
Biocontrol Potential of Bacteria Isolated from Vermicompost against Meloidogyne incognita on Tomato and Cucumber Crops
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms Isolated from Vermicompost
2.2. Preparation of Root-Knot Nematode Inoculum
2.3. Screening of Antagonistic Bacteria against Root-Knot Nematode
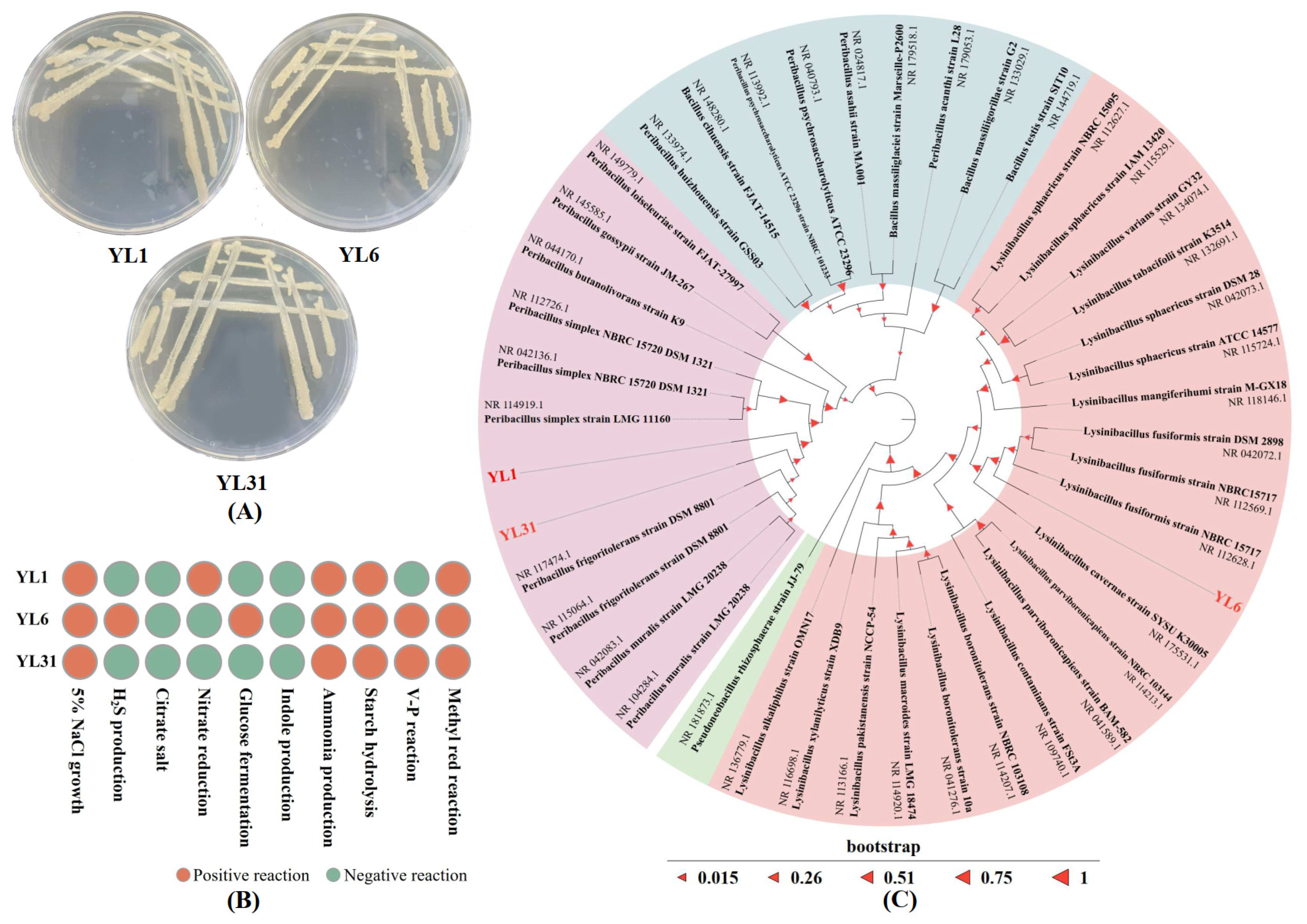
2.4. Identification of Antagonistic Bacteria in Root-Knot Nematode
2.5. Research on the Functions and Growth-Promoting Properties of Antagonistic Bacteria
2.5.1. Detection of the Protease and Chitinase Production Capacity of Antagonistic Bacteria
2.5.2. Phosphorus-Dissolving and Potassium-Dissolving Ability Tests of Antagonistic Bacteria
2.6. Effects of Antagonistic Bacteria on the Development of Root-Knot Nematode Eggs and J2
2.7. Pot Experiment
2.8. Field Tests
2.9. Data Analysis
3. Results
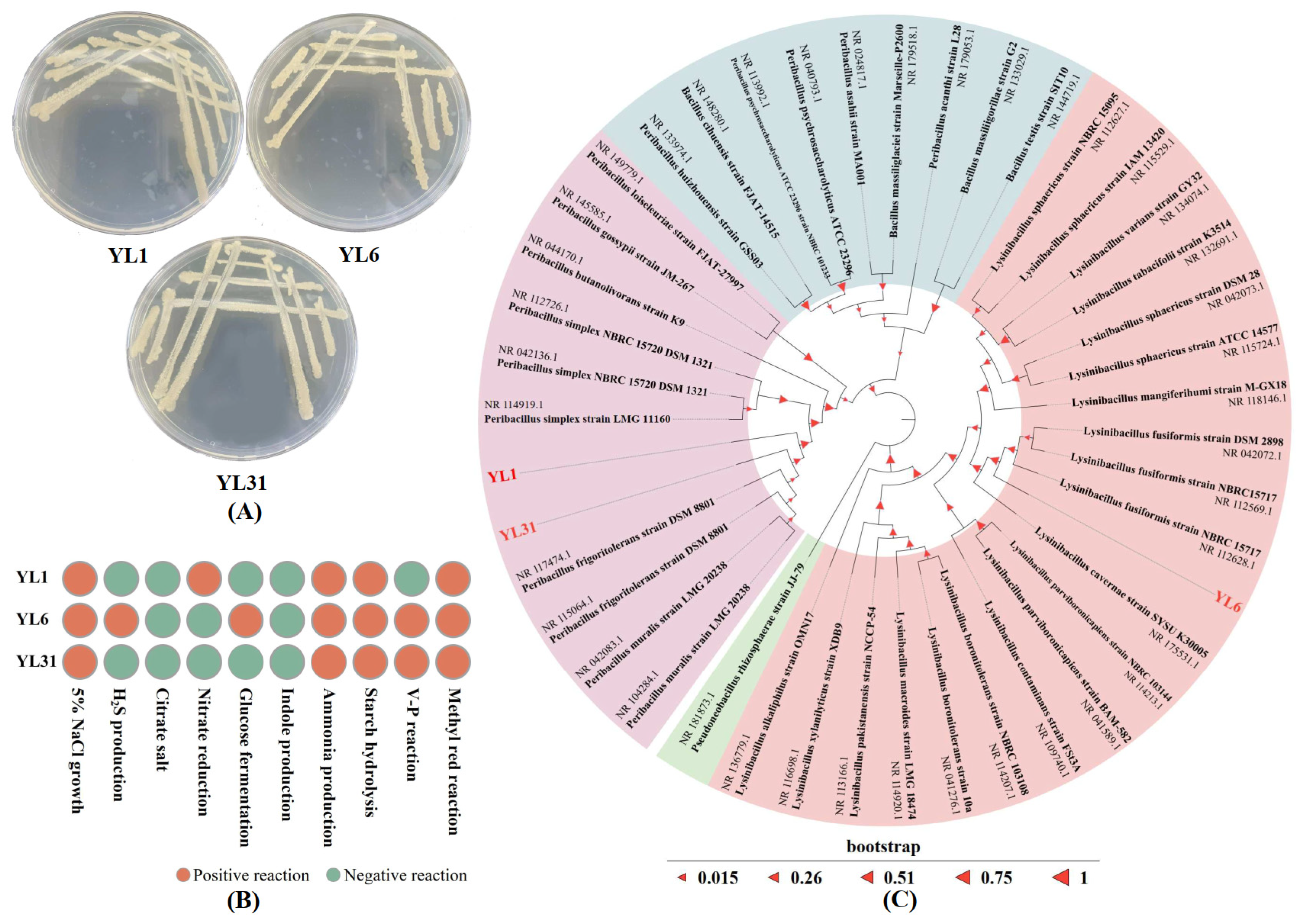
3.1. Isolation and Identification of Root-Knot Nematode Antagonistic Bacteria
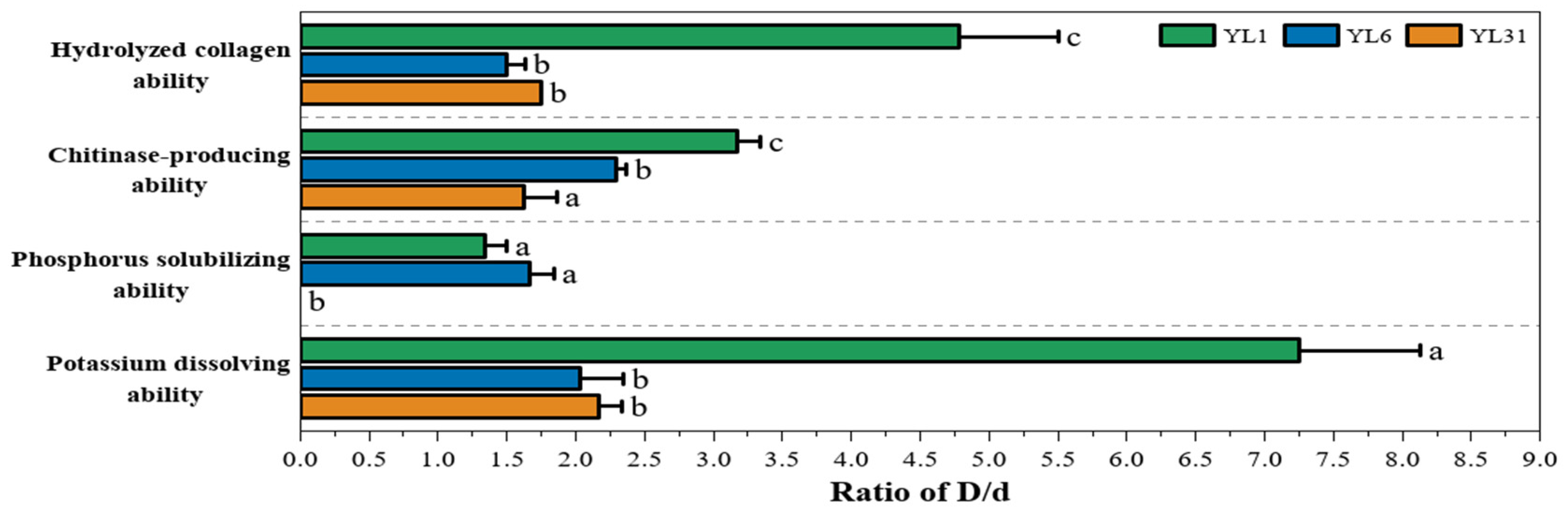
3.2. Functions and Growth-Promoting Properties of Antagonistic Bacteria
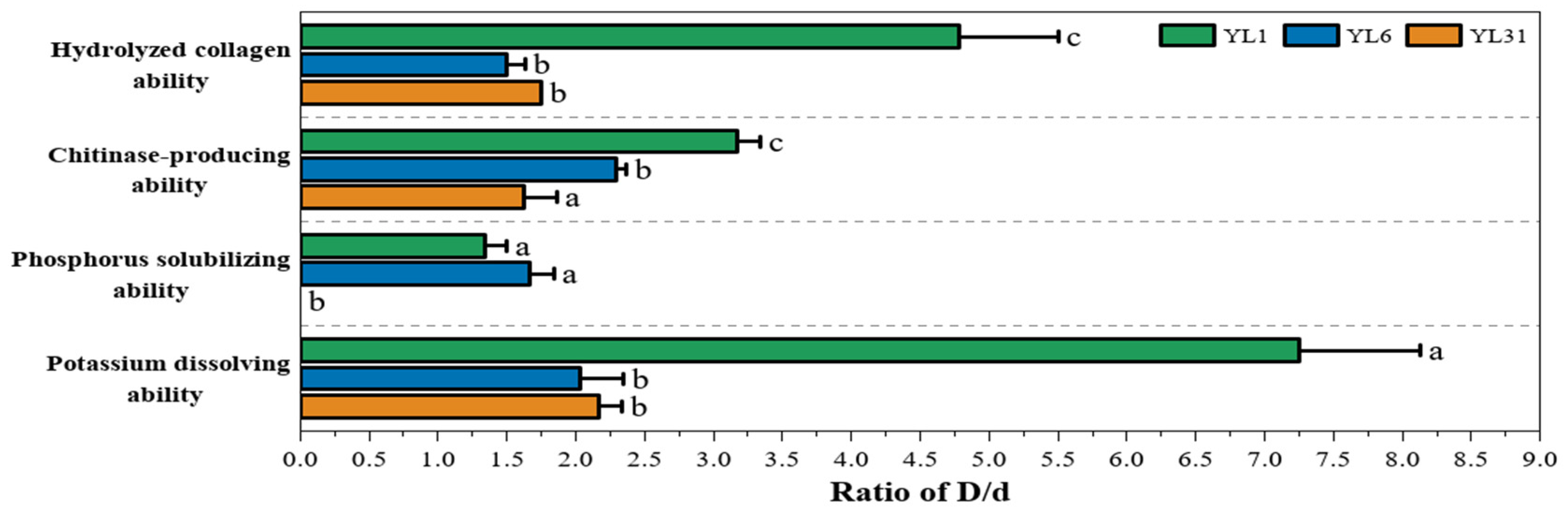
3.2.1. Protease and Chitinase Production Capacity of Antagonistic Bacteria
3.2.2. Phosphorus-Dissolving and Potassium-Dissolving Ability of Antagonistic Bacteria
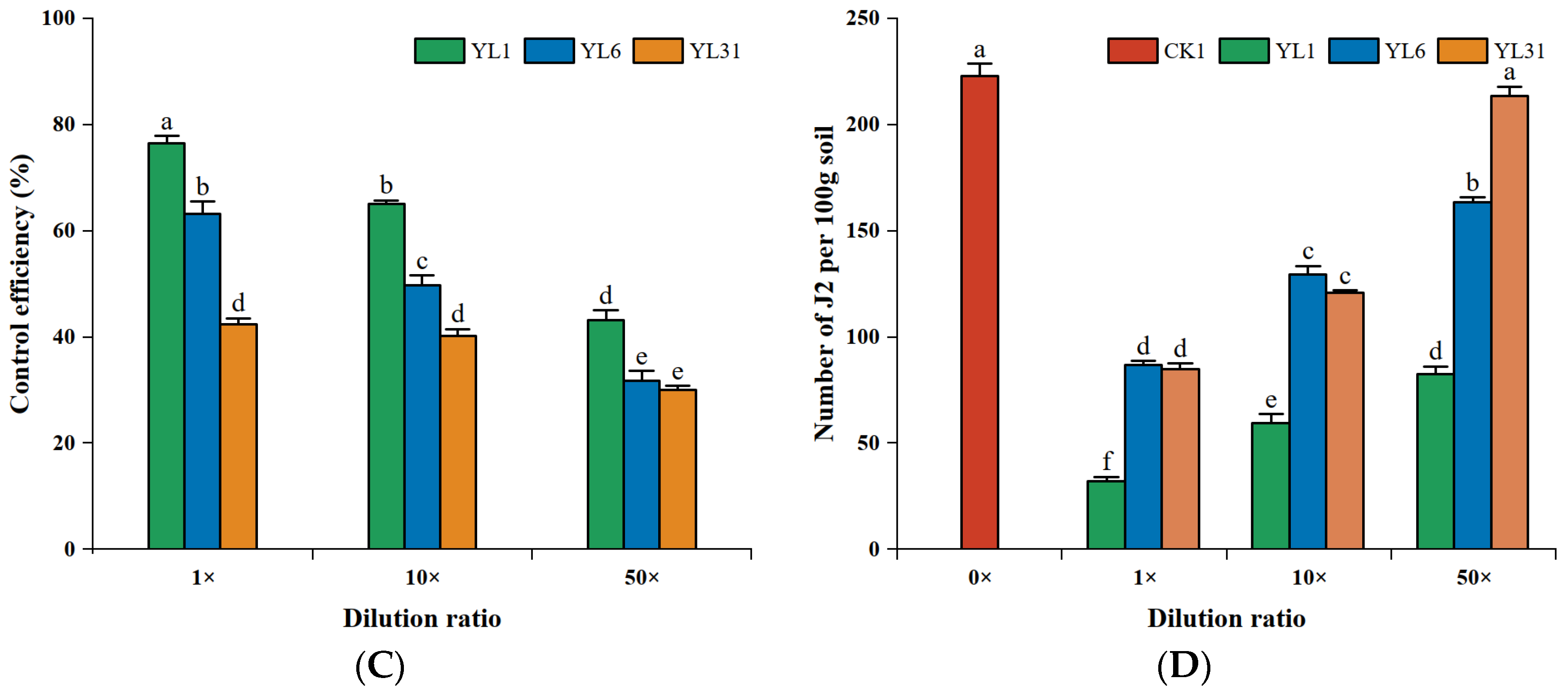
3.3. Effects of Antagonistic Bacteria on Root-Knot Nematode
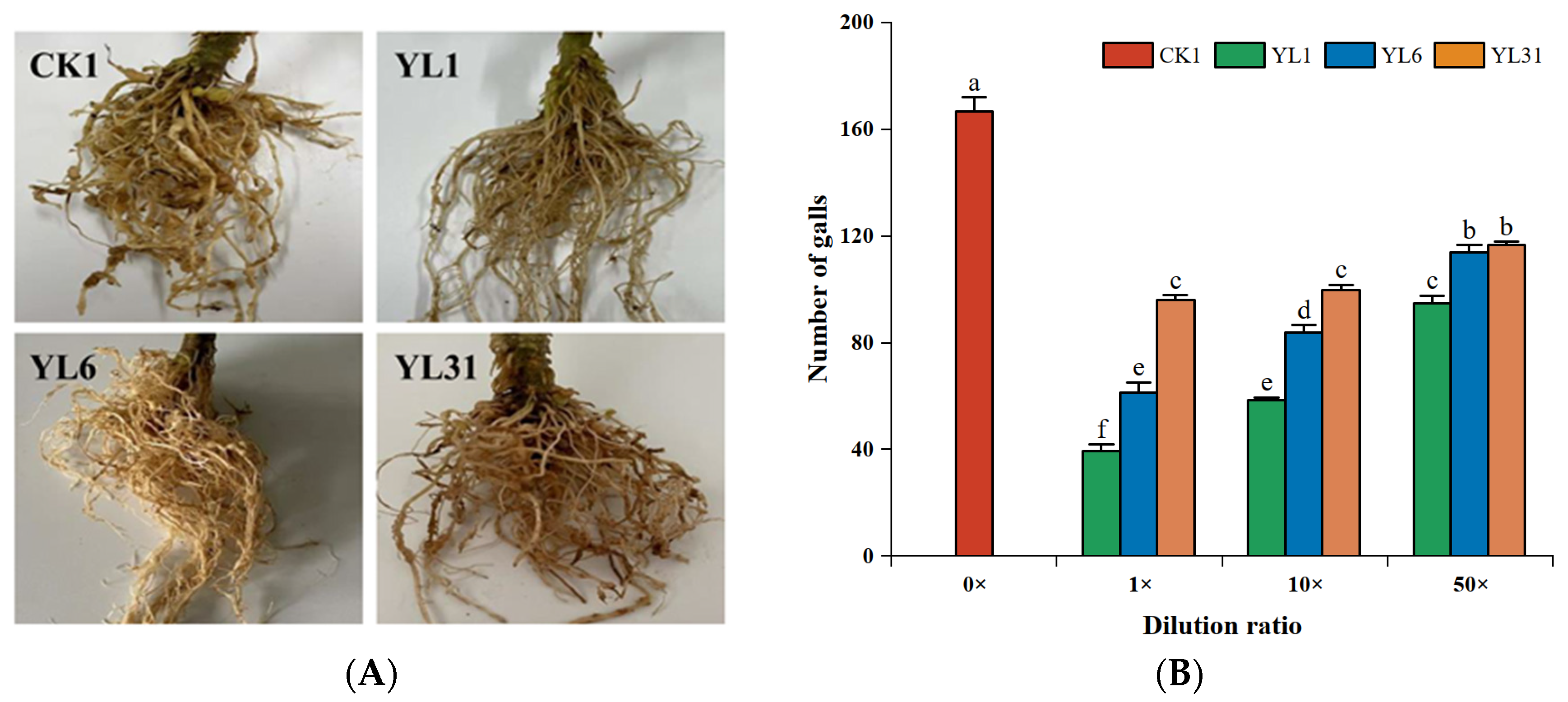
3.3.1. Root-Knot Nematode Egg Development
3.3.2. J2 of Root-Knot Nematode Development
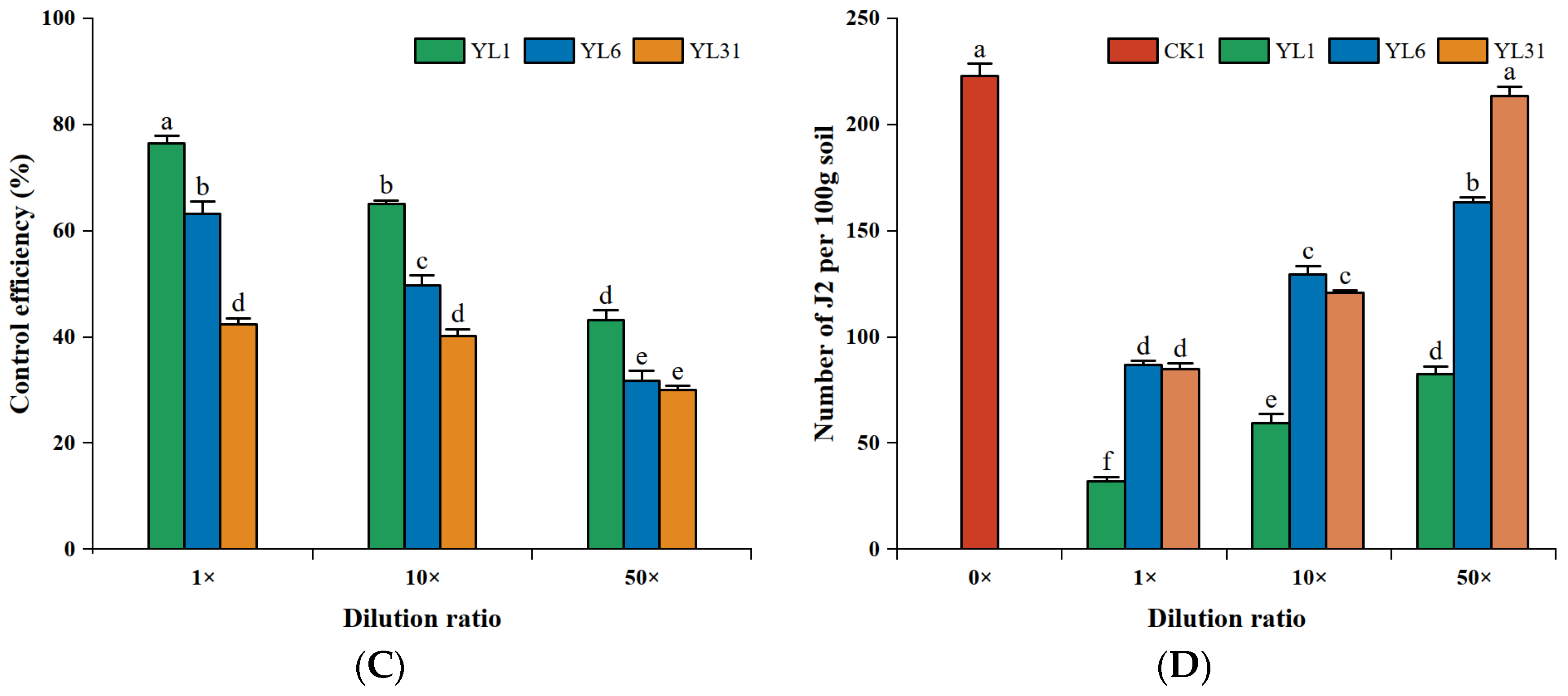
3.4. Efficacy in Controlling Nematodes of Tomato
3.4.1. Effects of Antagonistic Bacteria on Root-Knot Nematodes
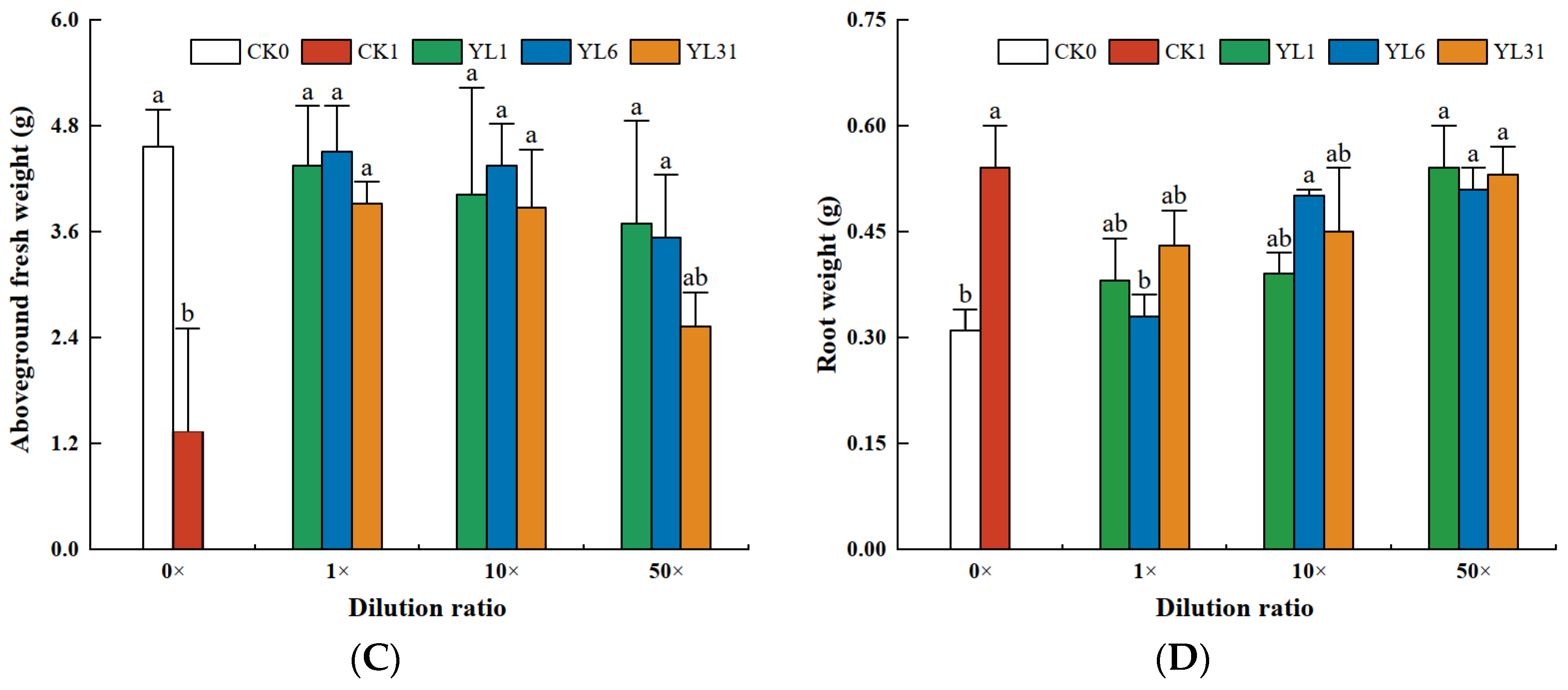
3.4.2. Effects of Antagonistic Bacteria on Tomato Growth
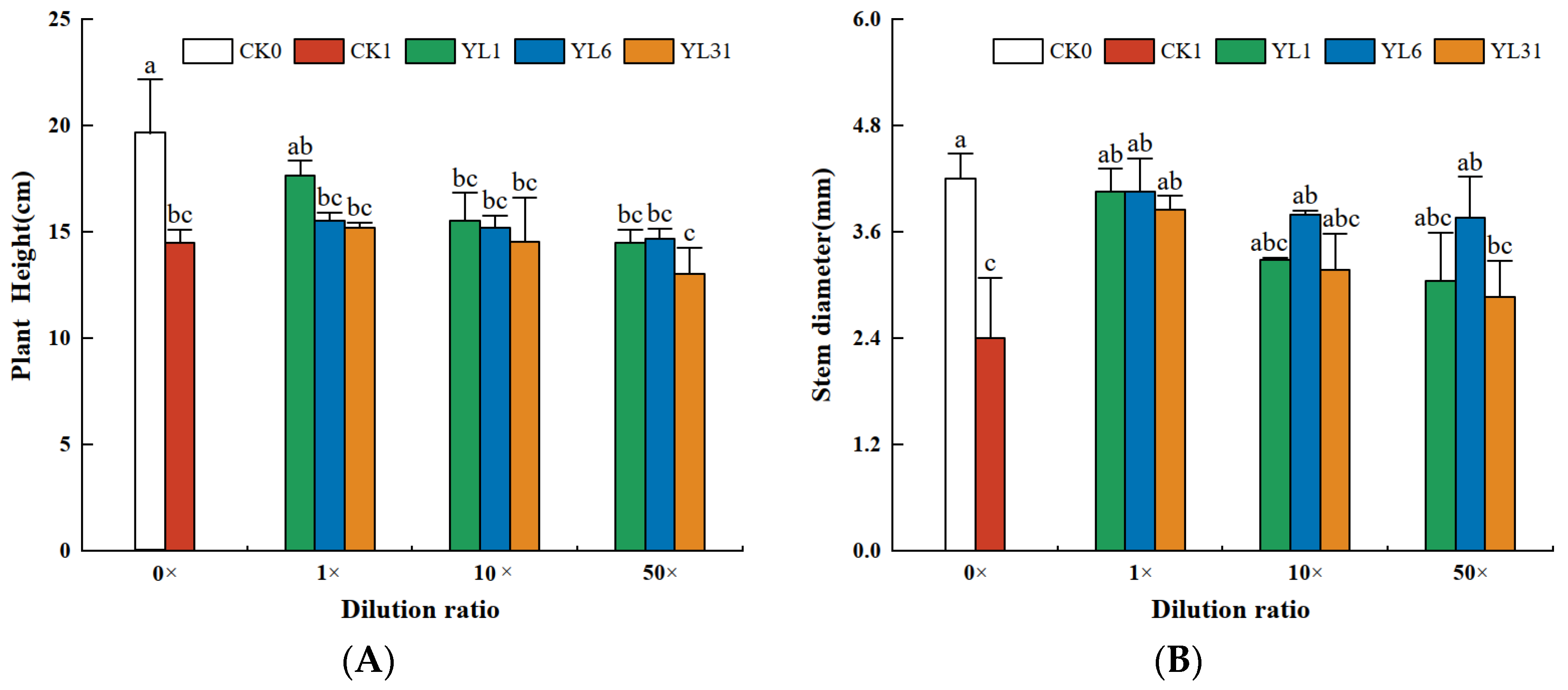
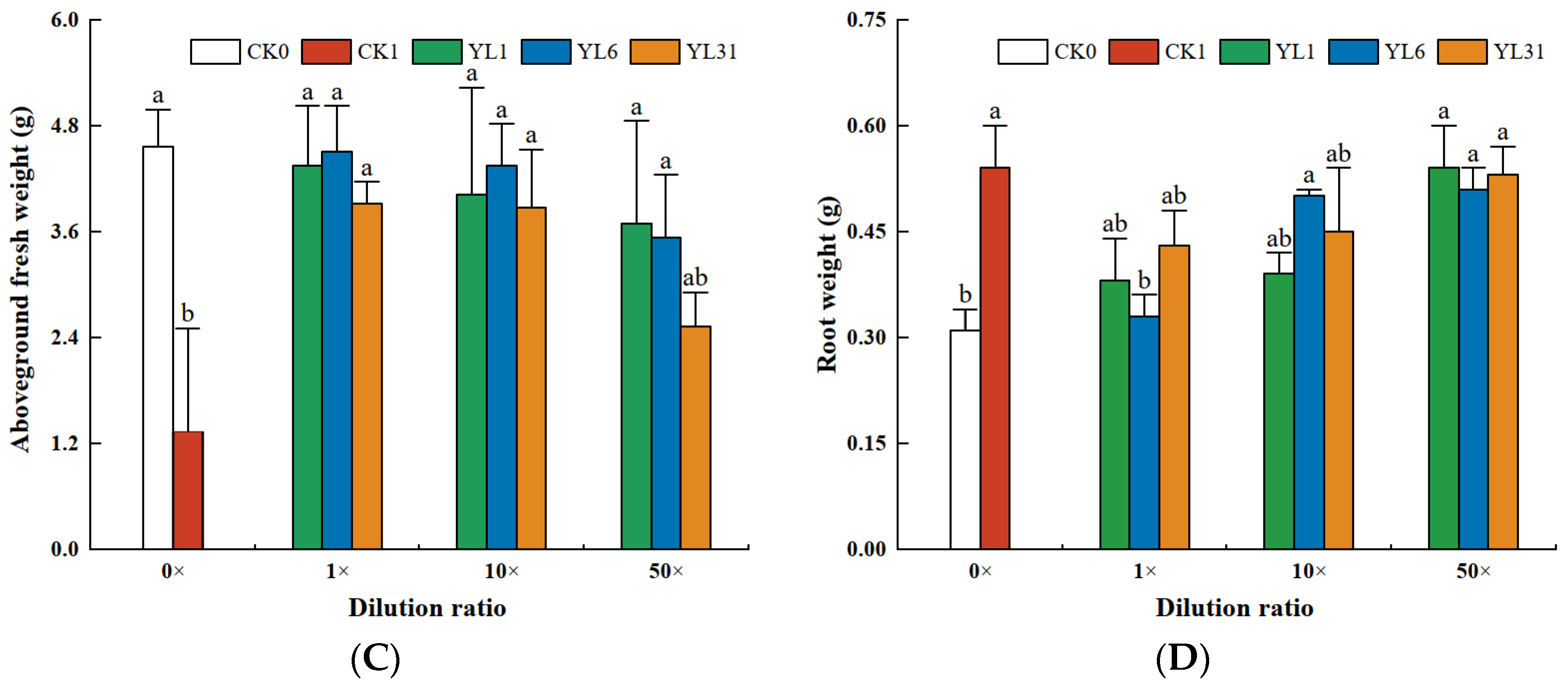
3.5. Efficacy in Controlling Nematodes of Cucumber
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mesa-Valle, C.M.; Garrido-Cardenas, J.A.; Cebrian-Carmona, J.; Talavera, M.; Manzano-Agugliaro, F. Global Research on Plant Nematodes. Agronomy 2020, 10, 1148. [Google Scholar] [CrossRef]
- Bernard, G.C.; Egnin, M.; Bonsi, C. The Impact of Plant-Parasitic Nematodes on Agriculture and Methods of Control. In Nematology—Concepts, Diagnosis and Control; Shah, M.M., Mahamood, M., Eds.; InTech: Rijeka, Croatia, 2017; pp. 121–151. [Google Scholar] [CrossRef]
