Comparative Study of Contact Repulsion in Control and Mutant Macrophages Using a Novel Interaction Detection †

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials
2.1. Fly Stocks and Preparation
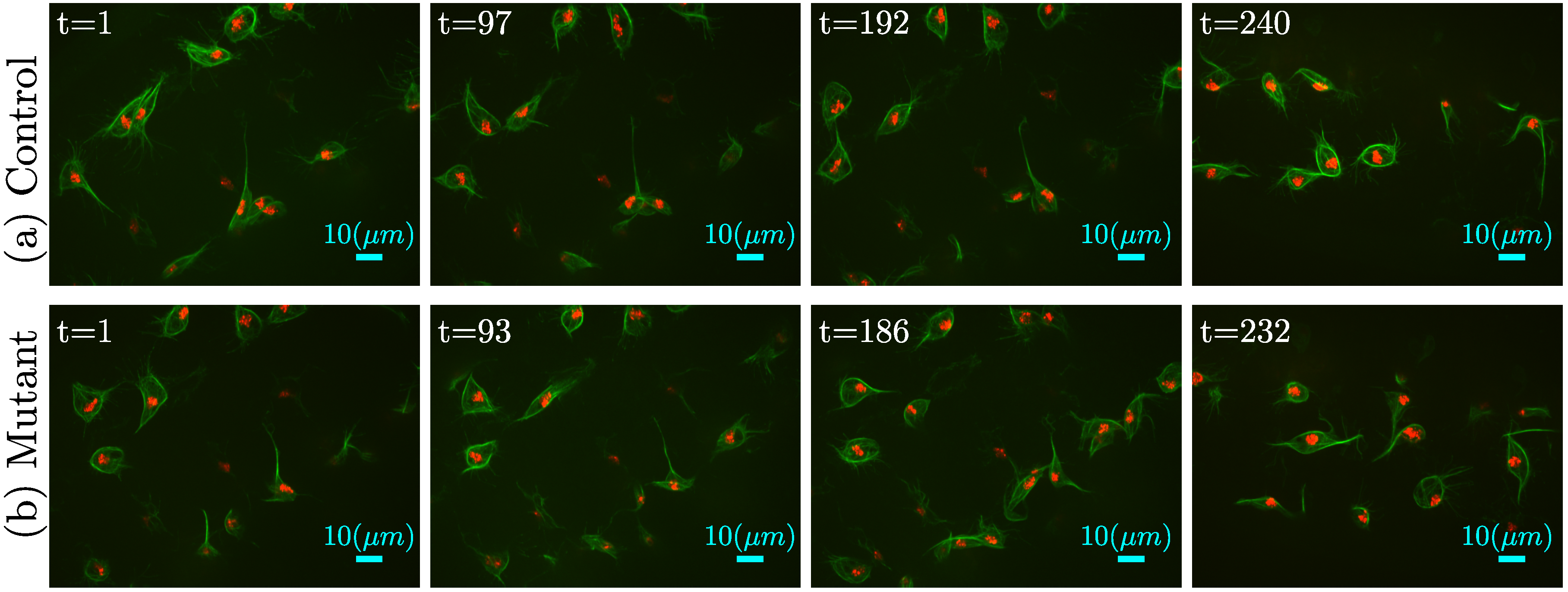
2.2. Microscopy
3. Methods
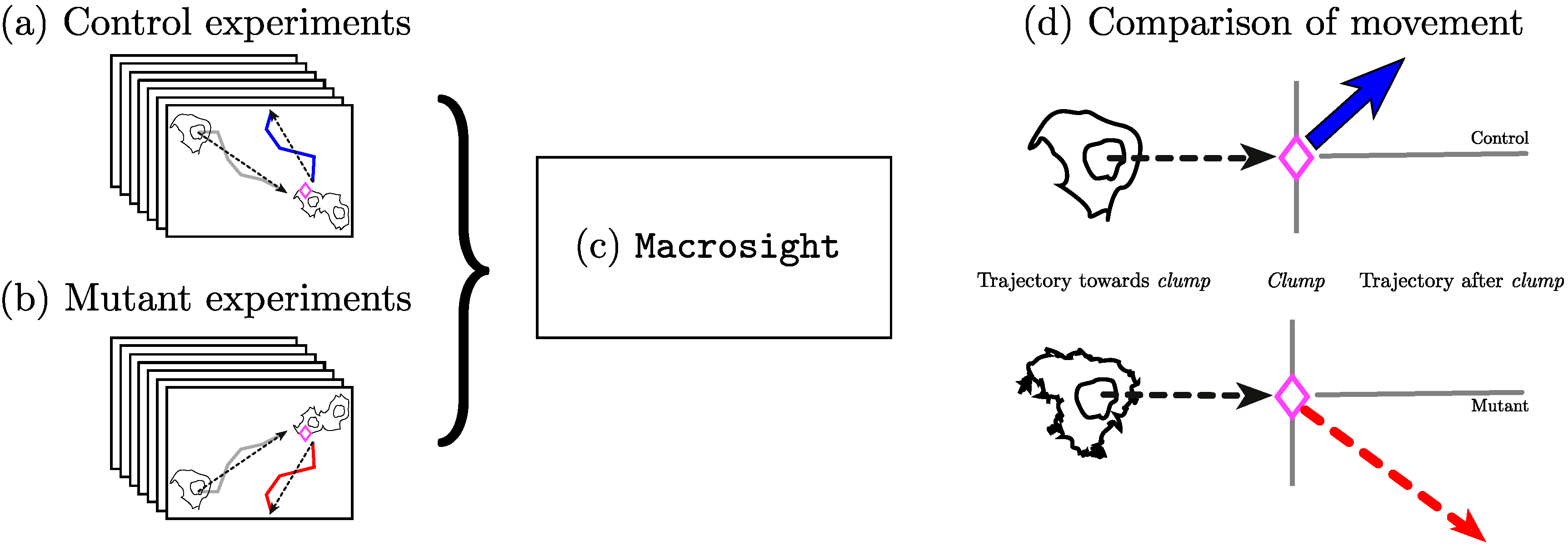
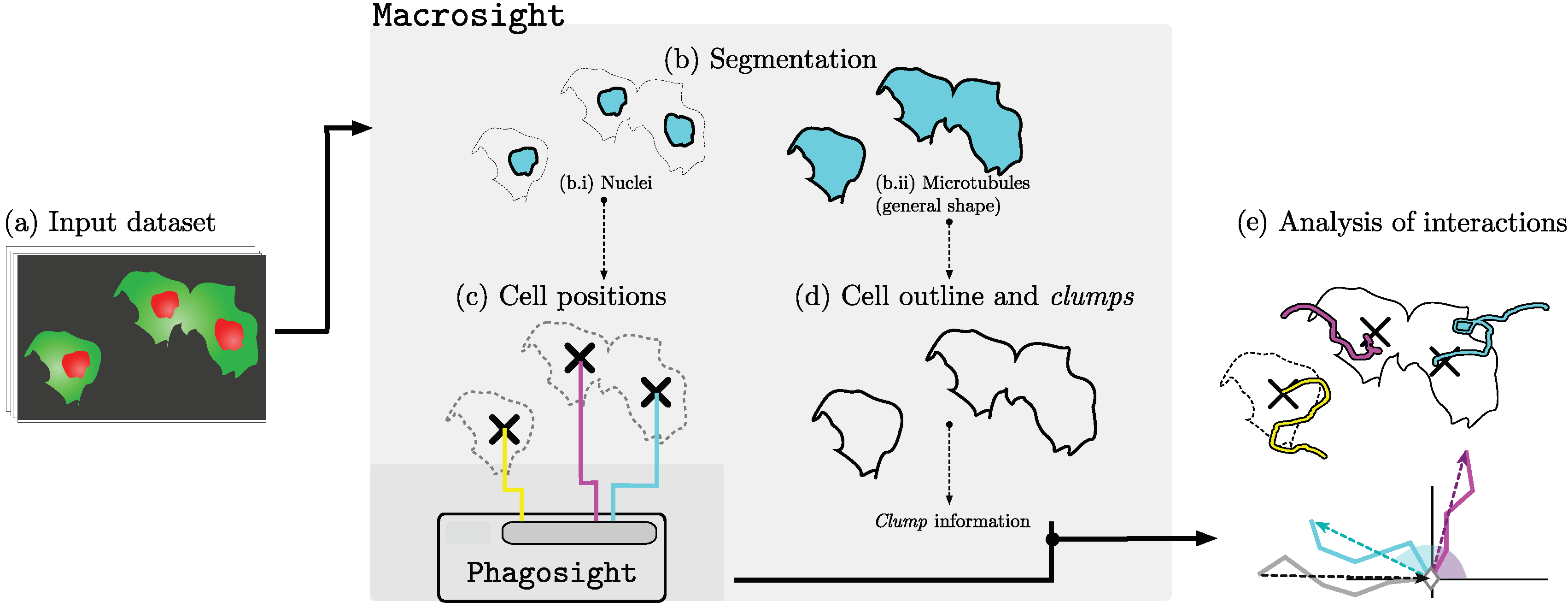
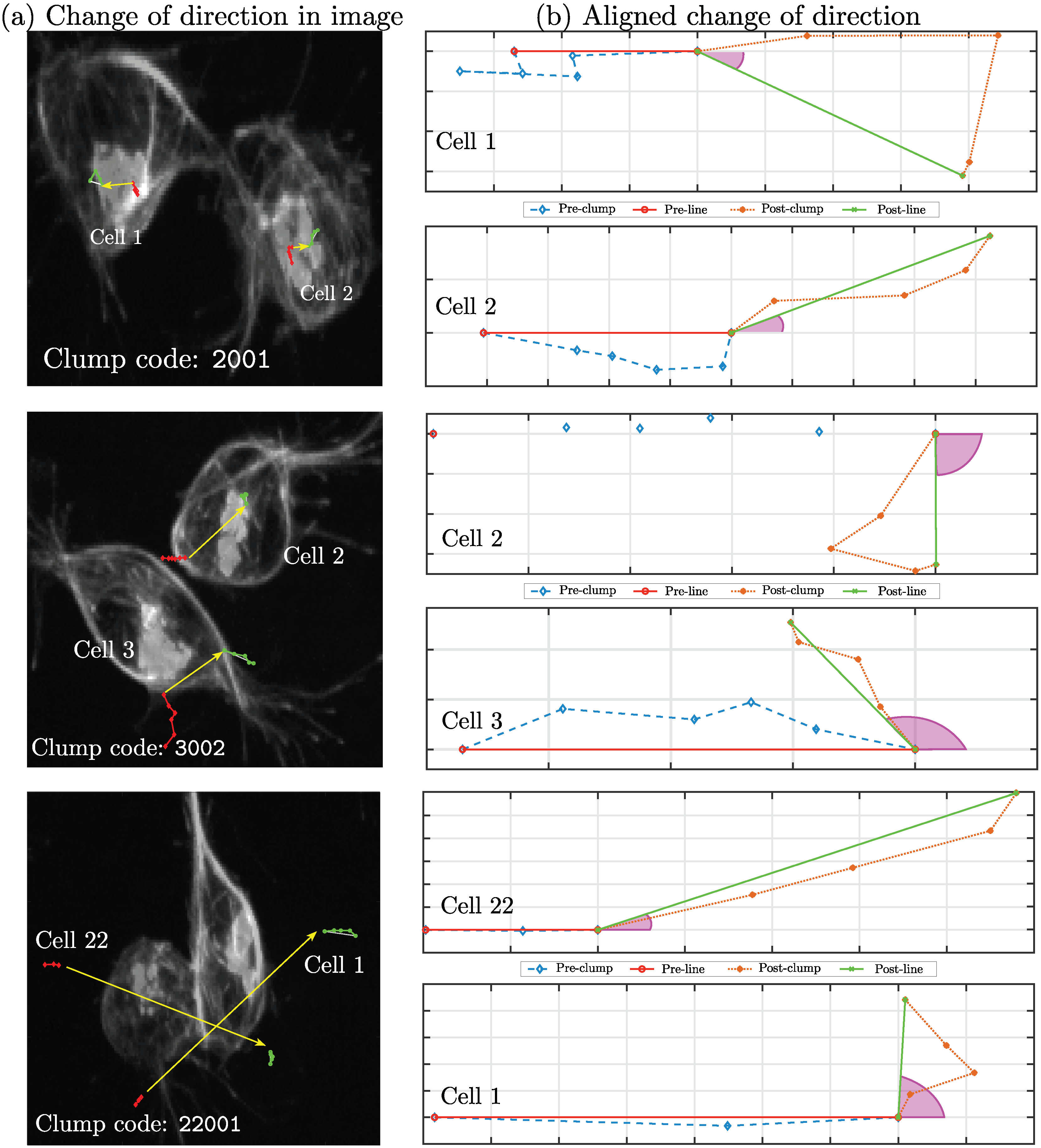
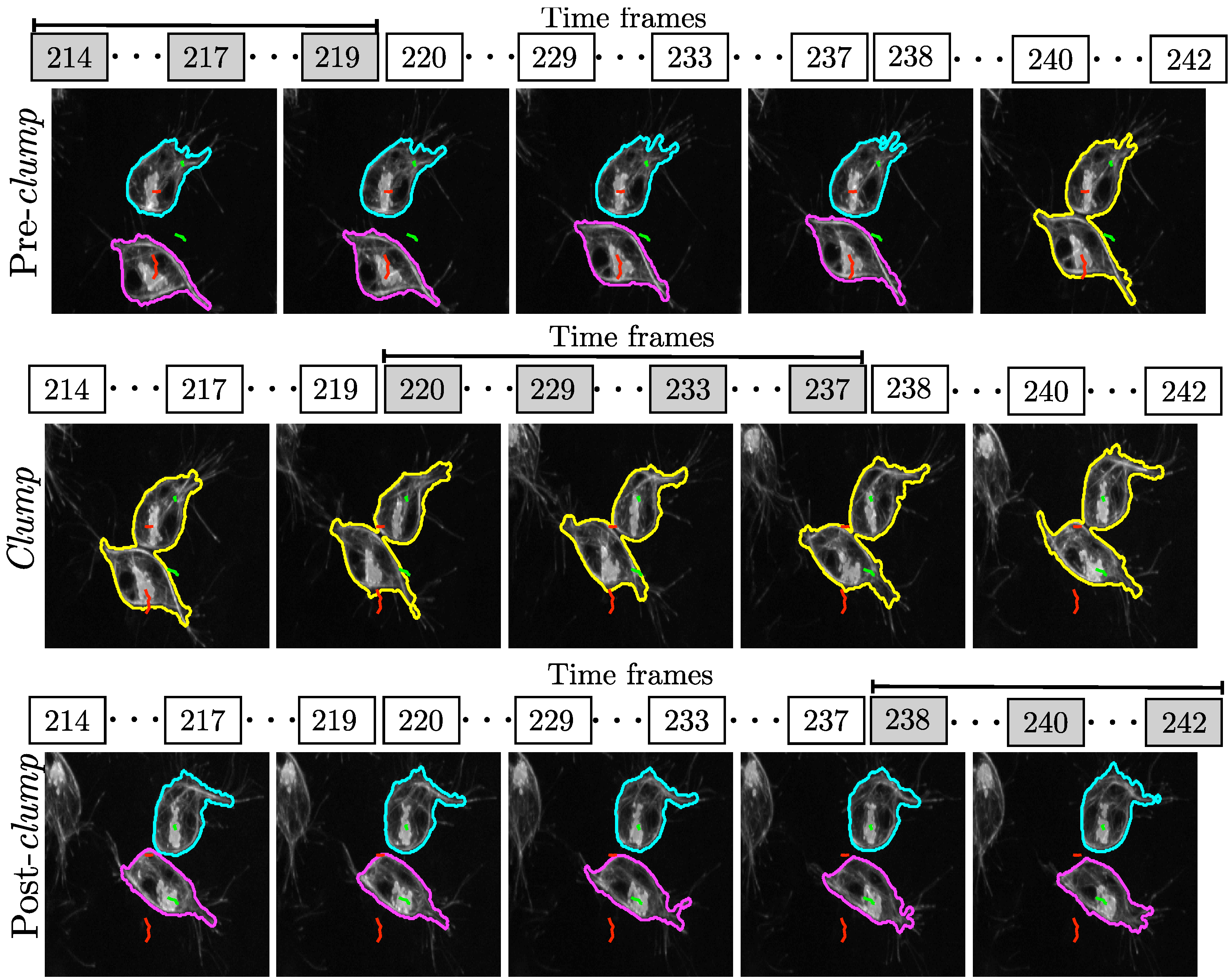
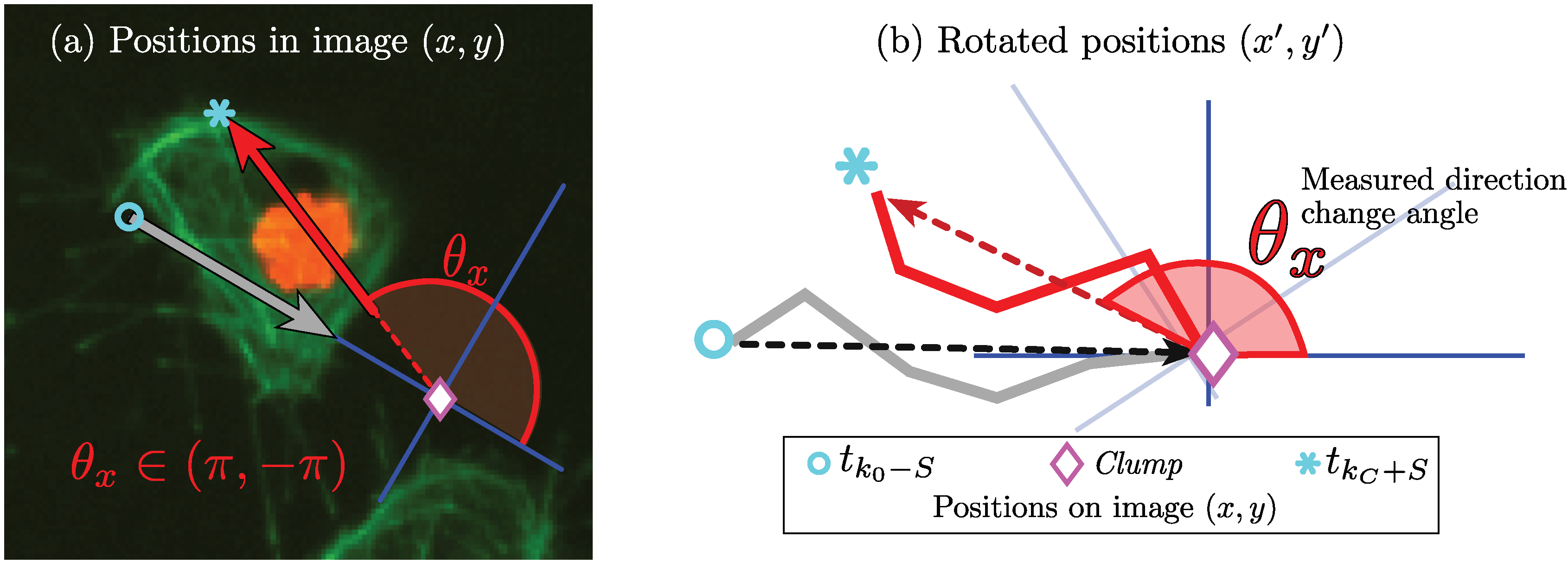
3.1. Analysis of Movements and Interactions
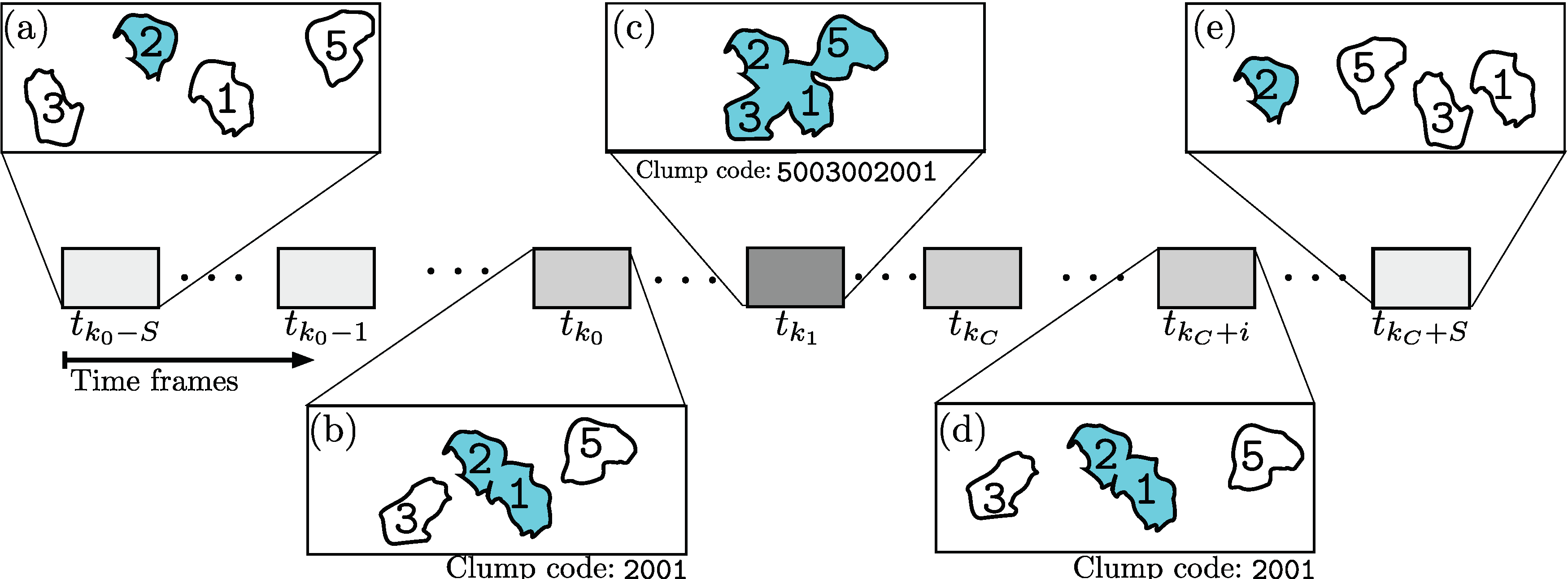
3.2. Selection of Interactions
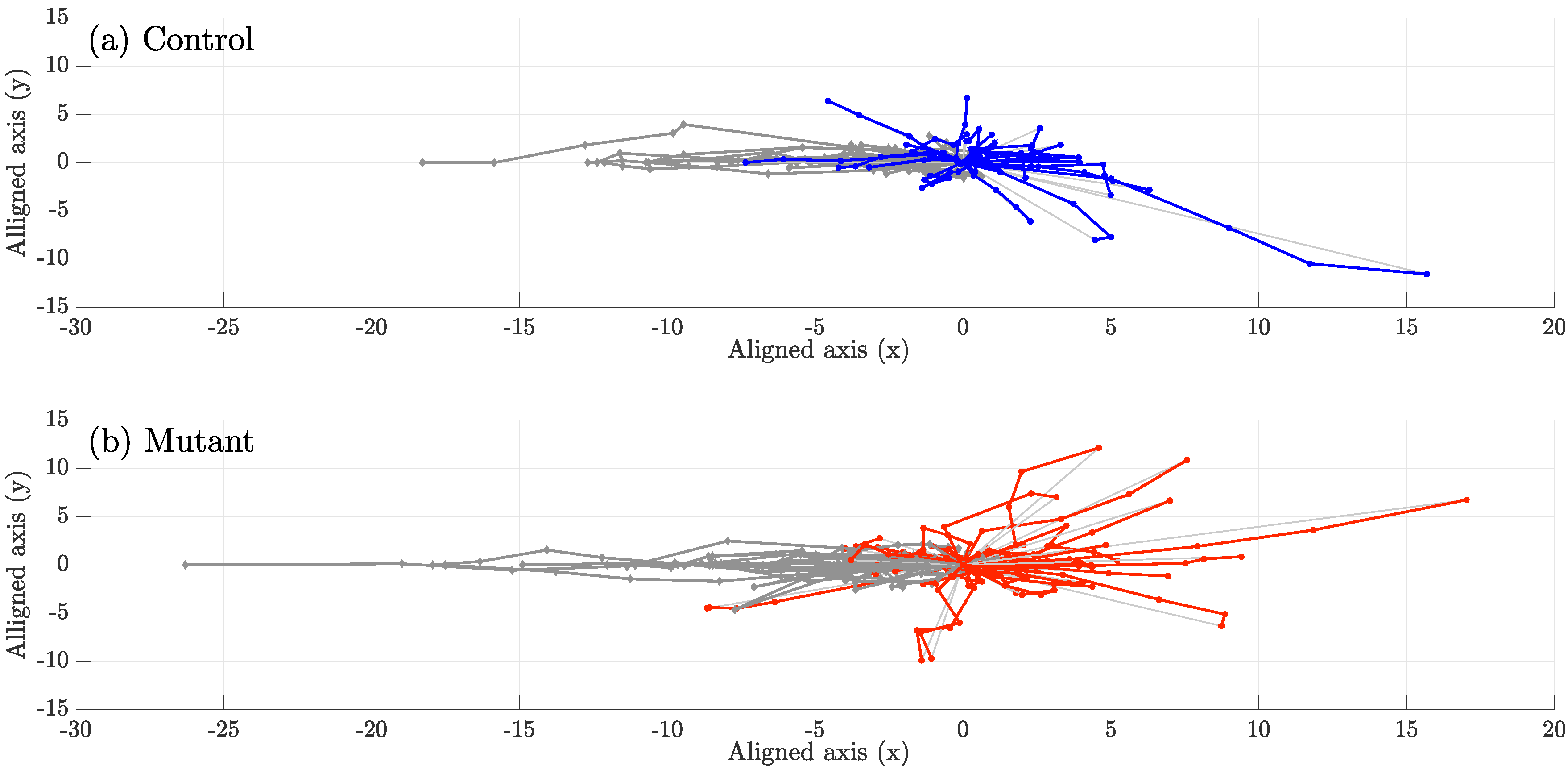
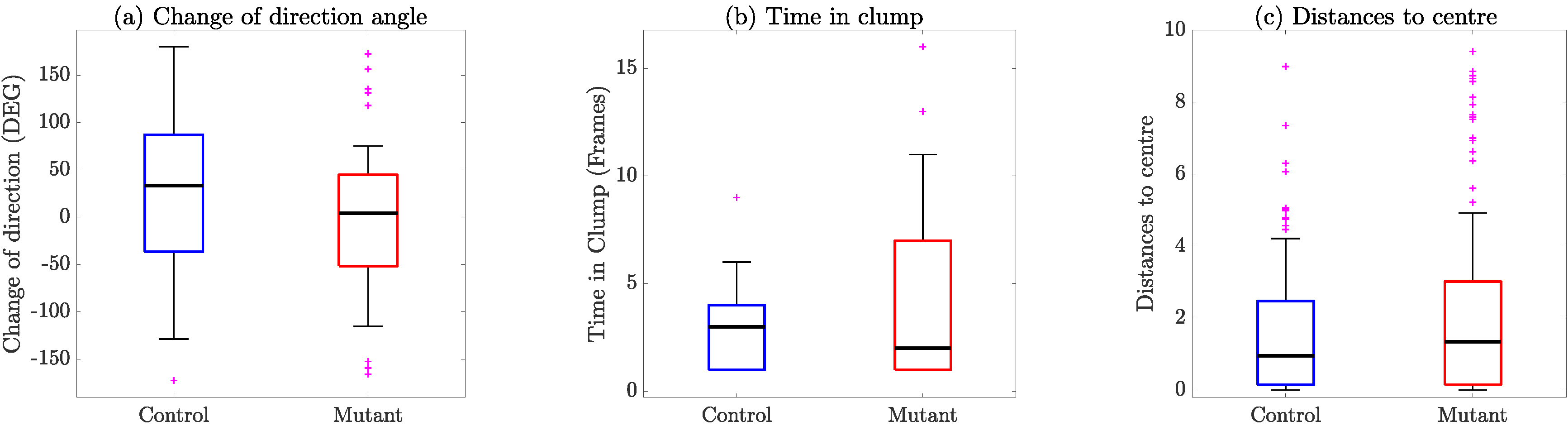
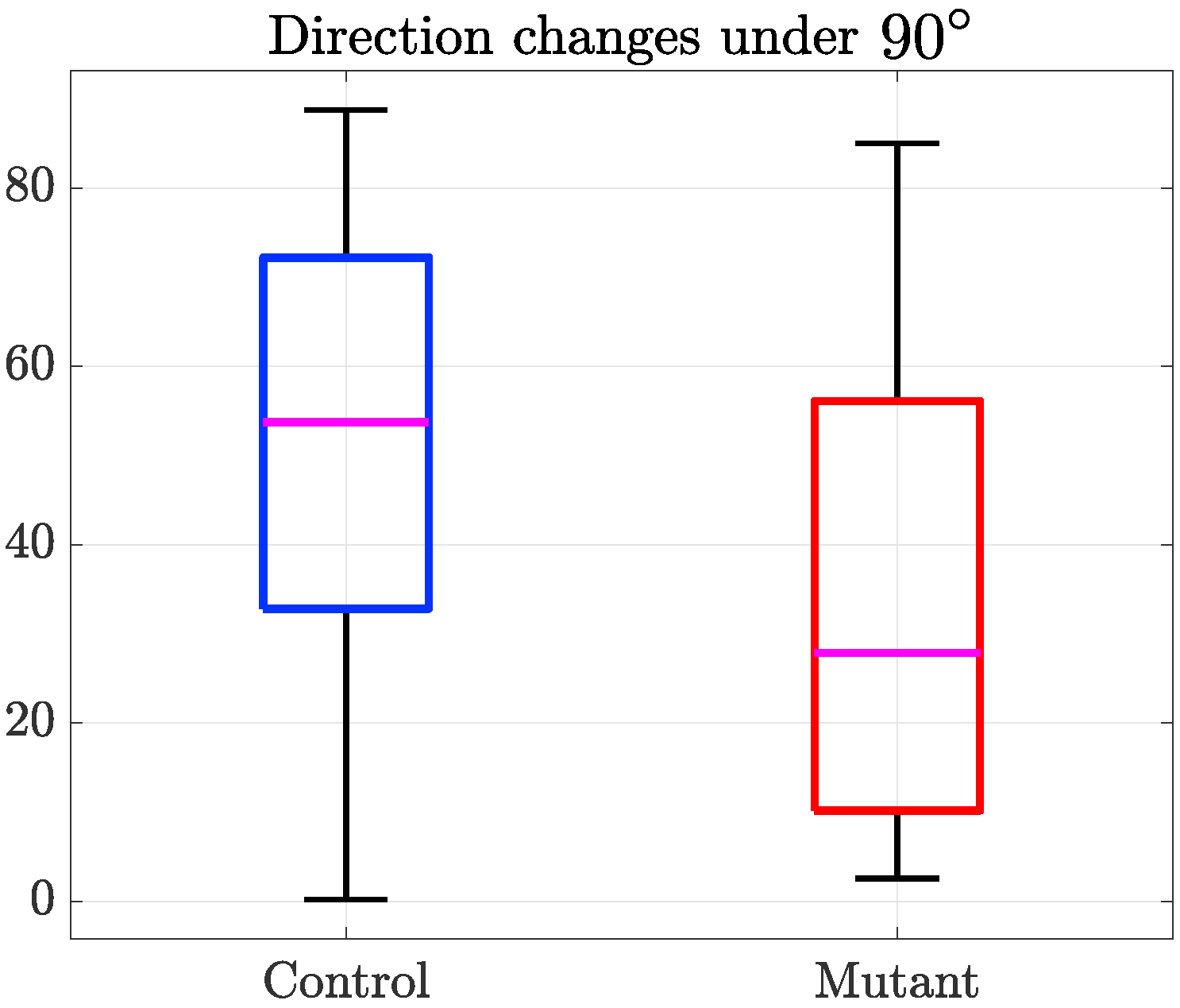
4. Results
5. Discussion
6. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| Direction | Change of direction |
| GFP | Green fluorescent probe |
| MIUA | Medical Image Understanding and Analysis |
| Track acquired with identifier i | |
| Time in clump |
References
- Lu, X.; Zhang, Y.; Liu, F.; Wang, L. Rac2 Regulates the Migration of T Lymphoid Progenitors to the Thymus during Zebrafish Embryogenesis. J. Immunol. 2020, 204, 2447–2454. [Google Scholar] [CrossRef]
- Duan, C.; Allard, J. Gonadotropin-releasing hormone neuron development in vertebrates. Gen. Comp. Endocrinol. 2020, 292, 113465. [Google Scholar] [CrossRef] [PubMed]
- Jadeski, L.C.; Hum, K.O.; Chakraborty, C.; Lala, P.K. Nitric oxide promotes murine mammary tumour growth and metastasis by stimulating tumor cell migration, invasiveness and angiogenesis. Int. J. Cancer 2000, 86, 30–39. [Google Scholar] [CrossRef]
