
Genetic Diversity and Population Differentiation of Pinus koraiensis in China



Abstract
1. Introduction
2. Results
2.1. Genetic Diversity at Different Loci among Populations
2.2. Genetic Diversity within Pinus koraiensis Populations
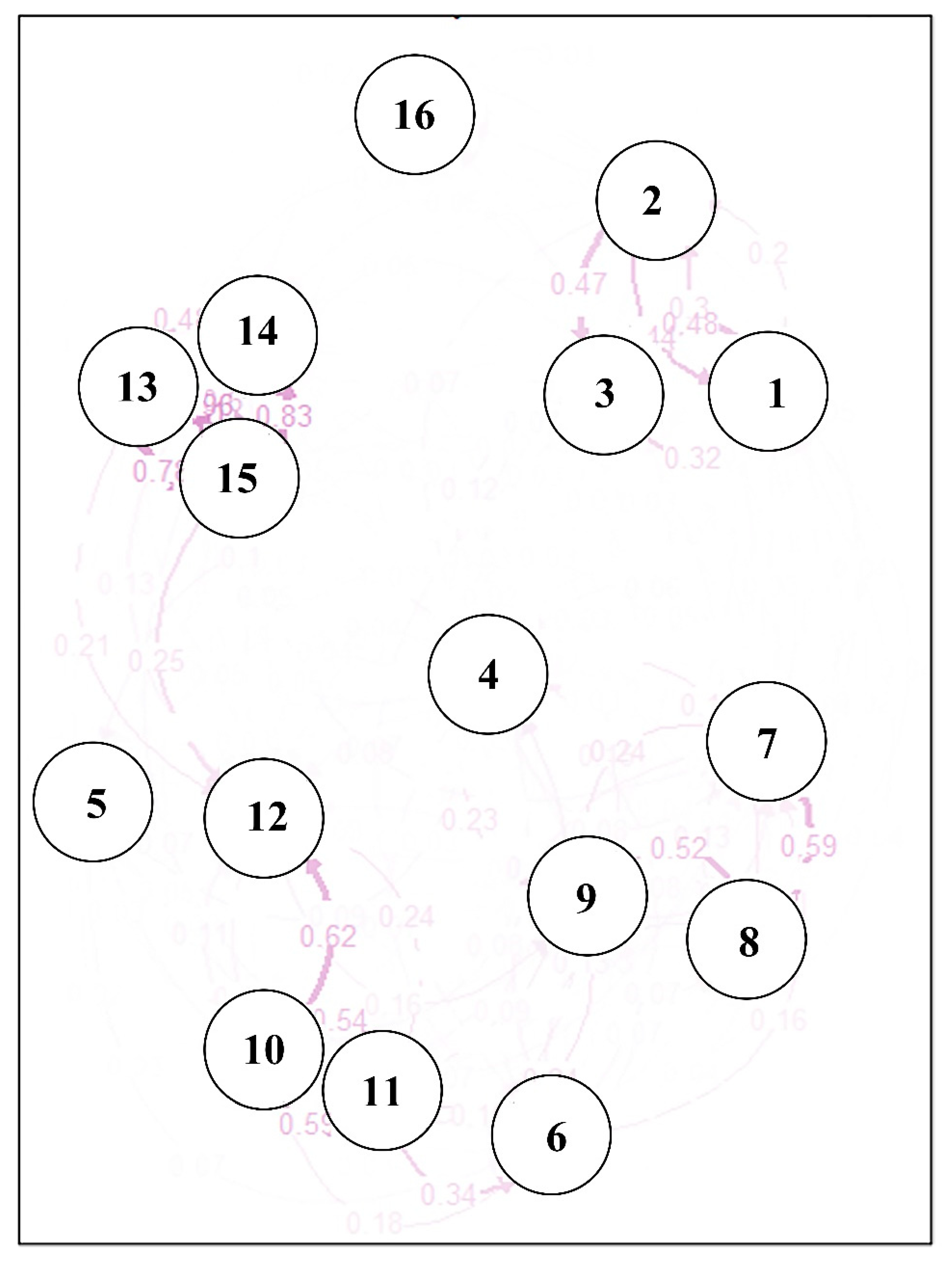
2.3. Genetic Variation among Pinus koraiensis Populations
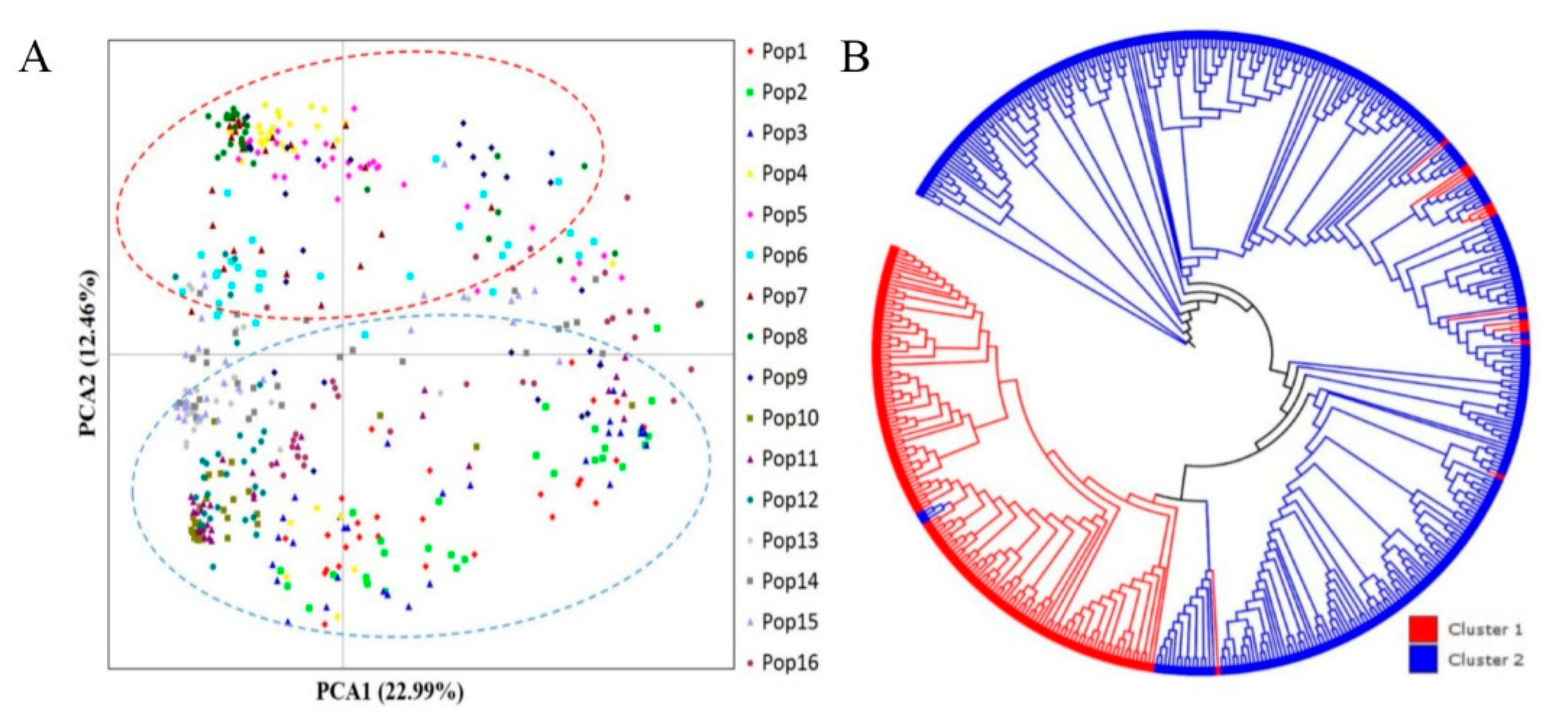
2.4. Population Structure
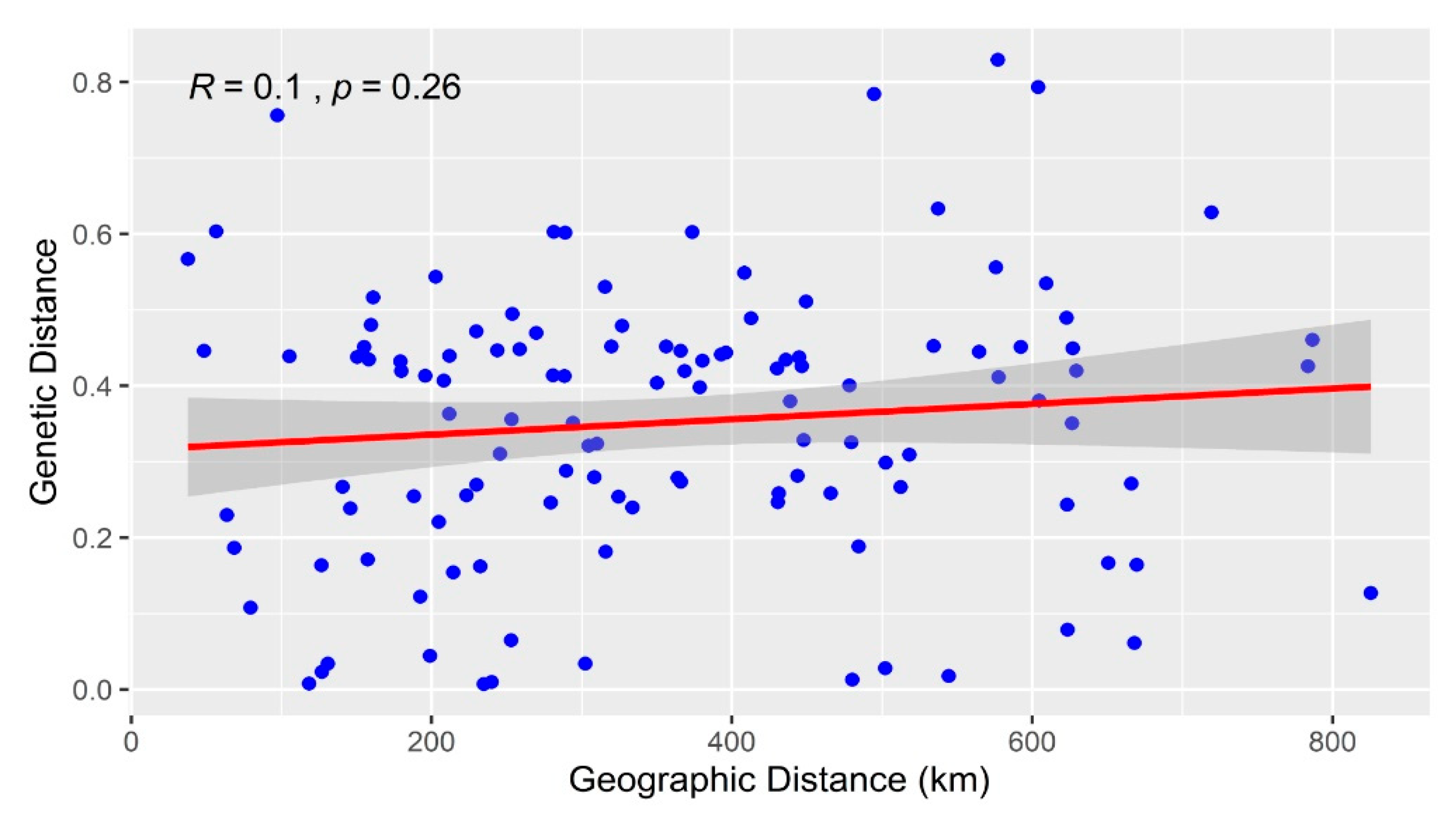
2.5. Correlations between Genetic Distance and Geographic Distance
3. Discussion
3.1. Genetic Diversity
3.2. Population Genetic Differentiation
3.3. Population Structure and Gene Flow
3.4. Conservation and Management Strategies
4. Materials and Methods
4.1. Plant Materials and Genomic DNA Extraction
4.2. PCR Amplification and SSR Analysis
4.3. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Choi, D.-S.; Jin, H.-O.; Lee, C.-H.; Kim, Y.-C.; Kayama, M. Effect of soil acidification on the growth of Korean pine (Pinus koraiensis) seedlings in a granite-derived forest soil. Environ. Sci. Int. J. Environ. Physiol. Toxicol. 2005, 12, 33–47. [Google Scholar]
- Jia, Y.; Zhu, J.; Wu, Y.; Fan, W.-B.; Zhao, G.-F.; Li, Z.-H. Effects of Geological and Environmental Events on the Diversity and Genetic Divergence of Four Closely Related Pines: Pinus koraiensis, P. armandii, P. griffithii, and P. pumila. Front. Plant Sci. 2018, 9, 1264. [Google Scholar] [CrossRef]
- Wang, F.; Chen, S.; Liang, D.; Qu, G.-Z.; Chen, S.; Zhao, X. Transcriptomic analyses of Pinus koraiensis under different cold stresses. BMC Genom. 2020, 21, 1–14. [Google Scholar] [CrossRef]
- Ma, J.L. The evolution of Korea Pine forest. J. Northeast. For. Univ. 1997, 25, 67–71. [Google Scholar]
- Wang, H.; Shao, X.-M.; Jiang, Y.; Fang, X.-Q.; Wu, S.-H. The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For. Ecol. Manag. 2013, 289, 333–340. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, K.; Sun, Y.; Yan, Q. Response of Pinus koraiensis seedling growth to different light conditions based on the assessment of photosynthesis in current and one-year-old needles. J. For. Res. 2014, 25, 53–62. [Google Scholar] [CrossRef]
- Fan, Y.; Moser, W.K.; Cheng, Y. Growth and Needle Properties of Young Pinus koraiensis Sieb. et Zucc. Trees across an Elevational Gradient. Forests 2019, 10, 54. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.T.; Wei, J.T.; Li, Y.; Tigabu, M.; Zhao, X.Y. Genetic improvement of Pinus koraiensis in China: Current situation and ffuture prospects. Forests 2020, 11, 148. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, Y.; Zhou, G.; Xiao, C. Composition and Structure of Pinus koraiensis Mixed Forest Respond to Spatial Climatic Changes. PLoS ONE 2014, 9, e97192. [Google Scholar] [CrossRef]
- Wang, X.C.; Zhang, M.H.; Ying, J.; Li, Z.S.; Zhang, Y.D. Temperature signals in tree-ring width and divergent growth of Ko-rean pine response to recent climate warming in northeast Asia. Trees 2016, 31, 1–13. [Google Scholar]
- Seo, Y.-W.; Balekoglu, S.; Choi, J.-K. Growth pattern analysis by stem analysis of Korean white pine (Pinus koraiensis) in the central northern region of Korea. For. Sci. Technol. 2014, 10, 220–226. [Google Scholar] [CrossRef][Green Version]
- Zhang, S.T.; Zhang, L.G.; Wang, L.; Zhao, Y.H. Total phenols, flavonoids, and procyanidins levels and total antioxidant activity of different Korean pine (Pinus koraiensis) varieties. J. For. Res. 2018, 30, 1743–1754. [Google Scholar] [CrossRef]