- Coyne, D.L.; Cortada, L.; Dalzell, J.J.; Claudius-Cole, A.O.; Haukeland, S.; Luambano, N.; Talwana, H. Plant-Parasitic Nematodes and Food Security in Sub-Saharan Africa. Annu. Rev. Phytopathol. 2018, 56, 381–403. [Google Scholar] [CrossRef] [PubMed]
- Inácio, M.L.; Faria, J.M.S.; Haukeland, S. Editorial: Novel approaches for sustainable crop yield and management of plant-parasitic nematodes. Front. Plant Sci. 2023, 14, 1274757. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 Plant-Parasitic Nematodes in Molecular Plant Pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Martin, F.N. Development of Alternative Strategies for Management of Soilborne Pathogens Currently Controlled with Methyl Bromide. Annu. Rev. Phytopathol. 2003, 41, 325–350. [Google Scholar] [CrossRef]
- Aubertot, J.N.; West, J.S.; Bousset-Vaslin, L.; Salam, M.U.; Barbetti, M.J.; Diggle, A.J. Improved Resistance Management for Durable Disease Control: A Case Study of Phoma Stem Canker of Oilseed Rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 91–106. [Google Scholar] [CrossRef]
- Damalas, C.A.; Eleftherohorinos, I.G. Pesticide Exposure, Safety Issues, and Risk Assessment Indicators. Int. J. Environ. Res. Public Health 2011, 8, 1402–1419. [Google Scholar] [CrossRef] [PubMed]
- Antil, S.; Kumar, R.; Pathak, D.V.; Kumar, A.; Panwar, A.; Kumari, A. Plant Growth-Promoting Rhizobacteria—Bacillus Cereus KMT-5 and B. Megaterium KMT-8 Effectively Suppressed Meloidogyne Javanica Infection. Appl. Soil Ecol. 2022, 174, 104419. [Google Scholar] [CrossRef]
- Mohiddin, F.A.; Khan, M.R. Efficacy of newly developed biopesticides for the management of wilt disease complex of chickpea (Cicer arietinum L.). Legume Res. 2019, 42, 550–556. [Google Scholar] [CrossRef]
- Soares, F.E.d.F.; Aguilar-Marcelino, L.; Braga, F.R. Editorial: Nematophagous fungi as nematode control agents. Front. Fungal Biol. 2024, 4, 1353132. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The Fight against Plant-Parasitic Nematodes: Current Status of Bacterial and Fungal Biocontrol Agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef] [PubMed]
- Maqsood, A.; Aslam, M.N.; Khaliq, H.; Shakeel, M.T.; Wu, H.Y.; Fahad, S. Endophytic Bacillus spp. Mediated Plant Growth Promotion of Tomato Seedlings and Suppression of Meloidogyne incognita and Fusarium oxysporum Disease Complex. J. Plant Growth Regul. 2024, 43, 1–16. [Google Scholar] [CrossRef]
