1. Introduction
The giant mottled eel,
Anguilla marmorata Quoy & Gaimard, 1824, has the widest known geographic distribution among the nineteen species/subspecies of the genus
Anguilla (Family Anguillidae) [
1,
2]. Unlike many anguillid species,
A. marmorata is believed to have at least four distinct spawning grounds, multiple population structure [
3,
4] and spawning throughout much of the year [
5,
6,
7,
8,
9,
10].
Due to the over-exploitation of temperate anguillid eel species (i.e.,
A. anguilla and
A. japonica) coupled with the drastic decline in juvenile recruitment in Europe and East Asia,
A. marmorata and many other tropical eel species in Southeast Asia have recently become targets to replace the temperate eels as species for human consumption [
11,
12,
13,
14]. Therefore, it is conceivable that tropical eel species will soon face an increased threat of extinction, as seen in their temperate congeners, if proper management and conservation policies are not implemented. In order to develop appropriate policies, data on genetic diversity and population structure of wild eel stocks are urgently needed [
15]. Indeed, the genetic population diversity of tropical anguillid eels has received intense research attention in recent years, with several studies published on
A. marmorata [
3,
4,
16,
17,
18],
A. bicolor [
19,
20],
A. bicolor bicolor [
21],
A. megastoma [
22] and
A. bicolor pacifica [
15]. Mitochondrial sequences, nuclear microsatellites and amplified fragment-length polymorphism have been used to determine the population genetic structure of
A. marmorata [
3,
4]. These methods revealed the existence of five populations, as determined on the basis of genetic differentiation indices, in the North Pacific Ocean, South Pacific Ocean, Southwestern Indian Ocean, Sumatra and Guam [
18].
Irrespective of the results obtained from these studies, future investigation into genetic diversity and population structure of tropical anguillid species in specific locations such as Vietnam is still required to gain insight into their migratory ecology and speciation mechanisms [
23].
Vietnam is known as one of the richest regions in the world in terms of marine biodiversity [
24]. There are at least three freshwater eel species (
A. marmorata,
A. japonica and
A. bicolor pacifica) found in the central parts of Vietnam [
25]. Of these species,
A. marmorata predominates and is consequently cultured widely throughout Vietnam, but farms depend entirely on wild-caught seed to stock tanks [
26]. There are currently no specific policies for active management or conservation of anguillid eels in Vietnam, mainly due to budget shortages [
26]. In the context of global conservation and management of anguillid eels, broad cooperation between countries is required [
14], and information about genetic diversity and population structure within a species is necessary for sustainable fisheries management [