- Lai, F.; Xie, H.; Wang, J. Functions of biology and disease control with Pycnogonid from pine bark extract. Chin. Tradit. Herb. Drugs 2005, 36, 127–131. [Google Scholar]
- Liu, M.; Mao, Z.J.; Li, Y.; Sun, T.; Li, X.H.; Huang, W.; Liu, R.P.; Li, Y.H. Response of radial growth of Pinus koraiensis in broad-leaved Korean pine forests with different latitudes to climatical factors. Ying Yong Sheng Tai Xue Bao J. Appl. Ecol. 2016, 27, 1341–1352. [Google Scholar]
- Kang, K.; Harju, A.; Lindgren, D.; Nikkanen, T.; Almqvist, C.; Suh, G. Variation in effective number of clones in seed orchards. New For. 2001, 21, 17–33. [Google Scholar] [CrossRef]
- Han, S.U.; Kang, K.S.; Kim, C.S.; Kim, T.S.; Song, J.H. Effect of top-pruning in a clonal seed orchard of Pinus koraiensis. Ann. For. Res. 2008, 51, 155–156. [Google Scholar]
- Park, J.M.; Kwon, S.H.; Lee, H.J.; Na, S.J.; El-Kassaby, Y.A.; Kang, K.S. Integrating fecundity variation and genetic related-ness in estimating the gene diversity of seed crops: Pinus koraiensis seed orchard as an example. Can. J. Forest. Res. 2016, 47, 366–370. [Google Scholar] [CrossRef]
- Guan, G.Y. Effects of different scion types on grafting survival rate and growth potential of Pinus koraiensis. Prot. For. Sci. Technol. 2019, 000, 44–45. [Google Scholar]
- Xia, D.; Yang, S.W.; Yang, C.P.; Lv, Q.Y.; Liu, G.F.; Zhang, P.G. Study of Pinus koraiensis provenance test (1): Provenance re-gionalization. J. Northeast. Univ. 1991, 19, 122–128. [Google Scholar]
- Wang, Y.X.; Dong, Y.H.; Yang, H.; Wang, H.Z.; Xu, J.M.; Shan, M.G.; Zhang, G.L. Open pollination progeny test of clonal seed orchard of Pinus koraiensis. For. Sci. Technol. 2005, 34, 20–21. [Google Scholar]
- Liang, D.; Ding, C.; Zhao, G.; Leng, W.; Zhang, M.; Zhao, X.; Qu, G. Variation and selection analysis of Pinus koraiensis clones in northeast China. J. For. Res. 2017, 29, 611–622. [Google Scholar] [CrossRef]
- Chen, M.-M.; Feng, F.; Sui, X.; Li, M.-H.; Zhao, D.; Han, S. Construction of a framework map for Pinus koraiensis Sieb. et Zucc. using SRAP, SSR and ISSR markers. Trees 2010, 24, 685–693. [Google Scholar] [CrossRef]
- Li, X.; Liu, X.; Wei, J.; Li, Y.; Tigabu, M.; Zhao, X. Development and Transferability of EST-SSR Markers for Pinus koraiensis from Cold-Stressed Transcriptome through Illumina Sequencing. Genes 2020, 11, 500. [Google Scholar] [CrossRef]
- Kwon, S.; Kim, S.; Kim, J.; Kang, W.; Park, K.-H.; Kim, C.-B.; Girona, M.M. Predicting Post-Fire Tree Mortality in a Temperate Pine Forest, Korea. Sustainability 2021, 13, 569. [Google Scholar] [CrossRef]
- Li, J.Q.; Wang, L.H.; Zhan, Q.W.; Liu, Y.L.; Yang, X.C. Transcriptome characterization and functional marker development in Sorghum sudanense. PLoS ONE 2016, 11, e0154947. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Song, P.; Koo, D.-H.; Guo, L.; Li, Y.; Sun, S.; Weng, Y.; Yang, L. Genome wide characterization of simple sequence repeats in watermelon genome and their application in comparative mapping and genetic diversity analysis. BMC Genom. 2016, 17, 1–17. [Google Scholar] [CrossRef]
- Zhao, X.; Li, C.; Wan, S.; Zhang, T.; Yan, C.; Shan, S. Transcriptomic analysis and discovery of genes in the response of Arachis hypogaea to drought stress. Mol. Biol. Rep. 2018, 45, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Geng, Y.; Song, M.; Zhang, P.; Hou, J.; Wang, W. Genetic Structure and Diversity of Glycyrrhiza Populations Based on Transcriptome SSR Markers. Plant Mol. Biol. Rep. 2019, 37, 401–412. [Google Scholar] [CrossRef]
