
Detection and Characterization of the Soybean Allergen Gly m 7 in Soybeans and Processed Soybean Foods



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Anti-Gly m 7 Antibodies
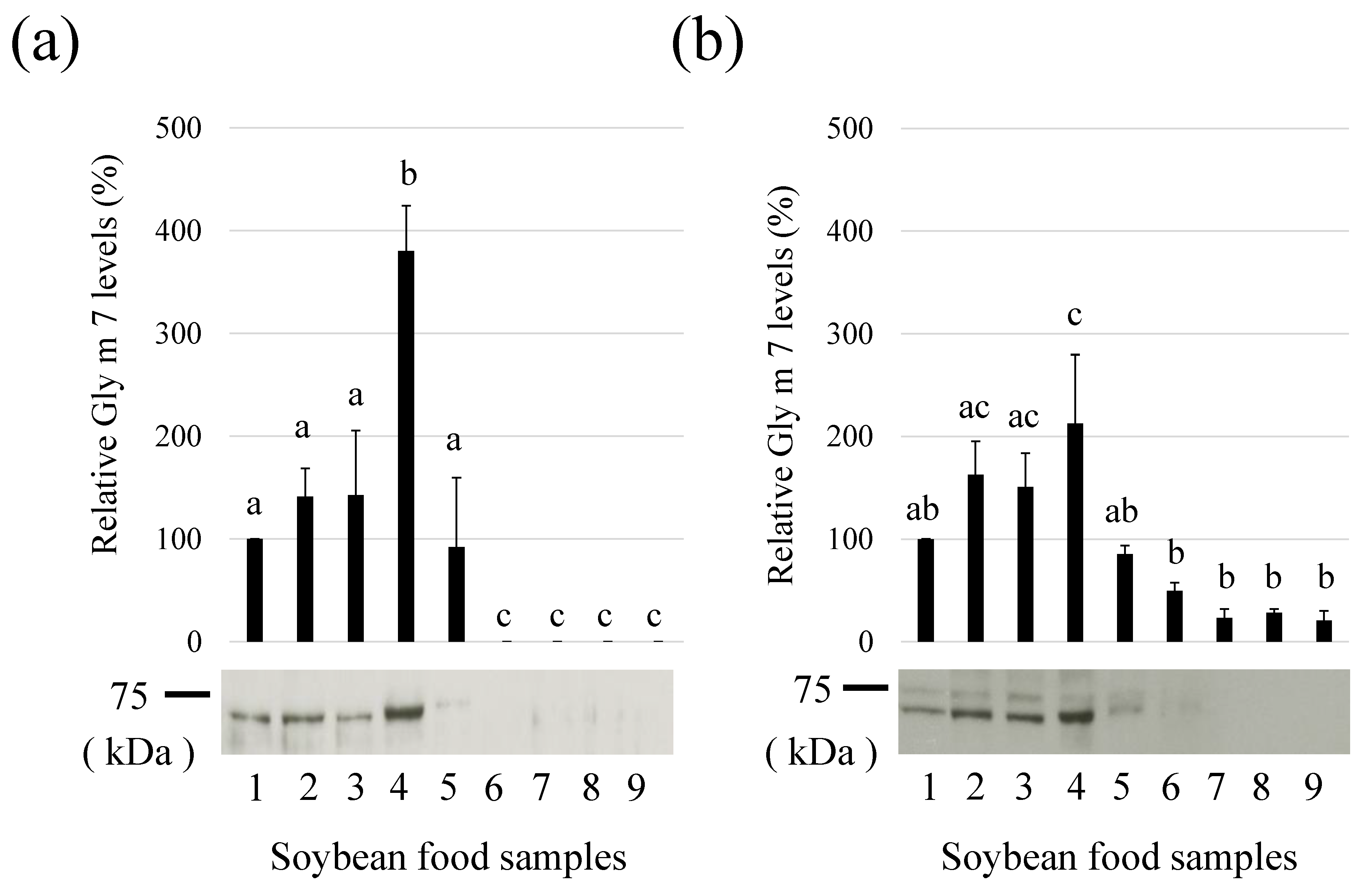
2.3. Preparation of Food Samples
2.4. Preparation of Extracts from Dried Soybeans and Protein Analysis
2.5. Antibodies to Soybean Allergens Other Than Gly m 7
2.6. Electrophoresis and Western Blotting
2.7. Detection of Biotin Moiety of Gly m 7
2.8. ELISA for Determination of Antibody Titers
2.9. Statistical Analysis
3. Results
3.1. Preparation and Assessment of Anti-Gly m 7 Antibodies
3.2. Detection of the Biotin Moiety of Gly m 7
3.3. Detection of Gly m 7 in Various Processed Soybean Foods
3.4. Detection of Gly m 7 in Various Soymilks
3.5. Detection of Gly m 7 in Various Soybean Cultivars
3.6. Thermostability of Gly m 7
3.7. Effect of Worm Wounding on Soybean Gly m 7 Levels
3.8. Effect of Immersion Times of Soybeans on Gly m 7 and Other Allergens
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sirtori, C.R.; Agradi, E.; Conti, F.; Mantero, O.; Gatti, E. Soybean-protein diet in the treatment of type-II hyperlipoproteinaemia. Lancet 1977, 1, 275–277. [Google Scholar] [CrossRef]
- Sugano, M.; Koba, K. Dietary protein and lipid metabolism: A multifunctional effect. Ann. N. Y. Acad. Sci. 1993, 676, 215–222. [Google Scholar] [CrossRef]
- Torres, N.; Torre-Villalvazo, I.; Tovar, A.R. Regulation of lipid metabolism by soy protein and its implication in diseases mediated by lipid disorders. J. Nutr. Biochem. 2006, 17, 365–373. [Google Scholar] [CrossRef]
- Malav, O.P.; Talukder, S.; Gokulakrishnan, P.; Chand, S. Meat Analog: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1241–1245. [Google Scholar] [CrossRef]
- Singh, M.; Trivedi, N.; Enamala, M.K.; Kuppam, C.; Parikh, P.; Nikolova, M.P.; Chavali, M. Plant-based meat analogue (PBMA) as a sustainable food: A concise review. Eur. Food Res. Technol. 2021, 247, 2499–2526. [Google Scholar] [CrossRef]
- Ogawa, T.; Samoto, M.; Takahashi, K. Soybean allergens and hypoallergenic soybean products. J. Nutr. Sci. Vitaminol. 2000, 46, 271–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moroz, L.A.; Yang, W.H. Kunitz Soybean Trypsin Inhibitor—A Specific Allergen in Food Anaphylaxis. N. Engl. J. Med. 1980, 302, 1126–1128. [Google Scholar] [CrossRef]
- Ogawa, T.; Tsuji, H.; Bando, N.; Kitamura, K.; Zhu, Y.L.; Hirano, H.; Nishikawa, K. Identification of the soybean allergenic protein, Gly m Bd 30 K, with the soybean seed 34-kDa oil-body-associated protein. Biosci. Biotechnol. Biochem. 1993, 57, 1030–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogawa, T.; Bando, N.; Tsuji, H.; Okajima, H.; Nishikawa, K.; Sasaoka, K. Investigation of the IgE-binding protein in soybeans by immunoblotting with the sera of the soybean-sensitive patient with atopic dermatitis. J. Nutr. Sci. Vitaminol. 1991, 37, 555–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, H.; Bando, N.; Hiemori, M.; Yamanishi, R.; Kimoto, M.; Nishikawa, K.; Ogawa, T. Purification and Characterization of Soybean Allergen Gly m Bd 28K. Biosci. Biotechnol. Biochem. 1997, 61, 942–947. [Google Scholar] [CrossRef]