- Kumar, D. Effectiveness of various nematode-trapping fungi for biocontrol of the Meloidogyne incognita in tomato (Lycopersicion esculentum Mill.). Rhizosphere 2024, 29, 100845. [Google Scholar] [CrossRef]
- Mogollón-Ortiz, A.M.; Monteiro, T.S.A.; de Freitas, L.G.; de Queiroz, M.V. Potential of different species of actinobacteria in the management of Meloidogyne javanica. Arch. Microbiol. 2024, 206, 160. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Shakeel, A.; Waqar, S.; Handoo, Z.A.; Khan, A.A. Microbes vs. Nematodes: Insights into Biocontrol through Antagonistic Organisms to Control Root-Knot Nematodes. Plants 2023, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.W.; Chia, S.R.; Yen, H.W.; Nomanbhay, S.; Ho, Y.C.; Show, P.L. Transformation of Biomass Waste into Sustaina ble Organic Fertilizers. Sustainability 2019, 11, 2266. [Google Scholar] [CrossRef]
- Naghman, R.; Bhatti, M.T.; Najabat, Z.; Hyder, S.; Rizvi, Z.F.; Gondal, A.S.; Zafar, Z.; Malik, S.; Iqbal, R.; Hafeez, A.; et al. Organic amendments: A natural way to suppress phytopathogens, a sustainable approach to go green. Turk. J. Agric. For. 2023, 47, 602–622. [Google Scholar] [CrossRef]
- Rehman, S.U.; De Castro, F.; Aprile, A.; Benedetti, M.; Fanizzi, F.P. Vermicompost: Enhancing Plant Growth and Combating Abiotic and Biotic Stress. Agronomy 2023, 13, 1134. [Google Scholar] [CrossRef]
- Sonmez, S.; Asri, F.O. Importance of Vermicompost in Tomato Plant Cultivation and Improvement of Some Soil Properties. Compost. Sci. Util. 2023, 29, 44–56. [Google Scholar] [CrossRef]
- Zuhair, R.; Moustafa, Y.T.A.; Mustafa, N.S.A.; El-Dahshouri, M.F.; Zhang, L.X.; Ageba, M.F. Efficacy of amended vermicompost for bio-control of root knot nematode (RKN) Meloidogyne incognita infesting tomato in Egypt. Environ. Technol. Innov. 2022, 27, 102397. [Google Scholar] [CrossRef]
- Tikoria, R.; Ohri, P. Application of neem waste vermicompost in compensating nematode induced stress and upregulating physiological markers of tomato plants under glass house conditions after 10 days of exposure. Environ. Sci. Pollut. 2023, 30, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Liu, M.; Jiang, L.; Chen, X.; Griffiths, B.S.; Li, H.; Hu, F. Vermicompost Increases Defense against Root-Knot Nematode (Meloidogyne Incognita) in Tomato Plants. Appl. Soil Ecol. 2016, 105, 177–186. [Google Scholar] [CrossRef]
- Singh, B.; Devindrappa; Hazra, K.K.; Singh, U.; Gupta, S. Eco–Friendly Management of Meloidogyne Javanica in Chickpea (Cicer arietinum L.) Using Organic Amendments and Bio–Control Agent. J. Clean. Prod. 2020, 257, 120542. [Google Scholar] [CrossRef]