- Gouy, M.; Rousselle, Y.; Chane, A.T.; Anglade, A.; Royaert, S.; Nibouche, S.; Costet, L. Genome wide association mapping of agro-morphological and disease resistance traits in sugarcane. Euphytica 2014, 202, 269–284. [Google Scholar] [CrossRef]
- Huang, Z.; Zhao, N.; Qin, M.; Xu, A. Mapping of quantitative trait loci related to cold resistance in Brassica napus L. J. Plant Physiol. 2018, 231, 147–154. [Google Scholar] [CrossRef]
- Kim, Z.S.; Hwang, J.W.; Lee, S.W.; Yang, C.; Gorovoy, P.G. Genetic Variation of Korean Pine (Pinus koraiensis Sieb. et Zucc.) at Allozyme and RAPD Markers in Korea, China and Russia. Silvae Genet. 2005, 54, 235–246. [Google Scholar] [CrossRef]
- Acheampong, A.; Leveque, N.; Tchapla, A.; Heron, S. Application of SRAP in the genetic diversity of Pinus koraiensis of different provenances. Afr. J. Biotechnol. 2010, 8, 1000–1008. [Google Scholar]
- Chen, J.Y. Analysis of Genetic Diversity of Pinus koraiensis Plantation in CaoHekou Forest Farm by ISSR Marker. Bull. Bot. Res. 2009, 29, 633–636. [Google Scholar]
- Feng, F.-J.; Han, S.-J.; Wang, H.-M. Genetic diversity and genetic differentiation of natural Pinus koraiensis population. J. For. Res. 2006, 17, 21–24. [Google Scholar] [CrossRef]
- Tong, Y.W.; Lewis, B.J.; Zhou, W.M.; Mao, C.R.; Wang, Y.; Zhou, L.; Yu, D.P.; Dai, L.M.; Qi, L. Genetic Diversity and Population Structure of Natural Pinus koraiensis Populations. Forests 2020, 11, 39. [Google Scholar] [CrossRef]
- Halbauer, E.-M.; Bohinec, V.; Wittenberger, M.; Hansel-Hohl, K.; Gaubitzer, S.; Sehr, E.M. Genetic diversity of flax accessions originating in the Alpine region: A case study for an ex situ germplasm evaluation based on molecular marker. Euphytica 2017, 213, 120. [Google Scholar] [CrossRef]
- Krishnan, A.G.; John, R.; Cyriac, A.; Sible, G. Estimation of genetic diversity in nutmeg (Myristica fragrans Houtt.) selections using RAPD markers. Int. J. Plant Sci. 2017, 12, 102–107. [Google Scholar] [CrossRef]
- Thomas, L. Genetic variation in Turkish Red Pine (Pinus brutia Ten.) seed stands as determined by RAPD markers. Silvae Genet. 2017, 53, 169–175. [Google Scholar]
- Chung, M.Y.; Moon, M.O.; López-Pujol, J.; Chung, J.M.; Chung, M.G. Genetic diversity in the two endangered endemic species Kirengeshoma koreana (Hydrangeaceae) and Parasenecio pseudotaimingasa (Asteraceae) from Korea: Insights into population history and implications for conservation. Biochem. Syst. Ecol. 2013, 51, 60–69. [Google Scholar] [CrossRef]
- Sanaa, A.; Boulila, A.; Boussaid, M.; Ben Fadhel, N. Genetic diversity, population structure and linkage disequilibrium analysis in the endangered Tunisian Pancratium maritimum Linnaeus (Amaryllidaceae) populations. Afr. J. Ecol. 2016, 54, 379–382. [Google Scholar] [CrossRef]
- Duan, D.; Jia, Y.; Yang, J.; Li, Z.H. Comparative transcriptome analysis of male and female conelets and development of mi-crosatellite markers in Pinus bungeana, an Endemic Conifer in China. Genes 2017, 8, 393. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, H.; Chen, W.; Pang, X.; Li, Y. Genetic diversity and structure of native and non-native populations of the endangered plant Pinus dabeshanensis. Genet. Mol. Res. 2016, 15, 15027937. [Google Scholar] [CrossRef]
- Xu, Y.L.; Cai, N.H.; Woeste, K.; Kang, X.Y.; He, C.Z.; Li, G.Q.; Chen, S.; Duan, A.A. Genetic diversity and population structure of Pinus yunnanensis by simple sequence repeat markers. Forest. Sci. 2016, 62, 38–47. [Google Scholar] [CrossRef]