- Gamble, J.R.; Drew, J.; Trezise, L.; Underwood, A.; Parsons, M.; Kasminkas, L.; Rudge, J.; Yancopoulos, G.; Vadas, M.A. Angiopoietin-1 is an antipermeability and anti-inflammatory agent in vitro and targets cell junctions. Circ. Res. 2000, 87, 603–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renshaw, S.A.; Loynes, C.A.; Elworthy, S.; Ingham, P.W.; Whyte, M.K.B. Modeling inflammation in the zebrafish: How a fish can help us understand lung disease. Exp. Lung Res. 2007, 33, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Aldasoro, C.C.; Biram, D.; Tozer, G.M.; Kanthou, C. Measuring cellular migration with image processing. Electron. Lett. 2008, 44, 791–793. [Google Scholar] [CrossRef] [Green Version]
- Dekoninck, S.; Blanpain, C. Stem cell dynamics, migration and plasticity during wound healing. Nat. Cell Biol. 2019, 21, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Condeelis, J.S.; Wyckoff, J.; Segall, J.E. Imaging of cancer invasion and metastasis using green fluorescent protein. Eur. J. Cancer 2000, 36, 1671–1680. [Google Scholar] [CrossRef]
- Kanthou, C.; Dachs, G.U.; Lefley, D.V.; Steele, A.J.; Coralli-Foxon, C.; Harris, S.; Greco, O.; Dos Santos, S.A.; Reyes-Aldasoro, C.C.; English, W.R.; et al. Tumour cells expressing single VEGF isoforms display distinct growth, survival and migration characteristics. PLoS ONE 2014, 9, e104015. [Google Scholar] [CrossRef]
