1. Introduction
The migration of cells is of great importance in several biological processes, such as embryogenesis, wound healing and, more relevantly, the immune system [
1,
2,
3]. Macrophages are cells of the immune system that filter foreign particles when settled in lymphoid tissues and the liver [
1]. In homeostasis, or the tendency to an equilibrium of physiological processes, the role of macrophages ranges from tissue repair through to immune responses to pathogens [
4]. However, excessive migration can be related to autoimmune diseases and cancer [
5]. The common fruit fly,
Drosophila melanogaster, has been widely studied as a model organism on developmental and cellular processes relating to other organisms, including humans [
6]; such investigations have led to insights into how macrophages integrate external cues into migration [
2]. For example, in [
7,
8], previously unrevealed dynamics of cytoskeletal structures in macrophages were discovered; where certain events of cell–cell contact appeared to anticipate migration.
Cell tracking is defined as the linking between detected objects in one time frame to the objects in subsequent frames. In this work, tracking is defined as a function of segmentation, the correct identification of cells from the background and from each other. Both cell segmentation and tracking have been widely studied [
9,
10,
11,
12]. Cell-tracking studies of phase-contrast microscopy time sequences have been presented in [
9,
10], showing quantitative analysis of cell dynamics in vitro. In [
11,
12], several tracking algorithms were evaluated with a series migratory cells with very different conditions, not only in their ability to track detected objects, but also to identify important cellular events, like mitosis. Other cellular events, such as the interactions between cells are also of importance. To assess such events, a more thorough study of a tracks’ features is required.
The analysis of movement, defined as the analysis of track features, is performed to answer a specific research question about the phenomenon studied. For example, in [
13], tracks are classified based on their features such as speed and curvature. In a related implementation, an automated analysis of movement patterns provided a toxicological assessment of the environment [
14]. In this assessment, the tracks produced by the movement of marine crustaceans were examined to detect specific patterns of motion indicating levels of toxicity in the environment. Finally, contributions regarding the specific data analyzed in this work have been varied. Segmentation of macrophages in single frames was presented in [
15], showcasing the complex interactions which manifest as overlapping (
clumps). In [
16], the relationship between contiguous frames was incorporated to the segmentation of single cells, allowing for a controlled measurement of shape parameters between overlapping events.
In this work, a software framework to analyze the movement and the changes of shape of fluorescently labelled macrophages is presented. The framework, called
macrosight, consists of two stages. The first stage segments and tracks the cells [
15,
17]. The second stage, contains two types of analyses from the tracking results: (a) the shape tracking described in [
16] and (b) the analysis of interactions introduced in [
18]. In the latter, overlapped
clumps are considered moments of assumed interaction between the cells and thus the movement before and after contact is analyzed.
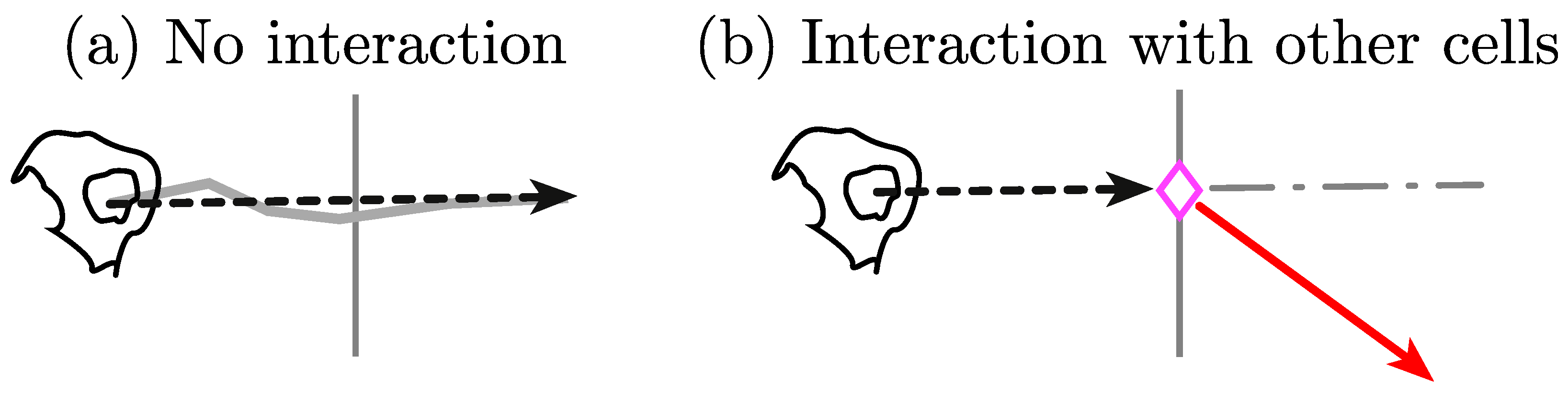
The two main hypotheses of this work are (i) that cell–cell contact accounts for an interaction between cells, and (ii) as a result of an interaction, one or both cells involved in the interaction will noticeably change the direction before and after contact.
Figure 1 shows a graphical abstract of the main contribution of the direction changing in the algorithm.
A preliminary version of this work was presented at the 22nd Medical Image Understanding and Analysis (MIUA) [
18]. The algorithms have been extended and several new experiments with new data are presented. Thus, this work now describes the following topics, not included previously: (i) description of the
macrosight framework, a complete framework for the analysis of movement of macrophages; (ii) a thorough descriptive statistical analysis of the cell–cell interactions, providing insights on interactions occurring after cell contact; (iii) more experiments performed on two additional datasets, comprising an increment of the sample size from 20 to over 50 cases analyzed.
2. Materials
Three different experiments of fluorescently labelled macrophages of the model organism
Drosophila melanogaster were observed, producing three distinct videos of migrating macrophages. In each of the experiments, the nuclei were labelled with CFP-Moesin, while the microtubules of the cells were labelled with Clip-GFP, a microtubule probe; the complete imaging methodology is thoroughly described in [
7,
8]. The images were obtained at a pixel density of 0.21
m/pixel and every 10 s. From the three datasets, RGB images were created by putting the nuclei into red layer and the microtubules into the green layer, resulting in a size of
and two layers of fluorescence. For a simpler identification, the datasets will be named as
MACROS1,
MACROS2 or
MACROS3, where the number of frames per dataset is 541, 361 and 462 frames, respectively.
Overlapping events, referred to as
clumps, are relevant to the study of the datasets.
