
A Comparative Study of the Influence of Soil and Non-Soil Factors on Seed Germination of Edible Salt-Tolerant Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Acquirement and Greenhouse Conditions
2.2. Germination Conditions
2.3. Seed Treatments
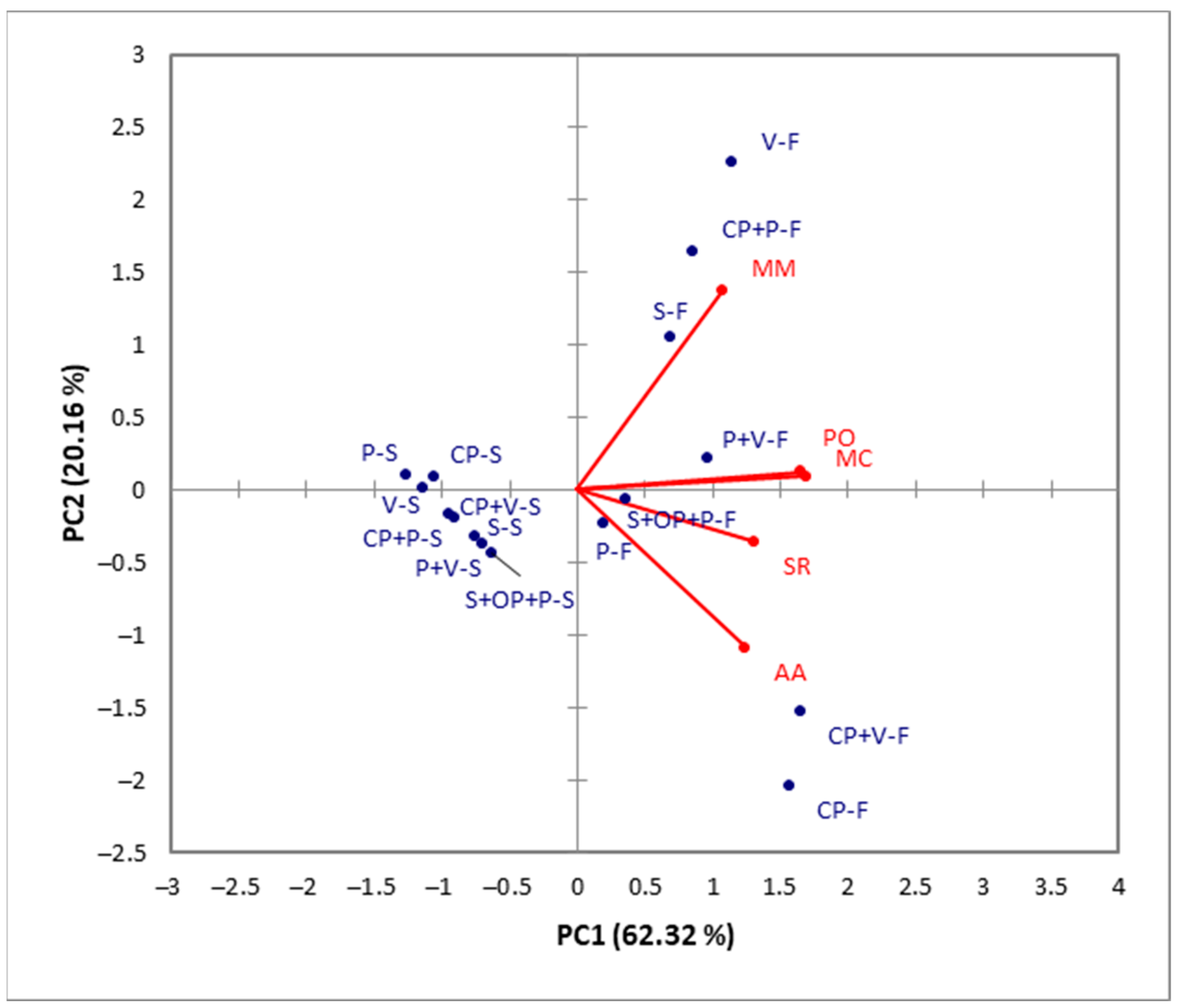
2.4. Statistical Analysis
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Soligno, I. Spatial and Temporal Patterns of Global Freshwater Use for Food Production and Trade. Ph.D. Thesis, Graduate School of Politecnico di Torino, Torino, Italy, 2019. [Google Scholar]
- Vanham, D.; Guenther, S.; Ros-Baró, M.; Bach-Faig, A. Which diet has the lower water footprint in Mediterranean countries? Resour. Conserv. Recycl. 2021, 171, 105631. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, S.S.; Likens, G.E.; Pace, M.L.; Reimer, J.E.; Maas, C.M.; Galella, J.G.; Utz, R.M.; Duan, S.; Kryger, J.R.; Yaculak, A.M. Freshwater salinization syndrome: From emerging global problem to managing risks. Biogeochemistry 2021, 154, 255–292. [Google Scholar] [CrossRef]
- Liu, L.; Wang, B. Protection of halophytes and their uses for cultivation of saline-alkali soil in China. Biology 2021, 10, 353. [Google Scholar] [CrossRef]
- FAO; UN. How to Feed the World in 2050. In Proceedings of the High Level Expert Forum—How to Feed the World in 2050, Rome, Italy, 12–13 October 2009. [Google Scholar]
- Vanham, D.; Leip, A.; Galli, A.; Kastner, T.; Bruckner, M.; Uwizeye, A.; Van Dijk, K.; Ercin, E.; Dalin, C.; Brandão, M. Environmental footprint family to address local to planetary sustainability and deliver on the SDGs. Sci. Total Environ. 2019, 693, 133642. [Google Scholar] [CrossRef]
- FAO/WHO. Sustainable Healthy Diets–Guiding Principles. In Proceedings of the Sustainable Healthy Diets–Guiding Principles, Rome, Italy, 29 October 2019. [Google Scholar]
- Song, J.; Wang, B. Using euhalophytes to understand salt tolerance and to develop saline agriculture: Suaeda salsa as a promising model. Ann. Bot. 2015, 115, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Barreira, L.; Resek, E.; Rodrigues, M.J.; Rocha, M.I.; Pereira, H.; Bandarra, N.; da Silva, M.M.; Varela, J.; Custódio, L. Halophytes: Gourmet food with nutritional health benefits? J. Food Compos. Anal. 2017, 59, 35–42. [Google Scholar] [CrossRef]
- Castañeda-Loaiza, V.; Oliveira, M.; Santos, T.; Schüler, L.; Lima, A.R.; Gama, F.; Salazar, M.; da Rosa Neng, N.; Nogueira, J.M.F.; Varela, J. Wild vs. cultivated halophytes: Nutritional and functional differences. Food Chem. 2020, 333, 127536. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C.F.R. Halophytic herbs of the Mediterranean basin: An alternative approach to health. Food Chem. Toxicol. 2018, 114, 155–169. [Google Scholar] [CrossRef]
- Custódio, L.; Rodrigues, M.J.; Pereira, C.G.; Castañeda-Loaiza, V.; Fernandes, E.; Standing, D.; Neori, A.; Shpigel, M.; Sagi, M. A review on Sarcocornia species: Ethnopharmacology, nutritional properties, phytochemistry, biological activities and propagation. Foods 2021, 10, 2778. [Google Scholar] [CrossRef]