27]. However, there is a severe lack of data on the
A. marmorata population in Vietnam.
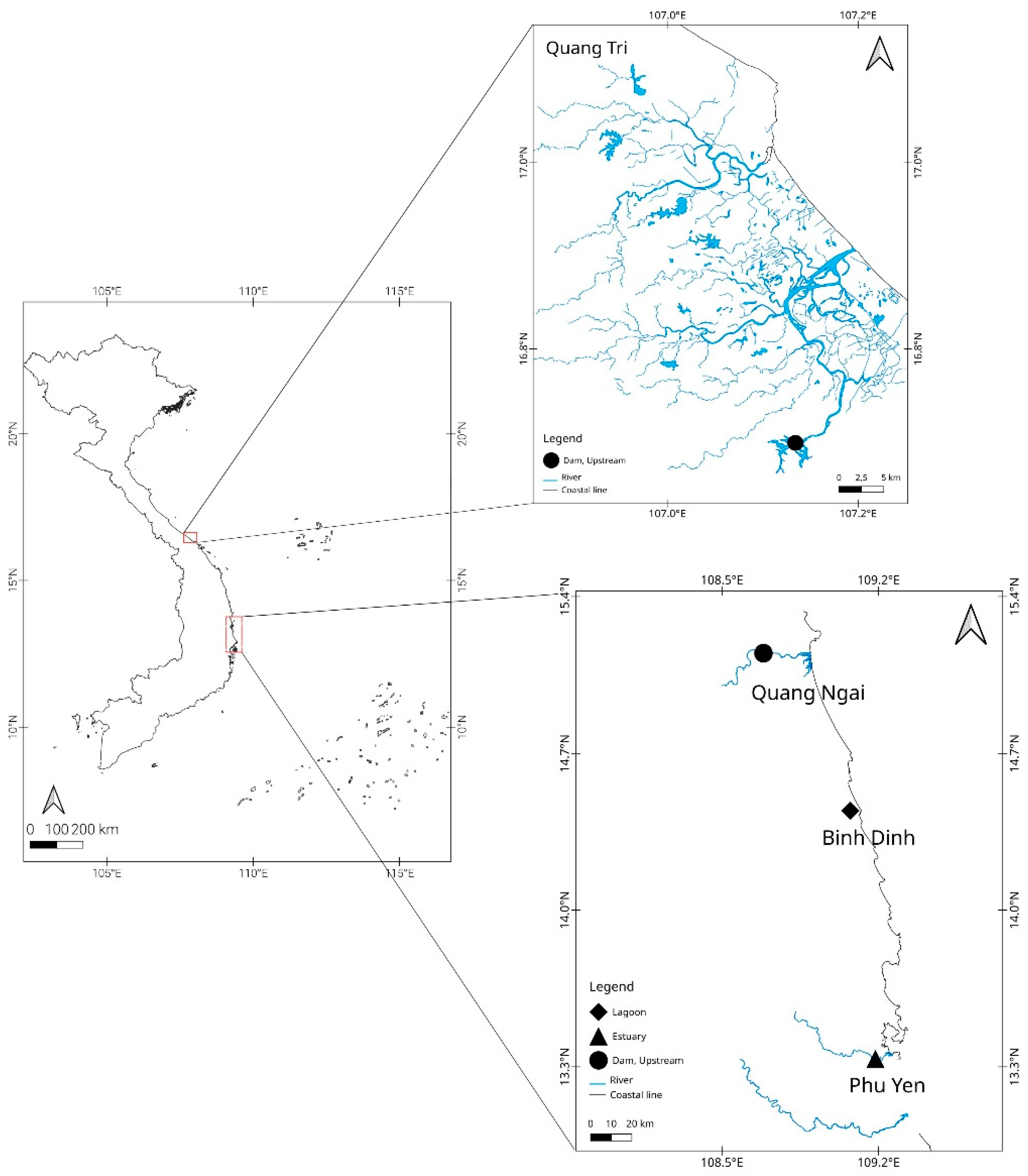
This study provides the first definitive data on population structure of freshwater eels in Vietnam, for use in eel conservation and management of aquaculture on a regional and/or global scale. By analyzing the sequences of the mitochondrial DNA control region (mtCR) for 176 individuals caught in locations throughout central Vietnam, we determined the population structure of A. marmorata in the area. Additionally, this study investigated the relationship of the Vietnam population to other A. marmorata populations around the world.
4. Discussion
Among freshwater eel species in Vietnam,
A. marmorata is the most economically important [
26], which is reflected by the fact that 95% of Vietnamese eel farms focus on the giant mottled eel [
37]. The inability to reliably propagate eels in captivity sees current farming practices relying solely on fattening of wild-caught glass eels. This over-exploitation of wild eel stocks has contributed heavily to declining populations and has left the species under threat. Information on the genetic status of
A. marmorata is necessary for effective resource management, but is currently limited. The present study examined the population structure of
A. marmorata in Vietnam and the relationship of this population to other previously researched populations around the globe.
Analysis of the complete mtCR sequence found high genetic diversity within the
A. marmorata population in central Vietnam. This was indicated by 165 distinct haplotypes and 306 polymorphic sites detected from 176 samples. Comparable variation has been reported in similar studies, which utilized the mtCR as a molecular marker to evaluate genetic population diversity of the giant mottled eel. For example, Ishikawa et al. [
3] identified 151 mt DNA haplotypes from 162
A. marmorata individuals, collected across ten localities (three in Japan, four in Indonesia and one each in Fiji, Tahiti and Madagascar). More recently, Minegishi et al. [
4] detected 267 separate haplotypes in a total of 290 individuals sampled from 13 different regions within the Pacific and Indian Oceans. Irrespective of rapid molecular evolution, the control region of the mtDNA is accepted as a suitable molecular marker for investigating intraspecific population relationships, due to its high mutation rate in comparison with other mtDNA regions [
38,
39,
40]. Therefore, the high value of haplotype diversity (hd = 0.999 ± 0.001) found in this study is reasonable. With regard to demographic estimators, the
A. marmorata population in Vietnam exhibited significant and negative Tajima’s D and Fu’s Fs values, indicating an excess of rare haplotypes and rapid population growth.
