665–685. [Google Scholar] [CrossRef]
- Harveson, R.M.; Kimbrough, J.W.; Hopkins, D.L. Novel use of a Pyrenomycetous Mycoparasite for management of fusarium wilt of watermelon. Plant Dis. 2002, 86, 1025–1030. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Zhuang, L.; Yu, Y.; Liu, J.; Zhang, L.; Gao, Z.; Wu, Y.; Gao, W.; Ding, G.; et al. A Rhizosphere-derived consortium of Bacillus subtilis and Trichoderma harzianum suppresses common scab of potato and increases yield. Comput. Struct. Biotechnol. J. 2019, 17, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Ünlü, E.; Çalış, Ö.; Say, A.; Karim, A.A.; Yetisir, H.; Yilmaz, S. Investigation of the effect of Bacillus subtilis and Bacillus thuringiensis as Bio-agents against powdery mildew (Podosphaera xanthii) disease in zucchini (Cucurbita pepo L.). Microb. Pathog. 2023, 185, 106430. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, H.; Behboudi, K.; Ahmadzadeh, M.; Javan-Nikkhah, M.; Zamioudis, C.; Pieterse, C.M.J.; Bakker, P.A. Induced systemic resistance in cucumber and Arabidopsis thaliana by the combination of Trichoderma harzianum Tr6 and Pseudomonas sp. Ps14. Biol. Control 2013, 65, 14–23. [Google Scholar] [CrossRef]
- Ma, Q.; Cong, Y.; Feng, L.; Liu, C.; Yang, W.; Xin, Y.; Chen, K. Effects of mixed culture fermentation of Bacillus amyloliquefaciens and Trichoderma longibrachiatum on its constituent strains and the biocontrol of tomato Fusarium wilt. J. Appl. Microbiol. 2022, 132, 532–546. [Google Scholar] [CrossRef] [PubMed]
- Staropoli, A.; Vassetti, A.; Salvatore, M.M.; Andolfi, A.; Prigigallo, M.I.; Bubici, G.; Scagliola, M.; Salerno, P.; Vinale, F. Improvement of Nutraceutical Value of Parsley Leaves (Petroselinum crispum) upon Field Applications of Beneficial Microorganisms. Horticulturae 2021, 7, 281. [Google Scholar] [CrossRef]
- Zhan, Y.; Wang, Q.; Chen, C.; Kim, J.; Zhang, H.; Yoza, B.; Li, Q. Potential of wheat bran to promote indigenous microbial enhanced oil recovery. J. Microbiol. Biotechnol. 2017, 44, 845–855. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Ke, Y.; Zhu, H.; Li, B.; Li, B.; Zhang, F.; Li, Y. Oleozon: A novel control strategy against powdery mildew in cucumber. J. Phytopathol. 2017, 165, 841–847. [Google Scholar] [CrossRef]
- Yang, J.; Zhu, B.; He, Y.; Ying, Q.S.; Zhang, L.S. Study on mechanical properties and fruit quality analysis of watermelon (Cirtullus lanatus). Acta Agric. Zhejiangensis 2017, 29, 1581–1588. [Google Scholar] [CrossRef]
- Sdiri, S.; Bermejo, A.; Aleza, P.; Navarro, P.; Salvador, A. Phenolic composition, organic acids, sugars, vitamin c and antioxidant activity in the juice of two new triploid late-season mandarins. Food Res. Int. 2012, 49, 462–468. [Google Scholar] [CrossRef]
- Tian, L.; Zhang, W.; Zhou, G.; Li, S.; Wang, Y.; Yang, B.; Bai, T.; Fan, H.; He, P.; Zheng, S. A biological product of Bacillus amyloliquefaciens QST713 strain for promoting banana plant growth and modifying rhizosphere soil microbial diversity and community composition. Front. Microbiol. 2023, 14, 1216018. [Google Scholar] [CrossRef]
- Hu, Y.; Li, Y.; Yang, X.; Li, C.; Wang, L.; Feng, J.; Chen, S.; Li, X.; Yang, Y. Effects of integrated biocontrol on bacterial wilt and rhizosphere bacterial community of tobacco. Sci. Rep. 2021, 11, 2653. [Google Scholar] [CrossRef] [PubMed]
- Bjørnsgaard, A.A.; Davey, M.L.; Kauserud, H. ITS all right mama: Investigating the formation of chimeric sequences in the ITS2 region by DNA metabarcoding analyses of fungal mock communities of different complexities. Mol. Ecol. Resour. 2017, 17, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Paruch, L.; Paruch, M.A.; Eiken, G.H.; Sørheim, R. Aquatic microbial diversity associated with faecal pollution of Norwegian waterbodies characterized by 16S rRNA gene amplicon deep sequencing. Microb. Biotechnol. 2019, 12, 1487–1491. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sun, Z.; Sang, Q.; Qin, C.; Kong, L.; Huang, X.; Liu, H.; Su, T.; Li, H.; He, M.; et al. Soybean reduced internode 1 determines internode length and improves grain yield at dense planting. Nat. Commun. 2023, 14, 7939. [Google Scholar] [CrossRef] [PubMed]
