
LbHSP17.9 Participated in the Regulation of Cold Stress in Cut Lily Flowers

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Treatment
2.2. Extraction and Reverse Transcription of mRNA
2.3. Reverse Transcription RT-PCR
2.4. Real-Time Fluorescence Quantitative PCR (qPCR)
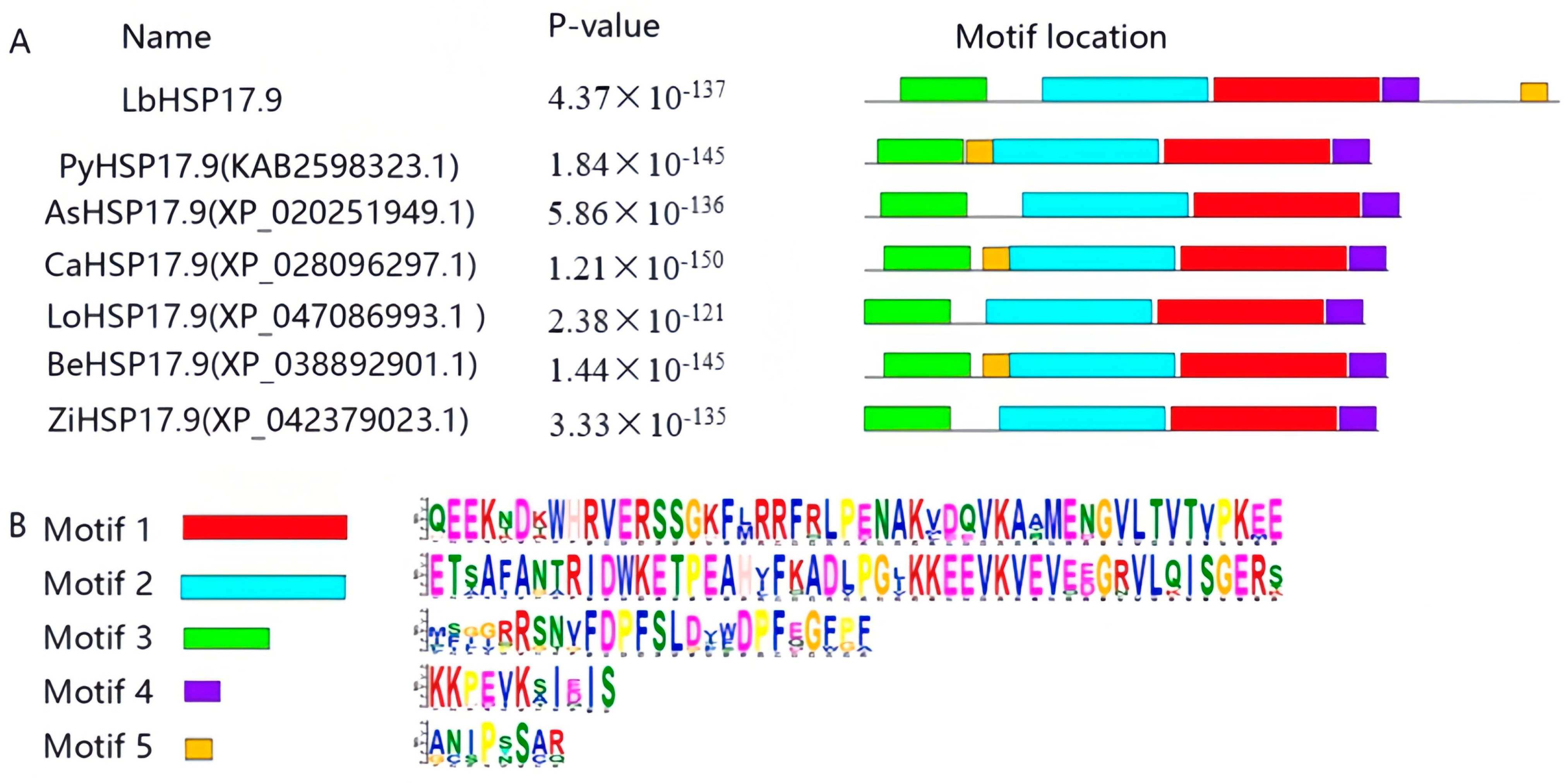
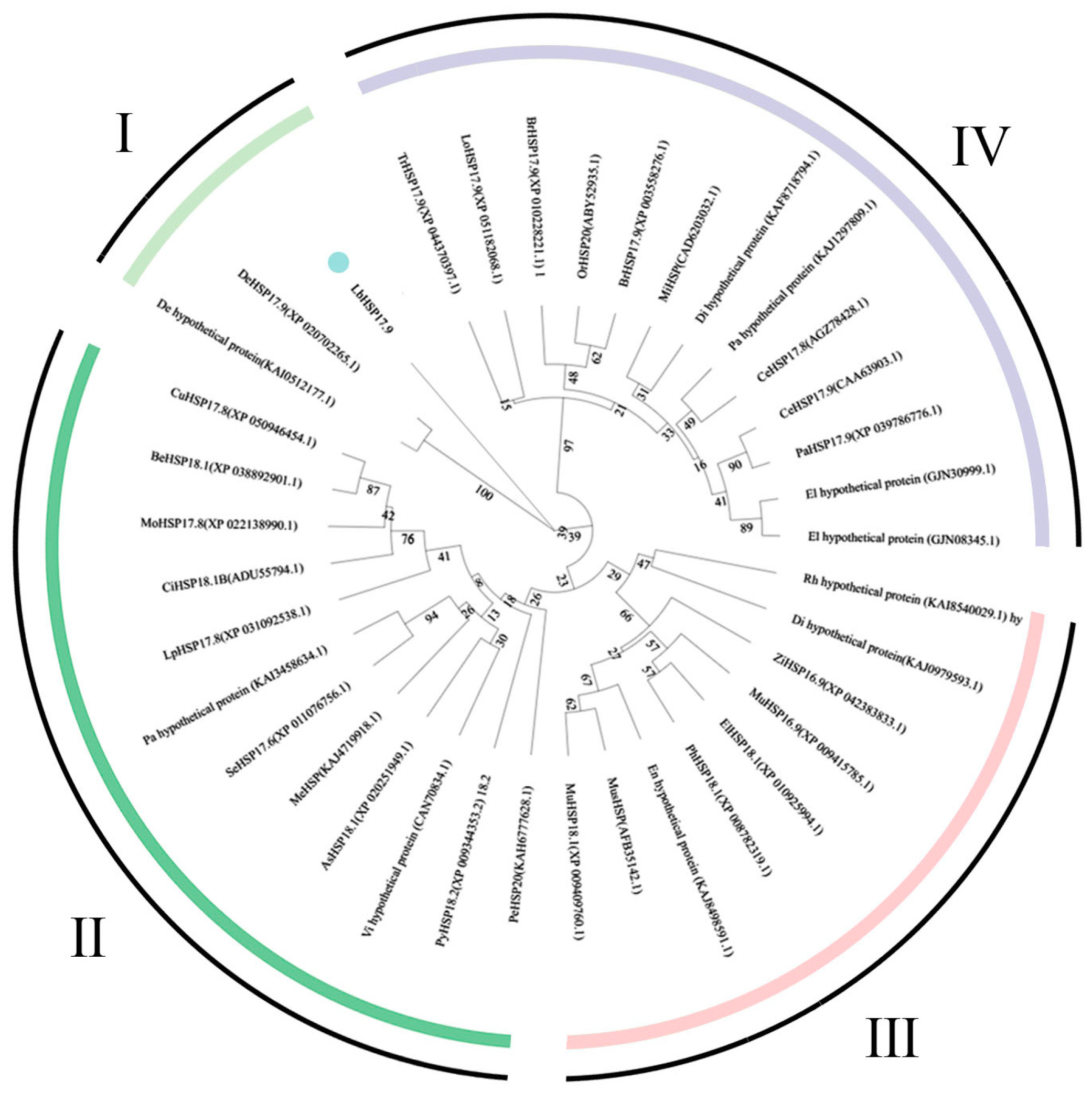
2.5. Bioinformatics Analysis of LbHSP17.9
2.6. Construction of Silencing Vector and Virus-Induced Gene Silencing (VIGS) Treatment
2.7. Construction of Overexpression Vector
2.8. Transient Overexpression of LbHSP17.9 in Lily Petal Disks
2.9. Acquisition of OE-LbHSP17.9 Transgenic A. thaliana
2.10. Cold Stress Treatment of A. thaliana
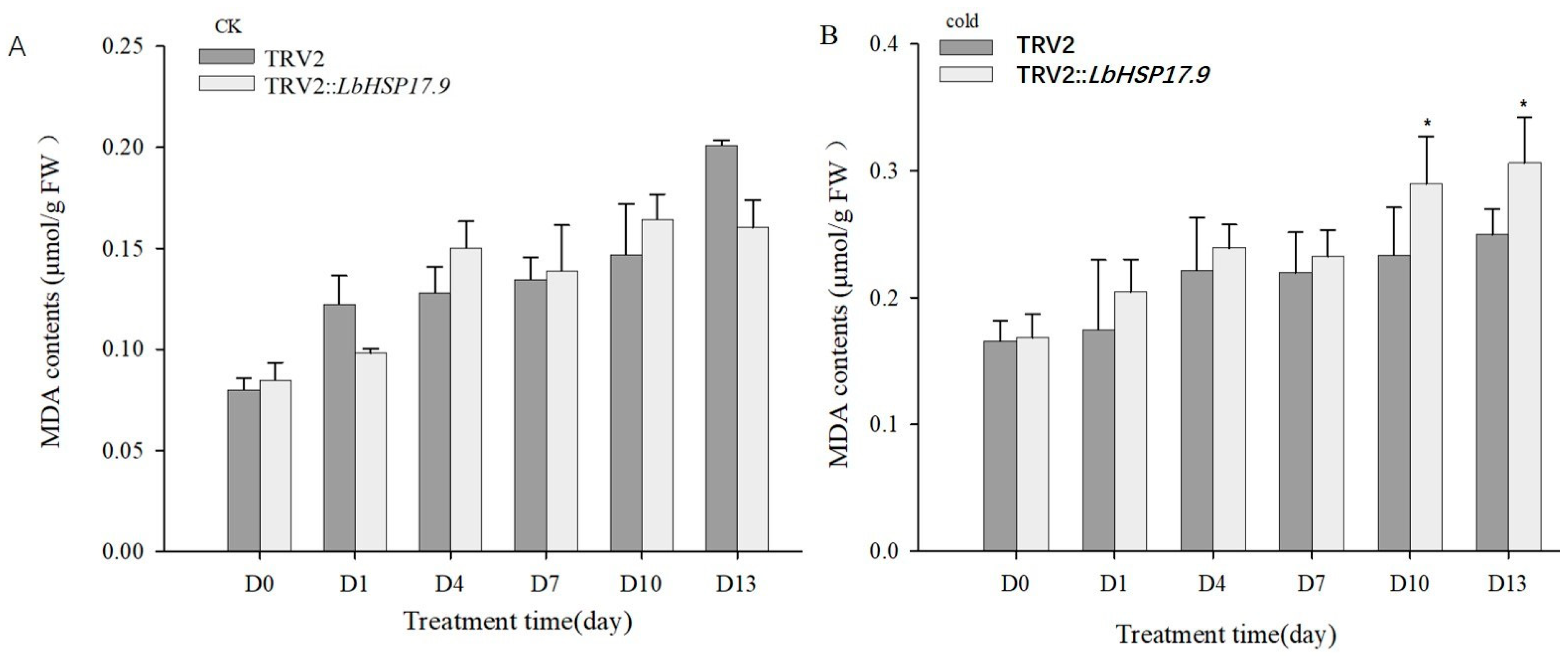
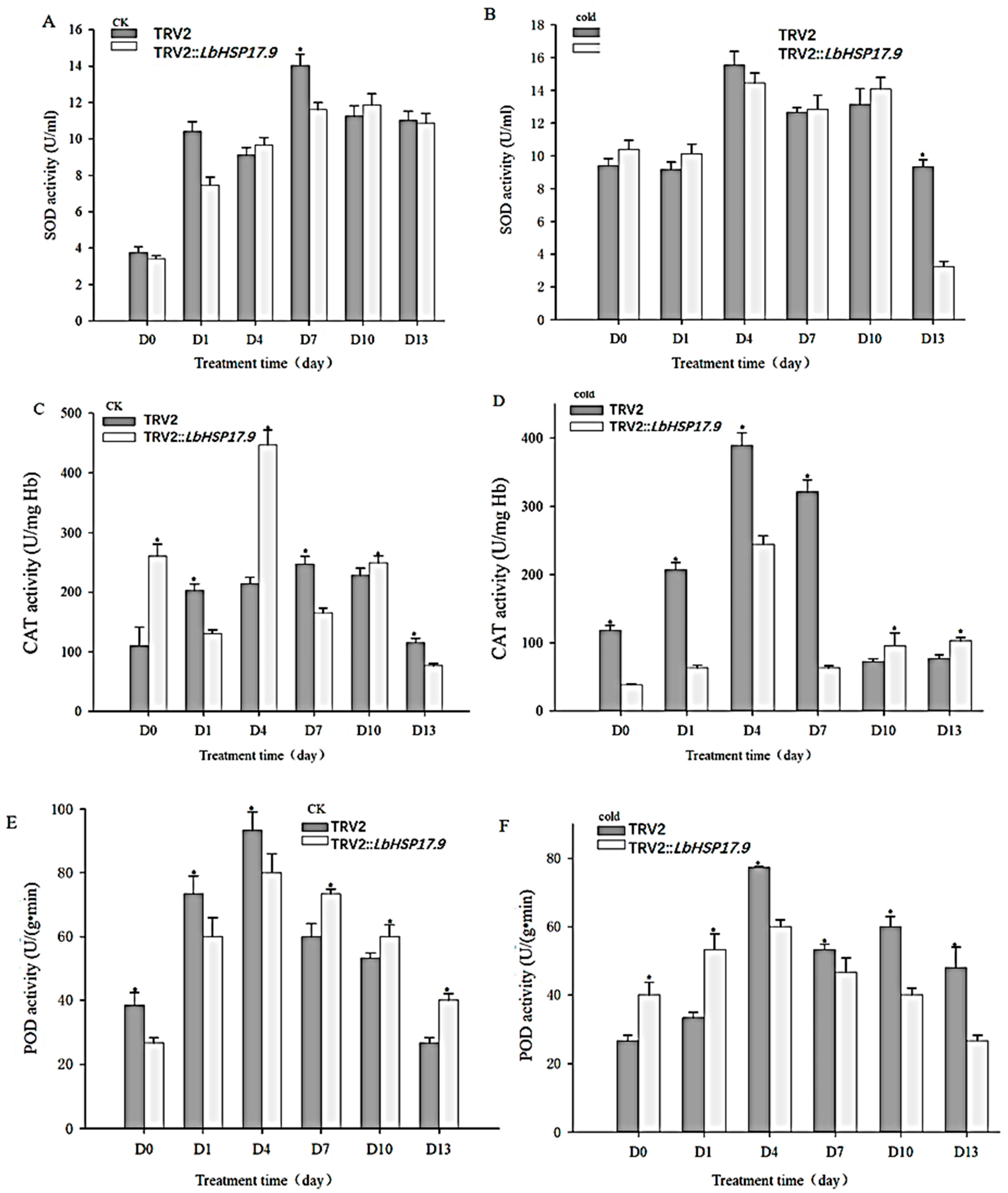
2.11. Determination of MDA Content and SOD, CAT and POD Activities
2.12. Determination of Relative Electrical Conductivity
2.13. Data Analysis
3. Results and Analysis
3.1. Cloning and Bio-Information Analysis of LbHSP17.9
3.2. Different Treatments Increased Expression Level of LbHSP17.9 in Cut Lily Flowers
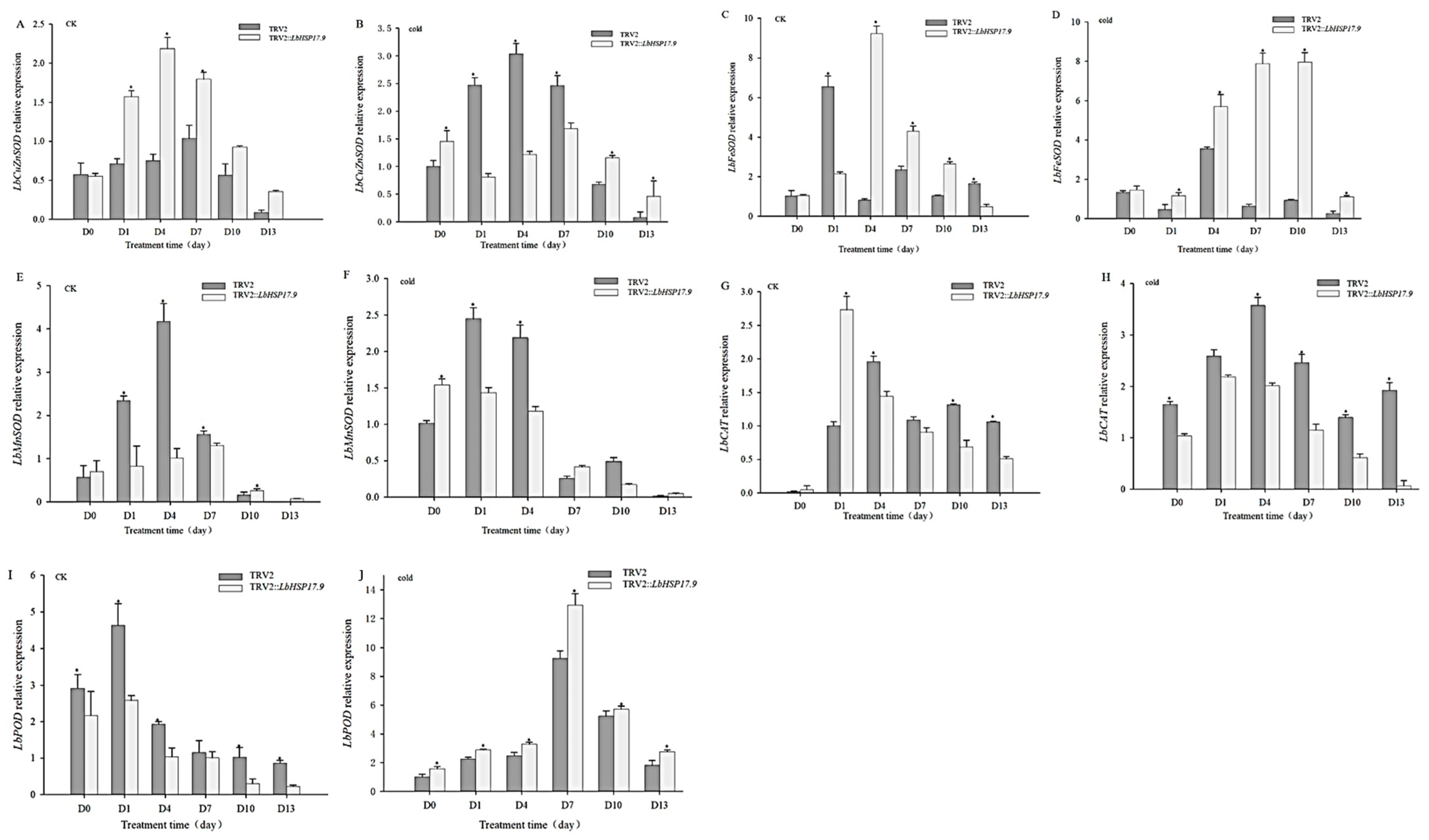
3.3. Silencing LbHSP17.9 Weakened the Cold Stress Resistance, Down-Regulated LbHSP17.9 Expression and Promoted MDA Content
