Abstract
Litchi downy blight (LDB) caused by Peronophythora litchii destroys 20–30% of litchi fruit every year and causes significant economic losses. Some Exiguobacterium strains exhibit considerable promise in both agricultural and industrial sectors. E. acetylicum SI17, isolated from the litchi fruit carposphere, demonstrated significant biocontrol activity against LDB through pre-harvest treatment. To elucidate its underlying regulatory mechanisms, the genome of SI17 was sequenced and analyzed, revealing a circular chromosome spanning 3,157,929 bp and containing 3541 protein-coding genes and 101 RNA genes. Notably, 94 genes were implicated in the production of secondary metabolites. Among the 29 Exiguobacterium strains so far sequenced, SI17 possessed the largest genome. In the phylogenomic analysis encompassing the entire genome, SI17 was clustered into Group I. Treating litchi fruit with SI17 before harvesting resulted in a decrease in H2O2 content in the fruit peel and an increase in superoxide dismutase activity, thus enhancing resistance to LDB. Interestingly, SI17 did not display plate antagonism against Peronophythora litchii SC18. It can be inferred that SI17 generates secondary metabolites, which enhance litchi’s resistance to LDB. This study represents the first documentation of an Exiguobacterium strain exhibiting a role in litchi plant disease and showcasing significant potential for the biological control of LDB.
1. Introduction
The litchi fruit (Litchi chinensis) belongs to the Sapindaceae family and is typically found in subtropical areas of Asia, South Africa, Australia, the United States (especially Hawaii), and Israel [1]. Originating from South China, it is predominantly cultivated in the southern provinces of Fujian, Guangdong, and Hainan. Unfortunately, harvested litchi fruit are highly susceptible to spoilage due to fungal diseases caused by pathogens such as Peronophythora litchii, Geotrichum candidum, and Colletotrichum gloeosporiodes, resulting in a 20–30% loss. Among these, litchi downy blight (LDB), caused by P. litchii, is a significant issue [2]. P. litchii infects litchi tender leaves, flowers, and mature fruits in the field and post-harvest storage. This hemibiotrophic, homothallic, and polycyclic oomycete produces oospores spherical in shape with smooth walls and a light-yellow color. Oospores can survive in the soil or plant debris for several years and germinate directly or indirectly under favorable conditions. Lemon-shaped deciduous sporangia with papillae at the apex are also produced. Sporangium, wind or water dispersal, germinates directly, producing germ tubes and penetration hyphae able to penetrate the receptive organs. Alternatively, sporangia can form secondary sporangia or release zoospores. Zoospores, kidney shaped, have two lateral flagella, and rapidly germinate in cool, wet conditions. P. litchii causes withering and watery brown spots on the infection sites of tender leaves or fruit and produces downy white sporangiophores, which trigger pre- and post-harvest fruit decay [3]. LDB also affects the growth and development of young leaves, although it is most detrimental during the period from bloom to fruit ripening, particularly in extremely humid and rainy conditions [4]. At present, the prevailing methods for managing LDB largely rely on post-harvest applications of chemical fungicides (like dimethomorph and propamocarb). Regrettably, these residues may persist in fruits, posing carcinogenic hazards to consumers and contributing to environmental pollution [5]. Hence, it is crucial and urgent to explore alternative techniques like biological control to minimize post-harvest decay of litchi fruit. This method provides safe, environmentally friendly, and sustainable results, in line with the goals of global strategic initiatives for eco-conscious and/or organic farming practices [6].
In recent years, there has been rapid progress in utilizing biological control agents (BCAs) to manage post-harvest diseases [7]. These applications have exhibited effectiveness in suppressing P. litchii and preventing decay in litchi fruit. Among the BCAs employed, there are various antagonistic bacteria like Bacillus subtilis [1,8,9], endophytic strains such as B. amyloliquefaciens TB2 and LY-1 [10,11], and Lactobacillus plantarum LAB [12]. Additionally, certain metabolites have demonstrated antagonistic effects against P. litchii, including hypothemycin from Paecilomyces sp. SC0924 [2], zeamines from Dickeya zeae [13], and 4-ethylphenol from Streptomyces fimicarius BWL-H1 [14]. The primary mechanisms through which BCAs inhibit diseases include the synthesis of antibiotics, release of extracellular enzymes such as protease, chitinase, cellulase, and β-1,3-glucanase, production of siderophores, and generation of indole acetic acid (IAA). They also occupy ecological niches, enhance microbial diversity in the phyllosphere and carposphere [15], and prompt plant defense responses such as systemic acquired resistance, induced systemic resistance, and/or priming [16].
Exiguobacterium exhibits significant promise for utilization across various sectors like industry and agriculture, owing to the presence of certain strains that possess diverse functionalities such as enzyme production, bioremediation, degradation of toxic substances, and facilitation of plant growth [17]. For instance, E. sp. VSG-1 produces hydrolytic enzymes like cellulase, pectinase, mannanase, xylanase, and tannase, which bolster the conversion of fermented sugar from sugarcane bagasse into bioethanol [18]. Additionally, E. profundum, known for its protease production ability, has been employed in the extraction of chitin from shrimp shells [19]. Furthermore, various other hydrolytic enzymes like lipase, amylase, and pullulanase have been isolated from different Exiguobacterium strains, further expanding their potential applications [20,21,22,23]. E. alkaliphilum B-3531D, E. sp. AO-11, and E. mexicanum M7 exhibit the capacity to metabolize crude oil, suggesting promise for the development of bioremediation solutions for oil-contaminated marine ecosystems [24,25,26]. Additionally, four Exiguobacterium isolates have demonstrated the ability to reduce Cr (VI) levels in polluted environments [27,28,29,30]. Moreover, Exiguobacterium exhibits significant potential for agricultural usage, with several strains possessing traits that promote plant growth across various crops [31,32,33,34,35]. While disease control is vital in agriculture, there have been no reported applications of Exiguobacterium in plant disease management thus far.
The genus Exiguobacterium includes species and strains engaged in industry and agriculture, comprising enzyme production, bioremediation, degradation of toxic substances, and plant growth-promoting properties [17]. Cellulase, pectinase, mannanase, xylanase, and tannase produced by E. sp. VSG-1 make steam-exploded sugarcane bagasse useful to biofuel fabrication by Saccharomyces cerevisiae fermentation [18]. A protease-producing strain of E. profundum has been employed in the extraction of chitin from shrimp shells [19]. Lipase, amylase, and pullulanase have been isolated from different Exiguobacterium strains, increasing their potential applications [20,21,22,23]. Exiguobacterium sp. AO-11, E. alkaliphilum B-3531D, and E. mexicanum M7 metabolize crude oil, suggesting promise for the bioremediation of oil-contaminated marine ecosystems [24,25,26]. Exiguobacterium strains ZM-2, GS1, PY14, and E. mexicanum from chromite mines reduce Cr (VI) levels in polluted environments [27,28,29,30]. Moreover, several Exiguobacterium strains exhibit significant potential for agricultural applications as plant growth promoting bacteria [31,32,33,34,35]. Exiguobacterium has been reported to suppress fungal diseases of cereal crops in Australia [36], and E. acetylicum strain 1P MTCC 8707 inhibited the in vitro growth of Rhizoctonia solani, Sclerotium rolfsii, Pythium sp., and Fusarium oxysporum [37]. No applications of Exiguobacterium in pre- and post-harvest plant disease management have been reported thus far.