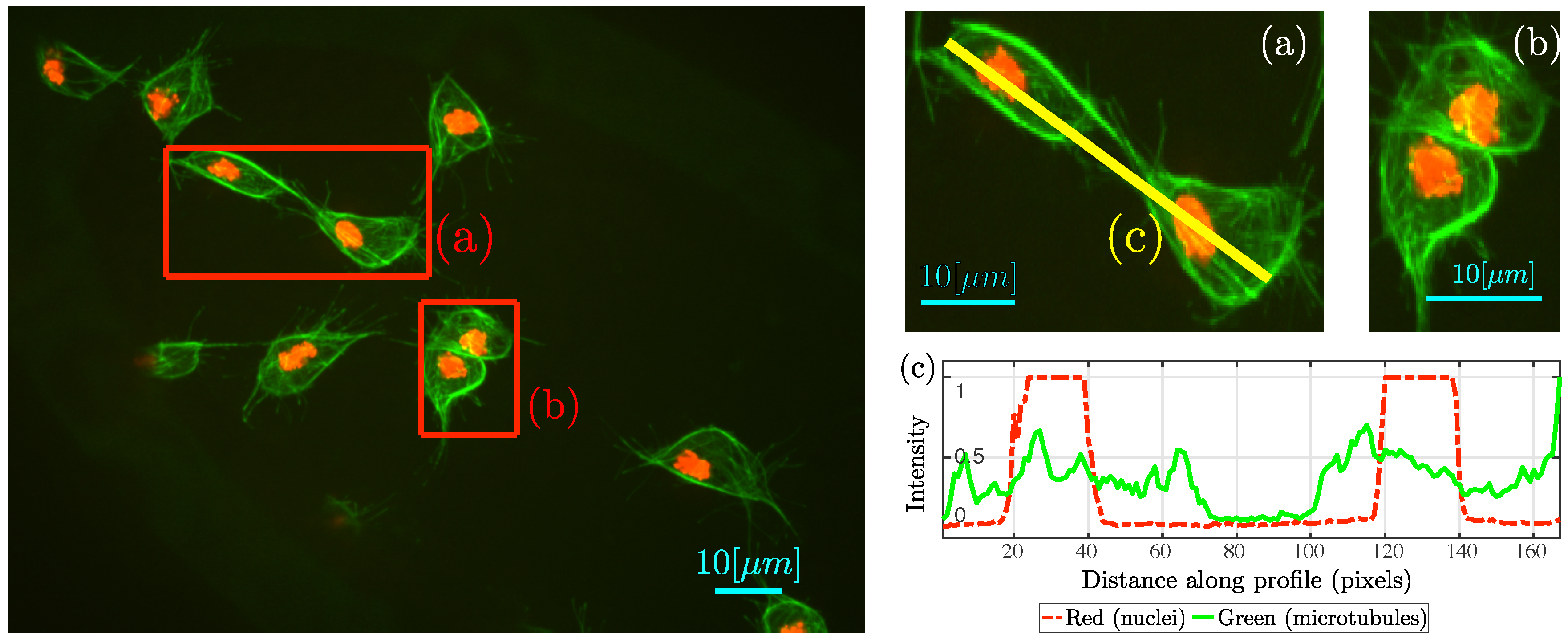
Figure 2 contains an example frame and detail of
clumps where cell–cell contact can be appreciated in two distinct ways:
Figure 2a shows a pair of cells where microtubular structures align;
Figure 2b shows two cells completely overlapped. Empirical observation of the data in [
8] suggest that cell–cell contact, similar to the one showed in
Figure 2a, could influence the migration pattern of the cells involved, with interactions considered to be under 3 min (18 frames). For example, the cells shown in
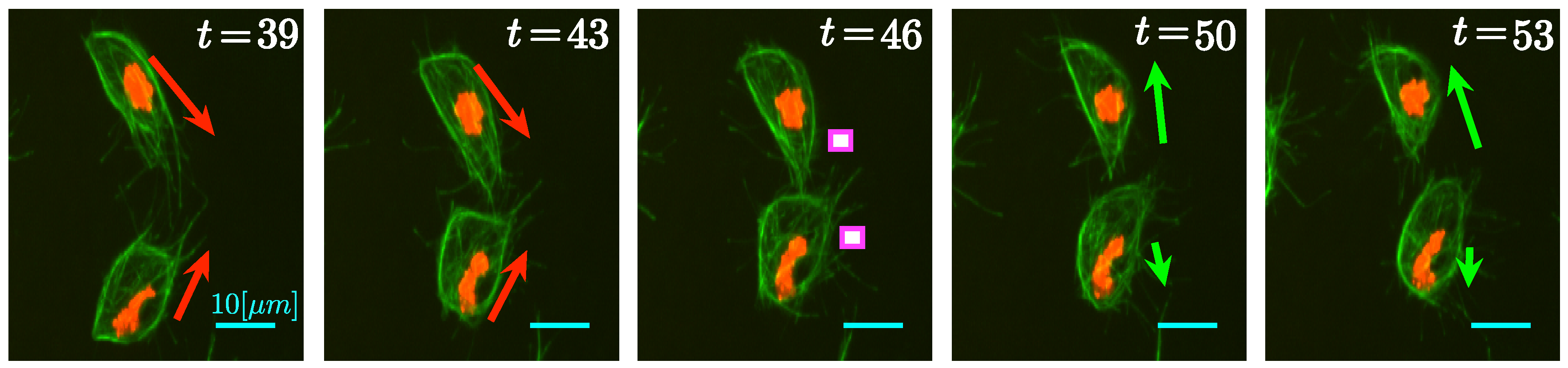
Figure 3, where a series of frames is shown, taken from two of cells that overlap and then appear to change direction over 15 frames.
3. Methods
In this section, the main functionalities of
macrosight, in the context of direction changes will be presented. Special emphasis will be given to the algorithms analyzing changes of cell direction as a result of cell–cell contact, which will be the focus of this work. In a previous development [
15],
clumps were studied and how to separate them. In this work,
clumps will be considered points of interaction between cells. In
Figure 3, arrows indicate the observed direction of movement of the cells. Notice the green microtubules aligning in the top cell on frames
and
, just before and during the contact between cells. Such alignment has been reported [
8] to indicate the change of trajectories between the cells in the following frames.