- Adam, M.A.M.; Phillips, M.S.; Blok, V.C. Molecular diagnostic key for identification of single juveniles of seven common and economically important species of root-knot nematode (Meloidogyne spp.). Plant Pathol. 2007, 56, 190–197. [Google Scholar] [CrossRef]
- Dong, X.Z.; Cai, M.Y. Identification System Manual of Common Bacteria; Science Press: Beijing, China, 2001. (In Chinese) [Google Scholar]
- Kumar, S.N.; Mohandas, C.; Nambisan, B. Purification, structural elucidation and bioactivity of tryptophan containing diketopiperazines, from Comamonas testosteroni associated with a rhabditid entomopathogenic nematode against major human-pa-thogenic bacteria. Peptides 2014, 53, 48–58. [Google Scholar] [CrossRef]
- Barker, K.R. Design of greenhouse and microplot experiments for evaluation of plant resistance to nematodes. In Plant Nematology Laboratory Manual; Zuckerman, B.M., Mai, W.F., Harrison, M.B., Station, M.A.E., Eds.; University of Massachusetts Amherst: Amherst, MA, USA, 1985; pp. 103–113. [Google Scholar]
- Arshad, K.; Amir, K.; Asgar, A.; Saba, F.; Ahmad, S.M. Root-Knot Nematodes (Meloidogyne spp.): Biology, Plant-Nematode Interactions and Their Environmentally Benign Management Strategies. Gesunde Pflanz. 2023, 75, 2187–2205. [Google Scholar] [CrossRef]
- Jenkins, W. A rapid centrifugal flotation technique for extracting nematodes from soil. Plant Dis. Report. 1964, 48, 692. [Google Scholar]
- Botelho, A.O.; Campos, V.P.; Da Silva, J.C.P.; Freire, E.S.; de Pinho, R.S.C.; Barros, A.F.; Oliveira, D.F. Physicochemical and biological properties of the coffee (Coffea arabica) rhizosphere suppress the root-knot nematode Meloidogyne exigua. Biocontrol Sci. Technol. 2019, 29, 1181–1196. [Google Scholar] [CrossRef]
- Chen, J.; Li, Q.X.; Song, B. Chemical nematicides: Recent research progress and outlook. J. Agric. Food Chem. 2020, 68, 12175–12188. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Wang, X.J.; Cao, Y.; Dong, Q.E.; Tong, J.Y.; Mo, M.H. Vermicomposting of Pleurotus eryngii spent mushroom substrates and the possible mechanisms of vermicompost suppressing nematode disease caused by Meloidogyne incognita. Heliyon 2023, 9, e15111. [Google Scholar] [CrossRef]
- Wharton, D. Nematode Egg-Shells. Parasitology 1980, 81, 447–463. [Google Scholar] [CrossRef] [PubMed]
