
Regional Population Structure of the European Eel at the Southern Limit of Its Distribution Revealed by Otolith Shape Signature


Abstract
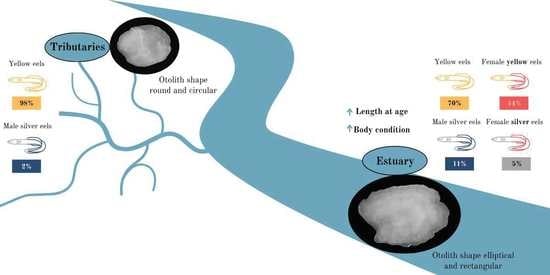
:
1. Introduction
2. Materials and Methods
2.1. Population Characterization
2.1.1. Study Area
2.1.2. Sampling
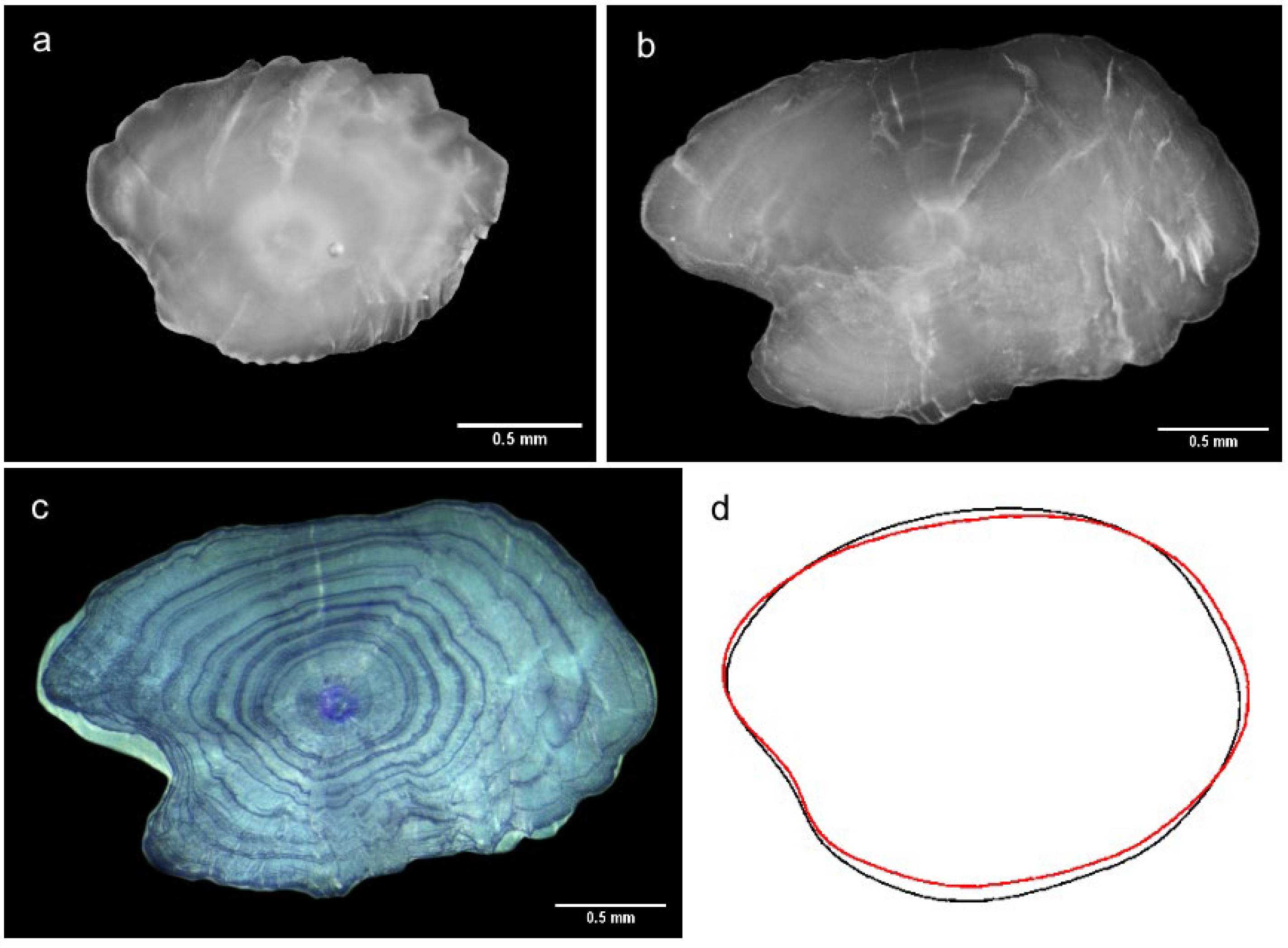
2.2. Otolith Analysis
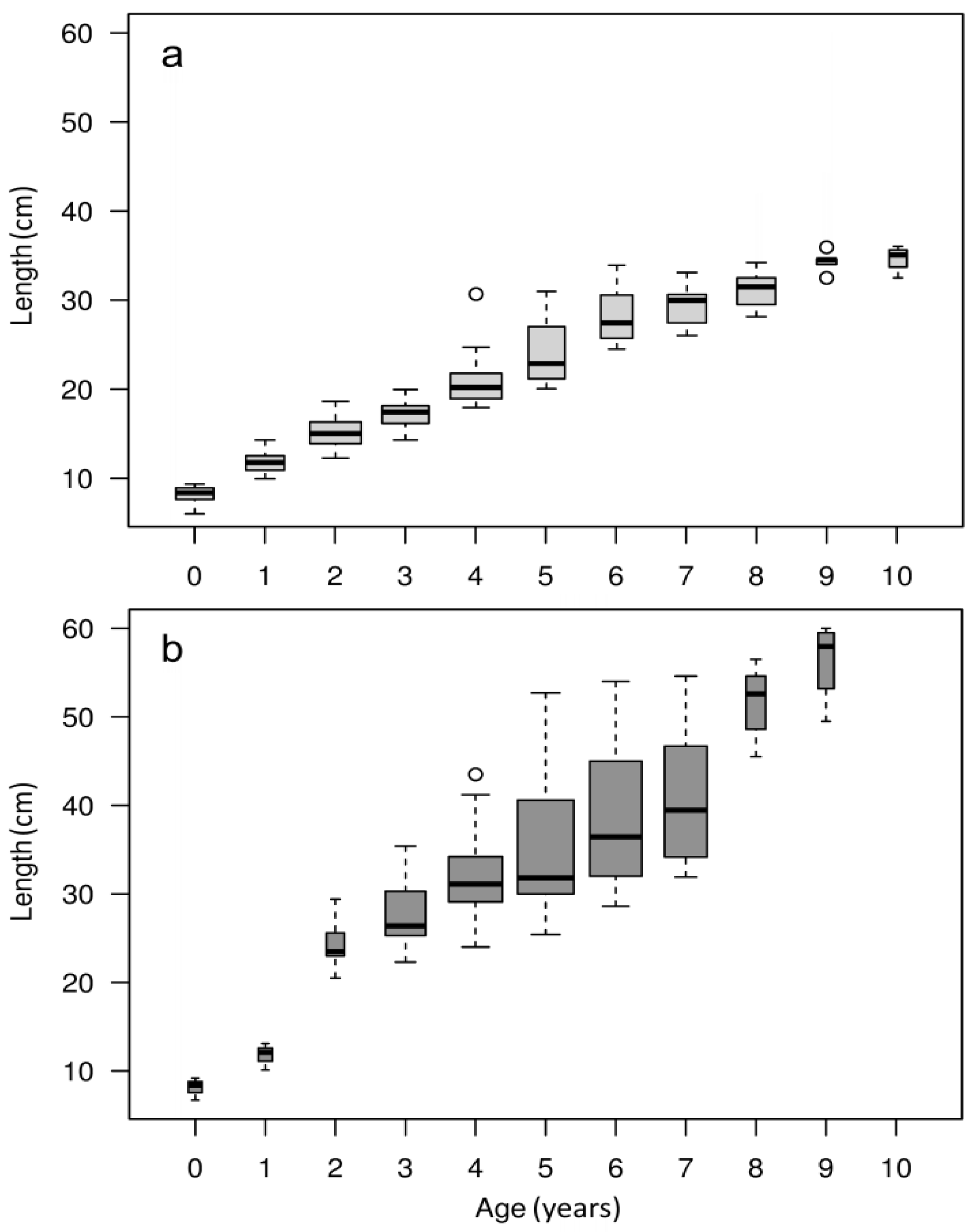
2.2.1. Age Estimation
2.2.2. Shape Descriptors
2.2.3. Statistical Analysis
3. Results
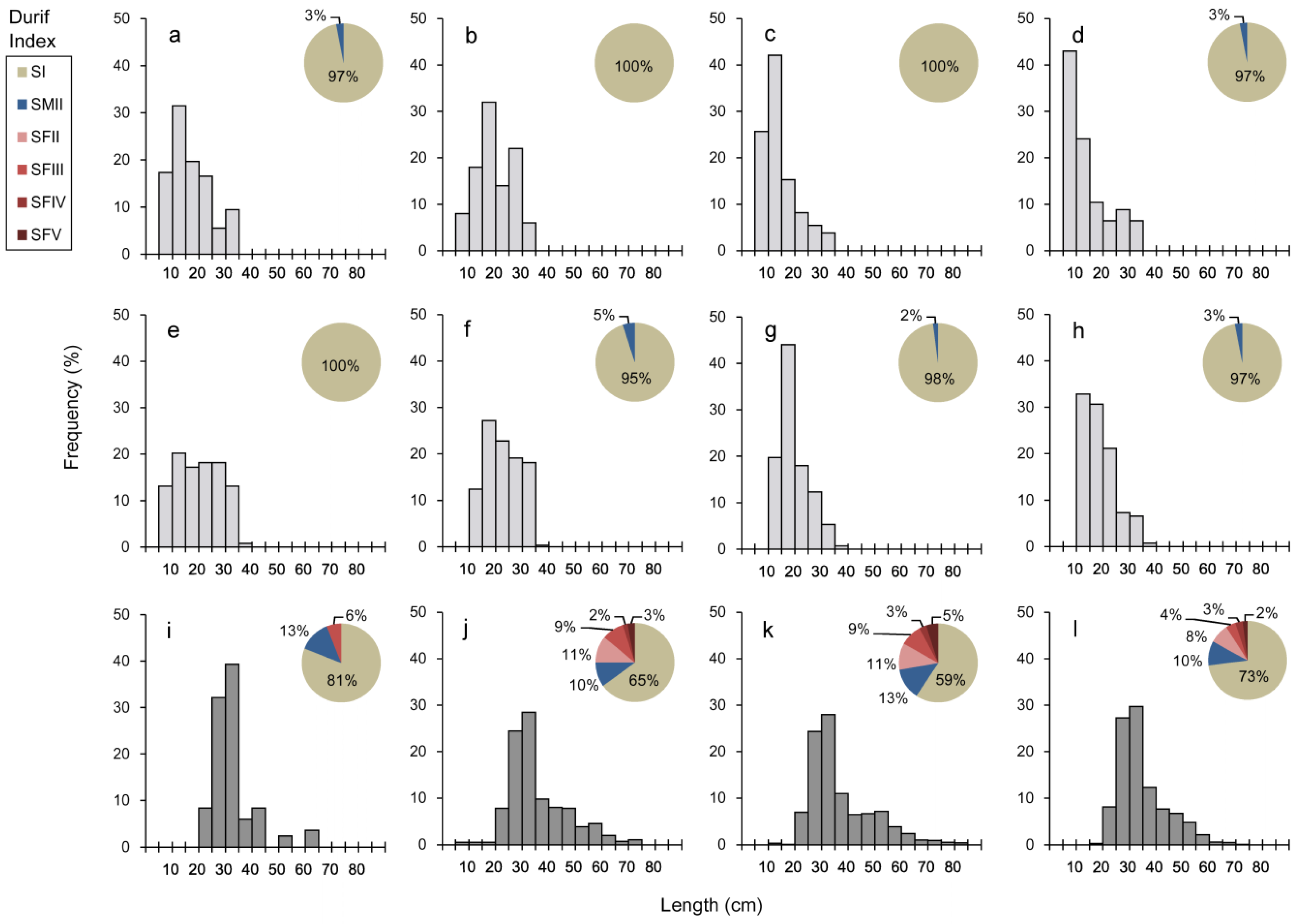
3.1. Population Characterization
3.2. Otolith Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- ICES. Report of the Joint EIFAAC/ICES/GFCM Working Group on Eel (WGEEL); ICES Document CM 2015/ACOM: 18; ICES: Antalya, Turkey, 2015. [Google Scholar]
- Moriarty, C.; Dekker, W. Management of European eel fisheries. Irish Fish. Bull. 1997, 15, 108. [Google Scholar]
- Dekker, W. Status of the European eel stock and fisheries. In Eel Biology; Springer: Tokyo, Japan, 2003. [Google Scholar]
- Bornarel, V.; Lambert, P.; Briand, C.; Antunes, C.; Belpaire, C.; Ciccotti, E.; Diaz, E.; Diserud, O.; Doherty, D.; Domingos, I.; et al. Modelling the recruitment of European eel (Anguilla anguilla) throughout its European range. ICES J. Mar. Sci. 2018, 75, 541–552. [Google Scholar] [CrossRef]
- Dekker, W. The history of commercial fisheries for European eel commenced only a century ago. Fish. Manag. Ecol. 2019, 26, 6–19. [Google Scholar] [CrossRef]
- Jacoby, D.; Gollock, M. The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2014. [Google Scholar]
- Jacoby, D.M.; Casselman, J.M.; Crook, V.; DeLucia, M.B.; Ahn, H.; Kaifu, K.; Kurwie, T.; Sasal, P.; Silfvergrip, A.M.; Smith, K.G.; et al. Synergistic patterns of threat and the challenges facing global anguillid eel conservation. Glob. Ecol. Conserv. 2015, 4, 321–333. [Google Scholar] [CrossRef]
- Miller, M.J.; Feunteun, E.; Tsukamoto, K. Did a “perfect storm” of oceanic changes and continental anthropogenic impacts cause northern hemisphere anguillid recruitment reductions? ICES J. Mar. Sci. 2016, 73, 43–56. [Google Scholar] [CrossRef] [Green Version]
- Dekker, W.; Beaulaton, L. Climbing back up what slippery slope? Dynamics of the European eel stock and its management in historical perspective. ICES J. Mar. Sci. 2016, 73, 5–13. [Google Scholar] [CrossRef] [Green Version]
- ICES. Joint EIFAAC/ICES/GFCM Working Group on Eels (WGEEL); ICES: Antalya, Turkey, 2019. [Google Scholar]
- The European Commission. Commission Decision 2010/93/EU of 18 December 2009 adopting a multiannual Community program for the collection, management and use of data in the fisheries sector for the period 2011–2013 (notified under document C (2009) 10121). Off. J. Eur. Union L 2009, 41, 8–71. [Google Scholar]
- ICES. Report of the Workshop on Designing an Eel Data Call (WKEELDATA); ICES CM 2017/SGIEOM: 30; ICES: Rennes, France, 2017. [Google Scholar]
- Tsukamoto, K.; Nakai, I. Do all freshwater eels migrate? Nature 1998, 396, 635–636. [Google Scholar] [CrossRef]
- Limburg, K.E.; Wickstrom, H.; Svedang, H.; Elfman, M.; Kristiansson, P. Do stocked freshwater eels migrate? Evidence from the Baltic suggests “yes”. In American Fisheries Society Symposium; American Fisheries Society: New York, NY, USA, 2003; pp. 275–284. [Google Scholar]