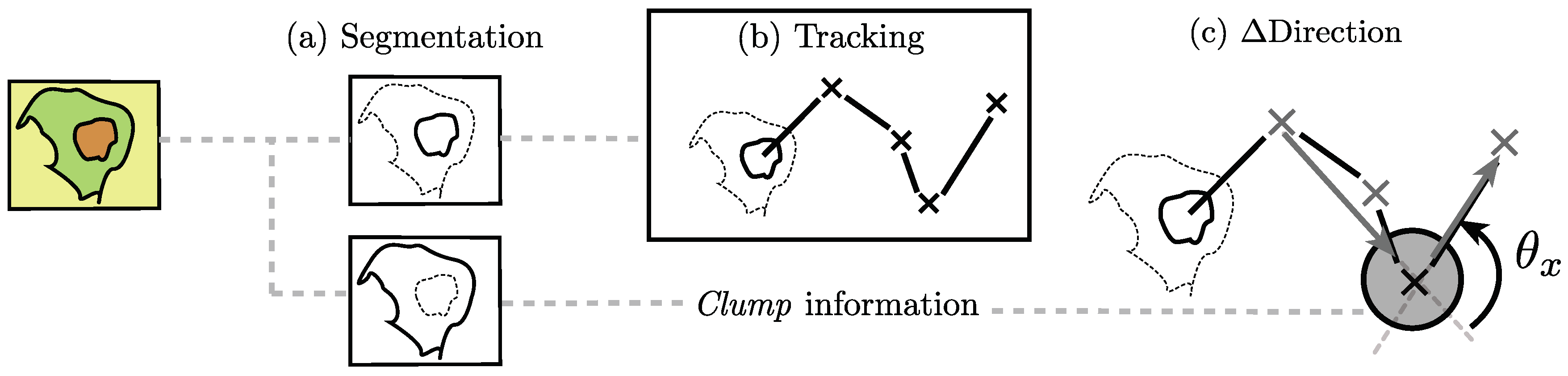
The methodology for cell–cell contact influence in the change of direction can be divided into four stages of analysis. First, the segmentation of each channel individually. Then, the tracking of the objects detected in the red channel is performed, and the detections of each channel are classified as
clumps or single cells. Finally, for each track, the change of direction (
Direction) is found before and after a given
clump.
Figure 4 shows a graphical representation of the procedures carried out in this work. Each stage is detailed in the following sections.
3.1. Segmentation of Fluorescence Intensity Channels
The segmentation procedure for segmenting each of the respective channels follows three steps, fully described in [
15]. Following a low-pass filtering, each channel was segmented by a hysteresis thresholding technique [
17]. Finally, a morphological opening with a disk structural element
was performed to remove noise and smooth the edges. Detection of
clumps was achieved by comparing the number of nuclei detected within the area covered by each segmentation of the green channel.
3.2. Tracking of the Nuclei and Incorporation of Clump Information
Tracking is defined as the linking of detected objects between consecutive time frames, namely: parent (
), present (
t) and child (
). Following segmentation, the tracking of the objects in the RED channel was performed through the PhagoSight software [
17,
19], a framework for cell tracking that uses the Keyhole algorithm [
20,
21]. The
macrosight framework incorporates the segmentation of the RED channel to PhagoSight, for tracking, and incorporates the information of the
clumps in the GREEN channel after the tracks have been calculated.
The Keyhole Tracking Algorithm. The algorithm [
17,
20] links the segmented objects in contiguous frames through the analysis of the velocity and direction of the object at frames
and
t, and estimates the position of the object at frame
. Apart from the estimation, the algorithm generates two regions of probability, which resemble an old-style keyhole, to anticipate changes in trajectory. Each track produced by PhagoSight includes the information of the cell’s nuclei that has been tracked and linked from one frame to others.
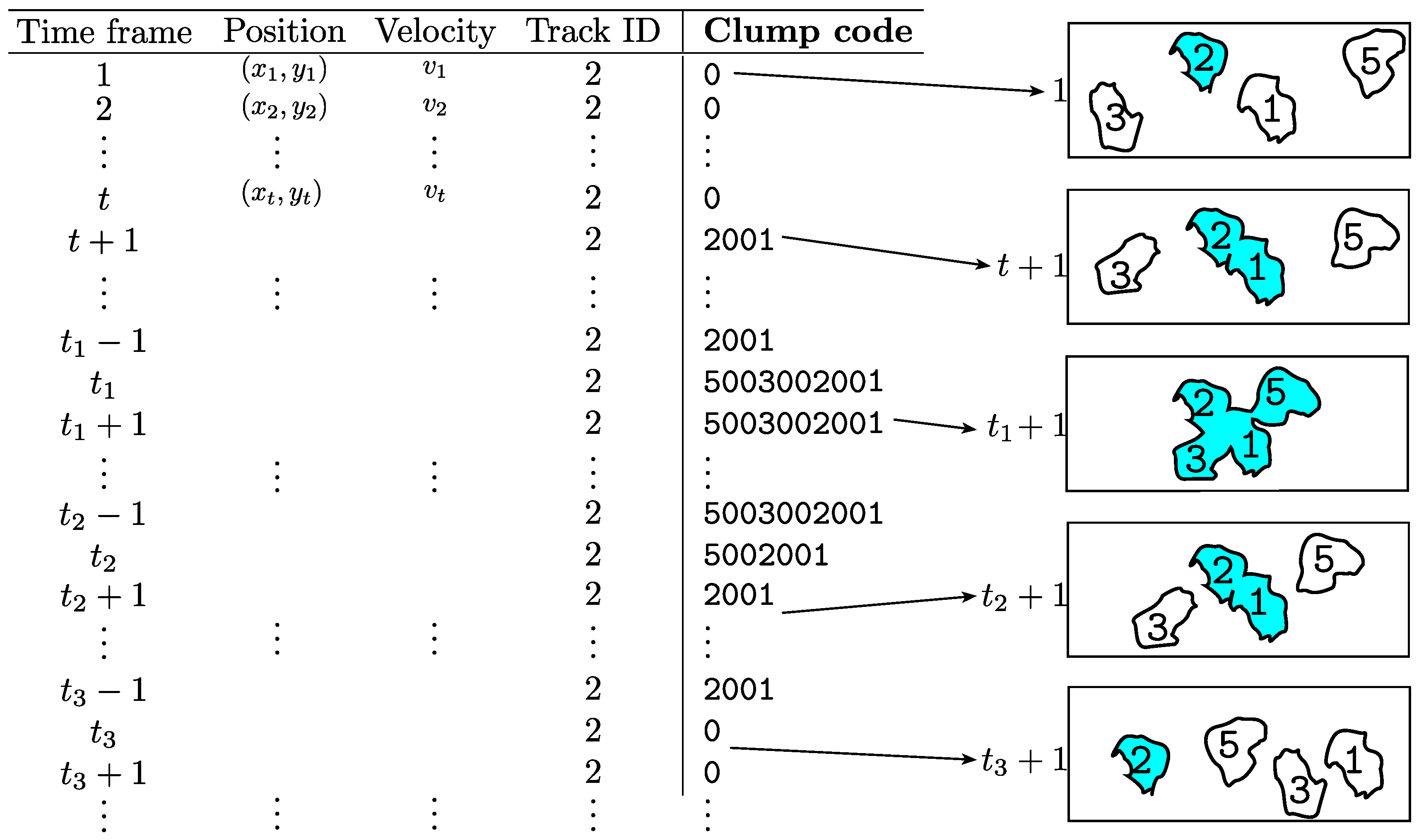
Addition of clump information bymacrosight. The information of interest to this work is explained in
Table 1, it includes parameters such as time frame, position, and velocity of each nuclei at each time frame. At each point in time, the presence of
clumps was detected by counting the number of nuclei contained within a single object detected in the green channel. A new parameter, called clump code, has been incorporated to
Table 1. Each nucleus within a
clump has a track associated with them, thus each
clump can be uniquely identified via a simple unique identifier number or for short
clump code, which includes the labels of the tracks contained within it. For instance, let
be the labels of two tracks (
) which at a certain point in time belong to a
clump, then the code
c is defined by
. The value of 1000 is chosen arbitrarily as a large number, larger than the total number of tracks. Notice how the tracks’ labels are arranged from left to right starting with the highest identifier to the lowest; for example, code
24013 would correspond to a
clump that at a certain frame contains tracks 24 and 13.