- Reyes-Aldasoro, C.C.; Akerman, S.; Tozer, G.M. Measuring the velocity of fluorescently labelled red blood cells with a keyhole tracking algorithm. J. Microsc. 2008, 229, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Seymour, J.F.; Lieschke, G.J.; Grail, D.; Quilici, C.; Hodgson, G.; Dunn, A.R. Mice lacking both granulocyte colony-stimulating factor (CSF) and granulocyte-macrophage CSF have impaired reproductive capacity, perturbed neonatal granulopoiesis, lung disease, amyloidosis, and reduced long-term survival. Blood 1997, 90, 3037–3049. [Google Scholar] [CrossRef] [PubMed]
- Mathew, L.K.; Sengupta, S.; Kawakami, A.; Andreasen, E.A.; Löhr, C.V.; Loynes, C.A.; Renshaw, S.A.; Peterson, R.T.; Tanguay, R.L. Unraveling tissue regeneration pathways using chemical genetics. J. Biol. Chem. 2007, 282, 35202–35210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, A.; Pollard, J.W.; Wynn, T.A. Macrophage biology in development, homoeostasis and disease. Nature 2013, 496, 445. [Google Scholar]
- Rubin, G.M. Drosophila melanogaster as an experimental organism. Science 1988, 240, 1453–1459. [Google Scholar] [CrossRef] [PubMed]
- Debec, A.; Kalpin, R.F.; Daily, D.R.; McCallum, P.D.; Rothwell, W.F. Live analysis of free centrosomes in normal and aphidicolin-treated drosophila embryos. J. Cell Biol. 1996, 134, 103–115. [Google Scholar] [CrossRef] [Green Version]