- Daverat, F.; Limburg, K.E.; Thibault, I.; Shiao, J.C.; Dodson, J.J.; Caron, F.; Tzeng, W.N.; Iizuka, Y.; Wickström, H. Phenotypic plasticity of habitat use by three temperate eel species, Anguilla anguilla, A japonica and A. rostrata. Mar. Ecol. Prog. Ser. 2006, 308, 231–241. [Google Scholar] [CrossRef]
- Als, T.D.; Hansen, M.M.; Maes, G.E.; Castonguay, M.; Riemann, L.; Aarestrup, K.I.M.; Munk, P.; Sparholt, H.; Hanel, R.; Bernatchez, L. All roads lead to home: Panmixia of European eel in the Sargasso Sea. Mol. Ecol. 2011, 20, 1333–1346. [Google Scholar] [CrossRef]
- Schmidt, J. On the distribution of the freshwater eels (Anguilla) throughout the world. I. Atlantic Ocean and adjacent regions. Medd. Fra Komm. Havunders. Seri Fisk. 1909, 3, 1–45. [Google Scholar]
- Tesch, F.W. Biology and Management of Anguillid Eels; CRC Press: Boco Raton, FL, USA, 1977. [Google Scholar]
- Dekker, W. On the distribution of the European eel (Anguilla anguilla) and its fisheries. Can. J. Fish. Aquat. Sci. 2003, 60, 787–799. [Google Scholar] [CrossRef]
- Antunes, C.; Tesch, F.W. A critical consideration of the metamorphosis zone when identifying daily rings in otoliths of European eel, Anguilla anguilla (L.). Ecol. Freshw. Fish 1997, 6, 102–107. [Google Scholar] [CrossRef]
- Edeline, E.; Dufour, S.; Elie, P. Role of glass eel salinity preference in the control of habitat selection and growth plasticity in Anguilla anguilla. Mar. Ecol. Prog. Ser. 2005, 304, 191–199. [Google Scholar] [CrossRef]
- Edeline, E.; Lambert, P.; Rigaud, C.; Elie, P. Effects of body condition and water temperature on Anguilla anguilla glass eel migratory behavior. J. Exp. Mar. Biol. Ecol. 2006, 331, 217–225. [Google Scholar] [CrossRef]
- Vøllestad, L.A. Geographic variation in age and length at metamorphosis of maturing European eel: Environmental effects and phenotypic plasticity. J. Anim. Ecol. 1992, 61, 41–48. [Google Scholar] [CrossRef]
- Bertin, L. Eels: A Biological Study; Cleaver-Hume Press: London, UK, 1956. [Google Scholar]
- Van Den Thillart, G.E.E.J.; Van Ginneken, V.; Körner, F.; Heijmans, R.; Van der Linden, R.; Gluvers, A. Endurance swimming of European eel. J. Fish Biol. 2004, 65, 312–318. [Google Scholar] [CrossRef]
- Helfman, G.S.; Facey, D.E.; Hales, L.S., Jr.; Bozeman, E.L., Jr. Reproductive ecology of the American eel. In American Fisheries Society Symposium; American Fisheries Society: New York, NY, USA, 1987; Volume 1, pp. 42–56. [Google Scholar]
- Daverat, F.; Beaulaton, L.; Poole, R.; Lambert, P.; Wickström, H.; Andersson, J.; Aprahamian, M.; Hizem, B.; Elie, P.; Yalçın-Özdilek, S.; et al. One century of eel growth: Changes and implications. Ecol. Freshw. Fish 2012, 21, 325–336. [Google Scholar] [CrossRef]
- Tesch, F.W. The Eel, 5th ed.; Blackwell Publishing: Oxford, UK, 2003. [Google Scholar]
- Jessop, B.M.; Shiao, J.C.; Iizuka, Y.; Tzeng, W.N. Variation in the annual growth, by sex and migration history, of silver American eels Anguilla rostrata. Mar. Ecol. Prog. Ser. 2004, 272, 231–244. [Google Scholar] [CrossRef] [Green Version]
- Krueger, W.H.; Oliveira, K. Evidence for environmental sex determination in the American eel, Anguilla rostrata. Environ. Biol. Fishes 1999, 55, 381–389. [Google Scholar] [CrossRef]
- Bevacqua, D.; Melia, P.; De Leo, G.A.; Gatto, M. Intra-specific scaling of natural mortality in fish: The paradigmatic case of the European eel. Oecologia 2011, 165, 333–339. [Google Scholar] [CrossRef]