- Sánchez-Faure, A.; Calvo, M.M.; Pérez-Jiménez, J.; Martín-Diana, A.B.; Rico, D.; Montero, M.P.; del Carmen Gómez-Guillén, M.; López-Caballero, M.E.; Martínez-Alvarez, O. Exploring the potential of common iceplant, seaside arrowgrass and sea fennel as edible halophytic plants. Food Res. Int. 2020, 137, 109613. [Google Scholar] [CrossRef]
- Sezgin, A.C.; Sanlier, N. A new generation plant for the conventional cuisine: Quinoa (Chenopodium quinoa Willd.). Trends Food Sci. Technol. 2019, 86, 51–58. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef]
- Hamed, K.B.; Castagna, A.; Ranieri, A.; Garcia-Caparros, P.; Santin, M.; Hernandez, J.A.; Espin, G.B. Halophyte based Mediterranean agriculture in the contexts of food insecurity and global climate change. Environ. Exp. Bot. 2021, 191, 104601. [Google Scholar] [CrossRef]
- Gairola, S.; Hameed, A.; Rasheed, A.; Alketbi, A.; Aljasmi, M.; El-Keblawy, A. Seed germination and salinity tolerance of habitat-indifferent halophytes as associated with geographical distribution. Seed Sci. Technol. 2022, 50, 125–140. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Gulzar, S.; Khan, M.A. Role of Dormancy Regulating Chemicals in Alleviating the Seed Germination of Three Playa Halophytes. Ekoloji Derg. 2014, 23, 1–7. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, S.; Zhang, F.; Si, L.; Yu, Y.; Zhao, X.; Zhang, D.; Wang, W. Inheritance of downy mildew resistance at different developmental stages in Chinese cabbage via the leaf disk test. Hortic. Environ. Biotechnol. 2012, 53, 397–403. [Google Scholar] [CrossRef]
- Di Lorenzo, R.; Pisciotta, A.; Santamaria, P.; Scariot, V. From soil to soil-less in horticulture: Quality and typicity. Ital. J. Agron. 2013, 8, e30. [Google Scholar] [CrossRef]
- Bilderback, T.E.; Warren, S.L.; Owen, J.S.; Albano, J.P. Healthy substrates need physicals too! Horttechnology 2005, 15, 747–751. [Google Scholar] [CrossRef]
- Rodrigues, M.J.; Monteiro, I.; Castañeda-Loaiza, V.; Placines, C.; Oliveira, M.C.; Reis, C.; Caperta, A.D.; Soares, F.; Pousão-Ferreira, P.; Pereira, C. Growth performance, in vitro antioxidant properties and chemical composition of the halophyte Limonium algarvense Erben are strongly influenced by the irrigation salinity. Ind. Crops Prod. 2020, 143, 111930. [Google Scholar] [CrossRef]
- Cai, J.-F.; Liu, X.-S.; Sun, K.; Jiang, F.; Zhang, M.-X.; Li, H.-L.; Yu, F.-H. Vermicompost Application Enhances Halophyte Suaeda salsa Performance and Improves Coastal Saline Soil Quality. J. Soil Sci. Plant Nutr. 2022, 22, 294–305. [Google Scholar] [CrossRef]
- Rozema, J.; Van Der List, J.C.; Schat, H.; van Diggelen, J.; Broekman, R.A. Ecophysiological response of Salicornia dolichostachya and Salicornia brachystachya to seawater inundation. In Vegetation between Land and Sea; Springer: Berlin/Heidelberg, Germany, 1987; pp. 180–186. [Google Scholar]