- Evans, I.R.; Zanet, J.; Wood, W.; Stramer, B.M. Live imaging of Drosophila melanogaster embryonic hemocyte migrations. J. Vis. Exp. JoVE 2010, 36, e1696. [Google Scholar] [CrossRef] [Green Version]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The Genome Sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.; Martin, P. Macrophage Functions in Tissue Patterning and Disease: New Insights from the Fly. Dev. Cell 2017, 40, 221–233. [Google Scholar] [CrossRef] [Green Version]
- Stramer, B.; Moreira, S.; Millard, T.; Evans, I.; Huang, C.Y.; Sabet, O.; Milner, M.; Dunn, G.; Martin, P.; Wood, W. Clasp-mediated microtubule bundling regulates persistent motility and contact repulsion in Drosophila macrophages in vivo. J. Cell Biol. 2010, 189, 681–689. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.R.; Luchici, A.; Mosis, F.; Thackery, J.; Salazar, J.A.; Mao, Y.; Dunn, G.A.; Betz, T.; Miodownik, M.; Stramer, B.M. Inter-Cellular Forces Orchestrate Contact Inhibition of Locomotion. Cell 2015, 161, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Dogterom, M.; Koenderink, G.H. Actin-microtubule crosstalk in cell biology. Nat. Rev. Mol. Cell Biol. 2019, 20, 38–54. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Nahm, M.; Lee, M.; Kwon, M.; Kim, E.; Zadeh, A.D.; Cao, H.; Kim, H.J.; Lee, Z.H.; Oh, S.B.; et al. The F-actin-microtubule crosslinker Shot is a platform for Krasavietz-mediated translational regulation of midline axon repulsion. Dev. Camb. Engl. 2007, 134, 1767–1777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Röper, K.; Brown, N.H. A spectraplakin is enriched on the fusome and organizes microtubules during oocyte specification in Drosophila. Curr. Biol. CB 2004, 14, 99–110. [Google Scholar] [CrossRef]