- Wang, M.B.; Hao, Z.Z. Rangewide Genetic Diversity in Natural Populations of Chinese Pine (Pinus tabulaeformis). Biochem. Genet. 2010, 48, 590–602. [Google Scholar] [CrossRef]
- Ferrer, M.M.; Eguiarte, L.E.; Montaña, C. Genetic structure and outcrossing rates in Flourensia cernua (Asteraceae) growing at different densities in the South-western Chihuahuan Desert. Ann. Bot. 2004, 94, 419–426. [Google Scholar] [CrossRef]
- Hellmann, J.J.; Pineda-Krch, M. Constraints and reinforcement on adaptation under climate change: Selection of genetically correlated traits. Biol. Conserv. 2007, 137, 599–609. [Google Scholar] [CrossRef]
- Wang, Z.; Kang, M.; Liu, H.; Gao, J.; Zhang, Z.; Li, Y.; Wu, R.; Pang, X. High-Level Genetic Diversity and Complex Population Structure of Siberian Apricot (Prunus sibirica L.) in China as Revealed by Nuclear SSR Markers. PLoS ONE 2014, 9, e87381. [Google Scholar] [CrossRef]
- Zu, Y.; Yu, J.; Wang, A. Study on pollination characteristics of natural population of Pinus koraiensis. Acta Ecol. Sin. 2000, 20, 430–433. [Google Scholar]
- Zhang, Z.; Zhang, H.G.; Mo, C.; Zhang, L. Transcriptome sequencing analysis and development of EST-SSR markers for Pinus koraiensis. Sci. Silvae Sin. 2015, 51, 114–120. [Google Scholar]
- Omelko, A.; Ukhvatkina, O.; Zhmerenetsky, A. Disturbance history and natural regeneration of an old-growth Korean pine-broadleaved forest in the Sikhote-Alin mountain range, Southeastern Russia. For. Ecol. Manag. 2016, 360, 221–234. [Google Scholar] [CrossRef]
- Parsons, B.J.; Newbury, H.J.; Jackson, M.T.; Ford-Lloyd, B.V. Contrasting genetic diversity relationships are revealed in rice (Oryza sativa L.) using different marker types. Mol. Breed. 1997, 3, 115–125. [Google Scholar] [CrossRef]
- Chen, W.; Hou, L.; Zhang, Z.; Pang, X.; Li, Y. Genetic Diversity, Population Structure, and Linkage Disequilibrium of a Core Collection of Ziziphus jujuba Assessed with Genome-wide SNPs Developed by Genotyping-by-sequencing and SSR Markers. Front. Plant Sci. 2017, 8, 575. [Google Scholar] [CrossRef] [PubMed]
- Kim, Z.S.; Lee, S.W.; Lim, J.H.; Hwang, J.W.; Kwon, W. Genetic diversity and structure of natural populations of Pinus koraiensis (Sieb. et Zucc.) in Korea. For. Genet. 1994, 1, 41–49. [Google Scholar]
- Potenko, V.V.; Velikov, A.V. Allozyme Variation and mating system of coastal populations of Pinus koraiensis sieb. et Zucc. in Russia. Silvae Genet. 2001, 50, 117–122. [Google Scholar]
- Wright, S. The interpretation of population structure by F-Statistics with special regard to systems of mating. Evolution 1965, 19, 395–420. [Google Scholar] [CrossRef]
- Holsinger, K.E.; Weir, B.S. Genetics in geographically structured populations: Defining, estimating and interpreting FST. Nat. Rev. Genet. 2009, 10, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, G.; Jueterbock, A.; Kraemer, P.; Deppermann, J.; Harmand, P. Calculations of population differentiation based on GST and D: Forget GST but not all of statistics! Mol. Ecol. 2010, 19, 3845–3852. [Google Scholar] [CrossRef]
- Waples, R.S. Separating the wheat from the chaff: Patterns of genetic differentiation in high gene flow species. J. Hered. 1998, 89, 438–450. [Google Scholar] [CrossRef]