The genomes of several Exiguobacterium strains have been sequenced. Based on the concatenated single-copy core genes and 16S rRNA gene sequences, the Exiguobacterium strains were clustered into two groups. Group I comprised strains capable of thriving at temperatures of 7 ℃ or lower, while Group II encompassed the remaining strains [38,39]. The proliferation of transporters, crucial for conveying vital substrates and conferring resilience against environmental stresses, was a key factor propelling the genome expansion in Group I strains, thus broadening their ecological niche [39]. Additionally, genome sequencing enables us to elucidate the agricultural utility of these strains.
E. acetylicum SI17, sourced from the carposphere of litchi fruit, has demonstrated efficacy in combating LDB through biocontrol measures [40]. The volatile organic compounds (VOCs) emitted by SI17, particularly those containing α-Farnesene (AF), exhibited the ability to hinder the growth of P. litchii, thus alleviating the severity of LDB [40]. The purpose of the study was to gain deeper insights into the attributes of SI17. We carried out genome sequencing and comparative analysis of its genome.
2. Materials and Methods
2.1. E. acetylicum SI17 Bioinformatics Analysis
2.1.1. Bacterial Culture and DNA Extraction
E. acetylicum SI17 was isolated from the interior of litchi fruit sarcocarp. The cells of E. acetylicum SI17 were cultured in a 500 mL flask containing 100 mL of Luria–Bertani (LB) medium (tryptone 10 g/L, yeast extract 5 g/L, and NaCl 10 g/L) at 28 °C with shaking at 200 rpm for 24 h. Subsequently, the cells were harvested at 4 °C through centrifugation at 10,000× g for 5 min. Genomic DNA extraction was carried out immediately using the method outlined by Hoffman and Winston [41]. The purity and integrity of the DNA were evaluated by agarose gel electrophoresis, and its concentration was measured using a Qubit™ fluorometer (Invitrogen, Carlsbad, CA, USA).
2.1.2. Genome Sequencing, Assembly, Annotation, and Phylogenomic Analyses
The genome of E. acetylicum SI17 was sequenced, assembled, and annotated following the methodology outlined by Zheng et al. [42]. The genome was sequenced on the Nanopore PromethION. Further, twenty-eight complete amino acid sequences of various Exiguobacterium strains were retrieved from the NCBI database (https://www.ncbi.nlm.nih.gov/genome/ (accessed on 24 May 2021)). A phylogenomic tree, based on whole genomes, was then constructed using CVTree3 [43], employing Bacillus subtilis as the out-group.
2.2. Biocontrol (Antagonistic) Activity on Litchi Fruit
2.2.1. Inoculation Experiments on Litchi Fruit
The pathogen P. litchii SC18 and the biocontrol agent E. acetylicum SI17 were cultured using the method described by Zheng et al. [40]. The pathogen was cultured on carrot agar (CA) medium for 7 days. “Feizixiao” litchi fruits, approximately 60% ripe (about 65 days after flowering), were treated by spraying with E. acetylicum SI17 suspension at a concentration of 5 × 107 CFU/mL or a 1/10 dilution of LB broth (as control) until completely covered. After seven days, the harvested fruits were moved to a greenhouse maintained at a steady temperature of 25 °C, 95% humidity, with 12 h day and night cycles. Twenty-four hours later, the treatments were sprayed with 30 mL of P. litchii SC18 at a concentration of 5 × 104 sporangia/mL. Disease severity was measured every 12 h from 60 to 96 hpi. Each treatment contained 3 replicates, with 30 fruits per replicate. The disease index was calculated following the procedure outlined by Zheng et al. [40].
2.2.2. Relative Quantification of P. litchii SC18 in Pericarp
Total RNA was extracted from the litchi pericarp by using the RNAprep Pure Plant Kit (Tiangen, Beijing, China). The purity and integrity of the RNA were assessed using agarose gel electrophoresis and the Agilent 2100 system (Agilent Technologies, Santa Clara, CA, USA), while the RNA concentration was measured with the Nanodrop (Thermo Scientific, Waltham, CA, USA) and Qubit 2.0 (Life Technologies, Carlsbad, CA, USA) systems. For cDNA synthesis, one microgram of RNA was used with the PrimeScriptTM RT Reagent Kit with gDNA Eraser (Takara, Kusatsu, Japan). The qRT-PCR assay was carried out using TB GreenTM Premix Ex TaqTM Ⅱ (Takara, Kusatsu, Japan), and the data were obtained using the Thermal CyclerTM Dice Real Time System III (Takara, Kusatsu, Japan). The relative expression levels were determined by calculating the fold change based on the 2−△△Ct method. The primers were designed with Primer 3.0 (ABI, Carlsbad, CA, USA), and the sequences were as follows: P. litchii Actin-F: ACATTGCCCTGGACTTCG, P. litchii Actin-R: AGCTCCTTGGTCATACGC, litchi Actin-F: CGGGAAATTGTCCGTGAC, litchi Actin-R: GAGGACTTCTGGGCAACG. P. litchii Actin was quantified using litchi Actin as a reference gene, and the results indicated the relative amount of P. litchii SC18 in the litchi peel.
2.2.3. Enzyme Activity in the Pericarp
In 2021, samples were collected from Litchi cv. “Feizixiao” fruits. Nine fruits were taken for each treatment at each time point (0, 48, 84, 96, 108, and 120 h post-inoculation (hpi)). The pericarp was then collected, promptly enveloped in tin foil, flash-frozen in liquid nitrogen, and stored at −80 °C until required for experimental analyses. The levels of H2O2 and the activities of the enzymes catalase (CAT, EC 1.11.1.6), superoxide dismutase (SOD, EC 1.15.1.1), and peroxidase (POD, EC 1.11.1.7) were meticulously measured. The detection kits for these analyses were obtained from Nanjing Jiancheng Biological Engineering Institute, Nanjing, China (http://www.njjcbio.com/ (accessed on 18 November 2021)).
2.3. Data Analysis
All statistical analyses were performed using SPSS 25.0 (IBM, Armonk, NY, USA). Significant differences between more than two groups were determined using one-way ANOVA and Duncan’s multiple comparison test. Statistical significance was set at p < 0.05. Histograms were constructed using Microsoft Office Excel 2016 (Microsoft, Los Angeles, CA, USA).
3. Results
3.1. Genome and Phylogenomic Analysis of E. acetylicum SI17
3.1.1. Genome Sequencing, Assembly, and Functional Annotation
The data gathered from the Nanopore platform were used to construct a circular between chromosomes, with a length of 3,157,929 base pairs (bp) and a GC content of 47.28% (Figure 1, Table 1). Additionally, four plasmids were detected, measuring 119,838 bp, 43790 bp, 6694 bp, and 68,355 bp in length, with GC contents of 40.05%, 43.16%, 37.50%, and 37.92%, respectively (Figure 1, Table 1). The genome comprises 3541 predicted protein-coding sequences (CDS), accounting for 88.98% of the total genomic content. These sequences possess an average length of 846 bp, as shown in Table 1. Analysis of the genome components revealed the presence of 154 interspersed repeats, 105 tandem repeats (TR), 70 tRNA genes, 27 rRNA genes, 1 sRNA gene, 3 other RNA genes, 6 genomic islands (GIs), and 5 prophages. Notably, no clustered regularly interspaced short palindromic repeats (CRISPR) sequences were identified, as specified in Table 2.