- Ogawa, T.; Bando, N.; Tsuji, H.; Nishikawa, K.; Kitamura, K. α-Subunit of β-Conglycinin, an allergenic protein recognized by IgE Antibodies of soybean-sensitive patients with atopic dermatitis. Biosci. Biotechnol. Biochem. 1995, 59, 831–833. [Google Scholar] [CrossRef] [Green Version]
- Adachi, A.; Horikawa, T.; Shimizu, H.; Sarayama, Y.; Ogawa, T.; Sjolander, S.; Tanaka, A.; Moriyama, T. Soybean beta-conglycinin as the main allergen in a patient with food-dependent exercise-induced anaphylaxis by tofu: Food processing alters pepsin resistance. J. Clin. Exp. Allergy 2009, 39, 167–173. [Google Scholar] [CrossRef]
- Holzhauser, T.; Wackermann, O.; Ballmer-Weber, B.K.; Bindslev-Jensen, C.; Scibilia, J.; Perono-Garoffo, L.; Utsumi, S.; Poulsen, L.K.; Vieths, S. Soybean (Glycine max) allergy in Europe: Gly m 5 (β-conglycinin) and Gly m 6 (glycinin) are potential diagnostic markers for severe allergic reactions to soy. J. Allergy Clin. Immunol. 2009, 123, 452–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maruyama, N.; Sato, S.; Cabanos, C.; Tanaka, A.; Ito, K.; Ebisawa, M. Gly m 5/Gly m 8 fusion component as a potential novel candidate molecule for diagnosing soya bean allergy in Japanese children. Clin. Exp. Allergy 2018, 48, 1726–1734. [Google Scholar] [CrossRef]
- Kleine-Tabbe, J.; Vogel, L.; Crowell, D.N.; Haustein, U.F.; Vieths, S. Severe oral allergy syndrome and anaphylactic reactions caused by a Bet v 1- related PR-10 protein in soybean, SAM22. J. Allergy Clin. Immunol. 2002, 110, 797–804. [Google Scholar] [CrossRef]
- Mittag, D.; Vieths, S.; Vogel, L.; Becker, W.M.; Rihs, H.P.; Helbling, A.; Wüthrich, B.; Ballmer-Weber, B.K. Soybean allergy in patients allergic to birch pollen: Clinical investigation and molecular characterization of allergens. J. Allergy Clin. Immunol. 2004, 113, 148–154. [Google Scholar] [CrossRef]
- Rihs, H.P.; Chen, Z.; Ruëff, F.; Petersen, A.; Rozynek, P.; Heimann, H.; Baur, X. IgE binding of the recombinant allergen soybean profilin (rGly m 3) is mediated by conformational epitopes. J. Allergy Clin. Immunol. 1999, 104, 1293–1301. [Google Scholar] [CrossRef]
- Riascos, J.J.; Weissinger, S.M.; Weissinger, A.K.; Kulis, M.; Burks, A.W.; Pons, L. The Seed Biotinylated Protein of Soybean (Glycine max): A Boiling Resistant New Allergen (Gly m 7) with the Capacity to Induce IgE Mediated Allergic Responses. J. Agric. Food Chem. 2016, 64, 3890–3900. [Google Scholar] [CrossRef]