- Boulenger, C.; Acou, A.; Gimenez, O.; Charrier, F.; Tremblay, J.; Feunteun, E. Factors determining survival of European eels in two unexploited sub-populations. Freshw. Biol. 2016, 61, 947–962. [Google Scholar] [CrossRef]
- Leo, G.D.; Gatto, M. A size and age-structured model of the European eel (Anguilla anguilla L.). Can. J. Fish. Aquat. Sci. 1995, 52, 1351–1367. [Google Scholar] [CrossRef]
- Poole, W.R.; Reynolds, J.D. Variability in growth rate in European eel Anguilla anguilla (L.) in a western Irish catchment. In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 1998; pp. 141–145. [Google Scholar]
- Parsons, J.; Vickers, K.U.; Warden, Y. Relationship between elver recruitment and changes in the sex ratio of silver eels Anguilla anguilla L. migrating from Lough Neagh, Northern Ireland. J. Fish Biol. 1977, 10, 211–229. [Google Scholar] [CrossRef]
- Walsh, C.T.; Pease, B.C.; Booth, D.J. Variation in the sex ratio, size and age of long finned eels within and among coastal catchments of south-eastern Australia. J. Fish Biol. 2004, 64, 1297–1312. [Google Scholar] [CrossRef]
- Kettle, A.J.; Asbjørn Vøllestad, L.; Wibig, J. Where once the eel and the elephant were together: Decline of the European eel because of changing hydrology in southwest Europe and northwest Africa? Fish Fish. 2011, 12, 380–411. [Google Scholar] [CrossRef]
- Vøllestad, L.A.; Jonsson, B. A 13-year study of the population dynamics and growth of the European eel Anguilla anguilla in a Norwegian river: Evidence for density-dependent mortality, and development of a model for predicting yield. J. Anim. Ecol. 1988, 57, 983–997. [Google Scholar] [CrossRef]
- Sakai, M.; Iwabuchi, K. Unique habitat and macroinvertebrate assemblage structures in spring-fed streams: A comparison among lowland tributaries and mainstreams in northern Japan. bioRxiv 2020, 22, 193–202. [Google Scholar] [CrossRef]
- Post, D.M.; Pace, M.L.; Hairston, N.G. Ecosystem size determines food-chain length in lakes. Nature 2000, 405, 1047–1049. [Google Scholar] [CrossRef] [PubMed]
- Maceda-Veiga, A.; Mac Nally, R.; de Sostoa, A. Environmental correlates of food-chain length, mean trophic level and trophic level variance in invaded riverine fish assemblages. Sci. Total Environ. 2018, 644, 420–429. [Google Scholar] [CrossRef]
- Pimm, S.L.; Lawton, J.H. Number of trophic levels in ecological communities. Nature 1977, 268, 329–331. [Google Scholar] [CrossRef]
- Panfili, J.; Ximénès, M.C.; Crivelli, A.J. Sources of variation in growth of the European eel (Anguilla anguilla) estimated from otoliths. Can. J. Fish. Aquat. Sci. 1994, 51, 506–515. [Google Scholar] [CrossRef] [Green Version]
- Campana, S.E.; Thorrold, S.R. Otoliths, increments, and elements: Keys to a comprehensive understanding of fish populations? Can. J. Fish. Aquat. Sci. 2001, 58, 30–38. [Google Scholar] [CrossRef]
- Lin, Y.J.; Ložys, L.; Shiao, J.C.; Iizuka, Y.; Tzeng, W.N. Growth differences between naturally recruited and stocked European eel Anguilla anguilla from different habitats in Lithuania. J. Fish Biol. 2007, 71, 1773–1787. [Google Scholar] [CrossRef]
- Thorrold, S.R.; Campana, S.E.; Jones, C.M.; Swart, P.K. Factors determining δ13C and δ18O fractionation in aragonitic otoliths of marine fish. Geochim. Cosmochim. Acta 1997, 61, 2909–2919. [Google Scholar] [CrossRef]