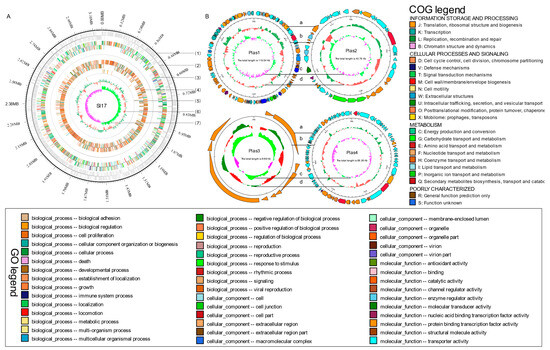
Figure 1.
Map of chromosome and four plasmids of Exiguobacterium acetylicum SI17. (A) chromosome, from outside to inside, the map shows the (1) position of the genome, (2) coding genes on the + and –, strands, (3) COG annotation, (4) GO annotation, (5) non-coding RNA, and (6) GC content. The outer green indicates that the GC content in this region is higher than the genome-wide average, while the inner red indicates the opposite. The height of the peak represents the magnitude of the difference from the average GC content. Higher peaks indicate greater differences from the average GC content. (7) GC skew value (G–C/G + C). When the value is positive, there is a high possibility that CDS is transcribed from the positive chain; Otherwise, there is a high possibility that CDS is transcribed from the negative chain. (B) plasmids, from outside to inside, the map shows the (a) COG annotation, (b) position of the genome, (c) GC content, (d) GC skew value.

Table 1.
Genomic features of strain SI17.
Table 2.
Genome component analyses of the strain SI17.
Functional annotation was conducted by aligning the predicted gene protein sequences using the diamond alignment tool against several frequently used databases. In the context of Clusters of Orthologous Groups (COG) analysis, a total of 2574 proteins were sorted into 23 distinct functional groups. Apart from the general function prediction category, both translation, ribosomal structure, and biogenesis, as well as the transcription category, exhibited a higher percentage among the 22 categories, encompassing 237 and 230 genes, respectively (Figure S1). In the Gene Ontology (GO) analysis, the 2409 proteins were categorized into three primary groups: biological processes, molecular functions, and cellular components. Interestingly, categories of metabolic processes and cellular processes, catalytic activity and binding, as well as cell part and cell, demonstrated higher gene abundance (Figure S2). Moreover, in the context of Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, the metabolism pathway emerged as particularly notable (Figure S3). The Carbohydrate-Active enZYmes (CAZy) database, dedicated to carbohydrate enzymes, covers a broad spectrum of enzyme families pivotal in catalyzing the degradation, alteration, and biosynthesis of carbohydrates. It features five main groups: glycoside hydrolase (GH), glycosyl transferase (GT), polysaccharide lyase (PL), carbohydrate esterase (CE), and auxiliary activity (AA). Notably, there were 45, 36, 1, 5, 0, and 27 genes categorized under GH, GT, PL, CE, AA, and CBM (carbohydrate-binding module), respectively (Figure S4). Additionally, the Transporter Classification Database (TCDB) analysis (Figure S5) showed that genes classified as porters, including uniporters, symporters, and antiporters (94 genes), along with P-P-bond-hydrolysis-driven transporters (129 genes), collectively represented 74.33% of all predicted transporters (300 genes).
3.1.2. Phylogenomic and Comparative Genomic Analysis
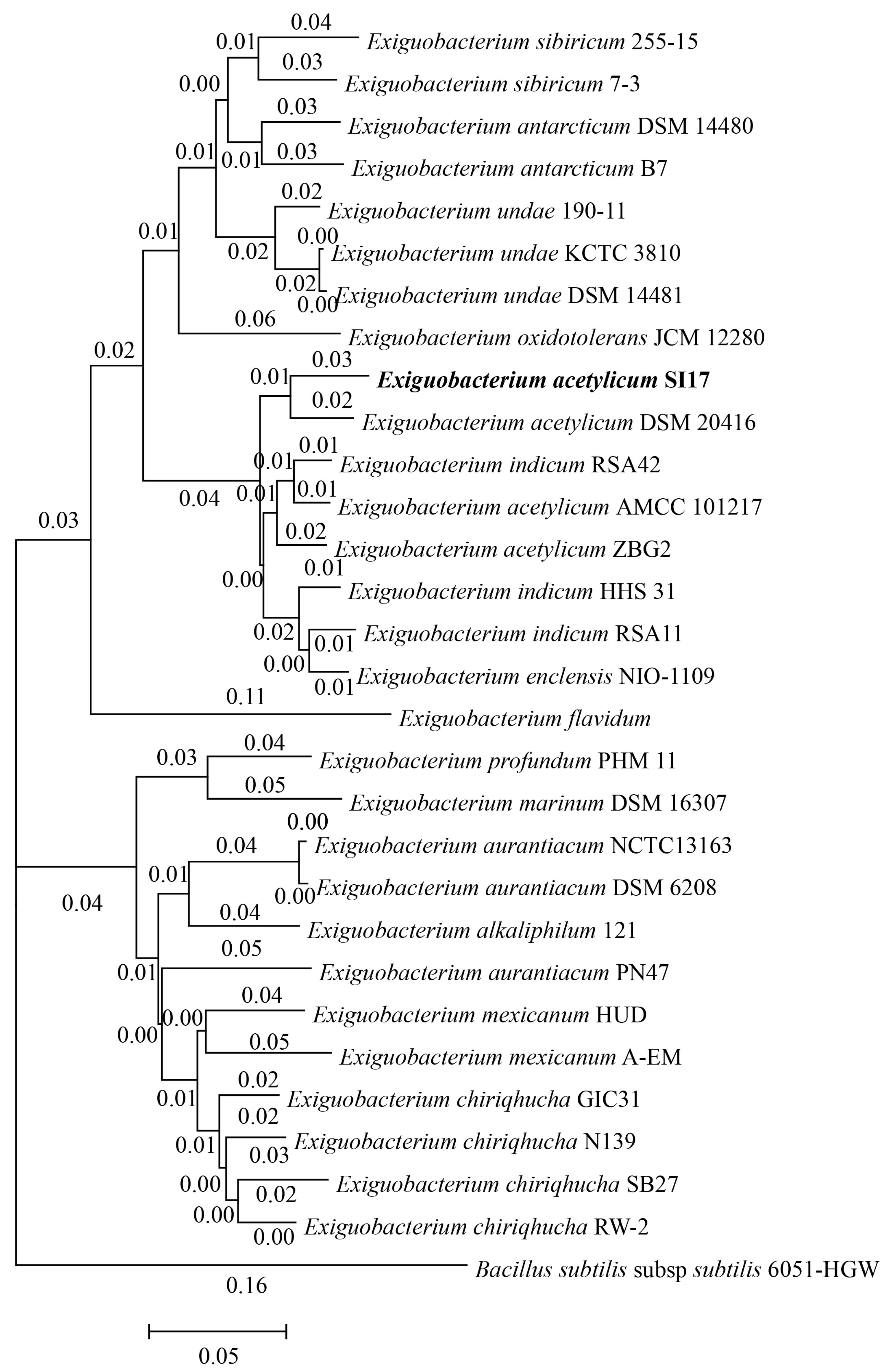
The 29 Exiguobacterium genomes were divided into two groups. Notably, E. acetylicum SI17 was clustered in the first group, showing greater homology to E. indicum and E. enclensis (Figure 2). Despite differences in GC content, there were no significant variations in genome size, gene count, proteins, rRNAs, or tRNAs between the two groups (Table S1). To the best of our knowledge, this is the first documented instance of using Exiguobacterium strains for controlling plant diseases. This suggests that the Group I strains may have potential applications in agriculture.