- Nashchekin, D.; Fernandes, A.R.; St Johnston, D. Patronin/Shot Cortical Foci Assemble the Noncentrosomal Microtubule Array that Specifies the Drosophila Anterior-Posterior Axis. Dev. Cell 2016, 38, 61–72. [Google Scholar] [CrossRef] [Green Version]
- Zimmer, C.; Labruyere, E.; Meas-Yedid, V.; Guillen, N.; Olivo-Marin, J.C. Segmentation and tracking of migrating cells in videomicroscopy with parametric active contours: A tool for cell-based drug testing. IEEE Trans. Med. Imaging 2002, 21, 1212–1221. [Google Scholar] [CrossRef]
- Hand, A.; Sun, T.; Barber, D.; Hose, D.; MacNeil, S. Automated tracking of migrating cells in phase-contrast video microscopy sequences using image registration. J. Microsc. 2009, 234, 62–79. [Google Scholar] [CrossRef] [PubMed]
- Ulman, V.; Maška, M.; Magnusson, K.E.G.; Ronneberger, O.; Haubold, C.; Harder, N.; Matula, P.; Matula, P.; Svoboda, D.; Radojevic, M.; et al. An objective comparison of cell-tracking algorithms. Nat. Methods 2017, 14, 1141. [Google Scholar] [CrossRef]
- Maška, M.; Ulman, V.; Svoboda, D.; Matula, P.; Matula, P.; Ederra, C.; Urbiola, A.; España, T.; Venkatesan, S.; Balak, D.M.W.; et al. A benchmark for comparison of cell tracking algorithms. Bioinformatics Oxf. Engl. 2014, 30, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- Schoenauer Sebag, A.; Plancade, S.; Raulet-Tomkiewicz, C.; Barouki, R.; Vert, J.P.; Walter, T. A generic methodological framework for studying single cell motility in high-throughput time-lapse data. Bioinformatics 2015, 31, i320–i328. [Google Scholar] [CrossRef] [Green Version]