The previous definition can be extended for an arbitrary number of labels
m interacting in the same
clump as
, where all labels in the
clump are ordered
. Each
clump can be uniquely identified based on the tracks contained in it.
Table 2 shows a simple example of the creation of the
clump codes. The inclusion of the codes facilitates the analysis of the cells that interact with each other. four cells
Figure 5 represents the information from each track, as produced by PhagoSight, where the
clump code is incorporated by
macrosight. The information of a single track (track ID = 2) is shown, at different time frames. When the cell is not part of a
clump, the variable
clump code has a
0. At a specific frame
, the cells in the green channel come in contact, forming
clump 2001. At frame
, two more cells come in contact with
clump 2001, thus forming
clump 5003002001.
3.3. Measuring the Change of Direction before and after a Clump
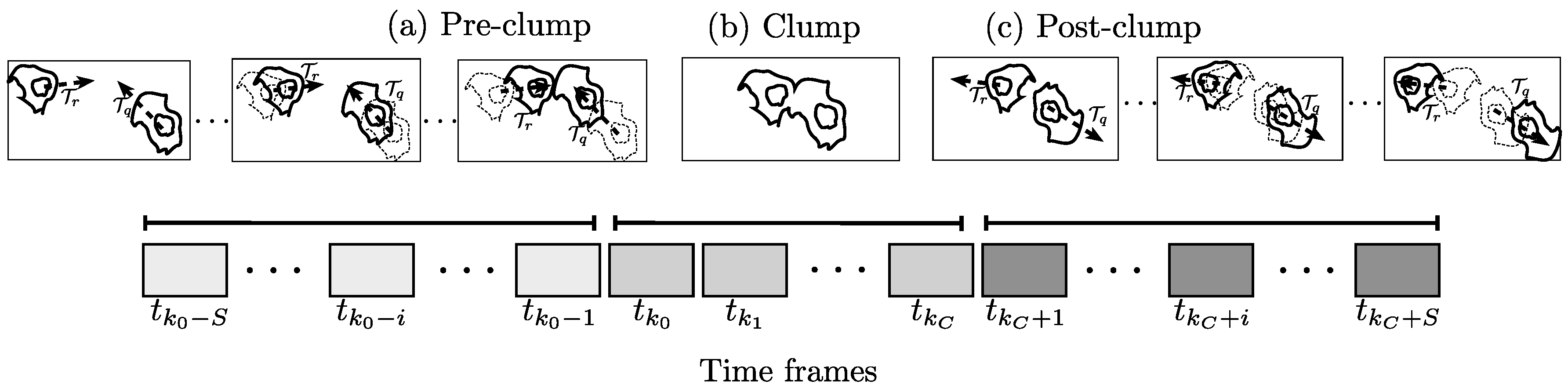
The algorithm developed in this work, estimates the angle formed between the direction of the cell prior to an interaction (clump) and the direction of the cell once the interaction is over and it does not belong to any clump. Let be the angle that measures the direction change ( direction). Let a track with label r, given by , interact with another through a clump with code , such that the overlap between the two cells happens at time frames . Let be the angle that measures the direction change ( direction). Let a track with label r, given by , interact with another through a clump with code , such that the overlap between the two cells happens at time frames .
The determination of
involves analyzing the tracks
starting
S frames before
and finalizing at
S frames after
. Frames
will be referred to as the
clump span; likewise, the time frames where the tracks are interacting,
, will be referred to as
time in clump.
Figure 6 shows a schematic of the tracks analyzed and the choice of the time frames. The relationship between the frames
,
S and
can be clearly observed as the moments in the
clump span containing tracks
and
. The moments are called (a) pre-
clump, (b)
clump and (c) post-
clump.