- Elsdon, T.S.; Gillanders, B.M. Reconstructing migratory patterns of fish based on environmental influences on otolith chemistry. Rev. Fish Biol. Fish. 2003, 13, 217–235. [Google Scholar] [CrossRef]
- Begg, G.A.; Campana, S.E.; Fowler, A.J.; Suthers, I.M. Otolith research and application: Current directions in innovation and implementation. Mar. Freshw. Res. 2005, 56, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Campana, S.E. Otolith science entering the 21st century. Mar. Freshw. Res. 2005, 56, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Moreira, C.; Froufe, E.; Vaz-Pires, P.; Correia, A.T. Otolith shape analysis as a tool to infer the population structure of the blue jack mackerel, Trachurus picturatus, in the NE Atlantic. Fish. Res. 2019, 209, 40–48. [Google Scholar] [CrossRef]
- Moura, A.; Muniz, A.A.; Mullis, E.; Wilson, J.M.; Vieira, R.P.; Almeida, A.A.; Pinto, E.; Brummer, G.J.A.; Gaever, P.V.; Gonçalves, J.M.S.; et al. Population structure and dynamics of the Atlantic mackerel (Scomber scombrus) in the North Atlantic inferred from otolith chemical and shape signatures. Fish. Res. 2020, 230, 105621. [Google Scholar] [CrossRef]
- Muniz, A.A.; Moura, A.; Triay-Portella, R.; Moreira, C.; Santos, P.T.; Correia, A.T. Population structure of the chub mackerel (Scomber colias) in the North-east Atlantic inferred from otolith shape and body morphometrics. Mar. Freshw. Res. 2020, 72, 341–352. [Google Scholar] [CrossRef]
- Jones, C.M. Development and application of the otolith increment technique. In Otolith Microstructure Examination and Analysis; Canadian Special Publication of Fisheries and Aquatic Sciences 117; Publishing Supply and Services Canada: Ottawa, ON, Canada, 1992; Volume 117, pp. 1–11. [Google Scholar]
- Lombarte, A.; Torres, G.J.; Morales-Nin, B. Specific Merluccius otolith growth patterns related to phylogenetics and environmental factors. Marine Biological Association of the United Kingdom. J. Mar. Biol. Assoc. 2003, 83, 277. [Google Scholar] [CrossRef] [Green Version]
- Bacha, M.; Jemaa, S.; Hamitouche, A.; Rabhi, K.; Amara, R. Population structure of the European anchovy, Engraulis encrasicolus, in the SW Mediterranean Sea, and the Atlantic Ocean: Evidence from otolith shape analysis. ICES J. Mar. Sci. 2014, 71, 2429–2435. [Google Scholar] [CrossRef]
- Vieira, A.R.; Neves, A.; Sequeira, V.; Paiva, R.B.; Gordo, L.S. Otolith shape analysis as a tool for stock discrimination of forkbeard (Phycis phycis) in the Northeast Atlantic. Hydrobiologia 2014, 728, 103–110. [Google Scholar] [CrossRef]
- Jemaa, S.; Bacha, M.; Khalaf, G.; Dessailly, D.; Rabhi, K.; Amara, R. What can otolith shape analysis tell us about population structure of the European sardine, Sardina pilchardus, from Atlantic and Mediterranean waters? J. Sea Res. 2015, 96, 11–17. [Google Scholar] [CrossRef]
- Capoccioni, F.; Costa, C.; Aguzzi, J.; Menesatti, P.; Lombarte, A.; Ciccotti, E. Ontogenetic and environmental effects on otolith shape variability in three Mediterranean European eel (Anguilla anguilla, L.) local stocks. J. Exp. Mar. Biol. Ecol. 2011, 397, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Milošević, D.; Bigović, M.; Mrdak, D.; Milašević, I.; Piria, M. Otolith morphology and microchemistry fingerprints of European eel, Anguilla anguilla Linnaeus, 1758) stocks from the Adriatic Basin in Croatia and Montenegro. Sci. Total Environ. 2021, 786, 147478. [Google Scholar] [CrossRef]