- Weeplian, T.; Yen, T.-B.; Ho, Y.-S. Growth, development, and chemical constituents of edible ice plant (Mesembryanthemum crystallinum L.) produced under combinations of light-emitting diode lights. HortScience 2018, 53, 865–874. [Google Scholar] [CrossRef]
- Ameixa, O.M.C.C.; Marques, B.; Fernandes, V.S.; Soares, A.M.V.M.; Calado, R.; Lillebø, A.I. Dimorphic seeds of Salicornia ramosissima display contrasting germination responses under different salinities. Ecol. Eng. 2016, 87, 120–123. [Google Scholar] [CrossRef]
- Figueira, C.; Ferreira, M.J.; Silva, H.; Cunha, A. Improved germination efficiency of Salicornia ramosissima seeds inoculated with Bacillus aryabhattai SP1016-20. Ann. Appl. Biol. 2019, 174, 319–328. [Google Scholar] [CrossRef]
- Scippa, G.S.; Petrollini, E.; Trupiano, D.; Rocco, M.; Falco, G.; Di Michele, M.; Chiatante, D. Dormancy of Medicago marina (L.) seed. Environ. Exp. Bot. 2011, 72, 320–329. [Google Scholar] [CrossRef]
- Bendimered, F.Z.; Mehdadi, Z.; Benhassaini, H. Étude de la germination et de la croissance foliaire de l’oyat (Ammophila arenaria (L.) Link) en conditions contrôlées. Acta Bot. Gall. 2007, 154, 129–140. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Ansari, F. Effects of site of origin, time of seed maturation, and seed age on germination behavior of Portulaca oleracea from the Old and New Worlds. Can. J. Bot. 2000, 78, 279–287. [Google Scholar]
- Chauhan, B.S.; Johnson, D.E. Seed germination ecology of Portulaca oleracea L.: An important weed of rice and upland crops. Ann. Appl. Biol. 2009, 155, 61–69. [Google Scholar] [CrossRef]
- Franco, J.A.; Cros, V.; Vicente, M.J.; Martínez-Sánchez, J.J. Effects of salinity on the germination, growth, and nitrate contents of purslane (Portulaca oleracea L.) cultivated under different climatic conditions. J. Hortic. Sci. Biotechnol. 2011, 86, 1–6. [Google Scholar] [CrossRef]
- Shaygan, M.; Baumgartl, T.; Arnold, S. Germination of Atriplex halimus seeds under salinity and water stress. Ecol. Eng. 2017, 102, 636–640. [Google Scholar] [CrossRef]
- Osman, A.E.; Ghassali, F. Effects of storage conditions and presence of fruiting bracts on the germination of Atriplex halimus and Salsola vermiculata. Exp. Agric. 1997, 33, 149–155. [Google Scholar] [CrossRef]
- Hamza, L.; Mederbal, K.; Regagba, Z.; Dahou, A.A.; Medjahed, M. Effect of Salinity and Temperature on Seed Germination of Atriplex halimus L.(halimus and schweinfurthii subspecies) Harvested in Western Algerian Region. Indian J. Agric. Res. 2023, 57, 435–441. [Google Scholar] [CrossRef]
- Brenchley, J.L.; Probert, R.J. Seed germination responses to some environmental factors in the seagrass Zostera capricorni from eastern Australia. Aquat. Bot. 1998, 62, 177–188. [Google Scholar] [CrossRef]