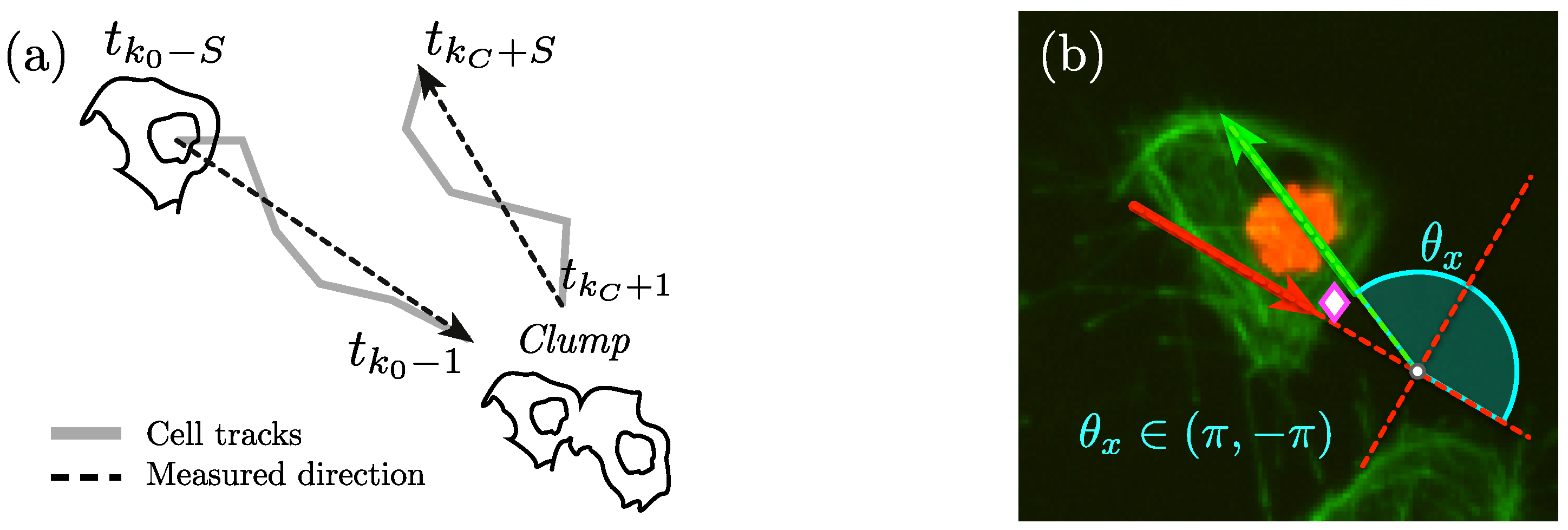
Once the tracks involved, and the span have been manually selected, the calculation of the change of direction angle is done by selecting a vector oriented towards the
clump and another one leaving it.
Figure 7 displays the process of selecting the lines from which direction before and after the
clump will be selected, and the way the angle will be measured. The estimation assesses the change of direction of each cell that exits a
clump, relative to the orientation it has entering it. To calculate the angle, vectors must be aligned and rotated from the original positions in the image
to a new set of rotated axes
. This is performed in an intermediate step, where the incidence angle is calculated and all the points in the track are rotated to the new axes.
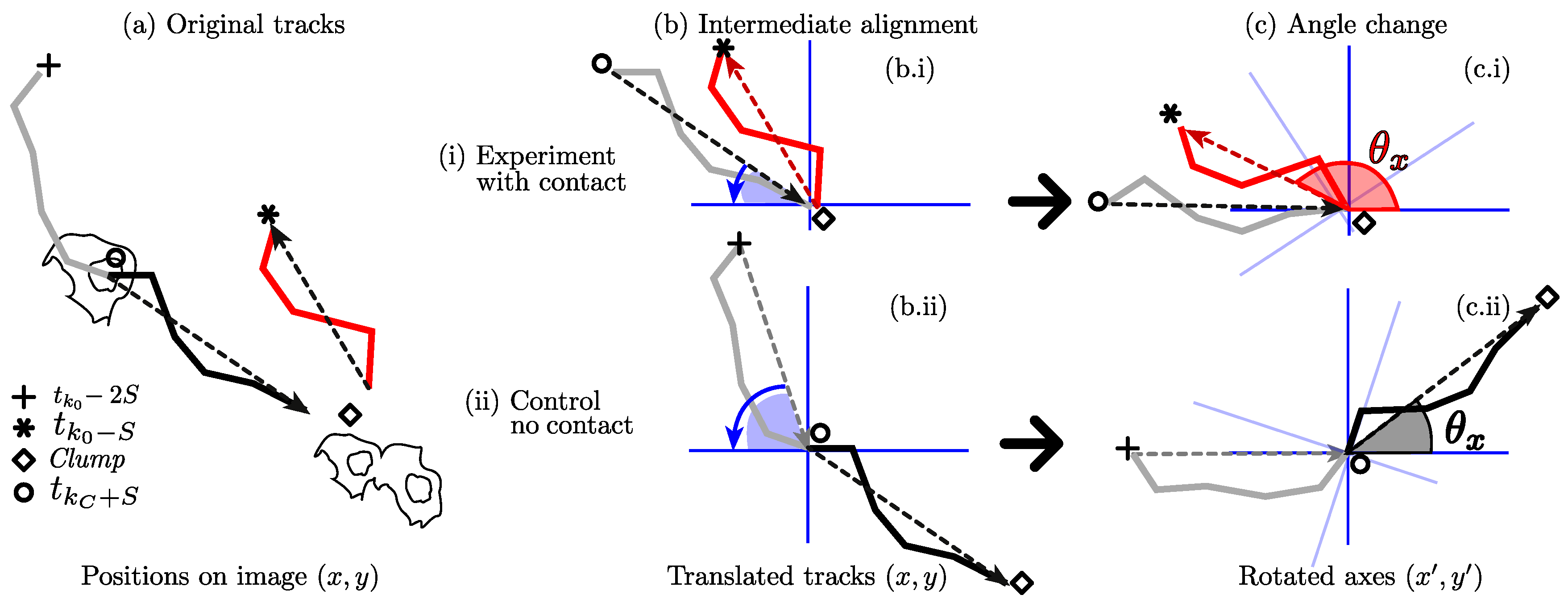
Figure 8 represents the calculation of the angles compared for the interactions and the control experiments. Notice that the new axes
in
Figure 8b can be interpreted as a new frame of reference, containing all the positions rotated and aligned.
3.4. Experiments
All three datasets were segmented in both and tracked. The tracks’ information was searched to find cases of clumps that fulfil the following criteria:
Only two cells interacting. There are cases where more than one cell integrates a clump. These cases were excluded from the analysis as it is not clear whether the interaction of more than two cells would be different from the interaction between a pair.
In and out cases. The cases selected only involved cells with a well-defined clump span, in which the cell would enter the clump and exit it without disappearing or interacting with other clumps.
Immediate reaction. A small value for was chosen to define the clump span, as the interest of this paper is to study the immediate reaction of a cell after interacting within a clump.
Both cells in clump. Cases where both tracks in the clump had a well-defined clump span were preferred as they would allow an analysis per clump.
Once the tracks were selected, the angles were calculated for each case. Additionally, control movements for each track, consisting of the time frames leading up to the clump, were selected to allow a comparison of the change of direction with a cell that has not interacted with another one.