3.4. Silencing LbHSP17.9 Lowered Activities and Transcription of Antioxidant Enzymes
3.5. LbHSP17.9 Overexpression Enhances Resistance to Cold Stress of Lily Petal Segments and Enhances Resistance to Low-Temperature Stress
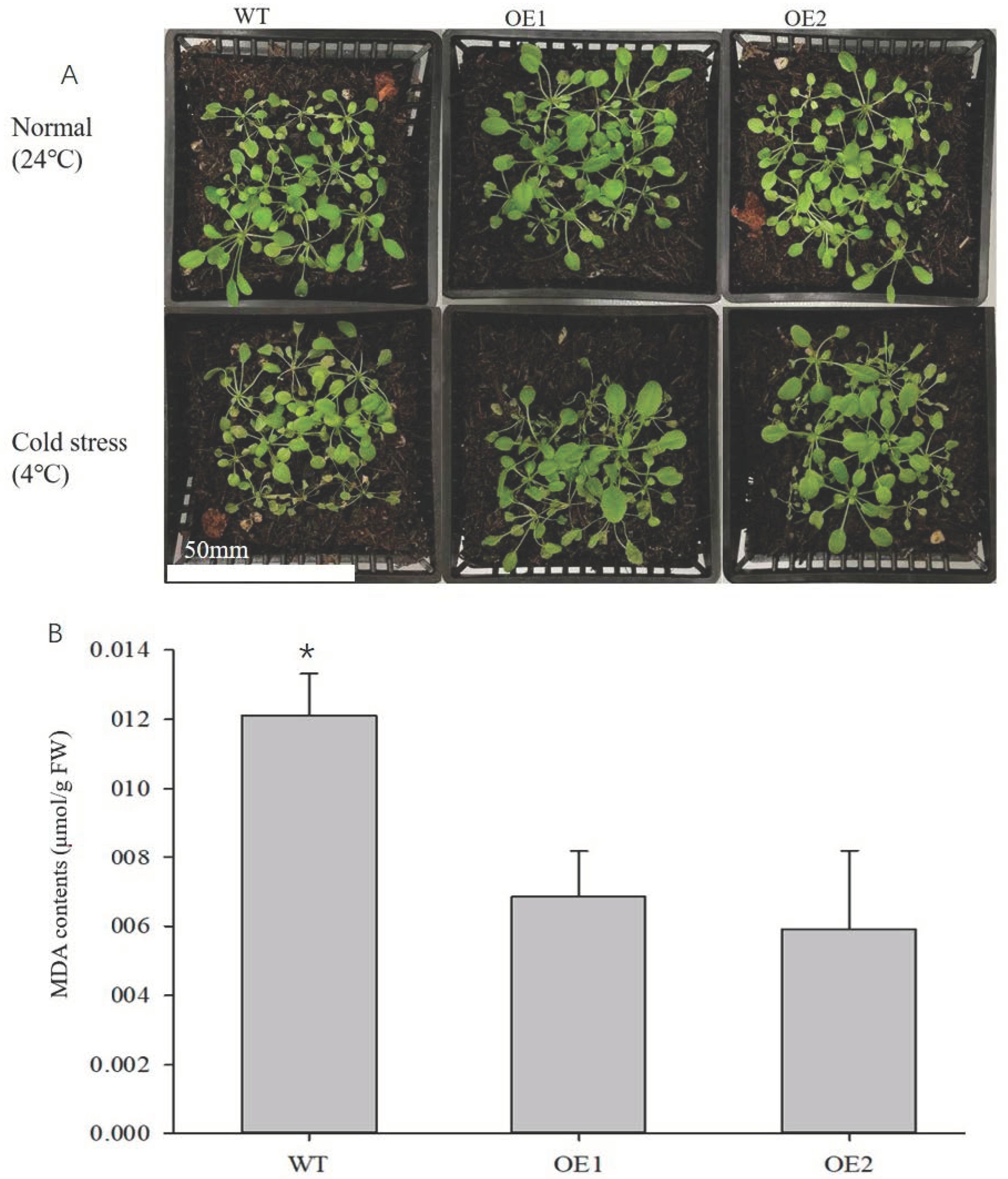
3.6. Overexpression of LbHSP17.9 in A. thaliana Enhances Resistance to Low-Temperature Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhao, Z.; Zhao, J.; Peng, C.; Duan, X.; Deng, M.; Wen, J. Comparative analysis of petal epidermal wax composition and loss-water resistance in five cut lily cultivars (lilium spp). Sci. Hortic. 2023, 311, 11805. [Google Scholar] [CrossRef]
- Yu, T.; Zhou, H.; Liu, Z.; Zhai, H.; Liu, Q. The sweet potato transcription factor IbbHLH33 enhances chilling tolerance in transgenic tobacco. Czech J. Genet. Plant Breed. 2022, 58, 210–222. [Google Scholar] [CrossRef]
- Lopez-Matas, M.A.; Nuez, P.; Soto, A.; Allona, I.; Gomez, L. Protein cryoprotective activity of a cytosolic small heat shock protein that accumulates constitutively in chestnut stems and is up-regulated by low and high temperatures. Plant Physiol. 2004, 134, 1708–1717. [Google Scholar] [CrossRef]