- Solis-Lemus, J.A.; Huang, Y.; Wlodkowic, D.; Reyes-Aldasoro, C.C. Microfluidic environment and tracking analysis for the observation of Artemia Franciscana. In Proceedings of the Workshop: Machine Vision of Animals and their Behaviour (MVAB 2015). British Machine Vision Association, Swansea, UK, 10 September 2015; pp. 4.1–4.8. [Google Scholar]
- Solís-Lemus, J.A.; Stramer, B.; Slabaugh, G.; Reyes-Aldasoro, C.C. Segmentation and Shape Analysis of Macrophages Using Anglegram Analysis. J. Imaging 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Solís-Lemus, J.A.; Stramer, B.; Slabaugh, G.; Reyes-Aldasoro, C.C. Shape analysis and tracking of migrating macrophages. In Proceedings of the 2018 IEEE 15th International Symposium on Biomedical Imaging (ISBI 2018), Washington, DC, USA, 4–7 April 2018; pp. 1006–1009. [Google Scholar]
- Solís-Lemus, J.A.; Stramer, B.; Slabaugh, G.; Reyes-Aldasoro, C.C. Macrosight: A Novel Framework to Analyze the Shape and Movement of Interacting Macrophages Using Matlab. J. Imaging 2019, 5, 17. [Google Scholar] [CrossRef] [Green Version]
- Solí s-Lemus, J.A.; Sánchez-Sánchez, B.J.; Marcotti, S.; Burki, M.; Stramer, B.; Reyes-Aldasoro, C.C. Comparison of Interactions Between Control and Mutant Macrophages. In Medical Image Understanding and Analysis; Zheng, Y., Williams, B.M., Chen, K., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 470–477. [Google Scholar]