According to Frankham et al. [
41], an F
ST value > 0.15 implies significant genetic differentiation among sampling groups. In the present study, the F
ST value was very low (F
ST = 0.007) and non-significant (
p = 0.096). Moreover, there were only two pairs (BD1 and BD2; BD1 and QN) showing significant differences in the pairwise comparisons of F
ST values (see
Figures S2 and S3 for haplotype networks between these populations). Panmixia and a lack of significant population genetic structuring seem likely for
A. marmorata across our sampling sites. This assumption was supported by findings from Minegishi et al. [
4], in which the North Pacific population was considered to be fully panmictic.
Mitochondrial DNA markers are widely accepted in evolutionary and population genetics studies [
40]. However, the employment of an mtDNA marker in the present study inevitably comes with limitations. In particular, mtDNA has a high evolutionary rate and an effective population size, approximately one-quarter that of nuclear markers. This can lead to an underestimation of genetic diversity, unwanted biases in the genealogical analysis due to missing links in mitochondrial haplotypes and the limited detection of remote population processes [
42]. These weaknesses may have contributed to the identification of significant differences in the pairwise comparisons of F
ST values in only two pairs: BD1 and BD2; and BD1 and QN. Moreover, mtDNA is maternally inherited (i.e., lacks recombination) and inherently reflects only the matrilineal history instead of that of the overall population or species [
42]. Due to this restriction, it can be beneficial to combine multiple molecular markers, such as microsatellites, which can provide data on gene flow in the present day or in recent generations [
4]; this is something that should be incorporated into future studies in order to obtain a comprehensive understanding (i.e., parentage, relatedness and inbreeding depression) of the population genetic structure of
A. marmorata in Vietnam.
When comparing eels collected in Vietnam to other
A. marmorata populations around the world, most were genetically similar to eels found in Japan, Taiwan and the Philippines; all of these eels were assigned to the North Pacific population by Ishikawa et al. [
3] and Minegishi et al. [
4]. This was supported by high genetic identity (approximately 97%) and low F
ST values (−0.0068 to 0.0083,
p > 0.05) when comparing the Vietnam population to the three aforementioned populations (c.f.
Table 8). The findings from our study further reinforce this sentiment, as all individuals from the Vietnam population, except for one sample from Quang Tri and two samples from Binh Dinh (see below), formed a single large clade with other populations from the North Pacific (c.f. the neighbor-joining tree).
Miller et al. [
43] and Tsukamoto et al. [
44] proposed for the spawning area of
A. marmorata to be in the western North Pacific, close to the spawning ground of
A. japonica. Closeness of the spawning grounds is reflected, at least in part, in the well-documented sympatry of both species in China [
45], Taiwan and southern Japan (e.g., Wakiya et al. [
46]). Indeed, we have previously reported the coexistence of
A. marmorata and
A. japonica in the inland waters of Vietnam [
25]. Based on the currents in the South China Sea, we assume that recruitment of
A. marmorata along the coast of central Vietnam is influenced by two main currents, the South China Sea Warm Current and the Kuroshio Intrusion Current. Both of these currents originate from the main body of the Kuroshio Current [
47,
48] which is known to be one of the main currents transporting larval
A. marmorata throughout the Pacific [
2,
23].
Interestingly, we identified three individuals (QT-219, BD1-07 and BD1-013) which were more genetically similar to the South Pacific populations than to the North Pacific populations. Not only did these individuals show high values of genetic identity with
A. marmorata populations from Tahiti, Fiji, New Caledonia and Papua New Guinea, but they were also nested within the South Pacific clade of our neighbor-joining tree. Our findings are supported by
A. marmorata belonging to South Pacific populations found in areas of Southeast Asia, such as Indonesia (GenBank KU695248, KU692251 and KU692252) and Taiwan (KU885607), as documented in Arai et al. [
49].