- Raaijeveld-Smit, F.J.L.K.; Eebee, T.J.C.B.; Riffiths, R.A.G.; Oore, R.D.M.; Schley, L. Low gene flow but high genetic diversity in the threatened Mallorcan midwife toad Alytes muletensis. Mol. Ecol. 2005, 14, 3307–3315. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.M.; Jacklyn, P.; Korb, J. Isolated in an ocean of grass: Low levels of gene flow between termite subpopulations. Mol. Ecol. 2013, 22, 2096–2105. [Google Scholar] [CrossRef]
- Zhao, F.; Yang, J.; He, H.S.; Dai, L. Effects of Natural and Human-Assisted Regeneration on Landscape Dynamics in a Korean Pine Forest in Northeast China. PLoS ONE 2013, 8, e82414. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Kudureti, A.; Bai, W.; Chen, R.; Wang, T.; Wang, H.; Ge, J. Contributions of multiple refugia during the last glacial period to current mainland populations of Korean pine (Pinus koraiensis). Sci. Rep. 2015, 5, 18608. [Google Scholar] [CrossRef]
- Schoettle, A.W.; Goodrich, B.A.; Hipkins, V.D.; Richards, C.; Kray, J. Geographic patterns of genetic variation and population structure in Pinus aristata, Rocky Mountain bristlecone pine. Can. J. For. Res. 2011, 42, 23–37. [Google Scholar] [CrossRef]
- Shamseldeen, E.; Sallam, A.; Belamkar, V.; Emara, H.A.; Nower, A.A.; Salem, K.F.; Poland, J.; Baenziger, P.S. Genetic diversity and population structure of F3:6 nebraska winter wheat genotypes using genotyping-by-sequencing. Front. Genet. 2018, 9, 76. [Google Scholar]
- Luo, Z.N.; Brock, J.; Dyer, J.M.; Kutchan, T.; Schachtman, D.; Augustin, M.; Ge, Y.F.; Fahlgren, N.; Abdel-Haleem, H. Genetic diversity and population structure of a camelina sativa spring panel. Front. Plant Sci. 2019, 10, 184. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Vieira, F.G.; Crawford, J.E.; Chu, C.; Nielsen, R. Asian wild rice is a hybrid swarm with extensive gene flow and feralization from domesticated rice. Genome Res. 2017, 27, 1029–1038. [Google Scholar] [CrossRef]
- Peng, L.P.; Cai, C.F.; Zhong, Y.; Xu, X.X.; Xian, H.L.; Cheng, F.Y.; Mao, J.F. Genetic analyses reveal independent domestication origins of the emerging oil crop Paeonia ostii, a tree peony with a long-term cultivation history. Sci. Rep. 2017, 7, 5340. [Google Scholar] [CrossRef]
- Chung, M.Y.; Chung, M.G. Large effective population sizes and high levels of gene flow between subpopulations of Lilium cernuum (Liliaceae). Biochem. Syst. Ecol. 2014, 54, 354–361. [Google Scholar] [CrossRef]
- Bartkowska, M.P.; Wong, A.Y.C.; Sagar, S.P.; Zeng, L.; Eckert, C.G. Lack of spatial structure for phenotypic and genetic variation despite high self-fertilization in Aquilegia canadensis (Ranunculaceae). Heredity 2018, 121, 605–615. [Google Scholar] [CrossRef]
- Herman, A.; Brandvain, Y.; Weagley, J.; Jeffery, W.R.; Keene, A.C.; Kono, T.J.Y.; Bilandžija, H.; Borowsky, R.; Espinasa, L.; O’Quin, K.; et al. The role of gene flow in rapid and repeated evolution of cave-related traits in Mexican tetra, Astyanax mexcanus. Mol. Ecol. 2018, 27, 4397–4416. [Google Scholar] [CrossRef]
- Yerka, M.K.; De Leon, N.; Stoltenberg, D.E. Pollen-Mediated Gene Flow in Common Lambsquarters (Chenopodium album). Weed Sci. 2012, 60, 600–606. [Google Scholar] [CrossRef]
- Hernando, R.C.; Ken, O.; Mauricio, Q.; Fuchs, E.J.; Antonio, G.R. Contrasting patterns of population history and seed-mediated gene flow in two endemic costa rican oak species. J. Hered. 2018, 109, 530–542. [Google Scholar]
- Aegisdóttir, H.H.; Patrick, K.; Jürg, S. Isolated populations of a rare alpine plant show high genetic diversity and considerable population differentiation. Ann. Bot. 2009, 104, 1313–1322. [Google Scholar] [CrossRef]