- Debez, A.; Ben Hamed, K.; Grignon, C.; Abdelly, C. Salinity effects on germination, growth, and seed production of the halophyte Cakile maritima. Plant Soil. 2004, 262, 179–189. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Aloisi, I.; Del Duca, S.; Canelo, V.; Torrigiani, P.; Silva, H.; Biondi, S. Salares versus coastal ecotypes of quinoa: Salinity responses in Chilean landraces from contrasting habitats. Plant Physiol. Biochem. 2016, 101, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Grzybowski, C.R.d.S.; Ohlson, O.d.C.; Silva, R.C.d.; Panobianco, M. Viability of barley seeds by the tetrazolium test. Rev. Bras. Sementes 2012, 34, 47–54. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Y.; Sun, J.; Meng, J.; Tao, J. Deterioration of orthodox seeds during ageing: Influencing factors, physiological alterations and the role of reactive oxygen species. Plant Physiol. Biochem. 2021, 158, 475–485. [Google Scholar] [CrossRef] [PubMed]
- Arora, S.; Rao, G.G. Bio-amelioration of salt-affected soils through halophyte plant species. In Bioremediation of Salt Affected Soils: An Indian Perspective; Springer: Berlin/Heidelberg, Germany, 2017; pp. 71–85. ISBN 9783319482576. [Google Scholar]
- Mandák, B. Germination requirements of invasive and non-invasive Atriplex species: A comparative study. Flora-Morphol. Distrib. Funct. Ecol. Plants 2003, 198, 45–54. [Google Scholar] [CrossRef]
- Mikhiel, G.S.; Meyer, S.E.; Pendleton, R.L. Variation in germination response to temperature and salinity in shrubby Atriplex species. J. Arid Environ. 1992, 22, 39–49. [Google Scholar] [CrossRef]
- Terrones, A.; Moreno, J.; Agulló, J.C.; Villar, J.L.; Vicente, A.; Alonso, M.Á.; Juan, A. Influence of salinity and storage on germination of Tamarix taxa with contrasted ecological requirements. J. Arid Environ. 2016, 135, 17–21. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Bailly, C.; El-Maarouf-Bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual role of reactive oxygen species in seed physiology. C. R. Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef]
- Panuccio, M.R.; Jacobsen, S.E.; Akhtar, S.S.; Muscolo, A. Effect of saline water on seed germination and early seedling growth of the halophyte quinoa. AoB Plants 2014, 6, plu047. [Google Scholar] [CrossRef] [PubMed]
- Ventura, Y.; Wuddineh, W.A.; Myrzabayeva, M.; Alikulov, Z.; Khozin-Goldberg, I.; Shpigel, M.; Samocha, T.M.; Sagi, M. Effect of seawater concentration on the productivity and nutritional value of annual Salicornia and perennial Sarcocornia halophytes as leafy vegetable crops. Sci. Hortic. 2011, 128, 189–196. [Google Scholar] [CrossRef]