- Kalousek, P.; Holátko, J.; Schreiber, P.; Pluháček, T.; Lónová1, K.; Radziemska, M.; Tarkowski, P.; Vyhnánek, T.; Hammerschmiedt, T.; Brtnický, M. The effect of chelating agents on the Zn-phytoextraction potential of hemp and soil microbial activity. Chem. Biol. Technol. Agric. 2024, 11, 23. [Google Scholar] [CrossRef]
- Park, C.J.; Seo, Y.S. Heat shock proteins: A review of the molecular chaperones for plant immunity. Plant Pathol. 2015, 31, 323–333. [Google Scholar] [CrossRef]
- Candido, E.P.M.; Grber, S.; Gaestel, M.; Buchner, J. Binding of non-native protein to hsp25 during heat shock creates a reservoir of folding intermediates for reactivation. Chemtracts 1998, 11, 166–172. [Google Scholar]
- Mogk, A.; Schlieker, C.; Friedrich, K.L.; Schnfeld, H.J.; Bukau, B. Refolding of substrates bound to small hsps relies on a disaggregation reaction mediated most efficiently by clpb/dnak. J. Biol.Chem. 2003, 278, 31033–31042. [Google Scholar] [CrossRef] [PubMed]
- Waters, E.R.; Lee, G.J.; Vierling, E. Evolution, structure and function of the small heat shock proteins in plants. J. Exp. Bot. 1996, 47, 325–338. [Google Scholar] [CrossRef]
- Zhang, J.H.; Wang, L.J.; Pan, Q.H.; Wang, Y.Z.; Zhan, J.C.; Huang, W.D. Accumulation and subcellular localization of heat shock proteins in young grape leaves during cross-adaptation to temperature stresses. Sci. Hortic. 2008, 117, 231–240. [Google Scholar] [CrossRef]