The spawning area of
A. marmorata in the South Pacific remains incompletely defined. However, a recent study has suggested that there could be at least two spawning areas within the New Caledonia to Samoa region, and another further east [
50]. Additionally, Schabetsberger et al. [
51] proposed that the spawning area of
A. marmorata is likely located somewhere near Tuvalu, and the larvae are transported towards Southeast Asia by the South Equatorial Current at a maximum speed of 0.15 ms
−1. We note that there are several complex seasonal currents connecting the western region of the South Pacific to the Indian Ocean through Indonesia [
50]. This is reinforced by the complex currents depicted by Lee et al. [
52] and Pattiaratchi & Siji [
53]. According to Ganachaud et al. [
54], the North Vanuatu Jet is supplied by waters from the South Equatorial Current flowing north of Fiji. To the northeast of Australia, the North Vanuatu Jet forks with the northern portion flowing directly into the Solomon Sea, the remainder continuing towards the Queensland coast before turning north to join the Gulf of Papua Current [
55]. Further, the waters from both the North and South Pacific make their way to the Indian Ocean through the Indonesian Throughflow. Previous studies indicate that the North and South Pacific waters reach the Indonesia Seas through Makassar Strait, with a smaller contribution entering through Lifamatola Passage, the South China Sea and Karimatra Strait [
56,
57,
58,
59]. Taken together, it seems very likely that these complex currents could transport
A. marmorata from Micronesia to the South China Sea, but the specific current(s) responsible for this transport is (are) presently unknown. According to Dao et al. [
60], there was no difference in genetic population structure of the ornate spiny lobster
, Panulirus ornatus, across a broad region of the Southeast ranging from Vietnam, Indonesia, Australia and Papua New Guinea, due to its long oceanic larval development phase and wide larval transport capability [
61]. Hence, the finding of
A. marmorata glass eels originating from the South Pacific populations in the present study is reasonable.
Notably, the South China Sea is subjected to seasonal monsoons; during the southwest monsoon (summer) the surface circulation is directed northward, while the direction of circulation reverses during the northeast monsoon (winter) [
47,
62]. The sampling interval of BD1-07 and BD1-013 (glass eels) coincided with the southwest monsoon, indicating that these individuals could have been transported via a northward current from the South Pacific. In addition,
A. marmorata from the Ambon region, an area where individuals of the North and South Pacific populations co-exist [
4], could be transported by the Indonesia Throughflow via the Makassar Strait [
63] towards Vietnam. To date, glass eels have not been found in Quang Tri [
26]. Indeed, the QT-219 sample was an elver collected during the northeast monsoon. Therefore, it is possible that the QT-219 individual may have made its way from Binh Dinh to Quang Tri via the northward circulation in the South China Sea. This assumption was reinforced by the differences in size among sampled
A. marmorata between Binh Dinh (glass eels) and Quang Tri (yellow eels). Interestingly, QT-219 was part of a clade with Taiwan-24 (AB279089.1), Taiwan-43 (AB279103.1) and other individuals from South Pacific populations. Therefore, we cannot rule out the possibility that BD1-07, BD1-013 and QT-219 were transported directly to coastal Vietnam from the spawning ground in the South Pacific Ocean by unknown northward flows. Regardless of the mechanism, we conclude that our evidence for some
A. marmorata in central Vietnam to have originated from the South Pacific population is compelling.
It is clear that A. marmorata from the North and South Pacific populations migrate to Vietnam via seasonally dependent routes. In order to combat increased population pressures resulting from overfishing of this species in the South East Asia region, we recommend that the Vietnamese government revise their regulations with regard to a restricted fishing season, imposing catch size limits or requiring fishing licenses. Additionally, introducing policies which facilitate the passage of glass eels over dams, reduce anthropogenic influences on glass eel recruiment and restrict the collection of glass eels will significantly aid in the management and conservation of A. marmorata in Vietnam.


 ,
, 


{kind=link}