- Saddhe, A.A.; Malvankar, M.R.; Karle, S.B.; Kumar, K. Reactive nitrogen species: Paradigms of cellular signaling and regulation of salt stress in plants. Environ. Exp. Bot. 2019, 161, 86–97. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Hameister, R.; Kaur, C.; Dheen, S.T.; Lohmann, C.H.; Singh, G. Reactive oxygen/nitrogen species (ROS/RNS) and oxidative stress in arthroplasty. J. Biomed. Mater. Res. Part B Appl. Biomater. 2020, 108, 2073–2087. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Anee, T.I.; Parvin, K.; Nahar, K.; Mahmud, J.A.; Fujita, M. Regulation of ascorbate-glutathione pathway in mitigating oxidative damage in plants under abiotic stress. Antioxidants 2019, 8, 384. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Öztürk, M. Ecophysiology, Abiotic Stress Responses and Utilization of Halophytes; Springer: Berlin/Heidelberg, Germany, 2019; ISBN 9811337624. [Google Scholar]
- Hussain, A.; Iqbal, K.; Aziem, S.; Mahato, P.; Negi, A.K. A review on the science of growing crops without soil (soilless culture)-a novel alternative for growing crops. Int. J. Agric. Crop Sci. 2014, 7, 833. [Google Scholar]
- Bewley, J.D.; Bradford, K.; Hilhorst, H. Chapter 4: Germination. In Seeds: Physiology of Development, Germination and Dormancy; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; pp. 133–181. ISBN 1461446929. [Google Scholar]
- Loi, A.; Howieson, J.G.; Nutt, B.J.; Carr, S.J. A second generation of annual pasture legumes and their potential for inclusion in Mediterranean-type farming systems. Aust. J. Exp. Agric. 2005, 45, 289–299. [Google Scholar] [CrossRef]
- Qu, X.-X.; Huang, Z.-Y.; Baskin, J.M.; Baskin, C.C. Effect of temperature, light and salinity on seed germination and radicle growth of the geographically widespread halophyte shrub Halocnemum strobilaceum. Ann. Bot. 2008, 101, 293–299. [Google Scholar] [CrossRef]
- Dashti, F.; Ghahremani-Majd, H.; Esna-Ashari, M. Overcoming seed dormancy of mooseer (Allium hirtifolium) through cold stratification, gibberellic acid, and acid scarification. J. For. Res. 2012, 23, 707–710. [Google Scholar] [CrossRef]
- Schelin, M.; Tigabu, M.; Eriksson, I.; Sawadogo, L.; Oden, P.C. Effects of scarification, gibberellic acid and dry heat treatments on the germination of Balanites aegyptiaca seeds from the Sudanian savanna in Burkina Faso. Seed Sci. Technol. 2003, 31, 605–617. [Google Scholar] [CrossRef]
- Babaei-Ghaghelestany, A.; Alebrahim, M.T.; MacGregor, D.R.; Khatami, S.A.; Hasani Nasab Farzaneh, R. Evaluation of ultrasound technology to break seed dormancy of common lambsquarters (Chenopodium album). Food Sci. Nutr. 2020, 8, 2662–2669. [Google Scholar] [CrossRef] [PubMed]
- Ashnagar, N.; Mir Mahmoodi, T.; Jalilnejhad, N. Effects of electromagnetic field and ultrasonic waves on seed germination, seedling characteristics and essence percent of thymes (Thymus vulgaris L.). J. Crop Ecophysiol. 2019, 13, 57–72. [Google Scholar]
- Machikowa, T.; Kulrattanarak, T.; Wonprasaid, S. Effects of ultrasonic treatment on germination of synthetic sunflower seeds. Int. J. Agric. Biosyst. Eng. 2013, 7, 1–3. [Google Scholar]
- Miano, A.C.; Forti, V.A.; Abud, H.F.; Gomes-Junior, F.G.; Cicero, S.M.; Augusto, P.E.D. Effect of ultrasound technology on barley seed germination and vigour. Seed Sci. Technol. 2015, 43, 297–302. [Google Scholar] [CrossRef]