- González-Gordo, S.; Palma, J.M.; Corpas, F.J. Small Heat Shock Protein (sHSP) Gene Family from Sweet Pepper (Capsicum annuum L.) Fruits: Involvement in Ripening and Modulation by Nitric Oxide (NO). Plants 2023, 12, 389. [Google Scholar] [CrossRef]
- Zhang, N.; Zhao, H.; Shi, J.; Wu, Y.; Jiang, J. Functional characterization of class i slhsp17.7 gene responsible for tomato cold-stress tolerance. Plant Sci. 2020, 298, 110568. [Google Scholar] [CrossRef]
- Neto, V.G.; Barbosa, R.R.; Carosio, M.G.A.; Ferreira, A.G.; Fernandez, L.G.; De Castro, R.D.; Ligterink, W.; Hilhorst, H.; Ribeiro, P.R. Sequence analysis of ricinus communis small heat-shock protein (shsp) subfamily and its role in abiotic stress responses. Ind. Crop. Prod. 2020, 152, 112541. [Google Scholar] [CrossRef]
- Sewelam, N.; Kazan, K.; Hüdig, M.; Maurino, V.G.; Schenk, P.M. The athsp17.4c1 gene expression is mediated by diverse signals that link biotic and abiotic stress factors with ros and can be a useful molecular marker for oxidative stress. Int. J. Mol. Sci. 2019, 20, 3201. [Google Scholar] [CrossRef]