- Stramer, B.; Reyes-Aldasoro, C.C.; Solis-Lemus, J.A.; Sanchez-Sanchez, B.; Marcotti, S.; Burki, M. Fluorescent Macrophages in Drosophila Embryo. BioRxiv 2020. [Google Scholar] [CrossRef]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development Camb. Engl. 1993, 118, 401–415. [Google Scholar]
- Bruckner, K.; Kockel, L.; Duchek, P.; Luque, C.M.; Rorth, P.; Perrimon, N. The PDGF/VEGF receptor controls blood cell survival in Drosophila. Dev. Cell 2004, 7, 73–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barolo, S.; Castro, B.; Posakony, J.W. New Drosophila transgenic reporters: Insulated P-element vectors expressing fast-maturing RFP. Biotechniques 2004, 36, 436–440, 442. [Google Scholar] [CrossRef] [Green Version]
- Stramer, B.; Wood, W. Inflammation and Wound Healing in Drosophila. In Chemotaxis; Methods in Molecular Biology™; Humana Press: Totowa, NJ, USA, 2009; pp. 137–149. [Google Scholar]
- Henry, K.M.; Pase, L.; Ramos-Lopez, C.F.; Lieschke, G.J.; Renshaw, S.A.; Reyes-Aldasoro, C.C. PhagoSight: An Open-Source MATLAB Package for the Analysis of Fluorescent Neutrophil and Macrophage Migration in a Zebrafish Model. PLoS ONE 2013, 8, e72636. [Google Scholar] [CrossRef]
- Otsu, N. A Threshold Selection Method from Gray-Level Histograms. IEEE Trans. Syst. Man Cyber. 1979, 9, 62–66. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset ID | n Interactions | Dataset ID | n Interactions | Dataset ID | n Interactions |