- Kaljund, K.; Jaaska, V. No loss of genetic diversity in small and isolated populations of Medicago sativa subsp. falcata. Biochem. Syst. Ecol. 2010, 38, 510–520. [Google Scholar] [CrossRef]
- Dostálek, T.; Münzbergová, Z.; Plačková, I. High genetic diversity in isolated populations of Thesium ebracteatum at the edge of its distribution range. Conserv. Genet. 2013, 15, 75–86. [Google Scholar] [CrossRef]
- Li, X.; Li, M.; Hou, L.; Zhang, Z.; Pang, X.; Li, Y. De Novo Transcriptome Assembly and Population Genetic Analyses for an Endangered Chinese Endemic Acer miaotaiense (Aceraceae). Genes 2018, 9, 378. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Xia, R.; Chen, H.; He, Y.H. TBtools, a Toolkit for Biologists integrating various HTS-data handling tools with a user-friendly interface. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Sándor, N.; Péter, P.; István, C.; Mousapour, G.A.; Géza, H.; János, T. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar]
- Hamrick, J.L.; Godt, M.J.W. Effects of life history traits on genetic diversity in plant species. Philos. Trans. R. Soc. B Biol. Sci. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of Population Structure Using Multilocus Genotype Data: Linked Loci and Correlated Allele Frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; Vonholdt, B.M. Structure harvester: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef]
- Keenan, K.; Mcginnity, P.; Cross, T.F.; Crozier, W.W.; Prodöhl, P.A. DiveRsity: An R package for the estimation and exploration of population genetics parameters and their associated errors. Methods Ecol. Evol. 2013, 4, 782–788. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A Language for Data Analysis and Graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Allele Size Range (bp) | Na | Ne | I | Ho | He | PIC | HWE | NRA | Fis | Fit | Fst | Nm |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NEPK-218 | 196–230 | 12 | 2.080 | 0.949 | 0.056 | 0.519 | 0.443 | *** | 6 | 0.647 | 0.893 | 0.695 | 0.109 |
| NEPK-40 | 196–238 | 13 | 1.170 | 0.399 | 0.063 | 0.145 | 0.142 | *** | 3 | 0.327 | 0.593 | 0.396 | 0.382 |
| NEPK-32 | 206–224 | 5 | 2.431 | 0.979 | 0.618 | 0.589 | 0.503 | *** | 1 | −0.236 | −0.053 | 0.148 | 1.438 |
| NEPK-53 | 197–218 | 5 | 1.902 | 0.697 | 0.008 | 0.474 | 0.367 | *** | 3 | 0.923 | 0.981 | 0.757 | 0.080 |
| NEPK-65 | 151–259 | 12 | 2.257 | 1.068 | 0.195 | 0.557 | 0.488 | *** | 5 | 0.259 | 0.649 | 0.526 | 0.225 |
| NEPK-71 | 215–239 | 7 | 2.013 | 0.772 | 0.635 | 0.503 | 0.392 | *** | 3 | −0.275 | −0.254 | 0.016 | 15.141 |
| NEPK-117 | 205–217 | 9 | 3.995 | 1.601 | 0.984 | 0.750 | 0.714 | *** | 2 | −0.594 | −0.290 | 0.191 | 1.058 |
| NEPK-72 | 222–234 | 7 | 1.649 | 0.637 | 0.043 | 0.394 | 0.324 | *** | 4 | 0.346 | 0.894 | 0.838 | 0.048 |
| NEPK-67 | 226–232 | 4 | 2.802 | 1.067 | 0.479 | 0.643 | 0.567 | *** | 1 | 0.034 | 0.259 | 0.233 | 0.822 |
| NEPK-38 | 229–255 | 15 | 1.211 | 0.498 | 0.096 | 0.174 | 0.172 | *** | 6 | 0.396 | 0.477 | 0.134 | 1.622 |
| NEPK-145 | 251–289 | 21 | 6.605 | 2.243 | 0.637 | 0.849 | 0.833 | *** | 6 | 0.229 | 0.309 | 0.104 | 2.154 |
| NEPK-168 | 241–301 | 18 | 3.550 | 1.602 | 0.279 | 0.718 | 0.674 | *** | 9 | 0.265 | 0.602 | 0.459 | 0.295 |
| NEPK-181 | 275–289 | 10 | 2.039 | 0.884 | 0.165 | 0.509 | 0.426 | *** | 5 | 0.375 | 0.667 | 0.467 | 0.285 |
| NEPK-213 | 276–298 | 12 | 1.846 | 1.039 | 0.421 | 0.458 | 0.435 | *** | 4 | −0.151 | 0.109 | 0.226 | 0.856 |
| NEPK-184 | 291–301 | 5 | 2.152 | 0.856 | 0.937 | 0.535 | 0.430 | *** | 0 | −0.743 | −0.719 | 0.014 | 17.691 |