- Sato, Y.; Yokoya, S. Enhanced tolerance to drought stress in transgenic rice plants overexpressing a small heat-shock protein, shsp17.7. Plant Cell Rep. 2008, 27, 329–334. [Google Scholar] [CrossRef]
- Yin, H.; Chen, L.; Li, X.; Chen, Q.; Yi, M. Analysis and improvement of high-quality RNA extraction in leaves of lily. J. CAU 2008, 13, 41–45. [Google Scholar]
- Liu, L.; Xie, T.; Zhao, Z.; Zhao, J.; Deng, M.; Wen, J. Involvement of a novel nap member lbnap in flower senescence and dehydration stress in cut lily. Sci. Hortic. 2023, 308, 111482. [Google Scholar] [CrossRef]
- Carra, S.; Alberti, S.; Arrigo, P.A.; Benesch, J.L.; Benjamin, I.J.; Boelens, W.; Bartelt-Kirbach, B.; Brundel, B.J.; Buchner, J.; Bukau, B.; et al. The growing world of small heat shock proteins: From structure to functions. Cell Stress. Chaperon. 2017, 22 Pt 13, 601–611. [Google Scholar] [CrossRef]
- Wan, X.; Yang, J.; Li, X.; Zhou, Q.; Guo, C.; Bao, M. Over-expression of pmhsp17.9 in transgenic arabidopsis thaliana confers thermotolerance. Plant Mol. Biol. Rep. 2016, 34, 899–908. [Google Scholar] [CrossRef]
- Basha, E.; Lee, G.J.; Breci, L.A.; Hausrath, A.C.; Buan, N.R.; Giese, K.C.; Vierling, E. The identity of proteins associated with a small heat shock protein during heat stress in vivo indicates that these chaperones protect a wide range of cellular functions. J. Biol. Chem. 2004, 279, 7566–7575. [Google Scholar] [CrossRef]