- Antunes, C.; Araújo, M.J.; Braga, C.; Roleira, A.; Carvalho, R.; Mota, M. Valorização dos recursos naturais da bacia hidrográfica do rio Minho. In Final Report from the Project Natura Miño-Minho; Centro interdisciplinar de Investigação Marinha e Ambiental; Universidade do Porto: Porto, Portugal, 2011. [Google Scholar]
- Vilas, F.; Somoza, L. El estuario del rio Miño: Observaciones previas de su dinâmica. Thalassas 1984, 2, 87–92. [Google Scholar]
- Sousa, R.; Dias, S.C.; Guilhermino, L.; Antunes, C. Minho River tidal freshwater wetlands: Threats to faunal biodiversity. Aquat. Biol. 2008, 3, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Dias, E.; Morais, P.; Cotter, A.M.; Antunes, C.; Hoffman, J.C. Estuarine consumers utilize marine, estuarine and terrestrial organic matter and provide connectivity among these food webs. Mar. Ecol. Prog. Ser. 2016, 554, 21–34. [Google Scholar] [CrossRef] [Green Version]
- Sousa, R.; Guilhermino, L.; Antunes, C. Molluscan fauna in the freshwater tidal area of the River Minho estuary, NW of Iberian Peninsula. Ann. Limnol.-Int. J. Limnol. 2005, 41, 141–147. [Google Scholar] [CrossRef]
- Mota, M.; Rochard, E.; Antunes, C. Status of the diadromous fish of the Iberian Peninsula: Past, present and trends. Limnetica 2016, 35, 1–18. [Google Scholar]
- Feunteun, E.; Acou, A.; Laffaille, P.; Legault, A. European eel (Anguilla anguilla): Prediction of spawner escapement from continental population parameters. Can. J. Fish. Aquat. Sci. 2000, 57, 1627–1635. [Google Scholar] [CrossRef]
- Durif, C.; Dufour, S.; Elie, P. The silvering process of Anguilla anguilla: A new classification from the yellow resident to the silver migrating stage. J. Fish Biol. 2005, 66, 1025–1043. [Google Scholar] [CrossRef]
- Durif, C.; Guibert, A.; Elie, P. Morphological discrimination of the silvering stages of the European eel. In American Fisheries Society Symposium; American Fisheries Society: New York, NY, USA, 2009; Volume 58, pp. 103–111. [Google Scholar]
- Fulton, T.W. The rate of growth of fishes. In Twenty-Second Annual Report; Fisheries Board of Scotland: Edinburgh, UK, 2009; pp. 141–241. [Google Scholar]
- Howard, S.W.; Crow, S.K.; Jellyman, P.G. Site-Specific Selectivity of Electric-Fishing Gear. New Zealand Fisheries Assessment Report. 2019. Available online: https://www.google.com.hk/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwi5zODSr5r4AhXzg2MGHff_B2cQFnoECAgQAQ&url=https%3A%2F%2Fwww.mpi.govt.nz%2Fdmsdocument%2F33166-far-201906-site-specific-selectivity-of-electric-fishing-gear&usg=AOvVaw3dPD5unARJBiQ69KpXsxdG (accessed on 22 May 2022).
- Souza, A.T.; Dias, E.; Nogueira, A.; Campos, J.; Marques, J.C.; Martins, I. Population ecology and habitat preferences of juvenile flounder Platichthys flesus (Actinopterygii: Pleuronectidae) in a temperate estuary. J. Sea Res. 2013, 79, 60–69. [Google Scholar] [CrossRef]
- ICES. Workshop on Age Reading of European and American Eel (WKAREA); ICES CM 2009\ACOM; ICES: Bordeaux, France, 2009; p. 48. [Google Scholar]
- ICES. Third Workshop on Age Reading of European and American Eel (WKAREA3); ICES: Bordeaux, France, 2020. [Google Scholar]
- Rasband, W. ImageJ v. 1.50i; National Institute of Health: Bethesda, MD, USA, 2009. Available online: http://imagej.nih.gov/ij/ (accessed on 22 May 2022).