- Burgwyn, B.; Nagel, B.; Ryerse, J.; Bolla, R.I. Heterodera Glycines: Eggshell Ultrastructure and Histochemical Localization of Chitinous Components. Exp. Parasitol. 2003, 104, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Girardi, N.S.; Sosa, A.L.; Etcheverry, M.G.; Passone, M.A. In vitro characterization bioassays of the nematophagous fungus Purpureocillium lilacinum: Evaluation on growth, extracellular enzymes, mycotoxins and survival in the surrounding agroecosystem of tomato. Fungal Biol. 2022, 126, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Nafady, N.A.; Sultan, R.; El-Zawahry, A.M.; Mostafa, Y.S.; Alamri, S.; Mostafa, R.G.; Hashem, M.; Hassan, E.A. Effective and Promising Strategy in Management of Tomato Root-Knot Nematodes by Trichoderma harzianum and Arbuscular Mycorrhizae. Agronomy 2022, 12, 315. [Google Scholar] [CrossRef]
- Chen, F.; Pang, C.Y.; Zheng, Z.Q.; Zhou, W.; Guo, Z.Q.; Xiao, D.Y.; Du, H.W.; Bravo, A.; Soberón, M.; Sun, M.; et al. Aminopeptidase MNP-1 triggers intestine protease production by activating daf-16 nuclear location to degrade pore-forming toxins in Caenorhabditis elegans. PLoS Pathog. 2023, 19, e1011507. [Google Scholar] [CrossRef]
- Horak, I.; Van Rensburg, P.J.J.; Claassens, S. Effect of cultivation media and temperature on metabolite profiles of three nematicidal Bacillus species. Nematology 2022, 24, 383–399. [Google Scholar] [CrossRef]
- Li, B.Y.; Wang, P.Y.; Yang, L.L.; Rang, X.; Zhou, W.Z.; Liu, Y.J. Chemotaxis of Meloidogyne incognita Response to Rhizosphere Bacteria. Microorganisms 2023, 11, 2271. [Google Scholar] [CrossRef] [PubMed]
- Fallahzadeh-Mamaghani, V.; Shahbazi-Ezmareh, R.; Shirzad, A.; Moslehi, S. Possible mechanisms of action of Bacillus wiedmannii AzBw1, a biocontrol agent of the root-knot nematode, Meloidogyne arenaria. Egypt. J. Biol. Pest Control 2023, 33, 28. [Google Scholar] [CrossRef]
- Karacic, V.; Miljakovic, D.; Marinkovic, J.; Ignjatov, M.; Milosevic, D.; Tamindzic, G.; Ivanovic, M. Bacillus Species: Excellent Biocontrol Agents against Tomato Diseases. Microorganisms 2024, 12, 457. [Google Scholar] [CrossRef]
- Díaz-Manzano, F.E.; Amora, D.X.; Martínez-Gómez, A.; Moelbak, L.; Escobar, C. Biocontrol of Meloidogyne spp. in Solanum lycopersicum using a dual combination of Bacillus strains. Front. Plant Sci. 2023, 13, 1077062. [Google Scholar] [CrossRef]
- Geng, Y.H. Application Effect of Bacillus in Tomato Root Knot Nematode Disease Control. J. Biobased Mater. Bioenergy 2024, 18, 172–180. [Google Scholar] [CrossRef]