- Yang, Z.; Du, H.; Xing, X.; Li, W.; Kong, Y.; Li, X.; Zhang, C. A small heat shock protein, GmHSP17. 9, from nodule confers symbiotic nitrogen fixation and seed yield in soybean. Plant biotechnol. J. 2022, 20, 103. [Google Scholar] [CrossRef]
- Zhong, L.; Zhou, W.; Wang, H.; Ding, S.; Lu, Q.; Wen, X.; Peng, L.; Zhang, L.; Lu, C. Chloroplast small heat shock protein HSP21 interacts with plastid nucleoid protein pTAC5 and is essential for chloroplast development in Arabidopsis under heat stress. Plant Cell 2013, 25, 2925–2943. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Shi, J.; Zhao, H.; Jiang, J. Activation of small heat shock protein (SlHSP17. 7) gene by cell wall invertase inhibitor (SlCIF1) gene involved in sugar metabolism in tomato. Gene 2018, 679, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yu, G.; Li, H.; Li, X.; Mu, C. Overexpression of small heat shock protein LimHSP16. 45 in Arabidopsis hsp17. 6II mutant enhances tolerance to abiotic stresses. Russ. J. Plant Physiol. 2020, 67, 231–241. [Google Scholar] [CrossRef]
- Ma, P.; Li, J.; Qi, L.; Dong, X. The archaeal small heat shock protein hsp17. 6 protects proteins from oxidative inactivation. Int. J. Mol. Sci. 2021, 22, 2591. [Google Scholar] [CrossRef]
- Sarkar, N.K.; Kotak, S.; Agarwal, M.; Kim, Y.K.; Grover, A. Silencing of class I small heat shock proteins affects seed-related attributes and thermotolerance in rice seedlings. Planta 2020, 251, 1–16. [Google Scholar] [CrossRef]
- Wu, J.; Gao, T.; Hu, J.; Zhao, L.; Chang, Y.; Ma, F. Research advances in function and regulation mechanisms of plant small heat shock proteins (sHSPs) under environmental stresses. Sci. Total Environ. 2022, 825, 154054. [Google Scholar] [CrossRef]
- Mu, C.; Zhang, S.; Yu, G.; Chen, N.; Li, X.; Liu, H.; Misra, R. Overexpression of small heat shock protein LimHSP16. 45 in Arabidopsis enhances tolerance to abiotic stresses. PLoS ONE 2013, 8, e82264. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, H.; Chu, P.; Li, Y.; Tan, B.; Ding, Y.; Tsang, E.W.T.; Jiang, L.; Wu, K.; Huang, S. NnHsp17.5, a cytosolic class II small heat shock protein gene from Nelumbo nucifera, contributes to seed germination vigor and seedling thermotolerance in transgenic Arabidopsis. Plant Cell Rep. 2012, 31, 379–389. [Google Scholar] [CrossRef]