- Sharififar, A.; Nazari, M.; Asghari, H.R. Effect of ultrasonic waves on seed germination of Atriplex lentiformis, Cuminum cyminum, and Zygophyllum eurypterum. J. Appl. Res. Med. Aromat. Plants 2015, 2, 102–104. [Google Scholar] [CrossRef]
- Tobe, K.; Li, X.; Omasa, K. Seed Germination and Radicle Growth of a Halophyte, Kalidium caspicum (Chenopodiaceae). Ann. Bot. 2000, 85, 391–396. [Google Scholar] [CrossRef]
- Kang, Y.-W.; Joo, N.-M. Comparative Analysis on Phytochemical Properties, Anti-Oxidative, and Anti-Inflammatory Activities of the Different Organs of the Common Ice Plant Mesembryanthemum crystallinum L. Appl. Sci. 2023, 13, 2527. [Google Scholar] [CrossRef]
- Bouftira, I.; Abdelly, C.; Sfar, S. Characterization of cosmetic cream with Mesembryanthemum crystallinum plant extract: Influence of formulation composition on physical stability and anti-oxidant activity. Int. J. Cosmet. Sci. 2008, 30, 443–452. [Google Scholar] [CrossRef]
- Atzori, G.; de Vos, A.C.; van Rijsselberghe, M.; Vignolini, P.; Rozema, J.; Mancuso, S.; van Bodegom, P.M. Effects of increased seawater salinity irrigation on growth and quality of the edible halophyte Mesembryanthemum crystallinum L. under field conditions. Agric. Water Manag. 2017, 187, 37–46. [Google Scholar] [CrossRef]
- Carapeto, A. Levantamento etnobotânico na reserva natural do sapal de castro marim e vila real de santo antónio. In Proceedings of the Relatório final do Projeto AGRO no 800 Rede Nacional para a Conservação e Utilização das Plantas Aromáticas e Medicinais, Castro Marim, Portugal, 29 June 2006. [Google Scholar]
- Lima, A.R.; Castañeda-Loaiza, V.; Salazar, M.; Nunes, C.; Quintas, C.; Gama, F.; Pestana, M.; Correia, P.J.; Santos, T.; Varela, J.; et al. Influence of cultivation salinity in the nutritional composition, antioxidant capacity and microbial quality of Salicornia ramosissima commercially produced in soilless systems. Food Chem. 2020, 333, 127525. [Google Scholar] [CrossRef]
- Ventura, Y.; Sagi, M. Halophyte crop cultivation: The case for Salicornia and Sarcocornia. Environ. Exp. Bot. 2013, 92, 144–153. [Google Scholar] [CrossRef]
- Flamini, G.; Luigi Cioni, P.; Morelli, I.; Ceccarini, L.; Andolfi, L.; Macchia, M. Composition of the essential oil of Medicago marina L. from the coastal dunes of Tuscany, Italy. Flavour. Fragr. J. 2003, 18, 460–462. [Google Scholar] [CrossRef]
- Wang, G.; Wang, J.; Liu, W.; Nisar, M.F.; El-Esawi, M.A.; Wan, C. Biological activities and chemistry of triterpene saponins from Medicago species: An update review. Evid.-Based Complement. Altern. Med. 2021, 2021, 6617916. [Google Scholar] [CrossRef] [PubMed]
- Salem, S.; Nasri, S.; Abidi, S.; Smaoui, A.; Nasri, N.; Mutjé, P.; Ben Hamed, K. Lignocellulosic Biomass from Sabkha Native Vegetation: A New Potential Source for Fiber-Based Bioenergy and Bio-Materials. In Sabkha Ecosystems; Springer: Berlin/Heidelberg, Germany, 2019; pp. 407–412. [Google Scholar]