- Gowda, A.P.A.; Pankaj, D.; Singh, D.; Singh, A.K.; Sowmya, R. Nematicidal potential of plant growth-promoting rhizobacteria against Meloidogyne incognita infesting tomato under protected cultivation. Egypt. J. Biol. Pest Control 2022, 32, 145. [Google Scholar] [CrossRef]
- Dinardo-Miranda, L.L.; Miranda, I.D.; Silva, H.D.S.; Fracasso, J.V. Biological control of phytoparasitic nematodes in sugarcane fields. Pesqui. Agropecuária Trop. 2022, 52, 1–7. [Google Scholar] [CrossRef]
- Wang, J.; Wen, C.; Liu, W.; Dong, D.; Qiu, J.; Liu, T. Screening and Identification of Biocontrol Bacterial Against Root-Knot Nematode. North. Hortic. 2016, 9, 121–124. [Google Scholar] [CrossRef]
- Settu, V.; Annaiyan, S.; Mannu, J. Revealing the genetic arsenal of Bacillus firmus TNAU1: Unleashing nematicidal and plant growth promotion traits. Physiol. Mol. Plant Pathol. 2023, 129, 102177. [Google Scholar] [CrossRef]
- Dai, M.-M.; Liu, R.; Jiang, H.; Zhang, X.-P.; Song, W.-W.; Zhang, J.; Liang, C.; Zhao, H.-H.; Shi, Q.-Q. Volatile Organic Compounds of Bacillus pumilus Strain S1-10 Exhibit Fumigant Activity Against Meloidogyne incognita. Plant Dis. 2023, 107, 3057–3063. [Google Scholar] [CrossRef] [PubMed]
- Zuo, M.; Tan, J.; Xiang, B.K.; Shi, H.L.; Chen, H.H.; Peng, W.X.; Yin, Z.C. Effects of different Bacillus on bacterial community of tobacco rhizosphere soil and control of root-knot nematodes. Tob. Sci. Technol. 2022, 55, 8–15. (In Chinese) [Google Scholar] [CrossRef]
- Liu, Z. Isolation and Identification of Salt-Tolerant Growth-Promoting Bacteria and the Effects on Brassica Campestris under Salt-Alkali Conditions. Master’s Thesis, Shandong Agricultural University, Taian City, China, 2022. (In Chinese). [Google Scholar]
- Lu, Z.H.; Ruan, Z.Y.; Zhao, B.Q.; Wang, H.M.; Zhao, J.C.; Zhao, W.Y.; Liu, H.; Sou, Z.; Wang, Y.W.; Liu, X.F. The Applications of Brevibacterium frigoritolerans and Microbial Agents. CN 103952348 B, 11 January 2017. (In Chinese) [Google Scholar]
- Yang, L.-L.; Huang, Y.; Liu, J.; Ma, L.; Mo, M.-H.; Li, W.-J.; Yang, F.-X. Lysinibacillus mangiferahumi sp. nov., a new bacterium producing nematicidal volatiles. Antonie Van Leeuwenhoek 2012, 102, 53–59. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, D.W.; Cheng, F.X.; Zhu, C.H.; Wang, Z.Y.; Shi, X.B.; Zheng, L.M.; Su, P.; Liu, Y.; Zhang, D.Y. Isolation, identification and application of a strain of Lysinibacillus macrolides against Meloidogyne incognita. J. South. Agric. 2021, 52, 2765–2775. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, Q. Using large-scale multi-module NRPS to heterologously prepare highly efficient lipopeptide biosurfactants in recombinant Escherichia coli. Enzym. Microb. Technol. 2022, 159, 110068. [Google Scholar] [CrossRef]