- Li, J.; Zhang, J.; Jia, H.; Li, Y.; Xu, X.; Wang, L.; Lu, M. The Populus trichocarpa PtHsp17.8 involved in heat and salt stress tolerances. Plant Cell Rep. 2016, 35, 1587–1599. [Google Scholar] [CrossRef] [PubMed]















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Yan, X.; Huang, W.; Liu, C.; Hao, X.; Gao, C.; Deng, M.; Wen, J. LbHSP17.9 Participated in the Regulation of Cold Stress in Cut Lily Flowers. Horticulturae 2024, 10, 287. https://doi.org/10.3390/horticulturae10030287
Zhao J, Yan X, Huang W, Liu C, Hao X, Gao C, Deng M, Wen J. LbHSP17.9 Participated in the Regulation of Cold Stress in Cut Lily Flowers. Horticulturae. 2024; 10(3):287. https://doi.org/10.3390/horticulturae10030287
Chicago/Turabian StyleZhao, Jiahui, Xinyu Yan, Wei Huang, Cheng Liu, Xuan Hao, Chengye Gao, Minghua Deng, and Jinfen Wen. 2024. "LbHSP17.9 Participated in the Regulation of Cold Stress in Cut Lily Flowers" Horticulturae 10, no. 3: 287. https://doi.org/10.3390/horticulturae10030287
APA StyleZhao, J., Yan, X., Huang, W., Liu, C., Hao, X., Gao, C., Deng, M., & Wen, J. (2024). LbHSP17.9 Participated in the Regulation of Cold Stress in Cut Lily Flowers. Horticulturae, 10(3), 287. https://doi.org/10.3390/horticulturae10030287