- Merchaoui, H.; Ben Mansour, R.; Oueslati, M.; Medini, F.; Hanana, M.; Ksouri, R. A Comparative Evaluation of Total Polyphenolic Content and Antioxidant Potential of Thirty Medicinal Halophytes from the Mediterranean Region. J. Agric. Sci. Technol. 2019, 21, 1433–1446. [Google Scholar]
- Samir, D.; Sara, C.; Widad, A. The Effects of aqueous leaf extract of Portulaca oleracea on haemato-biochemical and histopathological changes induced by Sub-chronic Aluminium toxicity in male wistar rats. Pharmacol. Res. Chin. Med. 2022, 4, 100101. [Google Scholar] [CrossRef]
- Nemzer, B.; Al-Taher, F.; Abshiru, N. Phytochemical composition and nutritional value of different plant parts in two cultivated and wild purslane (Portulaca oleracea L.) genotypes. Food Chem. 2020, 320, 126621. [Google Scholar] [CrossRef]
- Zeghib, K.; Boutlelis, D.A.; Menai, S.; Debouba, M. Protective effect of Atriplex halimus extract against benzene-induced haematotoxicity in rats. Ukr. Biochem. J. 2021, 93, 66–76. [Google Scholar] [CrossRef]
- Clauser, M.; Dall’Acqua, S.; Loi, M.C.; Innocenti, G. Phytochemical investigation on Atriplex halimus L. from Sardinia. Nat. Prod. Res. 2013, 27, 1940–1944. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Bellot, M.J.; Lorente, B.; Ortuño, M.F.; Medina, S.; Gil-Izquierdo, Á.; Bañón, S.; Sánchez-Blanco, M.J. Recycled wastewater and reverse osmosis brine use for halophytes irrigation: Differences in physiological, nutritional and hormonal responses of Crithmum maritimum and Atriplex halimus plants. Agronomy 2021, 11, 627. [Google Scholar] [CrossRef]

{kind=link}
| Substrate | Irrigation | M. crystallinum | S. ramosissima | M. marina | A. arenaria | P. oleracea | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| GR | MGT | GR | MGT | GR | MGT | GR | MGT | GR | MGT | ||
| Perlite | Freshwater | 43.3 ± 1.5 a | 6.3 a | 52.2 ± 1.5 abc | 9.2 b | - | - | - | - | 24.4 ± 1.5 d | 5.0 d |
| Saltwater | - | - | 5.6 ± 0.3 e | 9.4 ab | - | - | - | - | - | - | |
| Vermiculite | Freshwater | 35.6 ± 3.2 a | 6.0 a | 50.0 ± 1.5 abc | 12.2 ab | 11.1 ± 0.7 a | 17.4 ab | 2.2 ± 0.3 b | 29.0 a | 62.2 ± 0.7 b | 5.4 d |
| Saltwater | - | - | 14.4 ± 0.9 de | 12.6 ab | - | - | - | - | 1.1 ± 0.3 e | 13.3 ab | |
| Coco peat | Freshwater | 40.0 ± 2.1 a | 6.3 a | 57.8 ± 1.8 abc | 14.2 ab | - | - | 17.8 ± 0.9 a | 20.4 a | 95.6 ± 0.9 a | 5.2 d |
| Saltwater | - | - | 10.0 ± 1.2 de | 14.5 ab | - | - | - | - | 15.6 ± 0.7 de | 12.2 bc | |
| Sand | Freshwater | 35.6 ± 2.2 a | 10.6 a | 57.8 ± 0.9 abc | 16.7 ab | 5.6 ± 0.3 ab | 15.0 bc | - | - | 47.8 ± 1.2 bc | 5.5 d |
| Saltwater | - | - | 45.6 ± 2.0 abc | 17.1 ab | - | - | - | - | - | - | |
| Coco peat, perlite (1:1 v/v) | Freshwater | 38.9 ± 1.5 a | 6.2 a | 35.6 ± 1.7 bcd | 11.5 ab | 6.7 ± 0 ab | 21.3 a | - | - | 83.3 ± 0.6 a | 5.2 d |
| Saltwater | - | - | 31.1 ± 1.2 cde | 12.9 ab | - | - | - | - | - | - | |