- Mathivanan, K.; Chandirika, J.U.; Vinothkanna, A.; Govindarajan, R.K.; Meng, D.; Yin, H. Characterization and Biotechnological Functional Activities of Exopolysaccharides Produced by Lysinibacillus fusiformis KMNTT-10. J. Polym. Environ. 2021, 29, 1742–1751. [Google Scholar] [CrossRef]
- Wani, P.A.; Ebudola, A.N.; Olusebi, Y.K.; Rafi, N.; Fawzhia, O.S.J.; Oluwaseun, O.I. Hydrocarbon Utilizing and Metal Tolerant Bacteria Simultaneously Degrade Hydrocarbons and Detoxify Metals in Petroleum Contaminated Soil. Geomicrobiol. J. 2023, 40, 372–381. [Google Scholar] [CrossRef]
- Castro, N.R.; Zavala, J.; Gil-Kodaka, P.; Diringer, B. Evaluation of bacterial strains to improve the productivity of microalgae used in bivalve hatcheries in Peru. J. World Aquac. Soc. 2022, 53, 95–105. [Google Scholar] [CrossRef]
- Trukhachev, V.I.; Belopukhov, S.L.; Grigoryeva, M.; Dmitrevskaya, I.I. Study of the Sustainability of Ecological and Chemical Indicators of Soils in Organic Farming. Sustainability 2024, 16, 665. [Google Scholar] [CrossRef]
- Ghoreshizadeh, S.; Calvo-Pena, C.; Ruiz-Munoz, M.; Otero-Suarez, R.; Coque, J.J.R.; Cobos, R. Pseudomonas taetrolens ULE-PH5 and Pseudomonas sp. ULE-PH6 Isolated from the Hop Rhizosphere Increase Phosphate Assimilation by the Plant. Plants 2024, 13, 402. [Google Scholar] [CrossRef] [PubMed]
- Soumare, A.; Sarr, D.; Diedhiou, A.G. Potassium sources, microorganisms and plant nutrition: Challenges and future research directions. Pedosphere 2023, 33, 105–115. [Google Scholar] [CrossRef]
- Zapata, D.; López, J.E.; Saldarriaga, J.F. Plant Growth-Promoting and Biocontrol Potential of Aspergillus tubingensis and Talaromyces islandicus. J. Soil Sci. Plant Nutr. 2024, 1–17. [Google Scholar] [CrossRef]
- Song, Z.J.; Xiao, L.H.; Guo, X.L.; Zhu, Y.Z.; An, X.L.; Tan, Y.; Zhang, X.Y.; Wang, D.L. Effects of inoculating different mycorrhizal fungi on rhizosphere soil fungi and nutrient uptake of blueberry. Hortic. Environ. Biotechnol. 2024, 65, 29–41. [Google Scholar] [CrossRef]