|---|---|---|---|---|---|
| CONTROL01 | 14 | MUTANT03 | 10 | MUTANT07 | 3 |
| CONTROL02 | 4 | MUTANT04 | 2 | MUTANT08 | 2 |
| CONTROL04 | 4 | MUTANT05 | 2 | MUTANT10 | 4 |
| CONTROL05 | 2 | MUTANT06 | 9 | MUTANT11 | 7 |
| TOTAL | 24 | TOTAL | 39 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solís-Lemus, J.A.; Sánchez-Sánchez, B.J.; Marcotti, S.; Burki, M.; Stramer, B.; Reyes-Aldasoro, C.C. Comparative Study of Contact Repulsion in Control and Mutant Macrophages Using a Novel Interaction Detection. J. Imaging 2020, 6, 36. https://doi.org/10.3390/jimaging6050036
Solís-Lemus JA, Sánchez-Sánchez BJ, Marcotti S, Burki M, Stramer B, Reyes-Aldasoro CC. Comparative Study of Contact Repulsion in Control and Mutant Macrophages Using a Novel Interaction Detection. Journal of Imaging. 2020; 6(5):36. https://doi.org/10.3390/jimaging6050036
Chicago/Turabian StyleSolís-Lemus, José Alonso, Besaiz J Sánchez-Sánchez, Stefania Marcotti, Mubarik Burki, Brian Stramer, and Constantino Carlos Reyes-Aldasoro. 2020. "Comparative Study of Contact Repulsion in Control and Mutant Macrophages Using a Novel Interaction Detection" Journal of Imaging 6, no. 5: 36. https://doi.org/10.3390/jimaging6050036
APA StyleSolís-Lemus, J. A., Sánchez-Sánchez, B. J., Marcotti, S., Burki, M., Stramer, B., & Reyes-Aldasoro, C. C. (2020). Comparative Study of Contact Repulsion in Control and Mutant Macrophages Using a Novel Interaction Detection. Journal of Imaging, 6(5), 36. https://doi.org/10.3390/jimaging6050036