- Tuset, V.M.; Lozano, I.J.; González, J.A.; Pertusa, J.F.; García-Díaz, M.M. Shape indices to identify regional differences in otolith morphology of comber, Serranus cabrilla (L., 1758). J. Appl. Ichthyol. 2003, 19, 88–93. [Google Scholar] [CrossRef]
- Campana, S.E.; Casselman, J.M. Stock discrimination using otolith shape analysis. Can. J. Fish. Aquat. Sci. 1993, 50, 1062–1083. [Google Scholar] [CrossRef]
- Kuhl, F.P.; Giardina, C.R. Elliptic Fourier features of a closed contour. Comput. Graph. Image Process. 1982, 18, 236–258. [Google Scholar] [CrossRef]
- Ponton, D. Is geometric morphometrics efficient for comparing otolith shape of different fish species? J. Morphol. 2006, 267, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, G.J.; Ward, T.M.; Gillanders, B.M. Otolith shape and elemental composition: Complementary tools for stock discrimination of mulloway (Argyrosomus japonicus) in southern Australia. Fish. Res. 2011, 110, 75–83. [Google Scholar] [CrossRef]
- Pothin, K.; Gonzalez-Salas, C.; Chabanet, P.; Lecomte-Finiger, R. Distinction between Mulloidichthys flavolineatus juveniles from Reunion Island and Mauritius Island (south-west Indian Ocean) based on otolith morphometrics. J. Fish Biol. 2006, 69, 38–53. [Google Scholar] [CrossRef]
- Campana, S.E.; Chouinard, G.A.; Hanson, J.M.; Frechet, A.; Brattey, J. Otolith elemental fingerprints as biological tracers of fish stocks. Fish. Res. 2000, 46, 343–357. [Google Scholar] [CrossRef]
- Wilson, R.R., Jr. Depth-related changes in sagitta morphology in six macrourid fishes of the Pacific and Atlantic Oceans. Copeia 1985, 4, 1011–1017. [Google Scholar] [CrossRef]
- Campana, S.E.; Neilson, J.D. Microstructure of fish otoliths. Can. J. Fish. Aquat. Sci. 1985, 42, 1014–1032. [Google Scholar] [CrossRef]
- Morales-Nin, B. Review of the growth regulation processes of otolith daily increment formation. Fish. Res. 2000, 46, 53–67. [Google Scholar] [CrossRef]
- Schiavina, M.; Bevacqua, D.; Melia, P.; Crivelli, A.J.; Gatto, M.; De Leo, G.A. A user-friendly tool to assess management plans for European eel fishery and conservation. Environ. Model. Softw. 2015, 64, 9–17. [Google Scholar] [CrossRef]
- Bevacqua, D.; Melià, P.; Schiavina, M.; Crivelli, A.J.; De Leo, G.A.; Gatto, M. A demographic model for the conservation and management of the European eel: An application to a Mediterranean coastal lagoon. ICES J. Mar. Sci. 2019, 76, 2164–2178. [Google Scholar] [CrossRef]
- Edeline, E.; Elie, P. Is salinity choice related to growth in juvenile eel Anguilla anguilla. Cybium 2004, 28, 77–82. [Google Scholar]
- Daverat, F.; Tomas, J. Tactics and demographic attributes in the European eel Anguilla anguilla in the Gironde watershed, SW France. Mar. Ecol. Prog. Ser. 2006, 307, 247–257. [Google Scholar] [CrossRef]
- Dias, E.; Miranda, M.L.; Sousa, R.; Antunes, C. Riparian vegetation subsidizes sea lamprey ammocoetes in a nursery area. Aquat. Sci. 2019, 81, 1–13. [Google Scholar] [CrossRef]
- Rooney, N.; McCann, K.S. Integrating food web diversity, structure and stability. Trends Ecol. Evol. 2011, 27, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Rytwinski, T.; Harper, M.; Taylor, J.J.; Bennett, J.R.; Donaldson, L.A.; Smokorowski, K.E.; Clarke, K.; Bradford, M.J.; Ghamry, H.; Olden, J.D.; et al. What are the effects of flow-regime changes on fish productivity in temperate regions? A systematic map. Environ. Evid. 2020, 9, 1–26. [Google Scholar]
- Domingos, I.; Costa, J.L.; Costa, M.J. Factors determining length distribution and abundance of the European eel, Anguilla anguilla, in the River Mondego (Portugal). Freshw. Biol. 2006, 51, 2265–2281. [Google Scholar] [CrossRef]
- Gravato, C.; Guimarães, L.; Santos, J.; Faria, M.; Alves, A.; Guilhermino, L. Comparative study about the effects of pollution on glass and yellow eels (Anguilla anguilla) from the estuaries of Minho, Lima and Douro Rivers (NW Portugal). Ecotoxicol. Environ. Saf. 2010, 73, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Marohn, L.; Jakob, E.; Hanel, R. Implications of facultative catadromy in Anguilla anguilla. Does individual migratory behaviour influence eel spawner quality? J. Sea Res. 2013, 77, 100–106. [Google Scholar] [CrossRef]