The experiments were chosen in a semi-automatic way, using the information from the table in
Figure 5 to generate candidates of experiments and manually logging the starting and ending points
and
per experiment. It is important to notice that the experiments were chosen without considering the
time in clump. As highlighted in
Section 2, such parameter could be influential to the results, as the window where the interaction is observed occurs within a few minutes, which would be translated to a cell belonging a small number of time frames in
clump.
4. Results
All datasets were segmented and tracked. Tracks were selected based on the criteria described in
Section 3.4. In total, fifty-two cases were found with
respectively for each dataset. To represent the tracks and changes in directions for all datasets and compare them to the control tracks,
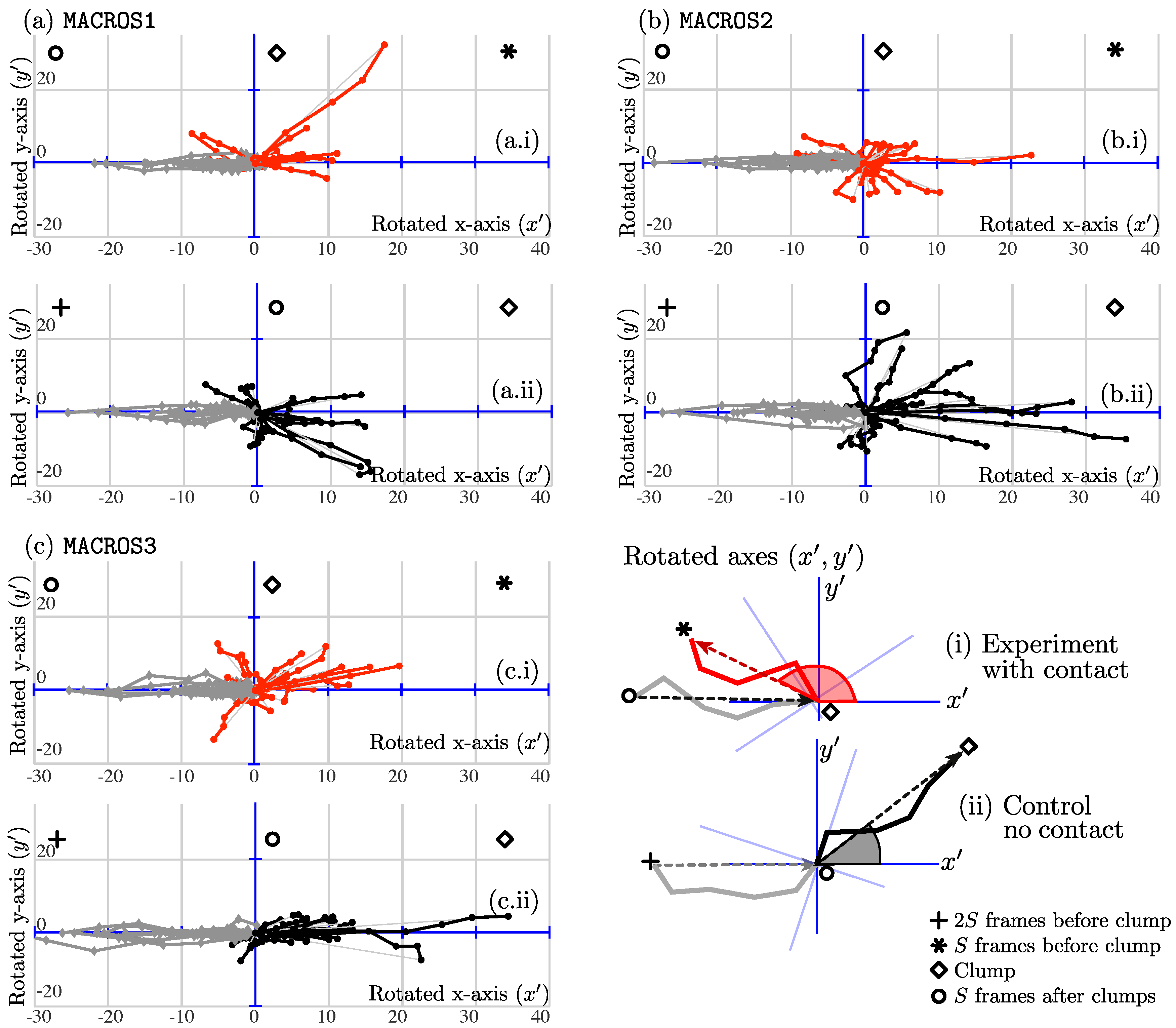
Figure 9 is presented to qualitatively show the hypothesis depicted in
Section 3.4 and
Figure 1. The figure contains a key taken from the explanatory
Figure 8.
Several differences can be observed in the tracks containing cell–cell contact, varying depending on the dataset. To assess the changes of angles of all experiments collected, the mean and median of the angle changes were compared from experiments to control. Referring to
Figure 9,
Table 3 summarizes the angle comparisons made with the mean and standard deviation in each case. The Wilcoxon Signed Rank test [
22] was implemented in all cases, to compare the median of the measurements and a normal
t-Test was implemented to compare the means.
A more thorough exploration of the cases and the tracks was implemented, considering the time in
clump (
) parameter. First, consider the average
per the datasets, where
MACROS1 had an average of
,
MACROS2 had an average of
and
MACROS3 had an average of
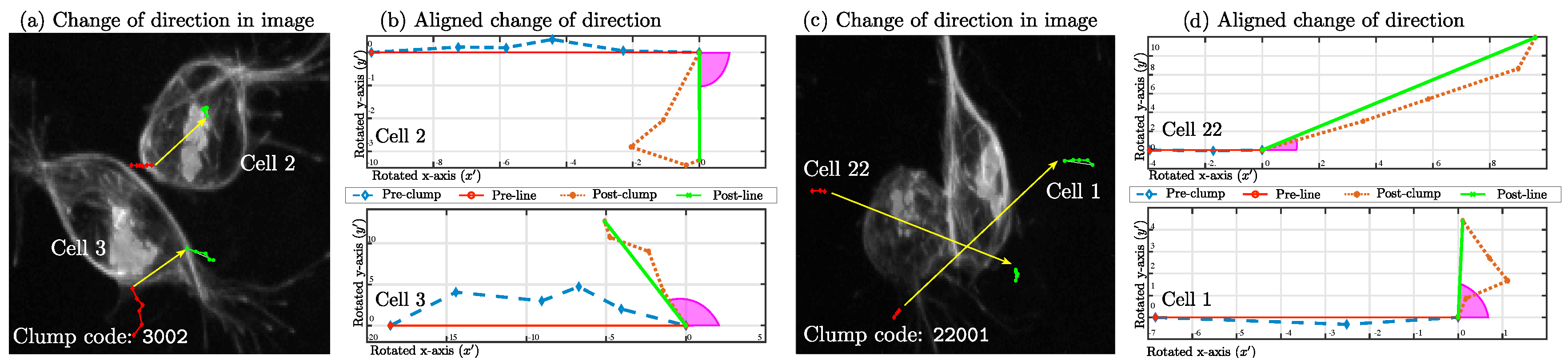
. Visually,
can be observed in
Figure 10, which displays two examples of cells interacting through a given
span and the orientation lines before (red) and after (green) the
clump.