| Average | 10.33 | 2.514 | 1.019 | 0.374 | 0.521 | 0.461 | 3.867 | 0.12 | 0.341 | 0.347 | 2.667 |
| Population | Size | Na | Ne | I | Ho | He | uHe | F (Null) | NRA |
|---|---|---|---|---|---|---|---|---|---|
| Liangzihe | 30 | 3.267 | 1.713 | 0.639 | 0.415 | 0.374 | 0.388 | −0.054 | 7 |
| Helong | 30 | 2.667 | 1.663 | 0.529 | 0.390 | 0.310 | 0.321 | −0.195 | 2 |
| Lushuihe | 30 | 3.133 | 1.829 | 0.628 | 0.394 | 0.350 | 0.359 | 0.015 | 1 |
| Heihe | 30 | 3.133 | 1.939 | 0.746 | 0.340 | 0.439 | 0.449 | 0.325 | 3 |
| Liangshui | 10 | 3.333 | 1.914 | 0.664 | 0.470 | 0.370 | 0.380 | −0.235 | 6 |
| Zhanhe | 30 | 4.133 | 2.009 | 0.782 | 0.356 | 0.413 | 0.422 | 0.156 | 12 |
| Tieli | 30 | 4.467 | 2.222 | 0.806 | 0.373 | 0.414 | 0.422 | 0.178 | 8 |
| Hegang | 30 | 3.533 | 1.702 | 0.634 | 0.401 | 0.354 | 0.361 | −0.088 | 7 |
| Linjiang | 28 | 3.667 | 2.257 | 0.860 | 0.470 | 0.488 | 0.502 | 0.018 | 4 |
| Jiaohe | 30 | 3.067 | 1.586 | 0.497 | 0.293 | 0.275 | 0.280 | 0.127 | 1 |
| Hulin | 29 | 2.733 | 1.877 | 0.523 | 0.324 | 0.294 | 0.300 | −0.025 | 0 |
| Boli | 30 | 3.400 | 1.970 | 0.666 | 0.328 | 0.361 | 0.367 | 0.116 | 2 |
| Muling | 30 | 3.000 | 1.840 | 0.525 | 0.335 | 0.285 | 0.290 | −0.013 | 3 |
| Maoershan | 30 | 3.000 | 1.784 | 0.539 | 0.368 | 0.293 | 0.300 | −0.186 | 2 |
| Fangzheng | 30 | 3.000 | 1.711 | 0.531 | 0.341 | 0.286 | 0.292 | 0.033 | 1 |
| Wangqing | 30 | 2.800 | 1.913 | 0.579 | 0.327 | 0.328 | 0.337 | 0.148 | 2 |
| Mean | 3.271 | 1.870 | 0.634 | 0.370 | 0.352 | 0.361 | 0.020 | 3.813 |
| Type | Source of Variation | d.f. | Sum of Squares | Variance Component | Percentage of Variation | Fixation Index |
|---|---|---|---|---|---|---|
| Variance partition a | Among populations | 15 | 2191.019 | 4.559 | 33.00 | FST = 0.516 |
| Within populations | 464 | 4311.567 | 9.292 | 67.00 | ||
| Total | 479 | 6502.585 | 13.851 | |||
| Variance partition b | Among groups | 1 | 115.322 | 0.266 | 26.67 | FST = 0.362 |
| Among populations within groups | 14 | 88.917 | 0.095 | 9.55 | FSC = 0.130 | |
| Within populations | 942 | 600.429 | 0.637 | 63.79 | FCT = 0.267 | |
| Total | 957 | 804.668 | 0.999 | |||
| Variance partition c | Among groups | 1 | 23.753 | 0.024 | 2.77 | FST = 0.264 |
| Among populations within groups | 14 | 180.487 | 0.205 | 23.64 | FSC = 0.243 | |
| Within populations | 942 | 600.429 | 0.637 | 73.59 | FCT = 0.028 | |
| Total | 957 | 804.668 | 0.866 |
| P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | P13 | P14 | P15 | P16 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | **** | 0.008 | 0.002 | 0.383 | 0.737 | 0.452 | 0.498 | 0.550 | 0.426 | 0.418 | 0.397 | 0.418 | 0.437 | 0.432 | 0.422 | 0.471 |
| P2 | 0.029 | **** | 0.007 | 0.443 | 0.813 | 0.403 | 0.520 | 0.619 | 0.458 | 0.395 | 0.388 | 0.406 | 0.439 | 0.466 | 0.434 | 0.423 |
| P3 | 0.017 | 0.021 | **** | 0.410 | 0.778 | 0.434 | 0.475 | 0.543 | 0.425 | 0.421 | 0.400 | 0.422 | 0.436 | 0.440 | 0.432 | 0.425 |
| P4 | 0.188 | 0.233 | 0.208 | **** | 0.048 | 0.154 | 0.106 | 0.167 | 0.104 | 0.368 | 0.337 | 0.285 | 0.258 | 0.244 | 0.245 | 0.612 |
| P5 | 0.298 | 0.348 | 0.322 | 0.054 | **** | 0.222 | 0.172 | 0.216 | 0.145 | 0.411 | 0.385 | 0.342 | 0.368 | 0.338 | 0.351 | 0.769 |
| P6 | 0.202 | 0.203 | 0.202 | 0.091 | 0.124 | **** | 0.093 | 0.207 | 0.143 | 0.317 | 0.297 | 0.260 | 0.255 | 0.266 | 0.234 | 0.430 |
| P7 | 0.221 | 0.245 | 0.220 | 0.070 | 0.104 | 0.051 | **** | 0.022 | 0.041 | 0.427 | 0.414 | 0.312 | 0.315 | 0.309 | 0.299 | 0.439 |