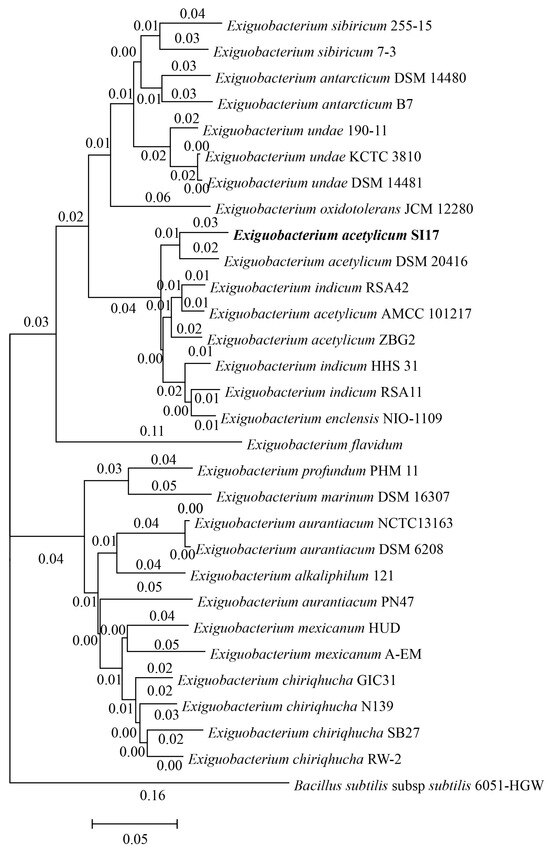
Figure 2.
Phylogenomic tree based on whole-genome sequences of 29 Exiguobacterium strains. Bacillus subtilis was used as an out-group. Strain E. acetylicum SI17 is highlighted in bold.
3.2. Biocontrol Activity of E. acetylicum SI17 on LDB
3.2.1. Pre-Harvest E. acetylicum SI17 Treatment Suppressed LDB
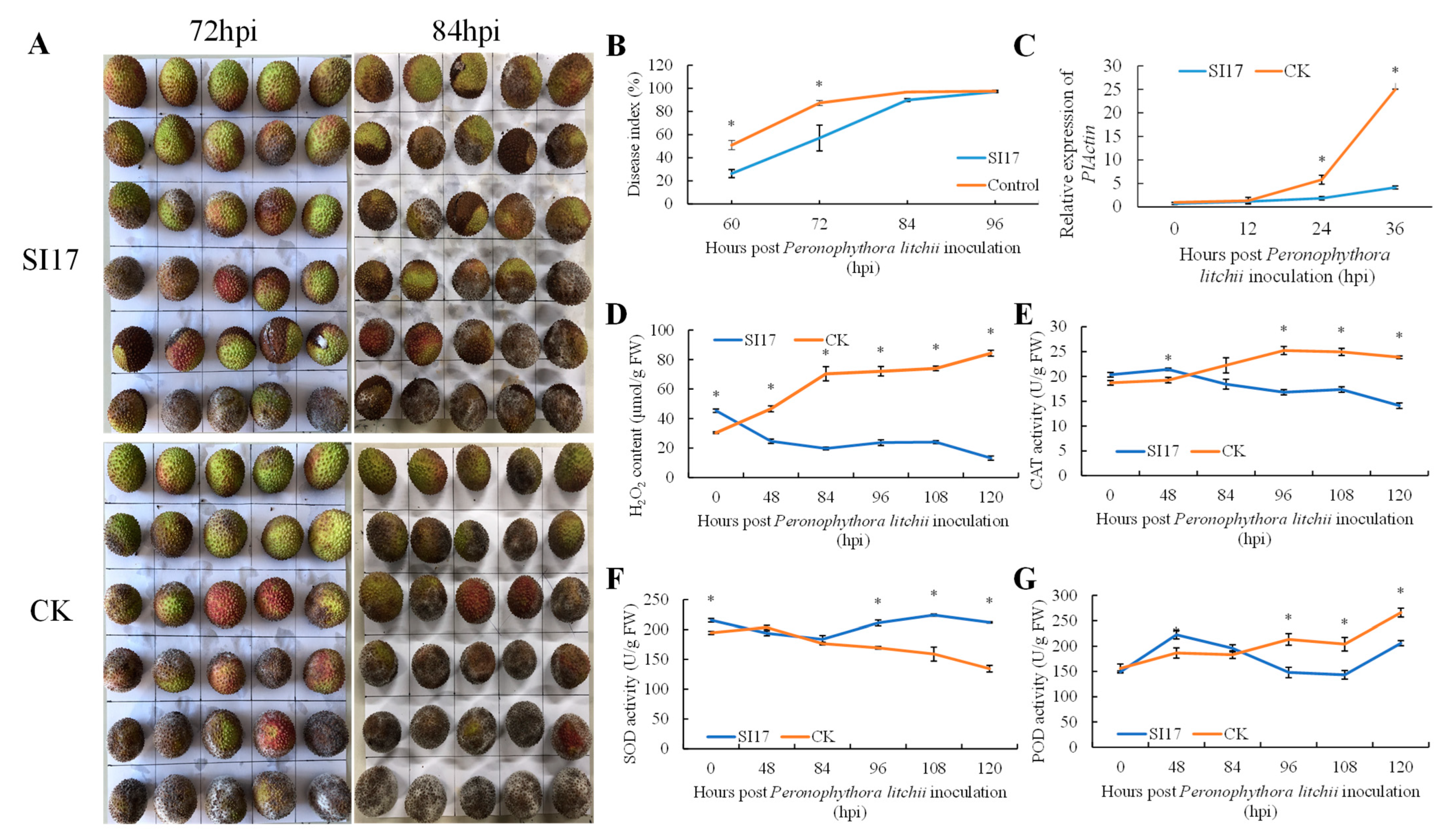
Pre-harvest inoculation experiments on litchi fruit indicated that E. acetylicum SI17-treated fruits had a significantly lower disease index at 60, 72, and 84 h, but this difference lessened over time (Figure 3A,B).
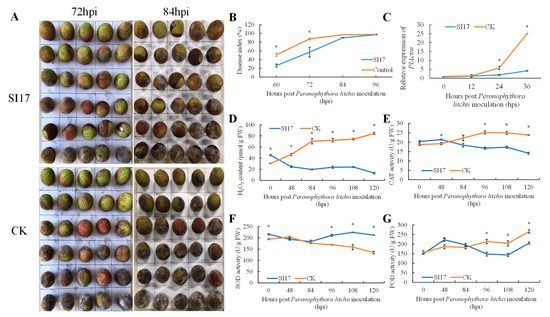
Figure 3.