Clumps shown contain the codes
3002 and
22001, which have two very different values of
, as can be observed from the yellow lines.
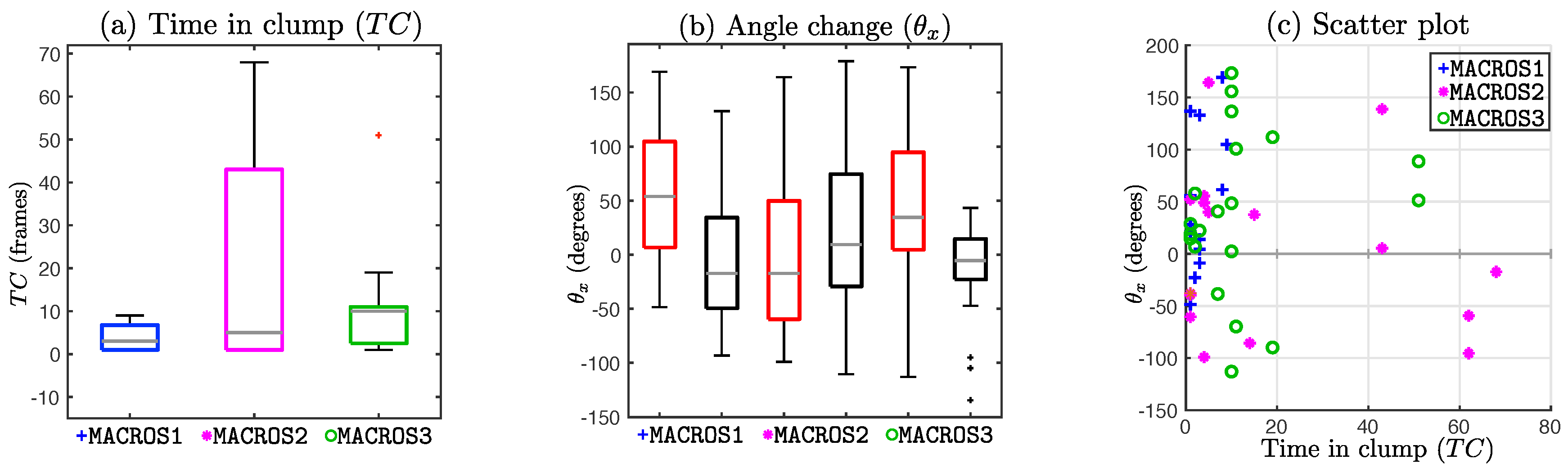
Figure 11 and
Figure 12 explore the differences between change in angle and time in
clump (
). From
Figure 11a, it can be observed that
range for datasets
MACROS1 and
MACROS3 is much smaller compared to the range of the
for dataset
MACROS2. The scatter plot
Figure 11c shows a distinct gap in the range
, and most of the cases in the range
.
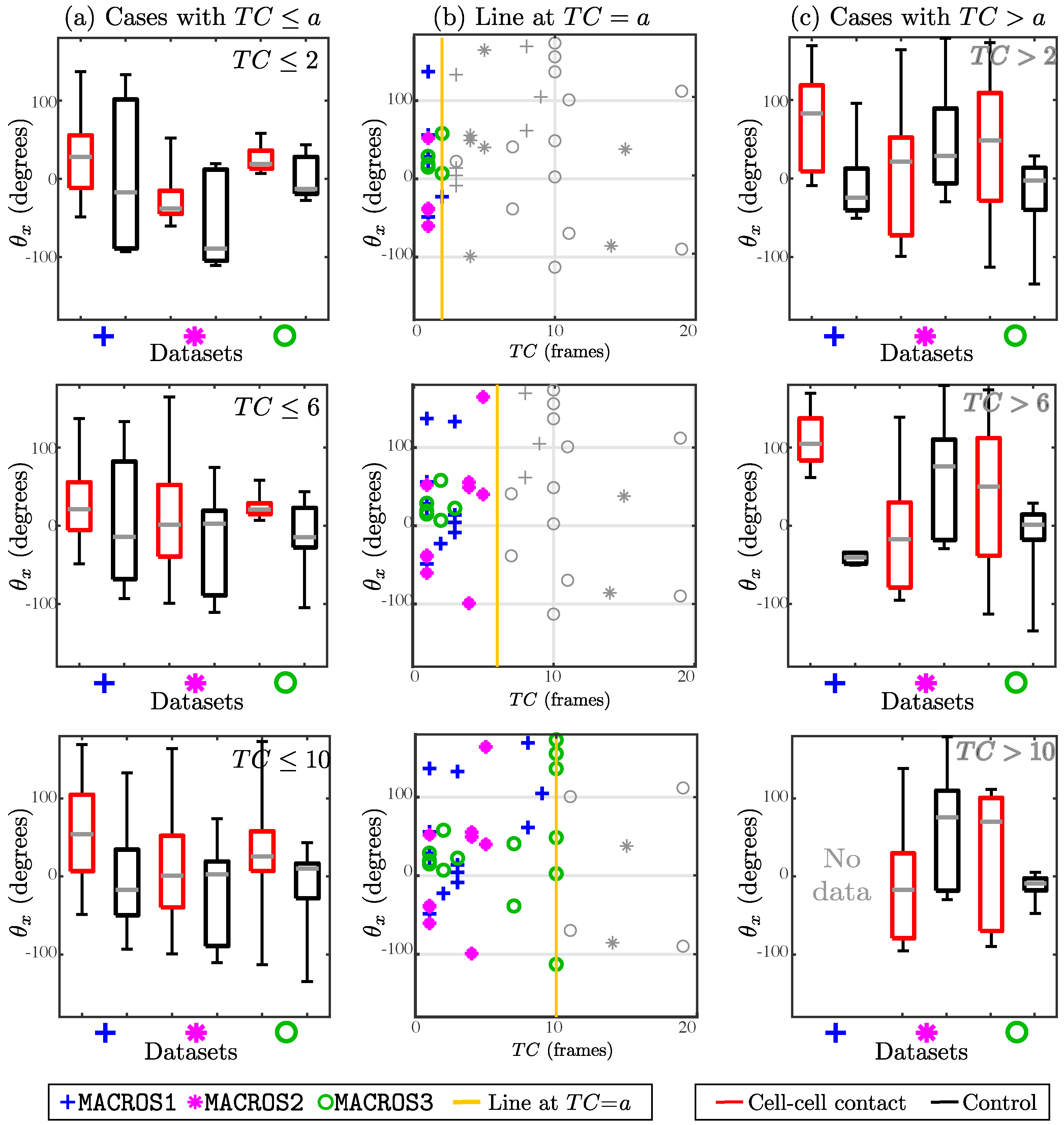
In
Figure 12, a comparison was made between the angle change depending on
. Each row represents the comparison of
between cases with cell–cell contact and control cases, when selecting only cases with
where
and 10. Although most cases are broadly similar, cases with
and
show a more distinctive difference between control and cell–cell contact cases, with control cases (black box plots) showing a greater range in almost all cases. A final experiment was run, in which the value of the angle change
was compared between the control and interaction experiments presenting a
. The cases satisfying
in all datasets were combined, giving a total of 33, where the mean for interaction cases was
and the control cases was
. The Wilcoxon Signed Rank test produced a
p-value of
, providing statistical significance to the difference.