| Coco peat, vermiculite (1:1 v/v) | Freshwater | 52.2 ± 2.2 a | 8.4 a | 66.7 ± 2.6 a | 13.2 ab | 2.2 ± 0.3 b | 10.5 cd | 16.7 ± 0.6 a | 22.8 a | 64.4 ± 0.9 b | 5.0 d |
| Saltwater | - | - | 33.3 ± 2.1 cde | 17.5 a | - | - | - | - | 1.1 ± 0.3 e | 19.0 a | |
| Perlite, vermiculite (1:1 v/v) | Freshwater | 45.6 ± 1.2 a | 6.0 a | 57.8 ± 0.7 abc | 10.4 ab | 4.4 ± 0.3 b | 9.5 d | 5.6 ± 0.3 b | 18.3 a | 44.4 ± 1.8 c | 6.9 cd |
| Saltwater | - | - | 50.0 ± 2.9 abc | 15.8 ab | - | - | - | - | - | - | |
| Sand, organic peat, perlite (1:1:1 v/v/v) | Freshwater | 30.0 ± 1.7 a | 7.9 a | 62.2 ± 0.9 ab | 9.9 ab | 1.1 ± 0.3 b | 9.9 d | - | - | 43.3 ± 1.2 c | 5.4 d |
| Saltwater | - | - | 55.6 ± 0.9 abc | 14.0 ab | - | - | - | - | - | - | |
| Type of Treatment | Specific Treatment | M. marina | A. arenaria | ||
|---|---|---|---|---|---|
| GR | RI | GR | RI | ||
| Chemical treatment | Gibberellic acid 1 g/L, 24 h | 35.0 ± 1.2 a | +23.9 | 26.7 ± 1.9 bcd | +8.9 |
| Chemical scarification | Sulfuric acid 50%, 10 min | 53.3 ± 1.7 a | +42.2 | 26.7 ± 1.3 abcd | +8.9 |
| Mechanical scarification | Ultrasounds, 15 min | 25.0 ± 1.2 a | +13.9 | 53.3 ± 1.2 a | +35.5 |
| Soaking | Water at RT, 24 h | 23.3 ± 0.7 a | +12.2 | 30.0 ± 1.2 abc | +12.2 |
| Thermal shock | W60 | 36.7 ± 1.3 a | +25.6 | 20.0 ± 0.6 d | +2.2 |
| W75 | 30.0 ± 1.0 a | +18.9 | 31.7 ± 1.7 abc | +13.9 | |
| W75W-4 | 38.3 ± 0.7 a | +27.2 | 53.3 ± 1.5 ab | +35.5 | |
| W-4W75 | 40.0 ± 1.7 a | +28.9 | 23.3 ± 0.9 cd | +5.5 | |
| W-4 | 26.7 ± 1.2 a | +15.6 | 1.7 ± 0.3 d | −16.1 | |
| Species | Irrigation | Substrate | Treatment |
|---|---|---|---|
| Mesembryanthemum crystallinum L. | Freshwater | Coco peat, vermiculite (1:1 v/v) | n.a. |
| Salicornia ramosissima J. Woods | Freshwater | Coco peat, vermiculite (1:1 v/v) | n.a. |
| Medicago marina L. | Freshwater | Vermiculite | Sulfuric acid 50%, 10 min |
| Ammophila arenaria (L.) Link | Freshwater | Coco peat | Ultrasounds, 15 min |
| Portulaca oleracea L. | Freshwater | Coco peat | n.a. |
| Atriplex halimus L. | No germination | No germination | No improvement |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castañeda-Loaiza, V.; Rodrigues, M.J.; Fernandes, E.; Custódio, L. A Comparative Study of the Influence of Soil and Non-Soil Factors on Seed Germination of Edible Salt-Tolerant Species. Horticulturae 2024, 10, 872. https://doi.org/10.3390/horticulturae10080872
Castañeda-Loaiza V, Rodrigues MJ, Fernandes E, Custódio L. A Comparative Study of the Influence of Soil and Non-Soil Factors on Seed Germination of Edible Salt-Tolerant Species. Horticulturae. 2024; 10(8):872. https://doi.org/10.3390/horticulturae10080872
Chicago/Turabian StyleCastañeda-Loaiza, Viana, Maria João Rodrigues, Eliana Fernandes, and Luísa Custódio. 2024. "A Comparative Study of the Influence of Soil and Non-Soil Factors on Seed Germination of Edible Salt-Tolerant Species" Horticulturae 10, no. 8: 872. https://doi.org/10.3390/horticulturae10080872