Interaction between Exiguobacterium acetylicum strain SI17 pre-harvest treatment and Peronophythora litchi on litchi cv. “Feizixiao” fruits: (A) litchi downy blight symptoms and (B) disease index (B) development, (C) PlActin expression, (D) H2O2 content, (E) catalase (CAT), (F) superoxide dismutase (SOD), (G) peroxidase (POD) activities. CK = control. The * above bars indicate significant differences at p < 0.05 in Duncan’s multiple range test.
3.2.2. Relative Quantification of P. litchii SC18 in Pericarp after E. acetylicum SI17 Treatment
Following the treatment with P. litchii SC18, the relative quantity of pathogens was measured using qRT-PCR. In both the E. acetylicum SI17 treatment and control groups, relative pathogen levels were low before 12 hpi (Figure 3C). By 24 hpi, pathogen levels in the E. acetylicum SI17 treatment group were three times lower than those in the control group, and by 36 hpi, they were six times lower (Figure 3C). As time progressed, the pathogen growth rate in the control group was much higher than that in the E. acetylicum SI17 treatment group.
3.2.3. Pre-Harvest E. acetylicum SI17 Treatment Enhanced the Activity of Defense-Related Enzymes
After inoculation with P. litchii, H2O2 levels in the E. acetylicum SI17-treated litchi pericarp steadily declined, unlike the control group, which showed a rapid H2O2 buildup before 84 h, followed by a gradual increase. Overall, H2O2 levels in the E. acetylicum SI17-treated litchi pericarp were lower compared to the control (Figure 3D). In the treatment group, the activity of SOD initially decreased before 84 h and then peaked at 108 h. Conversely, in the control group, SOD activity continued to decline over time (Figure 3F). CAT activity displayed an upward trend in the control group, whereas in the E. acetylicum SI17-treated litchi pericarp, it showed a downward trend (Figure 3E). Prior to 84 h, CAT activity in the E. acetylicum SI17 treatment group surpassed that of the control group but subsequently declined (Figure 3E). POD activity in the control group increased steadily over time, whereas in the E. acetylicum SI17 treatment group, it initially rose, then declined before rising again, peaking at 48 h and reaching its lowest point at 108 h (Figure 3F).
4. Discussion
E. acetylicum has been observed to enhance wheat seedling growth [32] and break down shrimp shell waste [44]. Yet, its impact on plant diseases remains unexplored until now. Our research reveals that E. acetylicum SI17 demonstrates efficacy in managing LDB. By sequencing the whole genome of E. acetylicum SI17, we identified the genetic basis for this functional trait. Compared to the other 28 Exiguobacterium genomes, E. acetylicum SI17 stands out with the largest genome size and the highest counts of genes, proteins, and tRNAs (Table S1). Exiguobacterium is clustered into two groups based on its ability to withstand temperatures as low as 7 °C, yet there was no significant difference in functionality between these groups. E. oxidotolerans (Group I) and E. profundum PHM11 (Group II) demonstrated the capacity to enhance the growth and yield of Bacopa monnieri and rice, respectively, particularly under conditions of salt stress [33,45]. Additionally, E. acetylicum (Group I) and E. marinum a-1 (Group II) exhibited capabilities in degrading harmful substances, such as cyanide-containing effluents and polypropylene, respectively [46,47]. This suggests that not only species within Group I but also those within Group II possess the ability to manage plant diseases.
Existing research has shown that using post-harvest E. acetylicum SI17 treatment is highly effective in controlling LDB. Additionally, exposing litchi fruits to SI17 VOCs before harvest significantly decreases the severity of LDB [40]. In this study, we examined the effectiveness of pre-harvest E. acetylicum SI17 treatment in controlling LDB to explore its potential for field application. Our findings indicate promising results (Figure 3A,B). H2O2, as a representative of reactive oxygen species (ROS), can be excessively generated in response to various biotic and abiotic stresses, resulting in its heightened reactivity and toxic effects on plants [48]. The accumulation of ROS is a widespread defense mechanism for higher plants to resist pathogen attacks. However, excessive ROS can cause plant damage and facilitate pathogen infection [49]. SOD, CAT, and POD are essential enzymes that act as antioxidants, responsible for neutralizing ROS [50]. Many studies have shown that biocontrol agents can effectively clear excess ROS that accumulate due to pathogen induction. For example, Bacillus velezensis could enhance the activity of ROS-scavenging enzymes, thereby enhancing the eggplant fruits’ disease resistance [51]. A similar situation occurred in Bacillus amyloliquefaciens GJ1 against citrus Huanglongbing [52] and Streptomyces griseorubiginosus LJS06 against cucumber anthracnose [53]. Litchi pre-treated with SI17 demonstrated an increased ability to scavenge H2O2 (Figure 3D), primarily due to heightened SOD activity rather than CAT and POD activities (Figure 3E–G).
Fermentation (C), suspension (F), and supernatant (Q) of E. acetylicum SI17 demonstrated no inhibitory effect on the growth of P. litchii SC18 (Figure S6). Therefore, it is speculated that E. acetylicum SI17 may control disease by enhancing plant resistance rather than directly suppressing P. litchii SC18 growth. This is one way for a biocontrol agent to resist disease. Biocontrol agent Pseudomonas chlororaphis PA23 could induce systemic acquired resistance against Sclerotinia sclerotiorum of Brassica napus [54]. Furthermore, various strains of Exiguobacterium have been found to produce diverse hydrolytic enzymes [17]. Genome analysis provided additional insights into the key features of E. acetylicum SI17 in its ability to combat LDB. Within this strain, four gene clusters, comprising a total of 94 genes, were identified to be linked with the biosynthesis of secondary metabolites (Table S2). These clusters comprised one non-ribosomal peptide synthetase (NRPS), one siderophore, and two terpenes (Table S2). Various metabolites produced by biocontrol agents showed disease resistance in plants. Phenazine-1-carboxylic acid produced by Pseudomonas chlororaphis YL-1 showed effectiveness against Acidovorax citrulli [55]. Nep1-like proteins generated by Pythium oligandrum enhanced plant resistance against Phytophthora pathogens and Sclerotinia sclerotiorum [56]. Hence, it can be deduced that E. acetylicum SI17 could enhance plant resistance through the production of diverse secondary metabolites, although the specific functionalities of these metabolites necessitate further verification.