| P8 | 0.239 | 0.271 | 0.239 | 0.098 | 0.124 | 0.104 | 0.025 | **** | 0.059 | 0.540 | 0.520 | 0.402 | 0.432 | 0.403 | 0.410 | 0.498 |
| P9 | 0.183 | 0.213 | 0.193 | 0.058 | 0.092 | 0.071 | 0.036 | 0.051 | **** | 0.2387 | 0.225 | 0.170 | 0.271 | 0.260 | 0.264 | 0.474 |
| P10 | 0.219 | 0.240 | 0.224 | 0.212 | 0.228 | 0.169 | 0.218 | 0.257 | 0.140 | **** | 0.005 | 0.025 | 0.248 | 0.258 | 0.262 | 0.533 |
| P11 | 0.211 | 0.233 | 0.218 | 0.202 | 0.222 | 0.164 | 0.213 | 0.251 | 0.134 | 0.014 | **** | 0.035 | 0.232 | 0.237 | 0.245 | 0.591 |
| P12 | 0.210 | 0.236 | 0.217 | 0.151 | 0.183 | 0.134 | 0.160 | 0.195 | 0.099 | 0.030 | 0.038 | **** | 0.146 | 0.152 | 0.155 | 0.457 |
| P13 | 0.229 | 0.258 | 0.232 | 0.161 | 0.206 | 0.142 | 0.171 | 0.216 | 0.147 | 0.156 | 0.147 | 0.097 | **** | 0.002 | 0.001 | 0.593 |
| P14 | 0.228 | 0.260 | 0.237 | 0.154 | 0.188 | 0.145 | 0.165 | 0.203 | 0.143 | 0.158 | 0.151 | 0.102 | 0.015 | **** | 0.001 | 0.591 |
| P15 | 0.229 | 0.247 | 0.237 | 0.158 | 0.196 | 0.134 | 0.163 | 0.208 | 0.145 | 0.155 | 0.155 | 0.097 | 0.014 | 0.014 | **** | 0.590 |
| P16 | 0.230 | 0.244 | 0.228 | 0.275 | 0.347 | 0.196 | 0.217 | 0.241 | 0.229 | 0.286 | 0.306 | 0.238 | 0.300 | 0.299 | 0.318 | **** |
| Population | Population Codes | Group | Sample Size | Latitude (N) | Longitude (E) | Average Altitude (m) | Province |
|---|---|---|---|---|---|---|---|
| Zhanhe | P6 | G1 | 30 | 48°1′23″ | 127°0′56.16″ | 358 | Heilongjiang |
| Heihe | P4 | G1 | 30 | 49°24′47″ | 126°36′47.16″ | 131 | Heilongjiang |
| Liangshui | P5 | G1 | 30 | 47°43′48″ | 128°55′12″ | 231 | Heilongjiang |
| Tieli | P7 | G1 | 29 | 47°58′48″ | 128°4′48″ | 213 | Heilongjiang |
| Hegang | P8 | G1 | 30 | 47°21′29″ | 129°33′50.03″ | 63 | Heilongjiang |
| Liangzihe | P1 | G2 | 30 | 47°1′54″ | 129°41′49.92″ | 500 | Heilongjiang |
| Lushuihe | P3 | G2 | 30 | 42°24′00″ | 127°28′59.88″ | 732 | Jilin |
| Linjiang | P9 | G2 | 30 | 42°0′36″ | 127°13′12″ | 342 | Jilin |
| Helong | P2 | G2 | 30 | 42°32′47″ | 129°1′3.72″ | 452 | Jilin |
| Wangqing | P16 | G2 | 31 | 43°19′26″ | 129°45′14.04″ | 232 | Jilin |
| Muling | P13 | G2 | 30 | 43°49′54″ | 129°45′19.08″ | 410 | Jilin |
| Jiaohe | P10 | G2 | 30 | 44°0′36″ | 127°25′12″ | 196 | Jilin |
| Maoershan | P14 | G2 | 30 | 45°16′22″ | 127°30′14.40″ | 536 | Heilongjiang |
| Hulin | P11 | G2 | 30 | 45°46′12″ | 132°58′48″ | 84 | Heilongjiang |
| Boli | P12 | G2 | 30 | 45°42′00″ | 130°25′12″ | 525 | Heilongjiang |
| Fangzheng | P15 | G2 | 30 | 45°49′48″ | 128°48′00″ | 111 | Heilongjiang |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhao, M.; Xu, Y.; Li, Y.; Tigabu, M.; Zhao, X. Genetic Diversity and Population Differentiation of Pinus koraiensis in China. Horticulturae 2021, 7, 104. https://doi.org/10.3390/horticulturae7050104
Li X, Zhao M, Xu Y, Li Y, Tigabu M, Zhao X. Genetic Diversity and Population Differentiation of Pinus koraiensis in China. Horticulturae. 2021; 7(5):104. https://doi.org/10.3390/horticulturae7050104
Chicago/Turabian StyleLi, Xiang, Minghui Zhao, Yujin Xu, Yan Li, Mulualem Tigabu, and Xiyang Zhao. 2021. "Genetic Diversity and Population Differentiation of Pinus koraiensis in China" Horticulturae 7, no. 5: 104. https://doi.org/10.3390/horticulturae7050104
APA StyleLi, X., Zhao, M., Xu, Y., Li, Y., Tigabu, M., & Zhao, X. (2021). Genetic Diversity and Population Differentiation of Pinus koraiensis in China. Horticulturae, 7(5), 104. https://doi.org/10.3390/horticulturae7050104