- Santiago, C.D.; Yagi, S.; Ijima, M.; Nashimoto, T.; Sawada, M.; Ikeda, S.; Asano, K.; Orikasa, Y.; Ohwada, T. Bacterial compatibility in combined inoculations enhances the growth of potato seedlings. Microbes Environ. 2017, 32, 14–23. [Google Scholar] [CrossRef]
- Izquierdo-García, L.F.; González-Almario, A.; Cotes, A.M.; Moreno-Velandia, C.A. Trichoderma virens Gl006 and Bacillus velezensis Bs006: A compatible interaction controlling Fusarium wilt of cape gooseberry. Sci. Rep. 2020, 10, 6857. [Google Scholar] [CrossRef]
- Zaim, S.; Bekkar, A.A.; Belabid, L. Efficacy of Bacillus subtilis and Trichoderma harzianum combination on chickpea Fusarium wilt caused by F. oxysporum f. sp. ciceris. Arch. Phytopathol. Plant Prot. 2018, 51, 217–226. [Google Scholar] [CrossRef]
- Maslennikova, V.S.; Tsvetkova, V.P.; Shelikhova, E.V.; Selyuk, M.P.; Alikina, T.Y.; Kabilov, M.R.; Dubovskiy, I.M. Bacillus subtilis and Bacillus amyloliquefaciens mix suppresses rhizoctonia disease and improves rhizosphere microbiome, growth and yield of potato (Solanum tuberosum L.). J. Fungi 2023, 9, 1142. [Google Scholar] [CrossRef]
- Marian, M.; Morita, A.; Koyama, H.; Suga, H.; Shimizu, M. Enhanced biocontrol of tomato bacteria wilt using the combined application of Mitsuaria sp. TWR114 and nonpathogenic Ralstonia sp. TCR112. J. Gen. Plant Pathol. 2019, 85, 142–154. [Google Scholar] [CrossRef]
- Lan, J.; Huang, J.; Huang, S.; Li, J.; Tian, K.; Liu, Q.; Wu, J.; Zheng, L. A preliminary study on the effects of combination of biocontrol agents against watermelon fusarium wilt caused by Fusarium oxysporum f. sp. Niveum. Guangdong Agric. Sci. 2022, 49, 94–104. [Google Scholar] [CrossRef]
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Curr. Res. Microb. Sci. 2021, 2, 100054. [Google Scholar] [CrossRef] [PubMed]
- Brussaard, L.; Ruiter, P.C.; Brown, G.G. Soil biodiversity for agricultural sustainability. Agric. Ecosyst. Environ. 2007, 121, 233–244. [Google Scholar] [CrossRef]
- Naumova, N.; Baturina, O.; Nechaeva, T.; Kabilov, M. Root and Rhizosphere Microbiome of Tomato Plants Grown in the Open Field in the South of West Siberia under Mineral Fertilization. Horticulturae 2022, 8, 1051. [Google Scholar] [CrossRef]
- Kruker, G.; Guidi, E.S.; Santos, J.M.d.S.d.; Mafra, Á.L.; Almeida, J.A.d. Quality of Bokashi-Type Biofertilizer Formulations and Its Application in the Production of Vegetables in an Ecological System. Horticulturae 2023, 9, 1314. [Google Scholar] [CrossRef]
- Yang, X.; Lai, J.; Zhang, Y.; Luo, X.; Han, M.; Zhao, S. Microbial community structure and metabolome profiling characteristics of soil contaminated by TNT, RDX, and HMX. Environ. Pollut. 2021, 285, 117478. [Google Scholar] [CrossRef] [PubMed]
- Lanzavecchia, G.; Frascarelli, G.; Rocchetti, L.; Bellucci, E.; Bitocchi, E.; Di Vittori, V.; Sillo, F.; Ferraris, I.; Carta, G.; Delledonne, M.; et al. Genotype Combinations Drive Variability in the Microbiome Configuration of the Rhizosphere of Maize/Bean Intercropping System. Int. J. Mol. Sci. 2024, 25, 1288. [Google Scholar] [CrossRef]
- Zhao, Y.; Sun, C.; Wang, S.; Zhang, M.; Li, Y.; Xue, Q.; Guo, Q.; Lai, H. Widely targeted metabolomic, transcriptomic, and metagenomic profiling reveal microbe-plant-metabolic reprogramming patterns mediated by Streptomyces pactum Act12 enhance the fruit quality of Capsicum annuum L. Food Res. Int. 2023, 166, 112587. [Google Scholar] [CrossRef]