5. Conclusions
The Exiguobacterium acetylicum SI17 exhibited a biocontrol effect on LDB caused by P. litchii. This study marks the initial documentation of an Exiguobacterium strain being employed for managing plant diseases. To deepen our comprehension of its regulatory mechanism, we conducted a full genome sequencing of E. acetylicum SI17, revealing a larger genome size with high numbers of genes, proteins, and tRNAs. Additionally, the genome was clustered into Group I. Litchi fruit pre-treated with E. acetylicum SI17 showed increased SOD activity and improved capacity to scavenge H2O2. Pre-harvest treatment with E. acetylicum SI17 suppressed LDB, suggesting promising prospects for its practical application. These findings lead to the hypothesis that E. acetylicum SI17 might generate a metabolite that enhances litchi’s resistance, highlighting the need for more detailed exploration and identification.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae10080888/s1, Table S1: Characteristic of Exiguobacterium genomes used for phylogenomic analysis; Table S2: Secondary metabolism gene clusters in E. acetylicum SI17; Figure S1: Clusters of Orthologous Groups (COG) function classifications. B: Chromatin structure and dynamics; C: Energy production and conversion; D: Cell cycle control, cell division, chromosome partitioning; E: Amino acid transport and metabolism; F: Nucleotide transport and metabolism; G: Carbohydrate transport and metabolism; H: Coenzyme transport and metabolism; I: Lipid transport and metabolism; J: Translation, ribosomal structure and biogenesis; K: Transcription; L: Replication, recombination and repair; M: Cell wall/membrane/envelope biogenesis; N: Cell motility; O: Posttranslational modification, protein turnover, chaperones; P: Inorganic ion transport and metabolism; Q: Secondary metabolites biosynthesis, transport and catabolism; R: General function prediction only; S: Function unknown; T: Signal transduction mechanisms; U: Intracellular trafficking, secretion, and vesicular transport; V: Defense mechanisms; W: Extracellular structures; X: Mobilome, prophages, transposons; Figure S2: Gene Ontology (GO) analysis. Biological process (from left to right): biological adhesion, biological regulation, cell proliferation, cellular component organization or biogenesis, cellular process, death, developmental process, establishment of localization, growth, immune system process, localization, locomotion, metabolic process, multi-organism process, multicellular organismal process; negative regulation of biological process, positive regulation of biological process, regulation of biological process, reproduction, reproductive process, response to stimulus; rhythmic process, signaling; viral reproduction; cellular component (from left to right): cell, cell junction, cell part, extracellular region, extracellular region part, macromolecular complex, membrane-enclosed lumen, organelle, organelle part, virion, virion part; molecular function (from left to right): Antioxidant activity, binding, catalytic activity, channel regulator activity, enzyme regulator activity, molecular transducer activity, nucleic acid binding transcription factor activity, protein binding transcription factor activity, structural molecule activity, transporter activity; Figure S3: Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway annotation. Cellular Processes (A): Transport and catabolism (1), Cellular community-prokaryotes (2), cell motility (3), cell growth and death (4); Environmental Information Processing (B): Signal transduction (5), Membrane transport (6); Genetic Information Processing (C): Translation (7), Transcription (8), Replication and repair (9), Folding, sorting and degradation (10); Human Diseases (D): Substance dependence (11), Neurodegenerative diseases (12), Infectious diseases (13), Immune diseases (14), Endocrine and metabolic diseases (15), Drug resistance (16), Cardiovascular diseases (17), Cancers (18); Metabolism (E): Xenobiotics biodegradation and metabolism (19), Nucleotide metabolism (20), Metabolism of terpenoids and polyketides (21), Metabolism of other amino acids (22), Metabolism of cofactors and vitamins (23), Lipid metabolism (24), Glycan biosynthesis and metabolism (25), Energy metabolism (26), Carbohydrate metabolism (27), Biosynthesis of other secondary metabolites (28), Amino acid metabolism (29); Organismal Systems (F): Nervous system (30), Immune system (31), Excretory system (32), Environmental adaptation (33), Endocrine system (34), Digestive system (35), Aging (36); Figure S4: Carbohydrate-Active enZYmes Database (CAZy) annotation. AA: auxiliary activity; CBM: carbohydrate-binding module; CE: carbohydrate esterase; GH: glycoside hydrolase; GT: glycosyl transferase; PL: polysaccharide lyase; Figure S5: Transporter Classification Database (TCDB) annotation. 1.A: alpha-Type Channels; 1.B: beta-Barrel Porins; 1.C: Pore-Forming Toxins (Proteins and Peptides); 1.E: Holins; 2.A: Porters (uniporters, symporters, antiporters); 3.A: P-P-bond-hydrolysis-driven transporters; 3.D: Oxidoreduction-driven transporters; 3.E: Light absorption- driven transporters; 4.A: Phosphotransfer-driven Group Translocators; 4.C: Acyl CoA ligase-coupled transporters; 5.A: Transmembrane 2-electron transfer carriers; 8.A: Auxiliary transport proteins; 9.A: Recognized transporters of unknown biochemical mechanism; 9.B: Putative transport proteins; Figure S6: Plate antagonism against Peronophythora litchii SC18 of fermentation (C), suspension (F), and supernatant (Q) of SI17.
Author Contributions
S.H.: data curation, methodology, investigation, writing—original draft, writing—review and editing; X.L.: data curation, methodology, supervision, writing—original draft; L.Z.: conceptualization, methodology, supervision, validation, funding acquisition, writing—review and editing; D.G.: conceptualization, data curation, formal analysis, funding acquisition, project administration, supervision, validation, writing—original draft, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by grants from Guangdong Basic and Applied Basic Research Foundation (2022A1515010470), Key Laboratory of Biology and Genetic Resources Utilization of South Tropical Fruit Trees, Ministry of Agriculture and Rural Affairs (202102), the Innovative team program of the Department of Education of Guangdong Province (2023KCXTD018) and Guangdong Province Rural Revitalization Strategic Project “Orchard Agricultural Machinery and Agronomy Integration and Production Management Informatization” (2024-TS-2-4).
Data Availability Statement
The Genbank accession number of Exiguobacterium acetylicum SI17 genome and four plastid sequences are CP075897.1-CP075901.1. All other datasets for this study are included in the article/Supplementary Materials.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jiang, Y.M.; Zhu, X.R.; Li, Y.B. Postharvest control of litchi fruit rot by Bacillus subtilis. Food Sci. Tech. 2001, 34, 430–436. [Google Scholar] [CrossRef]