- Pereira, L.; Braga, A.C.; Moura, A.; Antunes, C. Prevalence of the Anguillicola crassus parasite in the International Minho River. Environ. Smoke 2021, 17, 64–74. [Google Scholar] [CrossRef]
- Feunteun, E.; Acou, A.; Guillouët, J.; Laffaille, P.; Legault, A. Spatial distribution of an eel population (Anguilla anguilla L.) in a small coastal catchment of Northern Brittany (France). Consequences of hydraulic works. Bulletin Français de la Pêche et de la Pisciculture 1998, 349, 129–139. [Google Scholar] [CrossRef]
- Félix, P.M.; Costa, J.L.; Monteiro, R.; Castro, N.; Quintella, B.R.; Almeida, P.R.; Domingos, I. Can a restocking event with European (glass) eels cause early changes in local biological communities and its ecological status? Glob. Ecol. Conserv. 2020, 21, e00884. [Google Scholar] [CrossRef]
- Davey, A.J.; Jellyman, D.J. Sex determination in freshwater eels and management options for manipulation of sex. Rev. Fish Biol. Fish. 2005, 15, 37–52. [Google Scholar] [CrossRef]
- Taborsky, B. The influence of juvenile and adult environments on life-history trajectories. Proc. R. Soc. B Biol. Sci. 2005, 273, 741–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elie, P. Contribution a L’etude des Montees de Civelles Anguilla anguilla Linne (Poisson, Teleosteen, Anguilliforme), Dans L’estuaire de la Loire: Peche, Ecologie, Ecophysiologie et Elevage. Ph.D. Thesis, University of Rennes, Brittany, France, 1979. [Google Scholar]
- Charlon, N.; Blanc, J.M. Etude des civelles d´Anguilla anguilla L. dans la region du bassin de l´adour. I. Caracteristiques biometriques de longueur et poids en function de la pigmentation. Arch. Hydrobiol. 1982, 93, 238–255. [Google Scholar]
- Edeline, E.; Dufour, S.; Elie, P. Proximate and ultimate control of eel continental dispersal. In Spawning Migration of the European Eel; Springer: Dordrecht, The Netherlands, 2009; pp. 433–461. [Google Scholar]
- Johansen, S.J.S.; Ekli, M.; Stangnes, B.; Jobling, M. Weight gain and lipid deposition in Atlantic salmon, Salmo salar, during compensatory growth: Evidence for lipostatic regulation? Aquac. Res. 2002, 32, 963–974. [Google Scholar] [CrossRef]
- Zeller, M.; Koella, J.C. Effects of food variability on growth and reproduction of Aedes aegypti. Ecol. Evol. 2016, 6, 552–559. [Google Scholar] [CrossRef]
- Whigham, D.F.; Baldwin, A.H.; Barendregt, A. Tidal Freshwater Wetlands. In Coastal Wetlands; Elsevier: Amsterdam, The Netherlands, 2019; pp. 619–640. [Google Scholar]
- Laffaille, P.; Feunteun, E.; Baisez, A.; Robinet, T.; Acou, A.; Legault, A.; Lek, S. Spatial organization of European eel (Anguilla anguilla L.) in a small catchment. Ecol. Freshw. Fish 2003, 12, 254–264. [Google Scholar] [CrossRef]
- Melià, P.A.C.O.; Bevacqua, D.; Crivelli, A.J.; Panfili, J.; De Leo, G.A.; Gatto, M. Sex differentiation of the European eel in brackish and freshwater environments: A comparative analysis. J. Fish Biol. 2006, 69, 1228–1235. [Google Scholar] [CrossRef]
- Geffroy, B.; Bardonnet, A. Sex differentiation and sex determination in eels: Consequences for management. Fish Fish. 2016, 17, 375–398. [Google Scholar] [CrossRef]

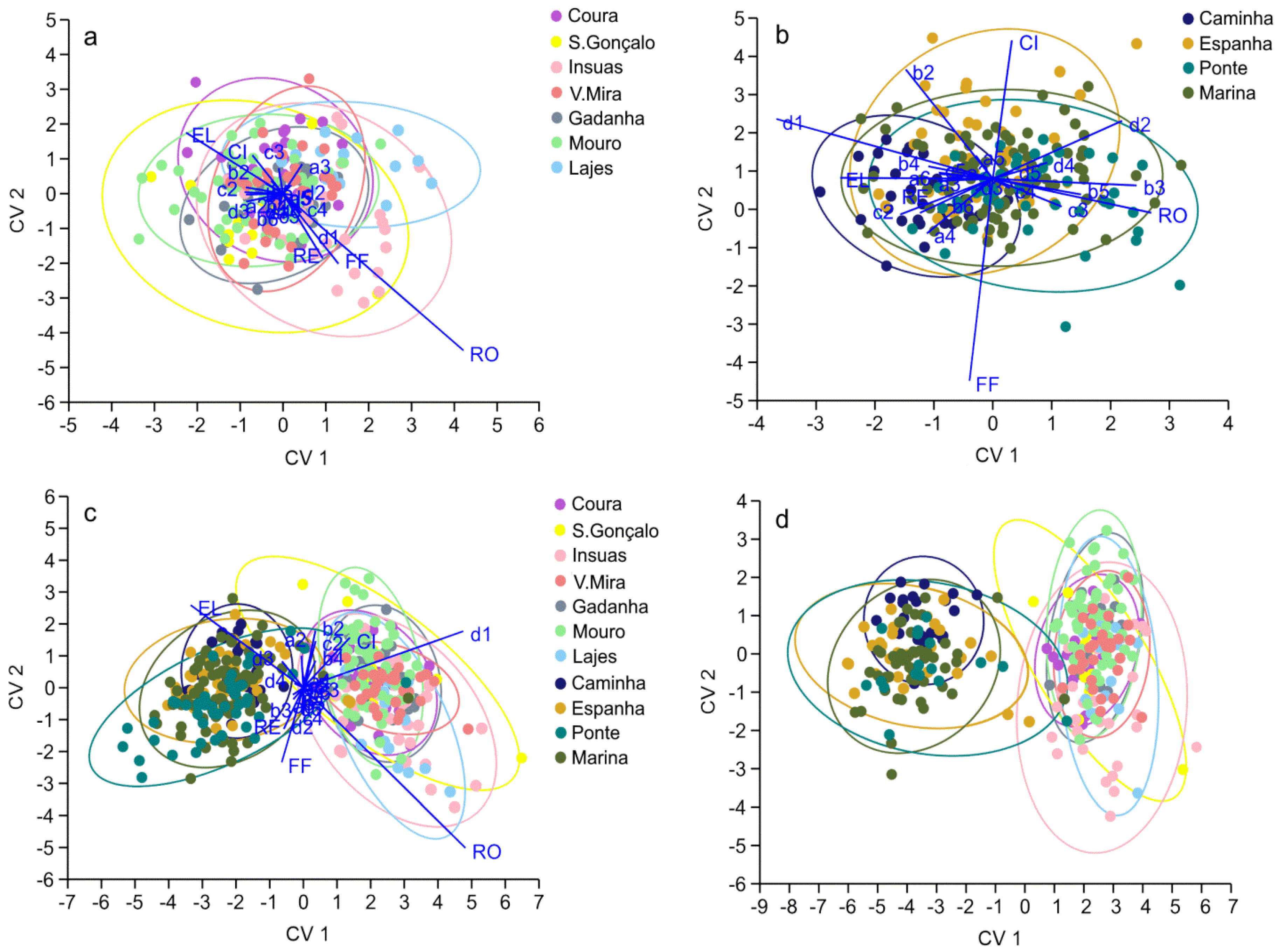

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling | N | Length (cm) | Durif Index-Length (cm, Mean ± SD) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Habitat | Location | Total | Silver | Min | Max | Mean ± SD | SI | SMII | SFII | SFIII | SFIV | SFIV |
| Coura | 127 | 3 | 6 | 34 | 17 ± 7.37 | 16 ± 6.86 | 33 ± 1.04 | - | - | - | - | |
| S.Gonçalo | 50 | 0 | 6 | 31 | 19 ± 6.72 | 19 ± 6.72 | - | - | - | - | - | |
| Campos | 184 | 0 | 6 | 34 | 14 ± 6.24 | 14 ± 6.24 | - | - | - | - | - | |
| Tributary | Insuas | 249 | 3 | 7 | 36 | 14 ± 8.07 | 14 ± 7.34 | 34 ± 1.46 | - | - | - | - |