- Costa, M.N.; Gil, T.; Teixeira, R.; dos Santos, A.S.R.; Romão, I.R.; López, C.S.; Vílchez, J.I. Combined Use of a Bacterial Consortium and Early-Colonizing Plants as a Treatment for Soil Recovery after Fire: A Model Based on Los Guajares (Granada, Spain) Wildfire. Biology 2023, 12, 1093. [Google Scholar] [CrossRef]
- Jha, Y.; Mohamed, H.I. Inoculation with Lysinibacillus fusiformis Strain YJ4 and Lysinibacillus sphaericus Strain YJ5 Alleviates the Effects of Cold Stress in Maize Plants. Gesunde Pflanz. 2023, 75, 77–95. [Google Scholar] [CrossRef]
- Santosh, S.; Velmourougane, K.; Idapuganti, R.G.; Manikandan, A.; Blaise, D. Potassium Solubilizing Potential of Native Bacterial Isolates from Cotton Rhizosphere of Rainfed Vertisols. Natl. Acad. Sci. Lett. 2022, 45, 209–212. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Root-Knot Index (%) | Control Efficacy (%) | Nematode Population/100 g Soil |
|---|---|---|---|
| CK | 71.9 ± 2.0 a | - | 1916.2 ± 46.4 a |
| YL1 | 21.1 ± 0.8 d | 70.6 ± 1.2 a | 653.5 ± 48.3 d |
| YL6 | 30.3 ± 0.3 c | 57.8 ± 0.4 b | 1075.5 ± 35.7 c |
| YL31 | 43.3 ± 1.2 b | 39.7 ± 1.7 c | 1223.8 ± 71.5 b |
| Treatment | Aboveground Weight (g) | Underground Weight (g) | Yield (kg ha−1) | ||
|---|---|---|---|---|---|
| Fresh | Dry | Fresh | Dry | ||
| CK | 246.8 ± 4.8 c | 22.1 ± 0.7 b | 15.8 ± 1.4 a | 1.4 ± 0.1 a | 102965.0 ± 39.4 d |
| YL1 | 434.3 ± 18.0 a | 33.4 ± 7.3 a | 10.5 ± 1.5 b | 0.9 ± 0.1 b | 118350.0 ± 48.2 a |
| YL6 | 379.4 ± 13.9 ab | 27.0 ± 3.0 a | 10.9 ± 0.8 b | 0.9 ± 0.1 b | 109911.3 ± 24.1 b |
| YL31 | 373.8 ± 25.1 b | 27.8 ± 4.1 a | 11.4 ± 1.7 ab | 1.0 ± 0.2 ab | 107338.8 ± 23.1 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, C.; Yang, D.; Dong, F.; Shang, J.; Niu, X.; Zhang, G.; Yang, L.; Wang, Y. Biocontrol Potential of Bacteria Isolated from Vermicompost against Meloidogyne incognita on Tomato and Cucumber Crops. Horticulturae 2024, 10, 407. https://doi.org/10.3390/horticulturae10040407
Liang C, Yang D, Dong F, Shang J, Niu X, Zhang G, Yang L, Wang Y. Biocontrol Potential of Bacteria Isolated from Vermicompost against Meloidogyne incognita on Tomato and Cucumber Crops. Horticulturae. 2024; 10(4):407. https://doi.org/10.3390/horticulturae10040407
Chicago/Turabian StyleLiang, Chen, Diwen Yang, Feiyu Dong, Jingxia Shang, Xuguang Niu, Guoxian Zhang, Lijuan Yang, and Yonghuan Wang. 2024. "Biocontrol Potential of Bacteria Isolated from Vermicompost against Meloidogyne incognita on Tomato and Cucumber Crops" Horticulturae 10, no. 4: 407. https://doi.org/10.3390/horticulturae10040407