- Xu, L.; Xue, J.; Wu, P.; Wang, D.; Lin, L.; Jiang, Y.; Duan, X.; Wei, X. Antifungal activity of hypothemycin against Peronophythora litchii in vitro and in vivo. J. Agric. Food Chem. 2013, 61, 10091–10095. [Google Scholar] [CrossRef] [PubMed]
- Situ, J.; Xi, P.; Lin, L.; Huang, W.; Song, Y.; Jiang, Z.; Kong, G. Signal and regulatory mechanisms involved in spore development of Phytophthora and Peronophythora. Front. Microbiol. 2022, 13, 984672. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, T.; Liu, G.; Hu, M.; Yun, Z.; Duan, X.; Cai, K.; Jiang, G. Inhibition of downy blight and enhancement of resistance in litchi fruit by postharvest application of melatonin. Food Chem. 2021, 347, 129009. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.M.; Wang, Y.; Song, L.; Liu, H.; Lichter, A.; Kerdchoechuen, O.; Joyce, D.C.; Shi, J. Postharvest characteristics and handling of litchi fruit-an overview. Aust. J. Exp. Agric. 2006, 46, 1541. [Google Scholar] [CrossRef]
- Dukare, A.S.; Paul, S.; Nambi, V.E.; Gupta, R.K.; Singh, R.; Sharma, K.; Vishwakarma, R.K. Exploitation of microbial antagonists for the control of postharvest diseases of fruits: A review. Crit. Rev. Food Sci. 2019, 9, 1498–1513. [Google Scholar] [CrossRef]
- Ippolito, A.; Nigro, F. Impact of preharvest application of biological control agents on postharvest diseases of fresh fruits and vegetables. Crop Prot. 2000, 19, 715–723. [Google Scholar] [CrossRef]
- Sivakumar, D.; Zeeman, K.; Korsten, L. Effect of a biocontrol agent (Bacillus subtilis) and modified atmosphere packaging on postharvest decay control and quality retention of litchi during storage. Phytoparasitica 2007, 35, 507–518. [Google Scholar] [CrossRef]
- Sivakumar, D.; Arrebola, E.; Korsten, L. Postharvest decay control and quality retention in litchi (cv. McLean’s Red) by combined application of modified atmosphere packaging and antimicrobial agents. Crop Prot. 2008, 27, 1208–1214. [Google Scholar] [CrossRef]
- Cai, X.Q.; Lin, N.; Chen, W.; Hu, F.P.; DongLiang, Q.; Mitra, S.K.; Diczbalis, Y. Control effects on litchi downy blight disease by endophytic bacterial strain TB2 and its pathogenesis-related proteins. Acta Hortic. 2010, 863, 631–636. [Google Scholar] [CrossRef]
- Wu, Y.; Lin, H.; Lin, Y.; Shi, J.; Xue, S.; Hung, Y.C.; Chen, Y.; Hui, W. Effects of biocontrol bacteria Bacillus amyloliquefaciens LY-1 culture broth on quality attributes and storability of harvested litchi fruit. Postharvest Biol. Tec. 2017, 132, 81–87. [Google Scholar] [CrossRef]
- Martínez-Castellanos, G.; Pelayo-Zaldívar, C.; Pérez-Flores, L.J.; López-Luna, A.; Gimeno, M.; Bárzana, E.; Shirai, K. Postharvest litchi (Litchi chinensis Sonn.) quality preservation by Lactobacillus plantarum. Postharvest Biol. Tec. 2011, 59, 172–178. [Google Scholar] [CrossRef]
- Liao, L.; Zhou, J.; Wang, H.; Fei, H.; Liu, S.; Jiang, Z.; Chen, S.; Zhang, L.H. Control of litchi downy blight by zeamines produced by Dickeya zeae. Sci. Rep. 2015, 5, 15719. [Google Scholar] [CrossRef]
- Xing, M.; Zheng, L.; Deng, Y.; Xu, D.; Xi, P.; Li, M.; Kong, G.; Jiang, Z. Antifungal activity of natural volatile organic compounds against litchi downy blight pathogen Peronophythora litchii. Molecules 2018, 23, 358. [Google Scholar] [CrossRef]
- Stéphane, H.; Schadt, C.W. Towards a holistic understanding of the beneficial interactions across the Populus microbiome. New Phytol. 2015, 205, 1424–1430. [Google Scholar]
- Conrath, U.; Beckers, G.J.; Flors, V.; Garcã-Agustã, P.; Jakab, G.; Mauch, F.; Newman, M.A.; Pieterse, C.M.J.; Poinssot, B.; Pozo, M.J.; et al. Priming: Getting ready for battle. Mol. Plant Microbe Interact. 2006, 19, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Kasana, R.C.; Pandey, C.B. Exiguobacterium: An overview of a versatile genus with potential in industry and agriculture. Crit. Rev. Biotechnol. 2018, 38, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Vijayalaxmi, S.; AnuAppaiah, K.A.; Jayalakshmi, S.K.; Mulimani, V.H.; Sreeramuluet, K. Production of bioethanol from fermented sugars of sugarcane bagasse produced by lignocellulolytic enzymes of Exiguobacterium sp. VSG-1. Appl. Biochem. Biotech. 2013, 2171, 246–260. [Google Scholar] [CrossRef]
- Xie, J.; Xie, W.; Yu, J.; Xin, R.; Shi, Z.; Song, L.; Yang, X. Extraction of chitin from shrimp shell by successive two-step fermentation of Exiguobacterium profundum and Lactobacillus acidophilus. Front. Microbiol. 2021, 12, 677126. [Google Scholar] [CrossRef]
- Ali, C.H.; Zhang, J.J.; Mbadinga, S.M.; Mu, B.Z. Screening, isolation and optimization of an extracellular lipase producing Exiguobacterium sp. BBXS-7 segregated from waste cooking oil contaminated sites. Wulfenia J. 2015, 2, 185–201. [Google Scholar]
- Mojallali, L.; ShahbaniZahiri, H.; Rajaei, S.; Noghabi, K.A.; Haghbeenet, K. A novel ~34-kDa a-amylase from psychrotroph Exiguobacterium sp. SH3: Production, purification, and characterization. Biotechnol. Appl. Bioc. 2014, 61, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Peng, Q.; Yan, J.; Wang, H.; Ding, H.; Shi, B. Gene cloning and enzymatic characterization of alkali-tolerant type I pullulanase from Exiguobacterium acetylicum. Lett. Appl. Microbiol. 2015, 60, 52–59. [Google Scholar] [CrossRef]
- Parthasarathy, A.; Miranda, R.R.; Eddingsaas, N.C.; Chu, J.; Freezman, I.M.; Tyler, A.C.; Hudson, A.O. Polystyrene degradation by Exiguobacterium sp. RIT 594: Preliminary evidence for a pathway containing an atypical oxygenase. Microorganisms 2022, 10, 1619. [Google Scholar] [CrossRef] [PubMed]
- Sakdapetsiri, C.; Kaokhum, N.; Pinyakong, O. Biodegradation of crude oil by immobilized Exiguobacterium sp. AO-11 and shelf life evaluation. Sci. Rep. 2021, 11, 12990. [Google Scholar] [CrossRef]
- Delegan, Y.; Kocharovskaya, Y.; Bogun, A.; Sizova, A.; Solomentsev, V.; Iminova, L.; Lyakhovchenko, N.; Zinovieva, A.; Goyanov, M.; Solyanikova, I. Characterization and genomic analysis of Exiguobacterium alkaliphilum B-3531D, an efficient crude oil degrading strain. Biotechnol. Rep. 2021, 32, e00678. [Google Scholar] [CrossRef]
- Barghoth, M.G.; Desouky, S.E.; Radwan, A.A.; Shah, M.P.; Salem, S.S. Characterizations of highly efficient moderately halophilic toluene degrading Exiguobacterium mexicanum M7 strain isolated from Egyptian saline sediments. Biotechnol. Genet. Eng. 2023, 2, 1–19. [Google Scholar] [CrossRef]
- Alam, M.Z.; Malik, A. Chromate resistance, transport and bioreduction by Exiguobacterium sp. ZM-2 isolated from agricultural soil irrigated with tannery effluent. J. Basic. Microb. 2008, 48, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Okeke, B.C. Bioremoval of hexavalent chromium from water by a salt tolerant bacterium, Exiguobacterium sp. GS1. J. Industrial Microbiol. Biot. 2008, 35, 1571–1579. [Google Scholar] [CrossRef]