- Li, Q.; You, P.; Hu, Q.; Leng, B.; Wang, J.; Chen, J.; Wan, S.; Wang, B.; Yuan, C.; Zhou, R.; et al. Effects of co-contamination of heavy metals and total petroleum hydrocarbons on soil bacterial community and function network reconstitution. Ecotoxicol. Environ. Saf. 2020, 204, 111083. [Google Scholar] [CrossRef]
- Mendes, L.; Kuramae, E.; Navarrete, A.; Veen, J.; Tsai, S. Taxonomical and functional microbial community selection in soybean rhizosphere. ISME J. 2014, 8, 1577–1587. [Google Scholar] [CrossRef] [PubMed]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Li, G.; Peng, Y.; Wang, A.; Zhu, J. Mixed bacterial fermentation can control the growth and development of Verticillium dahliae. Biotechnol. Biotechnol. Equip. 2020, 34, 58–69. [Google Scholar] [CrossRef]
- Wang, C.; Huang, Y.; Zhang, C.; Zhang, Y.; Yuan, K.; Xue, W.; Liu, Y.; Liu, Y.; Liu, Z. Inhibition effects of long-term calcium-magnesia phosphate fertilizer application Cd uptake in rice: Regulation of the iron-nitrogen coupling cycle driven by the soil microbial community. J. Hazard. Mater. 2021, 416, 125916. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Khurana, H.; Singh, D.N.; Negi, R.K. The genus Sphingopyxis: Systematics, ecology, and bioremediation potential—A review. J. Environ. Manag. 2021, 280, 111744. [Google Scholar] [CrossRef] [PubMed]
- Verastegui, Y.; Cheng, J.; Engel, K.; Kolczynski, D.; Mortimer, S.; Lavigne, J.; Montalibet, J.; Romantsov, T.; Hall, M.; McConkey, B.J.; et al. Multisubstrate isotope labeling and metagenomic analysis of active soil bacterial communities. mBio 2014, 5, e01157-14. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yoshida, S.; Mitani, N.; Egusa, M.; Takagi, M.; Izawa, H.; Matsumoto, T.; Kaminaka, H.; Ifuku, S. Disease resistance and growth promotion activities of chitin/cellulose nanofiber from spent mushroom substrate to plant. Carbohydr. Polym. 2022, 284, 119233. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Yang, S.; Chen, S.; Zhao, S.; Ai, Y.; Huang, D.; Yang, K.; Cheng, H. Soil microbial responses to simultaneous contamination of antimony and arsenic in the surrounding area of an abandoned antimony smelter in Southwest China. Environ. Int. 2023, 174, 107897. [Google Scholar] [CrossRef] [PubMed]
- Emenike, C.U.; Agamuthu, P.; Fauziah, S.H.; Omo-Okoro, P.N.; Jayanthi, B. Enhanced bioremediation of metal-contaminated soil by consortia of proteobacteria. Water Air Soil Pollut. 2023, 234, 731. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Bacillus velezensis LJ02 | Aspergillus aculeatus 9 | Aspergillus fumigatus F14 |
|---|---|---|---|
| CK | 0 | 0 | 0 |
| T1 | 2 L (1 × 107 CFU/mL) | 0 | 0 |
| T2 | 2 L (1 × 107 CFU/mL) | 2 L (1 × 107 conidia/mL) | 0 |
| T3 | 2 L (1 × 107 CFU/mL) | 0 | 2 L (1 × 107 conidia/mL) |
| Treatment | Shannon Index | Simpson Index | Chao1 Index |
|---|---|---|---|
| CK | 8.95918 | 0.967782 | 24,631.76 |
| T1 | 9.116933 | 0.971445 | 24,568.47 |
| T2 | 9.067654 | 0.970409 | 24,536.91 |
| T3 | 8.993019 | 0.968248 | 24,553.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, W.; Wu, T.; Chang, R.; Yuan, Y.; Wang, Y. The Effect of Bacillus velezensis LJ02 Compounded with Different Fungi on the Growth of Watermelon Seedlings and Microbial Community Structure. Horticulturae 2024, 10, 236. https://doi.org/10.3390/horticulturae10030236
Yu W, Wu T, Chang R, Yuan Y, Wang Y. The Effect of Bacillus velezensis LJ02 Compounded with Different Fungi on the Growth of Watermelon Seedlings and Microbial Community Structure. Horticulturae. 2024; 10(3):236. https://doi.org/10.3390/horticulturae10030236
Chicago/Turabian StyleYu, Weiwei, Tianyi Wu, Ruokui Chang, Yujin Yuan, and Yuanhong Wang. 2024. "The Effect of Bacillus velezensis LJ02 Compounded with Different Fungi on the Growth of Watermelon Seedlings and Microbial Community Structure" Horticulturae 10, no. 3: 236. https://doi.org/10.3390/horticulturae10030236