5. Discussion
Preliminary works in this field have provided separate analyses of macrophages data. First, ref. [
15] focusing only in the disambiguation of the lost information of overlapping regions, without including the temporal context. Then, ref. [
16] showed an analysis of the shape evolution of cells that do not overlap. Finally, the initial study to understand macrophages’ direction changes as a consequence of cell–cell contact was presented in [
18]. In this work,
macrosight is presented as a complete framework which includes the routines for data handling, segmentation as well as shape and tracks analysis. In particular, the methodologies described in this work analyze the movement of macrophages exploiting the overlapping of cells observed in the green channel.
This work is an extension to the work shown in [
18]. The principal extension made was the presentation of a full framework of routines. The main algorithm provides insights into the relationship between cell–cell contact events, interactions of cells and movement patterns, as shown by
Figure 9. The figure shows tracks to be different between the two cases. The cases of cell–cell contact show less movement before and after contact, shown by the smaller lines; a higher variability is observed in control cases. Each dataset also presents unique differences in the comparisons. In
MACROS2, for example, the tracks appear more curved after the interaction; and in
MACROS3, the ranges of change in direction are more distinct. Even though the differences between contact and control cases are noticeable, the characterization of the movement should not be reduced to the analysis of a single variable, in this case the angle of changed direction. More variables like speed, curvature could be incorporated. From a biological standpoint, the moment cells come in contact could be determined much sooner if the microtubules with lower intensities are also segmented and tracked.
The time in
clump,
, was found to be a relevant parameter for the measured angle change for different reasons. Firstly, the value of
per
clump appeared to be smaller where statistical significance was achieved, as seen in
Table 3. Secondly, from
Figure 11 and
Figure 12 display the differences in the angle changes and their ranges if the experiments are restricted to keeping a low time in
clump (
). Finally, as mentioned in
Section 2, each frame is taken every 10 s, and the time cells remain in contact should not exceed 1–2 min (
) to be considered an interaction relevant to explore. Through the analysis made in this work, the time in
clump was found to be helpful when assessing the change of direction in values of
. The result was not previously found in [
18], therefore it fuels the need for a more thorough analysis of the tracks’ parameters, attempting to characterize them.
The limitations of the algorithm involved the underlying limitations of the segmentation and tracking methods at dealing with complicated interactions. As mentioned in
Section 3.4, experiments were chosen manually, by looking at the
clump span of each case. Upon verification of the tracks, which consists of manually comparing the segmented nuclei and tracks, the dataset
MACROS1 presented some inconsistencies such as nuclei changing their track identifier, thus complicating the choosing of experiments. The problems with the tracks could be inherited from the limitations of the keyhole algorithm, which only considers velocity and previous direction to estimate the upcoming position. The reader is referred to
Figure 9b.i, where the red tracks appear clearly curved.
The problems could also be due to segmentation, as MACROS1 contains more variability in the intensities than MACROS2 and MACROS3, as well as a larger number of cells interacting in each clump. Future work could improve this by incorporating post-processing to the tracks.
6. Conclusions
This work presents a novel system for the analysis of movement of macrophages and other objects. The main contribution is the integration image analysis techniques into a robust framework to perform automated and semi-automated (such as the selection of the experiments) analyses of movement objects. In particular, this study introduced a methodology for the analysis of movement of macrophages, and the relationship between cell–cell contact and changes in the trajectories of the participating cells.
Despite some encouraging results, the differences shown by macrosight should not be interpreted as conclusive, but as encouraging insights into future research. Some future developments proposed include the following:
- (i)
A deeper understanding is needed in terms of the anticipation before recording an experiment, the number of frames to be taken before and after the start and end of the
clump,
S. As suggested by Stramer et al. [
8], cells appear to elongate before an interaction, thus such elongation could aid in determining the value of
S automatically.
Macrosight already contains the functionality to measure the elongation of the cells, as reported in [
16].
- (ii)
the time in clump, , was introduced, but not studied thoroughly. Clumps could introduce other variables not accounted for, or interactions not quantified
- (iii)
Cells involved in clumps where more than two cells come in contact were not considered, to control the variability of the experiments. However, such inclusion would allow several more cases of study, for instance, dataset MACROS1 contained over 40 cases including the 15 presented in this work.
- (iv)
More importantly, the calculation of the direction was done simply by taking two points before and after the
clump, and only one variable (change of direction) was measured. However, as mentioned before, the tracks have more complex parameters, such as speed, curvature, or acceleration. A thorough characterization of the tracks’ ontological properties, as mentioned in [
13]; could allow for the inclusion of several tracks in the analysis, aiding in a conclusive determination of cell movement changes influenced by contact.

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}