- Das, S.; Bikash, C.B.; Ranjan, K.M.; Biswaranjan, P.; Mathummal, S.; Anindita, C.; Hrudayanath, T. Reduction of hexavalent chromium by Exiguobacterium mexicanum isolated from chromite mines soil. Chemosphere 2021, 282, 131135. [Google Scholar] [CrossRef]
- Huang, Y.; Tang, J.; Zhang, B.; Long, Z.E.; Ni, H.; Fu, X.; Zou, L. Influencing factors and mechanism of Cr(VI) reduction by facultative anaerobic Exiguobacterium sp. PY14. Front. Microbiol. 2023, 14, 1242410. [Google Scholar] [CrossRef]
- Dastager, S.G.; Kumaran, D.C.; Pandey, A. Characterization of plant growth-promoting rhizobacterium Exiguobacterium NII-0906 for its growth promotion of cowpea (Vigna unguiculata). Biologia 2010, 65, 197–203. [Google Scholar] [CrossRef]
- Selvakumar, G.; Kundu, S.; Joshi, P.; Nazim, S.; Gupta, A.D.; Gupta, H.S. Growth promotion of wheat seedlings by Exiguobacterium acetylicum 1P (MTCC 8707) a cold tolerant bacterial strain from the Uttarakhand Himalayas. Indian J. Microbiol. 2010, 50, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Bharti, N.; Yadav, D.; Barnawal, D.; Maji, D.; Kalra, A. Exiguobacterium oxidotolerans, a halotolerant plant growth promoting rhizobacteria, improves yield and content of secondary metabolites in Bacopa monnieri (L.) Pennell under primary and secondary salt stress. World J. Microb. Biot. 2013, 29, 379–387. [Google Scholar] [CrossRef]
- Venkadesaperumal, G.; Amaresan, N.; Kumar, K. Plant growth promoting capability and genetic diversity of bacteria isolated from mud volcano and lime cave of Andaman and Nicobar Islands. Braz. J. Microbiol. 2014, 45, 1271–1281. [Google Scholar] [CrossRef]
- Marfetán, J.A.; Gallo, A.L.; Farias, M.E.; Vélez, M.L.; Pescuma, M.; Ordoñez, O.F. Exiguobacterium sp. as a bioinoculant for plant-growth promotion and selenium biofortification strategies in horticultural plants. World J. Microb. Biot. 2023, 39, 134. [Google Scholar] [CrossRef]
- Barnett, S.J.; Anstis, S.T.; Roget, D.K.; Ryder, M.H. Suppression of Rhizoctonia solani AG-8 induced disease on wheat by the interaction between Pantoea, Exiguobacterium, and Microbacteria. Aust. J. Soil. Res. 2006, 44, 331–342. [Google Scholar] [CrossRef]
- Selvakumar, G.; Joshi, P.; Nazim, S.; Mishra, P.K.; Kundu, S.; Gupta, H.S. Exiguobacterium acetylicum strain 1P MTCC 8707, a novel bacterial antagonist from the North Western Indian Himalayas. World J. Microbiol. Biotechnol. 2009, 25, 131–137. [Google Scholar] [CrossRef]
- Vishnivetskaya, T.A.; Kathariou, S.; Tiedje, J.M. The Exiguobacterium genus: Biodiversity and biogeography. Extremophiles 2009, 13, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zhu, Z.; Li, Y.; Li, X.; Guan, Z.; Zheng, J. Comparative genomics of Exiguobacterium reveals what makes a cosmopolitan bacterium. mSystems 2021, 6, e0038321. [Google Scholar] [CrossRef]
- Zheng, L.; Situ, J.; Zhu, Q.; Xi, P.; Zheng, Y.; Liu, H.; Zhou, X.; Jiang, Z.D. Identification of volatile organic compounds for the biocontrol of postharvest litchi fruit pathogen Peronophythora litchii. Postharvest Biol. Technol. 2019, 155, 37–46. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformation of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Zheng, L.; Huang, S.; Hsiang, T.; Yu, G.; Guo, D.; Jiang, Z.; Li, J. Biocontrol using Bacillus amyloliquefaciens PP19 against litchi downy blight caused by Peronophythora litchii. Front. Microbiol. 2021, 11, 619423. [Google Scholar] [CrossRef]
- Zuo, G.; Hao, B. CVTree3 Web server for whole-genome-based and alignment-free prokaryotic phylogeny and taxonomy. Genom. Proteom. Bioinf. 2015, 13, 321–331. [Google Scholar] [CrossRef]
- Sorokulova, I.; Krumnow, A.; Globa, L.; Vodyanoy, V. Efficient decomposition of shrimp shell waste using Bacillus cereus and Exiguobacterium acetylicum. J. Ind. Microbiol. Biot. 2009, 36, 1123–1126. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Srivastava, R.; Bharati, A.P.; Singh, A.K.; Sharma, A.; Das, S.; Tiwari, P.K.; Srivastava, A.K.; Chakdar, H.; Kashyap, P.L.; et al. Analysis of biosynthetic gene clusters, secretory, and antimicrobial peptides reveals environmental suitability of Exiguobacterium profundum PHM11. Front. Microbiol. 2022, 12, 785458. [Google Scholar] [CrossRef] [PubMed]
- Mekuto, L.; Alegbeleye, O.O.; Ntwampe, S.K.; Ngongang, M.M.; Mudumbi, J.B.; Akinpelu, E.A. Co-metabolism of thiocyanate and free cyanide by Exiguobacterium acetylicum and Bacillus marisflavi under alkaline conditions. 3 Biotech. 2016, 6, 173. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, Y.; Hao, X.; Zhang, X.; Ma, Y.; Niu, Z. A novel marine bacterium Exiguobacterium marinum a-1 isolated from in situ plastisphere for degradation of additive-free polypropylene. Environ. Pollut. 2023, 336, 122390. [Google Scholar] [CrossRef] [PubMed]
- Mhamdi, A.; Van Breusegem, F. Reactive oxygen species in plant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Li, J.; Huang, C.; Zhang, D.; Xu, Y.; Yang, X.; Song, J.; Wang, D.; Qiu, N.; Short, D.P.G.; et al. Cu/Zn superoxide dismutase (VdSOD1) mediates reactive oxygen species detoxification and modulates virulence in Verticillium dahliae. Mol. Plant Pathol. 2021, 22, 1092–1108. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Bioch. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Zhang, X.; Xin, Y.; Wang, J.; Dhanasekaran, S.; Yue, Q.; Feng, F.; Gu, X.; Li, B.; Zhao, L.; Zhang, H. Characterization of a Bacillus velezensis strain as a potential biocontrol agent against soft rot of eggplant fruits. Int. J. Food Microbiol. 2024, 410, 110480. [Google Scholar] [CrossRef] [PubMed]
- Nan, J.; Zhang, S.; Jiang, L. Antibacterial potential of Bacillus amyloliquefaciens GJ1 against citrus Huanglongbing. Plants 2021, 10, 261. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.H.; Hong, C.F.; Huang, J.W. Identification and characterization of a multifunctional biocontrol agent, Streptomyces griseorubiginosus LJS06, against cucumber anthracnose. Front. Microbiol. 2022, 13, 923276. [Google Scholar] [CrossRef]
- Duke, K.A.; Becker, M.G.; Girard, I.J.; Millar, J.L.; Dilantha Fernando, W.G.; Belmonte, M.F.; de Kievit, T.R. The biocontrol agent Pseudomonas chlororaphis PA23 primes Brassica napus defenses through distinct gene networks. BMC Genom. 2017, 18, 467. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, Y.; Qiao, J.; Yu, W.; Pan, X.; Zhang, T.; Liu, Y.; Lu, S.E. Phenazine-1-carboxylic acid produced by Pseudomonas chlororaphis YL-1 is effective against Acidovorax citrulli. Microorganisms 2021, 9, 2012. [Google Scholar] [CrossRef]
- Yang, K.; Chen, C.; Wang, Y.; Li, J.; Dong, X.; Cheng, Y.; Zhang, H.; Zhai, Y.; Ai, G.; Song, Q.; et al. Nep1-like proteins from the biocontrol agent Pythium oligandrum enhance plant disease resistance independent of cell death and reactive oxygen species. Front. Plant Sci. 2022, 13, 830636. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).