| V.Mira | 99 | 0 | 6 | 33 | 19 ± 7.70 | 19 ± 7.70 | - | - | - | - | - | |
| Gadanha | 298 | 12 | 9 | 39 | 23 ± 6.39 | 22 ± 6.09 | 33 ± 2.13 | - | - | - | - | |
| Mouro | 284 | 3 | 10 | 35 | 19 ± 5.37 | 19 ± 5.06 | 31 ± 2.50 | - | - | - | - | |
| Lajes | 137 | 1 | 11 | 35 | 19 ± 5.84 | 18 ± 5.40 | 32 ± 2.80 | - | - | - | - | |
| Caminha | 84 | 2 | 15 | 64 | 33 ± 8.80 | 29 ± 4.27 | 36 ± 3.85 | - | 63 ± 3.49 | - | - | |
| Estuary | Espanha | 551 | 18 | 8 | 73 | 35 ± 11.21 | 30 ± 6.01 | 34 ± 3.09 | 49 ± 5.68 | 53 ± 11.76 | 56 ± 8.46 | 60 ± 7.00 |
| Ponte | 967 | 50 | 6 | 88 | 36 ± 11.92 | 30 ± 5.05 | 36 ± 3.68 | 49 ± 4.38 | 54 ± 13.72 | 57 ± 8.63 | 61 ± 6.28 | |
| Marina | 1074 | 18 | 19 | 71 | 34 ± 8.95 | 30 ± 5.21 | 35 ± 3.74 | 46 ± 1.56 | 48 ± 3.83 | 55 ± 4.08 | 57 ± 6.00 | |
| Habitat | Location | N | Sexed | Age | W (g) | L (cm) | FU (Mean ± SE) |
| Coura | 30 | 2 * | 1–10 | 1.53–69.4 | 10.6–34.9 | 0.146 ± 0.004 | |
| S.Gonçalo | 15 | 0 | 0–8 | 0.23–70.8 | 6.3–32.7 | 0.151 ± 0.004 | |
| Tributary | Insuas | 30 | 2 * | 0–10 | 0.4–75.6 | 7–36 | 0.136 ± 0.005 |
| V.Mira | 30 | 0 | 0–8 | 1.25–55.3 | 9.5–31 | 0.144 ± 0.003 | |
| Gadanha | 15 | 12 * | 2–10 | 3.02–75.6 | 14.9–34.6 | 0.154 ± 0.006 | |
| Mouro | 60 | 3 * | 1–10 | 1.6–73.6 | 11.6–35.2 | 0.125 ± 0.003 | |
| Lajes | 15 | 1 * | 1–9 | 3.55–90.5 | 14.4–35.9 | 0.150 ± 0.007 | |
| Caminha | 30 | 3/1 * | 3–7 | 16.3–78.2 | 23.0–36.2 | 0.156 ± 0.004 | |
| Estuary | Espanha | 45 | 12 | 0–9 | 0.67–276 | 8.4–53 | 0.160 ± 0.004 |
| Ponte | 45 | 25 | 0–9 | 0.57–367 | 6.7–54.6 | 0.173 ± 0.003 | |
| Marina | 90 | 40/3 * | 2–9 | 15.4–483 | 20.5–60.5 | 0.168 ± 0.004 | |
| Habitat | Sex | Age | W(g) | L(cm) | Silvering | ||
| Tributary | Undifferentiated | 0–10 | 0.23–69.4 | 6.3–34.9 | All Yellow | ||
| Male | 6–10 | 32.1–90.5 | 29–36 | All Silver | |||
| Estuary | Undifferentiated | 0–7 | 0.57–98 | 6.7–36.5 | All Yellow | ||
| Female | 4–9 | 57.6–483 | 31.1–60 | 2 Silver | |||
| Male | 4–7 | 46.9–87 | 30.8–37.6 | All Silver |
| (a) Tributary | |||||||||
| Original Locations | Coura | S.Gonçalo | Insuas | V.Mira | Gadanha | Mouro | Lajes | % Correct | |
| Coura | 8 | 4 | 5 | 6 | 4 | 1 | 2 | 27 | |
| S.Gonçalo | 2 | 3 | 1 | 3 | 2 | 3 | 1 | 20 | |
| Insuas | 1 | 5 | 9 | 1 | 5 | 4 | 5 | 30 | |
| V.Mira | 5 | 7 | 1 | 5 | 7 | 1 | 4 | 17 | |
| Gadanha | 5 | 3 | 2 | 5 | 9 | 3 | 3 | 30 | |
| Mouro | 7 | 5 | 6 | 11 | 11 | 15 | 5 | 17 | |
| Lajes | 2 | 0 | 1 | 3 | 1 | 2 | 6 | 40 | |
| Total | 30 | 27 | 25 | 34 | 39 | 29 | 26 | 26 | |
| (b) Estuary | (c) Habitat | ||||||||
| Original Locations | Caminha | Ponte | Espanha | Marina | % Correct | Original Locations | Tributary | Estuary | % Correct |
| Caminha | 17 | 5 | 6 | 2 | 57 | Tributary | 210 | 0 | 100 |
| Ponte | 6 | 12 | 12 | 15 | 27 | Estuary | 8 | 202 | 96 |
| Espanha | 9 | 11 | 15 | 10 | 33 | Total | 218 | 202 | 98 |
| Marina | 13 | 28 | 21 | 28 | 31 | ||||
| Total | 45 | 56 | 54 | 55 | 34 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moura, A.; Dias, E.; López, R.; Antunes, C. Regional Population Structure of the European Eel at the Southern Limit of Its Distribution Revealed by Otolith Shape Signature. Fishes 2022, 7, 135. https://doi.org/10.3390/fishes7030135
Moura A, Dias E, López R, Antunes C. Regional Population Structure of the European Eel at the Southern Limit of Its Distribution Revealed by Otolith Shape Signature. Fishes. 2022; 7(3):135. https://doi.org/10.3390/fishes7030135
Chicago/Turabian StyleMoura, Ana, Ester Dias, Rodrigo López, and Carlos Antunes. 2022. "Regional Population Structure of the European Eel at the Southern Limit of Its Distribution Revealed by Otolith Shape Signature" Fishes 7, no. 3: 135. https://doi.org/10.3390/fishes7030135
APA StyleMoura, A., Dias, E., López, R., & Antunes, C. (2022). Regional Population Structure of the European Eel at the Southern Limit of Its Distribution Revealed by Otolith Shape Signature. Fishes, 7(3), 135. https://doi.org/10.3390/fishes7030135