- Hsing, Y.; Tsou, C.; Hsu, T.; Chen, Z.; Hsieh, K.; Hsieh, J.; Chow, T. Tissue- and stage-specific expression of a soybean (Glycine max L.) seed-maturation, biotinylated protein. Plant Mol. Biol. 1998, 38, 481–490. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Moriyama, T.; Machidori, M.; Ozasa, S.; Maebuchi, M.; Urade, R.; Takahashi, K.; Ogawa, T.; Maruyama, N. A novel enzyme-linked immunosorbent assay for quantification of soybean beta-conglycinin, a major soybean storage protein, in soybean and soybean food products. J. Nutr. Sci. Vitaminol. 2005, 51, 34–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanafusa, K.; Murakami, H.; Ueda, T.; Yano, E.; Zaima, N.; Moriyama, T. Worm wounding increases levels of pollen-related food allergens in soybean (Glycine max). Biosci. Biotechnol. Biochem. 2018, 82, 1207–1215. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Kyhse-Andersen, J. Electroblotting of multiple gels: A simple apparatus without buffer tank for rapid transfer of proteins from polycrylamide to nitrocellulose. J. Biochem. Biophys. Methods 1984, 10, 203–209. [Google Scholar] [CrossRef]
- Moriyama, T.; Yano, E.; Suemori, Y.; Nakano, K.; Zaima, N.; Kawamura, Y. Hypoallergenicity of Various Miso Pastes Manu-factured in Japan. J. Nutr. Sci. Vitaminol. 2013, 59, 462–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuo, A.; Matsushita, K.; Fukuzumi, A.; Tokumasu, N.; Yano, E.; Zaima, N.; Moriyama, T. Comparison of Various Soybean Allergen Levels in Genetically and Non-Genetically Modified Soybeans. Foods 2020, 9, 522. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Manufactures | Protein Concentrations (mg/mL) |
|---|---|---|
| 1 | company A | 38.5 |
| 2 | company A | 45.5 |
| 3 | company A | 12.0 |
| 4 | company A | 27.0 |
| 5 | company B | 36.0 |
| 6 | company B | 56.8 |
| 7 | company B | 27.0 |
| 8 | company C | 50.0 |
| 9 | company C | 47.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukuzumi, A.; Tokumasu, N.; Matsuo, A.; Yano, E.; Zaima, N.; Moriyama, T. Detection and Characterization of the Soybean Allergen Gly m 7 in Soybeans and Processed Soybean Foods. Allergies 2021, 1, 233-246. https://doi.org/10.3390/allergies1040022
Fukuzumi A, Tokumasu N, Matsuo A, Yano E, Zaima N, Moriyama T. Detection and Characterization of the Soybean Allergen Gly m 7 in Soybeans and Processed Soybean Foods. Allergies. 2021; 1(4):233-246. https://doi.org/10.3390/allergies1040022
Chicago/Turabian StyleFukuzumi, Ayano, Naoki Tokumasu, Ayato Matsuo, Erika Yano, Nobuhiro Zaima, and Tatsuya Moriyama. 2021. "Detection and Characterization of the Soybean Allergen Gly m 7 in Soybeans and Processed Soybean Foods" Allergies 1, no. 4: 233-246. https://doi.org/10.3390/allergies1040022
APA StyleFukuzumi, A., Tokumasu, N., Matsuo, A., Yano, E., Zaima, N., & Moriyama, T. (2021). Detection and Characterization of the Soybean Allergen Gly m 7 in Soybeans and Processed Soybean Foods. Allergies, 1(4), 233-246. https://doi.org/10.3390/allergies1040022